
Abstract
Plant growth, morphology and nutritive value under shade can differ between temperate grasses. Therefore, the aim of this study was to quantify the dry matter (DM) production, sward morphology, crude protein (CP%), organic matter digestibility (OMD) and macro-nutrient concentrations (P, K, Mg, Ca and S) in a grazed cocksfoot (Dactylis glomerata L.) pasture under 10-year-old Pinus radiata D. Don forest. Four levels of light intensity were compared: full sunlight (100% photosynthetic photon flux density-PPFD), open + wooden slats (∼43% PPFD), trees (∼58% PPFD) and tree + slats (∼24% PPFD). The mean total DM production was 8.2 t DM ha−1 yr−1 in the open and 3.8 t DM ha−1 yr−1 in the trees + slats treatment. The changes in cocksfoot leaf area index (LAI) were related to variations in morphological aspects of the sward such as canopy height and tiller population. CP% increased as PPFD declined with mean values of 18.6% in open and 22.5% in the trees + slats treatment. In contrast, the intensity of fluctuating shade had little effect on OMD with a mean value of 79 ± 3.2%. The mean annual macro-nutrient concentrations in leaves increased as the PPFD level declined mainly between the open and the trees + slats treatments. It therefore appears that heavily shaded dominant temperate pastures in silvopastoral systems limit animal production per hectare through lower DM production rates and per animal through reduced pre-grazing pasture mass of lower bulk density from the etiolated pasture.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The general aim of silvopastoral systems is to maximise profitability from a given area of land using combinations of compatible trees and pastures. Silvopastoralism in New Zealand with wide spaced conifers planted in native pasture (mainly ryegrass/white clover pasture) has generally resulted in trees of low timber quality and the pasture has been poorly adapted to tree competition (Hawke and Knowles 1997). Studies on different understory species indicate ways to maximise pasture production under the increasing area of trees on farms. Research with low density Pinus radiata D. Don stands (200–250 trees ha−1) has suggested that cocksfoot is suitable for silvopastoral systems due to its shade tolerance (Joshi et al. 1999; Peri et al. 2001).
Plant growth, morphology and nutritive value under shade can differ between temperate grasses (Devkota et al. 1997). Foliar nitrogen concentration has been shown to increase in shaded cocksfoot (Sheehy and Cooper 1973), but results were variable for herbage in vitro digestibility of grasses (Wong 1991).
A limited number of studies can be found emphasising effects of shading on nutritive value of temperate pastures. Most of these results have been generated by using continuous shading by cloth (Devkota et al. 1997). However, in silvopastoral systems understorey plants experience fluctuations in irradiance from full sun to shade (Peri et al. 2002), but the effects of this on pasture nutritive value have not been reported. Thus, the aim of this study was to quantify the response of cocksfoot dry matter (DM) production, plant morphology and nutritive value under different intensities of fluctuating light regimes compared with an adjacent open site as the control.
Materials and methods
Silvopastoral site
This study was conducted in the Lincoln University silvopastoral experimental area in Canterbury, New Zealand (43°38′ S and 172°28′ E). The original experiment was established in July 1990 to investigate soil/tree/pasture/sheep/climate interactions of five Pinus radiata genotypes and six understorey pasture treatments in a split-plot design with three replicates, being pasture species the main plots. The total area planted in trees is about 5.2 ha with 18 main pasture plots of 46.2 × 42.0 m (0.194 ha). After 11 years, the most persistent grass species was cocksfoot, which is the focus of this study. An adjacent 1 ha site without trees, on the same soil type, also had 18 pasture plots (plot size: 27.5 × 18 m) of the same age. Of these, three were cocksfoot plots, which were used to provide an open pasture comparison for the silvopastoral experiment. Additional areas of cocksfoot were available to allow 4–plot grazing rotations under trees and in the open. In all plots both open and under trees, herbage was cut and removed for silage during the first 3 years of the original experiment but since spring 1993 they have been grazed by sheep. The ‘Grasslands Wana’ cocksfoot pastures were originally sown with ‘Grasslands Pawera’ red clover (Trifolium pratense L.), ‘Grasslands Huia’ white clover (T. repens L.) and ‘Woogenellup’ subterranean clover (T. subterraneum L.).
The pine trees were planted at 1000 stems ha−1 (7 × 1.4 m) and were periodically thinned at 2.5, 3.5, 4.5 and 6.5 years after planting to the present uniform population of 200 stems ha−1 with 7 m between rows by 1996. In the first 2 years, tree rows were strip sprayed (1 m wide) with herbicide (hexazinone at 2.5 kg active ingredient ha−1) to assist tree establishment. Therefore, plots with trees had only 86% of their area occupied by sown pasture. Details of tree characteristics measured during this trial are given in Table 1. Crown closure had not occurred at age 10–11 years when the present experiment was carried out.
The soil is classified as a Templeton silt loam (Haplusteps) and consists of 1 to 2 m of fine alluvial sediments over gravels. It is medium to free-draining with a moderate capacity to hold moisture (320 mm in the top one meter). The site has only slight changes in topography, but there is variation in depth to the underlying gravels. Neither fertilizer, lime nor irrigation has been applied to the experimental area since its establishment. Thirty soil cores to 150 mm depth were taken at random within each cocksfoot plot in autumn 1999 and 2000 (Table 2). Soil tests indicated Olsen-P and S(SO4) were below optimum for maximum pasture production (Morton et al. 1994), but levels of Ca, K and Mg were adequate. In general, there were no differences in soil nutrients between cocksfoot plots in the open and in the silvopastoral site and to be consistent with the long-term experimental protocol no basal fertilisers were added to any of the pastures.
