
Abstract
Strong interest in organic produce has warranted great demand in organically produced forages. However, extremely limited research has focused on organic forage production and its ecological contribution. Grass-legume biculture has been suggested to reduce dependency on external N inputs, control weed encroachment, and achieve better soil health. A 2-year study was designed as a conservative organic system with minimum tillage and no external inputs to investigate productivity, botanical composition, nutritive value, mineral concentration, economic benefit, and soil total carbon changes in five certified forage biculture systems; including (1) one annual rotation of winter wheat-Austrian winter pea biculture followed by sorghum-sudangrass hybrid-cowpea biculture, and four perennial systems of (2) bermudagrass-alfalfa, (3) tall fescue-white clover, (4) old world bluestem-sainfoin, and (5) orchardgrass-alfalfa bicultures. Annual systems indicated the greatest productivity (12.7 Mg ha−1) followed by old world bluestem-sainfoin (11.3 Mg ha−1) and bermudagrass-alfalfa (10.3 Mg ha−1) bicultures; but with very limited nutritive value advantages, low mineral concentration, and poor economic return. Perennial systems, particularly old world bluestems-sainfoin biculture, indicated great adaptation, soil carbon enhancement capacity. Monthly biomass production was influential on several nutritive value indices (P < 0.05, R2 ≈ 0.5) within many forage systems. No treatment effects were detected on soil total carbon (P = 0.27), but significant year-wise increase was found under old world bluestems-sainfoin biculture (P < 0.05). Soil moisture levels were affected by treatments and correlated well to botanical composition. Path analysis indicated that Radiation Use Efficiency is the key driver for determining forage yield in the temperate environment.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Organic farming is an alternative agricultural system developed and adopted by producers in the early 1990s (Francis and Van Wart 2009), which encompasses a suite of agricultural practices designed to eliminate inorganic chemical inputs to sustain the Earth’s environment. Meanwhile, land used for permanent forage production and animal grazing has increased dramatically in the past decade. To date, grassland-based agriculture for forage production and grazing livestock ranks the first among all land uses globally (Willer and Lernound 2016). On average, ruminant animals consume more than 80% of diet as forages or even more when managed organically (Mitchell and Nelson 2003). Additionally, many organic program rules mandate a minimum level (e.g., > 30% in the US) of forage-based dry matter intake of a ruminant animal within a single grazing season. Therefore, high-quality forage is the key component for organic livestock/dairy production, and there is rapidly growing demand for certified forage products (e.g., hay, silage, etc.). However, the scarcity of research-based information on organic forage systems has been the major bottleneck for the development and expansion of the production scale and adoption by producers (Oberholtzer et al. 2012). For example, several studies evaluated organic forage production as a cover cropping component from a main grain crop system, thus, providing little information on the establishment, productivity, and nutritive value of the selected forage species (Delate and Cambardella 2004; Cavigelli et al. 2008). Meanwhile, many studies focused solely on a particular forage species that is of significant importance to organic dairy, such as alfalfa (Medicago Sativa L.; Mahoney et al. 2004; Archer et al. 2007; Delbridge et al. 2011). The knowledge gaps related to cool/warm season grasses as well as grass-legume biculture selection, management, and persistence remain large; particularly in the temperate subtropical regions, where climatic conditions tend to be variable and soil properties differ greatly.
One of the biggest concerns of organic forage production is how to maintain productivity and quality (Tu et al. 2006; Dawson et al. 2008). Previous studies have verified that grass-legume/forb bicultures could help enhance forage nutritive value (e.g., protein content) and biomass production through legume-associated symbiotic N2 fixation (Birkhofer et al. 2008; Cui et al. 2013) or enhanced P availability (Hinsinger 2001). Another challenge for organic crop production is how to control weeds with no inorganic chemical inputs and minimized economic hardship (Francis and Van Wart 2009; Liebman and Davis 2009). Certain small-grain annual forages such as cereal rye (Secale cereale L.) are well known for their weed suppression capacity through the production of allelopathic secondary metabolites (Li et al. 2013). Additionally, diversified forage systems (e.g., mixtures of grasses and legumes, cool-season and warm-season, perennial and annual, etc.) tend to provide better weed control than monocultures (Schoofs and Entz 2000; Sanderson et al. 2012). However, there is almost no published information relating to the dynamics of botanical composition and weed suppression capacity of organically managed perennial forage systems, and botanical composition is usually the main driver for changes in forage nutritive value and mineral status (Cui et al. 2014). Furthermore, given soils in many temperate regions are highly weathered with high acidity, and organic nutrient sources are usually hard to find and could incur careful scrutiny during the certifiers’ inspection process, N credit provided by legumes and proper management strategy might be the most effective way for enhancing nutrient availability and recycling in organic forage systems (Franzluebbers 2010). In a most recent transitional forage production study, Inwood et al. (2015) found that annual forage systems could provide excellent productivity and comparable soil carbon contribution compared with perennial systems in the southeastern US. However, no information was provided regarding forage nutritive value and radiation use efficiency, which is a crucial index evaluating forage productivity and cultivar selection especially in the radiation-limiting southeastern US environment. In another study, Gelley et al. (2016) systematically investigated nutritive value of several forage systems but none of which were managed organically. Likewise, Nave et al. (2020) evaluated two cowpea [Vigna unguiculata (L.) Walp.]-based grass-legume bicultures, and highlighted the nutritive contribution offered by legumes while managed conventionally. Furthermore, Cui et al. (2014) indicated that differences in root structure and root profile depth of the dominating forage groups within grass-legume bicultures can have significant impact on soil C and moisture status, which could