Experiment design with four light regimes
This experiment was set-up to measure DM production, morphology and changes in nutritive value of cocksfoot experiencing different levels of a fluctuating light regime from September 1999 to May 2001. Within each of the three main cocksfoot plots of the silvopastoral experiment, a study plot of 14.0 × 5.0 m was located in the middle of the 7.0 m wide inter-row under trees and also in the adjacent open pasture plots. Within these study areas, slatted shade structures measuring 3.0 × 2.1 m covered with pine wood slats (150 mm wide) and gaps between slats (150 mm wide) were used to reduce the total incidence of light by approximately 50% (Plate 1). This structure provided a bimodal light regime to represent the silvopastoral system (Peri et al. 2002). The shade structures were supported horizontally on a vertically adjustable metal frame, which allowed the shade source to be maintained at 0.3 m above the cocksfoot canopy. For the slatted shade structure, the objective was to create intervals of sunlight and shade similar to the shade pattern of the radiata pine in the silvopastoral area (Plate 1).

Cocksfoot pasture under 10-year-old radiata pine trees (200 stems ha−1, pruned up to 6 m height) at Lincoln University silvopastoral (Canterbury, New Zealand) experiment which provided a fluctuating light regime of ∼58% of open PPFD. In the middle of the 7 m inter-row, slatted shade structures were used to reduce the total incidence of light by approximately 50%. This structure provided a bimodal light regime
This experiment was arranged with open (100% transmittance) and silvopastoral (∼58% transmissivity) plots as main treatments with three replicates. Within each replicate a cocksfoot plot was split into two sub-plots: slatted shade and no slatted shade. This gave four light transmission regimes: (i) cocksfoot open pasture, (ii) cocksfoot pasture under slatted shade, (iii) cocksfoot pasture under tree shade, (iv) cocksfoot pasture under trees + slatted shade. The trees + slatted shade treatment extended the light regime beyond that experienced under the current silvopastoral situation.
The slatted shade structures were orientated in an East–West direction in the main plots with the slats North–South. They were set up continuously in the plots from September 1999 to May 2001. During periods when main plots were grazed, the shade frames were removed to avoid damage on plants through sheep using the structures as a camping area. Immediately after each grazing, plots were trimmed with a mower to an even height of 20 mm and slatted frames were replaced to their original positions.
Grazing management
A flock of Coopworth ewe lambs were rotationally grazed for 7 ± 1 days around the three cocksfoot main plots plus the extra cocksfoot area under trees (28 day rotation with 21 ± 1 days regrowth). A smaller flock of the same type of sheep was grazed in the same rotational pattern around the four adjacent open cocksfoot pastures. All pastures were grazed from 15 September 1999 (initial liveweight of 45 ± 3 kg) to 21 May 2000 and from 21 September 2000 (initial liveweight of 42 ± 5 kg) to 2 April 2001. Because pasture was drought stressed, grazing was stopped from 16 March to 15 April 2000 and from 26 January to 8 March 2001, to allow pasture to accumulate the minimum pre-grazing mass of 2.0 t ha−1. Stocking rate during grazing periods, over 2 years, under trees averaged 16 lambs ha−1 and 25 lambs ha−1 in the open. Stocking rate was adjusted when necessary after each liveweight measurement (37 ± 5 day intervals) to ensure a similar pasture allowance for both flocks (mean pasture allowance of 3.2 kg DM animal−1 d−1).
Physical environmental measurements
The air temperature measurements were taken on site in the open and under trees using digital temperature sensors (TDC-01A, Monitor Sensors, Queensland, Australia) located 1.5 m above ground, which logged every 6 min (resolution ± 0.2°C). Rainfall measurements were obtained from the Broadfields meteorological station located 3 km north of the silvopastoral site. The mean soil volumetric water content (VWC) in the top 500 mm was measured every 7 days with Time Domain Reflectometry (TDR, Trase Systems, Santa Barbara, USA).
Light intensity was monitored with quantum sensors (Li-cor LI-191SB, Lincoln, Nebraska, USA) installed above and below the slatted shade structures, but above cocksfoot canopy height. This gave a quantitative description of the four levels of shade used in the experiment. The quantum sensors measured the photosynthetic photon flux density (PPFD) in the 400–700 nm waveband every 5 min by a datalogger with mean PPFD recorded at 30 min intervals.
Spectral irradiance from 300 to 1100 nm wavelengths was measured with a Li-Cor LI-1800 spectro-radiometer (Lincoln, Nebraska, USA). Measurements were taken at noon and 17:00 h for a sunny day in spring, which corresponded to solar angle elevations of 46.5° and 17.6°, respectively. Also, measurements were taken at noon for a cloudy day. From the total spectral irradiance data, proportions of red (660 nm) to far-red (730 nm) wavelengths were calculated.
Biological measurements
Herbage measurements were taken prior to lambs grazing (21 ± 1 days regrowth). For all treatments, pasture samples for DM production were obtained from a 0.2 m2 quadrat cut to 20–25 mm stubble height. Canopy height was measured using a sward stick before herbage harvesting. The number of vegetative tillers was counted as new leaf extension above the grazed leaf sheath height within 3–5 days post-harvest using a circular 0.01 m2 quadrat.
Pasture samples were dried in a forced draught oven at 65°C to constant weight and ground in a mill containing a 1 mm stainless steel screen for crude protein (CP) and organic matter digestibility (OMD) determinations. The nitrogen content of leaves was determined using the Kjeldahl technique (CP% = N% × 6.25). Phosphorus was analysed according to method No. 964.06 (Padmore 1990). Calcium and magnesium analyses were done by atomic absorption after Kjeldahl digestion (Padmore 1990). Potassium analysis was done by the atomic emission method (Padmore 1990) and Sulphur analysis by Quin and Woods (1976).
The Li-cor LAI-2000 Plant Canopy Analyser (Lincoln, Nebraska, USA) was used to measure leaf area index (LAI). The Li-cor LAI-2000 is a hand-held instrument, with optical sensors that includes a fisheye lens and five silicon detectors allowing simultaneous measurement of the radiation coming from the upward hemisphere in five zenithal angles. There are difficulties in measuring total LAI for grasses because the optical sensor head of the instrument is 40 mm high. Therefore, aluminium trenches 40 mm deep × 30 mm wide × 1.2 m long were set up for all treatments so that the top of the sensor was at the soil surface. In this study, measurements were taken from one reading above the canopy followed by five readings beneath, along the trench (transect). As the Li-cor LAI-2000 requires diffuse light to give reliable measurements, the instrument was only used under uniform overcast conditions, or before sunrise and after sunset. To avoid contamination of the measurements by the operator, a 180° view cap was used.
Statistical analysis
Statistical analyses were carried out using the Genstat statistical package (Genstat 5, 1997). Standard error of means (sem) were used to evaluate least significant differences (lsd) at the 0.05 probability level for mean separation of the pasture variables. Significant differences for the experiment with four light regimes were determined for each rotation by analysis of variance (ANOVA) according to the split-plot design with three replicates. Pasture variables were also analysed by considering time as a factor. Thus, this analysis was carried out to detect potential interactions between a pasture variable (such as DM growth rate) and the main environmental factors (such as temperature) which vary with time (seasons). DM yield and LAI data from vegetative cocksfoot pastures obtained from each harvest of the experiment (80 data points) were analysed using non-linear regression analysis. The fitted parameters for each treatment of both experiments were compared using an ANOVA.
Results
Environment
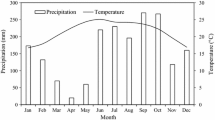
The mean daily temperature was similar in the open and under trees (Fig. 1). In both summers, (December–February 1999/2000 and 2000/2001), the mean temperature under trees was 0.4°C warmer than in the open, and during winter (June–August 2000) it was 0.2°C warmer. However, during a sunny day in autumn-winter (maximum temperatures between 10 and 15°C) the temperature under trees was up to 3°C warmer at midday and morning (from 5:00), but the difference was reversed after sun set. In contrast, during sunny hot days in summer (>28°C) there was minimal difference in air temperature under trees and open pasture sites.

Rainfall (Ͷ) and mean monthly air temperature under trees (—) and in the adjacent open (··–··) pasture at the Lincoln University silvopastoral experiment (Canterbury, New Zealand) from September 1999 to May 2001
Rainfall was 17% less than the long-term mean over the study period. This was mainly because for March–April 2001, rainfall was only 9.2 mm, which was approximately 90% less than the long-term mean (Fig. 1). In spring and winter, the mean soil VWC in the top 500 mm was always above 24% and was therefore always greater than half the maximum available water content of the site (mean field capacity = 30%) indicating that the treatments were not moisture stressed during those periods (Fig. 2). However, in summer and autumn of both years, pastures were under water stress. On average, pastures under trees had 2.5% less soil VWC than open pastures. The shaded treatment (open + slats) had a higher soil VWC than open. Similarly, the treatment trees + slats had a higher soil VWC than the pasture under trees. This additional soil VWC under the slatted shade resulted in greater water recharge during winter. For example, in July 2000 the pastures in the open had a soil VWC of 30.5% compared with 32.0% in the open + slats treatment.

Mean soil volumetric water content in the top 500 mm (measured every 7 days) for four shaded treatments at the Lincoln University silvopastoral experiment (Canterbury, New Zealand): Open (—) (100% transmissivity), open + slats (▁) (∼43% transmissivity), under trees (...) (∼58% transmissivity) and trees + slats (---) (∼24% transmissivity). Bars indicate standard error of the mean (sem)
The daily PPFD was integrated to calculate the accumulated monthly photosynthetic photons per unit area (Fig. 3). The maximum photosynthetic photons reaching the cocksfoot pasture was in December (1715–1815 mol m−2 for open pastures) corresponding to the maximum noon solar angle elevation (69.8° at noon). The minimum (302 mol m−2 for open pastures) was in June with the lowest noon solar angle elevation of 23°. In December, pastures in the open received 720, 960 and 1220 mol photons m−2 more than pastures under trees, open + slats and trees + slats, respectively. However, these differences decreased in June. The daily PPFD integral in the open for a sunny day in spring or autumn (e.g. 21 September or 21 March at solar angle elevation of 46.5° at noon), summer (21 December at solar angle elevation of 69.8° at noon) and winter (21 June at solar angle elevation of 23.0° at noon), and over a range of cloudy days were used as a reference (100% transmissivity) to calculate the transmissivity of the shade treatments (Table 3). This was used to represent the relative reduction of photosynthetic photons in the shaded treatments compared with the open pasture. The total daily integral photosynthetic photons received in open pasture around the 21 December was 63.3 mol photons m−2 d−1, which was 6 times higher than in winter (21 June) (Table 3). For cloudy days (diffuse light) during summer and spring the total integral daily photosynthetic photons received in open pasture varied between 7 and 18 mol photons m−2 d−1 depending on the cloud type. The transmissivity under the 10 to 11-year-old trees measured in the middle of rows was 62% of the open over a sunny day in summer (at maximum solar elevation), with alternating periods of full sunlight and this decreased to 26% with the addition of the slatted structure. The transmissivity of the shaded treatments decreased with a decrease in solar angle elevation from summer to winter. The transmissivity of the tree-shaded treatments during cloudy days (58%) was lower than sunny days in spring and summer (60–62%), but under the slatted shade it remained at 45% between cloudy and sunny days (Table 3).

Mean monthly photosynthetic photons (400–700 nm waveband) received for cocksfoot pastures from the four shaded treatments at the Lincoln University silvopastoral experiment (Canterbury, New Zealand): open (◯), open + slats (∇), under trees (●) and trees + slats (▼)
The red–far red ratio (R:FR) decreased from sun to any of the shaded situations (Table 4). The minimum value of R:FR was 0.54 at noon in the middle of the tree shade. The R:FR also decreased under the shade of slats. There was a difference in R:FR within the tree shade with higher values along the perimeter (0.5 m inside the shaded zone). There was no difference in R:FR for two different solar angles elevation (noon and afternoon) for full sunlight conditions. However, under the tree shade, the R:FR increased at the lowest solar angle. At noon on the cloudy day, the R:FR was greater under trees and the slatted structure compared with the sunny day, but still less than the R:FR in open.
Pasture DM production and growth rate
The mean total DM production was 8.2 t DM ha−1 yr−1 in open, 7.3 t DM ha−1 yr−1 in open + wooden slats, 6.3 t DM ha−1 yr−1 under trees shade and 3.8 t DM ha−1 yr−1 in the trees + slats treatment. The mean clover percentage of the cocksfoot pasture for the two grazing seasons was 8% in open and 4% under trees with a mean of cocksfoot content over 80%. Differences in DM production were driven by DM growth rates (Fig. 4) being lower under trees and trees + slats compared with the full sunlight treatment in all seasons. The mean DM production rate of cocksfoot pasture for the grazing seasons (September–April) for the 2 years was 30 kg DM ha−1 d−1 in open, 26 kg DM ha−1 d−1 in open + slats, 21 kg DM ha−1 d−1 under trees and 14 kg DM ha−1 d−1 under trees + slats. For the dry period January–March 2001, pastures in the open under slat shade produced more than the adjacent full sunlight treatment. There was an interaction (P < 0.05) between treatments and time (rotations). This was expressed by seasonal fluctuations in pasture DM growth rates (Fig. 4). The highest (P < 0.05) growth rates occurred during November (mean of 48 kg DM ha−1 d−1 in open, 43 kg DM ha−1 d−1 in open+slats, 35 kg DM ha−1 d−1 under trees and 24 kg DM ha−1 d−1 under trees + slats) and there was a rapid decrease in summer (December–February) and winter (June–July). In autumn 2000 (April–May), there was a recovery after summer drought showing a typical bimodal annual growth curve. However, this trend did not occur during autumn 2001. DM production rate was higher in the second year during spring compared with the first year for pastures in the open and under trees, but lower for the shaded treatments; open + slats and trees + slats.

Cocksfoot dry matter growth rates (21±1 days regrowth) over time for four shade treatments at the Lincoln University silvopastoral experiment (Canterbury, New Zealand): open (◯) (100% PPFD), open + slats (∇) (∼43% transmissivity), under trees (●) (∼58% transmissivity) and trees + slats (▼) (∼24% transmissivity). Bars indicate standard error of the mean (sem)
Leaf area index (LAI)
As for DM production, LAI curves showed seasonal fluctuations (Fig. 5) which was indicated by the interaction (P < 0.05) between treatments and time (rotations). The greatest (P < 0.05) LAI occurred in spring during October–November (mean of 4.1 in open, 3.8 in open + slats, 3.0 under trees and 2.2 under trees + slats) and there was a rapid decrease in late summer and winter. The LAI was consistently lower under trees (P < 0.05) and trees + slats (P < 0.01) compared with the full sunlight treatment in all seasons (Fig. 5).

Cocksfoot leaf area index (LAI) (21 ± 1 days regrowth) over time for four shade treatments at the Lincoln University silvopastoral experiment (Canterbury, New Zealand): open (◯) (100% transmissivity), open + slats (∇) (∼43% transmissivity), under trees (●) (∼58% transmissivity) and trees + slats (▼) (∼24% transmissivity). Bars indicate standard error of the mean (sem)
Canopy pasture height and tiller population
The changes in cocksfoot LAI were related to variations in morphological aspects of the sward such as canopy pasture height and tiller population. Details of changes in pasture tiller population and canopy height over time for the experiment with four light regimes are given in Figs. 6 and 7, respectively. When water was non-limiting (soil VWC > 24%), the cocksfoot canopies under shade were taller (P < 0.05) than those grown in full sunlight. During the period of maximum increase in height (October-November), cocksfoot tillers under the shade of trees + slats were etiolated to be 60 mm taller than comparable tillers in full sunlight. In general, cocksfoot tiller population decreased (P < 0.05) as shade level increased with a mean vegetative tiller population per m2 of 5540 in the full sunlight, 5020 in the open + slats treatment, 4720 under trees and 3570 tillers m−2 in the tree + slats treatment.

Cocksfoot tiller population (21 ± 1 days regrowth) over time for four shade treatments at the Lincoln University silvopastoral experiment (Canterbury, New Zealand): open (◯) (100% transmissivity), open + slats (∇) (∼43% transmissivity), under trees (●) (∼58% transmissivity) and trees + slats (▼) (∼24% transmissivity). Bars indicate standard error of the mean (sem)

Cocksfoot canopy height (21 ± 1 days regrowth) over time for four shade treatments at the Lincoln University silvopastoral experiment (Canterbury, New Zealand): open (◯) (100% transmissivity), open + slats (∇) (∼43% transmissivity), under trees (●) (∼58% transmissivity) and trees + slats (▼) (∼24% transmissivity). Bars indicate standard error of the mean (sem)
Relationship between DM yield and LAI
The lack of significant differences in the slope of these relationships meant a single function could be used (Fig. 8). This relationship was described by an exponential function (Eq. 1), which resulted in an r2 of 0.93 and standard error of the estimate (ESE) of DM yield of 96.2 kg DM ha−1.
From 0.5 to 2.0 units of LAI, the relationship was approximately linear and increased at a rate of 216 kg DM ha−1 per unit of LAI. From this point to LAI = 4.5 the relationship was curvilinear (Fig. 8).

Accumulated dry matter (DM) yield (kg DM ha−1) against leaf area index (LAI) for vegetative cocksfoot pastures grown at the Lincoln University silvopastoral experiment from the four light regimes experiment (Canterbury, New Zealand). The line is for the fitted single exponential function (Eq. 1)
Nutritive value
CP percentage increased as PPFD declined (Fig. 9) with mean values of 18.6% in open, 21.2% in open + wooden slats, 19.5% under trees shade and 22.5% in the trees + slats treatment. The interaction (P < 0.05) between treatments and time showed the increase in CP percentage with shade under trees showed less difference during severe drought in both summers and autumn 2001 (Fig. 9).

Cocksfoot crude protein (CP%) content over time for four shade treatments at the Lincoln University silvopastoral experiment (Canterbury, New Zealand): open (◯) (100% transmissivity), open + slats (∇) (∼43% transmissivity), under trees (●) (∼58% transmissivity) and trees + slats (▼) (∼24% transmissivity). Bars indicate standard error of the mean (sem)
In contrast, the intensity of fluctuating shade had little effect on OMD with a mean value of 79 ± 3.2%, but with a fluctuating (P < 0.05) seasonal variation which ranged from 72.0 to 85.8% (Fig. 10). Maximum values corresponded to spring, winter and autumn regrowth and minimum values when cocksfoot pastures were under water stress (soil VWC < 18%).

Cocksfoot organic matter digestibility (OMD%) over time for four shade treatments at the Lincoln University silvopastoral experiment (Canterbury, New Zealand): open (◯) (100% transmissivity), open + slats (∇) (∼43% transmissivity), under trees (●) (∼58% transmissivity) and trees + slats (▼) (∼24% transmissivity). Bars indicate standard error of the mean (sem)
Table 5 indicates that mean macro-nutrient concentrations of cocksfoot leaves increased (P < 0.05) as the PPFD level declined (Table 5), mainly between the open (100% transmissivity) and the trees + slats (∼24% transmissivity) treatments (P 4.2 vs 5.2, Mg 1.6 vs 2.1, K 29.4 vs 33.9, Ca 4.7 vs 5.3, S 2.6 vs 3.6 g kg−1 DM). However, there was an interaction (P < 0.05) between treatments and time (seasons) with shade under trees which showed less difference during severe drought in both summers and autumn 2001. Thus, macro-nutrient concentrations were higher (except Ca) in spring and winter (means P = 5.1, Mg = 1.9, K = 32.4, Ca = 4.9, S = 3.4 g kg−1 DM) compared with summer and autumn (means P = 4.4, Mg = 1.8, K = 29.6, Ca = 5.2, S = 3.0 g kg−1 DM).
Discussion
Effect of shade on DM production
The specific component unique to silvopastoral systems is the light regime compared with open pastures. In this study, the tree canopy and slatted structures reduced and modified the light available to the understorey cocksfoot pasture. Specifically, the daily PPFD integral for a sunny day in summer (around 21 December at solar angle elevation of 69.8° at noon) was 63.3 mol photons m−2 d−1 (100% transmissivity) and this was reduced by 38% under trees and 74% under the slatted structures in the silvopastoral system (Fig. 3). The reduction in available light quantity for the understorey pasture also changed with cloudy conditions and differences in solar angle elevation throughout the seasons. As a consequence, cocksfoot DM growth rate decreased by 13% under slat shade in the open, 22% under tree shade and 48% under the trees + slats shade compared with the full sunlight pastures during periods of non-limiting water (soil VWC >25%) (September–November 1999 and 2000). This reduction in DM growth is in the range reported in the literature. For example, Joshi et al. (1999) reported that yield of irrigated cocksfoot pasture under 650 (18% of the open PPFD) and 300 (40% of the open PPFD) trees ha−1 was reduced by 55% and 16% compared with open pasture, respectively. Similarly, in northern Greece, Braziotis and Papanastasis (1995) reported that cocksfoot DM production during spring was reduced by 55% under a 20-year-old maritime pine (Pinus pinaster Aiton) plantation thinned to 1750 trees ha−1 (mean light intensity of 31% of the open area) compared with pastures under 1000 trees ha−1 (mean light intensity of 41% of the open area). Hawke and Knowles (1997) reported that DM production of ryegrass (Lolium perenne L.)/white clover (Trifolium repens L.) pastures at Tikitere (Rotorua, North Island, New Zealand) was 25% of the open pasture production at age 13 years for 200 trees ha−1. Similar results have been reported for perennial ryegrass in South Otago (New Zealand) (Cossens 1984) and in nine silvopastoral environments in the United Kingdom (Sibbald et al. 1991).
Interaction with time
Changes in environmental factors over time (seasons) had a strong influence on DM production. For example, the mean daily temperature during this experiment ranged from 6°C in winter to 16°C in summer (Fig. 1) with daily minimum temperatures of 1.4°C and daily maximum temperatures of 22.6°C. In addition, as a result of seasonal effects, the soil VWC in the top 500 mm varied from 33 to 8.5% (Fig. 2). These changes provided a wide range of cocksfoot DM growth rates from 2.1 to 51.3 kg DM ha−1 d−1.
The decrease in DM production with shade intensity showed seasonal variation responses (Fig. 4) with less difference during winter (mean daily air temperatures <8°C) and during severe drought (soil VWC < 15%). This indicates that pasture production during winter was limited mainly by low temperatures and by soil water stress in dry conditions. Similarly, Korte et al. (1987) reported that low levels of solar radiation do not appear to limit unshaded pasture production in winter. Low temperature is considered to be the major environmental variable limiting pasture production for this season in temperate latitudes. In addition, trees in the silvopastoral plots reduced the soil VWC in all seasons with a mean reduction of 2.5% compared with open pastures due to root competition and the interception of rainfall. These probably also contributed to a reduction in DM growth rate in addition to shade. However, there was some evidence that shade assisted soil moisture conservation during drought periods. For example, from January to April 2000, the open + slats treatment produced 15% more DM than open pastures as a consequence of 2.2% more soil VWC (Fig. 4).
Leaf area index (LAI), canopy pasture height and tiller population
LAI, which depends on the rate of leaf appearance, growth and death of individual tillers and leaves and their morphological changes, has been reported to be dependent on temperature, irradiance, water status (Davies 1988) and light quality (Casal et al. 1987).
Change in light quantity and quality (mainly the decrease of the R:FR ratio) under trees can modify LAI because stem elongation can be promoted and tillering and branching inhibited (Casal et al. 1987; Garnier and Roy 1988). In this study, where flowering of cocksfoot pasture was not allowed under grazing, shade encouraged plants to become etiolated (Fig. 7) where the taller growth may be an effort to gain greater access to available light in competition with neighbouring plants and tillers. For example, when water was non-limiting (September–November), shade increased canopy height by approximately 23% for both open + slats and under trees pastures, and by 41% for the trees + slats treatment. Anderson (1978) found that etiolation of cocksfoot was due to cell elongation under shaded environments. According to Kephart and Buxton (1993) etiolation occurs at the expense of root growth, increasing consequently the plant shoot/root ratio under shade.
It also appears that the etiolation of shaded cocksfoot plants, responded to a reduction in the R:FR ratio. In the current study, differential absorption of red and far-red light from the tree canopies produced a 56% decrease in the R:FR ratio in the middle of the tree shade compared with full sunlight (Table 4). The changes in R:FR ratio are perceived by understorey plants through the phytocrome system which may change morphogenetic characters in plants (Smith 1982). It is likely that leaf area of shaded cocksfoot leaf blades is maintained or increased to maximise light interception at the expense of leaf thickness, resulting in leaves being longer, narrower, and thinner than when grown in full sunlight conditions. This is consistent with Devkota et al. (2000) who reported that plants from 10 cocksfoot selections increased the specific leaf area with shade from 15.9 mm2 mg−1 under 73% of the open PPFD to 21.3 mm2 mg−1 under 24% of the open PPFD.
Reduced light intensity and changes in light quality have been reported to reduce tillering and are therefore likely to reduce LAI. In this study, cocksfoot vegetative tiller population decreased from 5540 in the full sunlight to 3570 tillers m−2 under severe shade (∼24% transmissivity) (Fig. 6). Similarly, Garnier and Roy (1988) reported a 36% reduction of cocksfoot tiller population in France under 33% transmissivity oak tree shade compared with open pasture. Devkota et al. (1997) reported for a range of cocksfoot cultivars that the mean tiller number declined 25–30% as the shade environment fell from 77 to 17% PPFD of full sunlight. Deregibus et al. (1983) showed that after 28 days, the mean number of new tillers per plant of Lolium sp. was 16 with a R:FR of 2.2 and decreased to 11 tiller per plant when R:FR declined to 1.1 of similar light intensity. A similar response was reported by Casal et al. (1987). The physiological basis for the reduction in tillering is that under low irradiance a reduced supply of current assimilate is preferentially allocated to existing tillers at the expense of axillary buds (Robson et al. 1988). Thus, the effect of low light intensity is not on the rate of site production, but rather on the extent to which sites are filled.
Relationship between DM yield and LAI
The non-linear relationship between DM yield and LAI (Eq. 1) for the vegetative cocksfoot sward, indicated that the LAI of the sward increased more slowly in relative terms than the biomass when LAI was greater than 2 units (Fig. 8). This was consistent with Duru et al. (1997) who reported, for cocksfoot, a unique exponential function between LAI and DM. One reason for the non-linear relationship, mainly from LAI below 2, would be that the pseudo-stem (sheath) length and weight increased with LAI (or time of regrowth) and consequently decreased the leaf:pseudo-stem ratio. As a consequence, the proportion of green leaf was reduced. Thus, the increase in the more vertical and heavier pseudo-stem component and the relative decrease in the green leaf component over time gives a greater proportion of DM with a smaller increase in LAI.
The importance of this single relationship between DM yield and LAI is that it includes differences in the cocksfoot canopy due to changes over time in morphological aspects of the sward, such as tiller population, pasture height (Figs. 6, 7) and leaf size caused by environmental and management factors. This relationship between DM production and LAI can then be used together with a canopy photosynthesis model to determine the foliage (LAI) increment for each day of growth.
Nutritive value
Feeding value of pasture is the product of voluntary feed consumption and the digestibility of nutrients consumed. The increase in CP percentage as PPFD declined (Fig. 9) may be attributed to either a decrease in photosynthates, with a consequent rise in the nitrogen concentration, or to an increase in soil organic matter mineralisation under trees that provided more nitrogen for grass uptake (Wilson and Ludlow 1991). The major effects of different light transmission intensity on the mean N concentration of six tropical grasses was in plants under moderate shade (68% light transmission) compared with pastures grown at 100% light transmission, but differences between species were much larger than any shade effects observed in those studies (Norton et al. 1991). In our study, there was a seasonal decrease in CP percentage when water stress was severe (soil moisture < 18%) with less difference between open and shaded treatments. Similarly, Whitehead (1995) reported that dry conditions may retard both pasture uptake and soil mineralisation of N during the season.
There were small differences between open and shaded treatments for organic matter digestibility (OMD) (Fig. 10). Many results have shown that in vitro digestibility of grasses is reduced under low light intensity and this is associated with reduced digestibility of cell wall constituents (Hight et al. 1968). However, other studies have obtained less conclusive results with both increases and decreases in digestibility of herbage grown in a shaded environment (Wilson 1988). Cell walls are the major fraction available for digestion and it is important to determine whether shade affects their composition (Norton et al. 1991). Shading usually reduces the total non-structural carbohydrate of grasses, but has variable (positive and negative) effects on cell wall content and composition, lignin and in vitro digestibility of plant dry matter (Wilson 1988). According to Hight et al. (1968), soluble carbohydrate content decreased by 3.7% units and dried forage digestibility by 0.6–3.6% units during a short shading period of time (2–3 days at 22% light transmission). However, Norton et al. (1991) found a small non-significant decrease in soluble carbohydrate and a decrease in in vitro dry matter digestibility.
In the present study, the mean annual macro-nutrient concentrations in leaves increased as the PPFD level declined (Table 5). One reason for this may be because heavily shaded pasture (i.e. less than 30–40% light transmission) has low dry matter contents or higher water contents (Reynolds 1995). In Vanatu, the dry matter percentage of shaded T-grass (Paspalum conjugatum) Berg. is often below 18%, especially under heavy shade. Burton et al. (1959) reported that severe shade reduced the yield of DM by about one-third, the concentration of P, Ca and Mg in Coastal Bermudagrass increases by 25–50%.
Furthermore, active mineralisation of soil organic matter by pine roots or their mycorrhiza was suggested as a likely mechanism accounting for elevated inorganic and available nutrient levels under pines in some New Zealand grassland soils and therefore more nutrient availability for pastures in a silvopastoral system (Davis 1994). Hawke and O’Connor (1993) reported that while soil potassium remained relatively constant (average of 7.8 mg kg−1 at 400 sph), magnesium, sulphate-sulphur and phosphorus levels tended to increase at higher tree stocking rates (14 mg kg−1, 20 mg kg−1 and 50 mg kg−1, respectively under an 18 year-old radiata pine stand). These changes in topsoils were attributed to nutrient uptake, organic matter mineralisation, and the effect of organic acids under the radiata pine stand.
There was an interaction between treatments and time (seasons). Shade under trees showed less difference during severe drought in both summers and autumn 2001. Thus, macro-nutrient concentrations were higher (except Ca) in spring and winter (soil moisture > 24%) compared with summer (soil moisture < 18%). In general, water stress may reduce the capacity of roots to absorb nutrients if it reduces transpiration and growth, and therefore plant demand for nutrients (Lambers et al. 1998). While concentration of P in grasses is reduced by drought, the effect on other elements is less consistent (Rahman et al. 1971). The increase in Ca concentration in summer for the drought-resistant cultivar KM2 cocksfoot population could reflect a greater density of roots in deeper soil horizons (Volaire and Thomas 1995).
Macro-nutrient concentrations increased as PPFD declined The minimum values measured mainly in the open treatment (100% transmissivity) satisfy the demand for optimum animal feed requirements. According to Kemp et al. (1999) the optimum concentration of mineral elements in temperate pasture species for sheep growth are P = 2.0, S = 1.15, K = 3.6, Ca = 2.9 and Mg = 1.2 g kg−1 DM. However, according to Kemp et al. (1999) leaf nutrient concentrations in the present study were below the optimum for pasture growth mainly for the open treatment and during summer drought (P = 4.0, S = 3.2, K = 25.0, Ca = 3.0 and Mg = 2.0 g kg−1 DM).
Given CP percentage and macro-nutrient concentrations (where all pasture samples were higher than the optimum required for sheep growth) increased slightly with shade level, digestibility values were not greatly influenced by shade, and clover content in cocksfoot pastures were low (<7%), it seems likely that shaded pastures would limit animal production primarily due to lower DM production rates. This may lower pre-grazing pasture mass and reduce bulk density and bite size from the etiolated pasture. This is consistent with Peri et al. (2001) who reported for the same experimental area that apparent intakes were reduced by 0.3 kg DM animal−1 d−1 for lambs grazing cocksfoot pastures under trees. This was attributed to a reduction in pasture bulk densities. Thus, grazing animals had greater potential intake in open pastures than under trees because the amount harvestable in a single bite for livestock grazing lower density pasture is smaller (Gong et al. 1996). Reduction in bite size leads to reduced daily pasture intake and LWG because the animal normally cannot sufficiently compensate by increasing the rate of biting or grazing time per day. Alternatively, long grazing rotations could be used under shaded pastures to provide time to accumulate sufficient pasture mass. However, problems from longer spelling times would be: (i) taller pasture and increased reproductive development that results in reduced bulk density, and (ii) older forage of lower nutritive value.
In summary, while nutritive value of cocksfoot pastures under fluctuating light regimes increased as the PPFD level declined, it appears that heavily shaded dominant temperate pastures in silvopastoral systems limit animal production per hectare through lower DM production rates and per animal through reduced pre-grazing pasture mass of lower bulk density from the etiolated pasture.
References
Anderson GW (1978) Productivity of crops and pastures under trees. In: Howes KMW, Rummery RA (eds) Integrated agriculture and forestry. CSIRO, Division of Land Resource Management, Perth, pp 59–63
Braziotis DC, Papanastasis VP (1995) Seasonal changes of understorey herbage yield in relation to light intensity and soil moisture content in a Pinus pinaster plantation. Agroforest Syst 29:91–110
Burton GW, Jackson JE, Knox FE (1959) The influence of light reduction upon the production, persistence and chemical composition of Coastal Bermudagrass, Cynodon dactylon. Agron J 51:537–542
Casal JJ, Deregibus VA, Sanchez RA (1987) Tillering responses of Lolium multiflorum plants to changes of red/far-red ratio typical of sparse canopies. J Exp Bot 38:1432–1439
Cossens GG (1984) Grazed pasture production under Pinus radiata in Otago. In: Bilbrough GW (ed) Proceedings of a Technical Workshop on Agroforestry. Dunedin, New Zealand, May 1984
Davies A (1988) The regrowth of grass swards. The grass plant – its form and function. In: Jones MB, Lazenby A (eds) The grass crop: The physiological basis of production. Chapman and Hall, London, p 85
Davis MR (1994) Topsoil properties under tussock grassland and adjoining pine forest in Otago, New Zealand. New Zeal J Agr Res 37:465–469
Deregibus VA, Sanchez RA, Casal JJ (1983) Effects of light quality on tiller production in Lolium spp. Plant Physiol 72:900–902
Devkota NR, Kemp PD, Hodgson J (1997) Screening pasture species for shade tolerance. Proc Agron Soc New Zeal 27:119–128
Devkota NR, Kemp PD, Valentine I, Hodgson J (2000) Shade tolerance of pasture species in relation to deciduous tree, temperate silvopastoral systems. Proc Agron Soc New Zeal 30:101–107
Duru M, Lemaire G, Cruz P (1997) Grasslands. In: Lemaire G (ed) Diagnosis of the nitrogen status in crops. Springer, New York
Garnier E, Roy J (1988) Modular and demographic analysis of plant leaf area in sward and woodland populations of Dactylis glomerata and Bromus erectus. J Ecol 76:729–743
Gong Y, Hodgson J, Lambert MG, Gordon IL (1996) Short-term ingestive behaviour of sheep and goats grazing grasses and legumes. 2. Quantitative relationships between sets of sward and ingestive behaviour variables. New Zeal J Agr Res 39:75–82
Hawke MF, Knowles RL (1997) Temperate agroforestry in New Zealand. In: Gordon AM, Newman SM (eds) Temperate agroforestry systems. CAB International, UK, Wallingford
Hawke MF, O’Connor MB (1993) Soil pH and nutrient levels at Tikitere agroforestry research area. New Zeal J For Sci 23:40–48
Hight GK, Sinclair DP, Lancaster RJ (1968) Some effects of shading and of nitrogen fertiliser on the chemical composition of freeze-dried and oven-dried herbage, and on the nutritive value of oven-dried herbage fed to sheep. New Zeal J Agr Res 11:286–302
Joshi MR, Lucas RJ, Sedcole JR, Pollock KM, Moot DJ, Mead DJ (1999) Shading effects of Pinus radiata on productivity and feeding value of orchardgrass pasture. Agroforest Forum 9:17–19
Kemp PD, Condron LM, Matthew C (1999) Pastures and soil fertility. In: White J, Hodgson J (eds) New Zealand pastures and crop science. Oxford University Press, Auckland
Kephart KD, Buxton DR (1993) Forage quality responses of C3 and C4 perennial grasses to shade. Crop Sci 33:831–837
Korte CJ, Chu ACP, Field TRO (1987) Pasture production. In: Nicol AM (ed) Feeding livestock on pasture. New Zealand Society of Animal Production, New Zealand
Lambers H, Chapin III FS, Pons TJ (1998) Plant physiological ecology. Springer-Verlag, New York
Morton J, Roberts AHC, Edmeades DC (1994) Fertiliser use on sheep and beef farms. New Zealand Fertiliser Manufacturers Research Association, Auckland
Norton BW, Wilson JR, Shelton HM, Hill KD (1991) The effect of shade on forage quality. In: Shelton HM, Stur WW (eds) Forages for plantation crops. ACIAR, Canberra
Padmore JM (1990) Animal feed. In: Helrich K (ed) Official methods of analysis. Association of Official Analytical Chemists, Arlington
Peri PL Varella AC, Lucas RJ, Moot DJ (2001) Cocksfoot and lucerne productivity in a Pinus radiata silvopastoral system: a grazed comparison. Proc New Zeal Grassland Assoc 63:139–147
Peri PL, McNeil DL, Moot DJ, Varella AC, Lucas RJ (2002) Net photosynthetic rate of cocksfoot leaves under continuous and fluctuating shade conditions in the field. Grass Forage Sci 57:157–170
Quin BF, Woods PH (1976) Rapid manual determination of sulphur and phosphorus in plant material. Soil Sci Plant Anal 7:415–426
Rahman AAA, Shalaby AF, El Monayeri MO (1971) Effect of moisture stress on metabolic products and ion accumulation. Plant soil 34:65–90
Reynolds SG (1995) Pasture-Cattle-Coconut Systems. FAO, RAPA (Regional Office for Asia and the Pacific), Bangkok
Robson MJ, Ryle GJA, Woledge J (1988) The grass plant – its form and function. In: Jones MB, Lazenby A (eds) The grass crop. Chapman and Hall, London
Sheehy JE, Cooper JP (1973) Light interception, photosynthetic activity, and crop growth rate in canopies of six temperate forage grasses. J Appl Ecol 10:239–250
Sibbald AR, Griffiths JH, Elston DA (1991) The effects of the presence of widely spaced conifers on under-storey herbage production in the U.K. Forest Ecol Manag 45:71–77
Smith H (1982) Light quality, photoperception, and plant strategy. Ann Rev Plant Physiol 33:481–518
Volaire F, Thomas H (1995) Effects of drought on water relations, mineral uptake, water-soluble carbohydrate accumulation and survival of two contrasting populations of cocksfoot (Dactylis glomerata L.). Ann Bot-LONDON 75:513–524
Whitehead DC (1995) Grassland nitrogen. CAB International, Wallingford
Wilson JR (1988) Ecological constraints to production and nutritive quality of pastures under tree crops. In: Livestock-tree cropping production systems. Proc FAO/MARDI Int Workshop, 5–9 December 1988, Serdang, Malaysia
Wilson JR, Ludlow MM (1991) The environment and potential growth of herbage under plantations. In: Shelton HM, Stur WW (eds) Forages for plantation crops. ACIAR, Canberra
Wong CC (1991) Shade tolerance of tropical forage: a review. In: Shelton HM, Stur WW (eds) Forages for plantation crops. ACIAR, Canberra
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Peri, P.L., Lucas, R.J. & Moot, D.J. Dry matter production, morphology and nutritive value of Dactylis glomerata growing under different light regimes. Agroforest Syst 70, 63–79 (2007). https://doi.org/10.1007/s10457-007-9029-x
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-007-9029-x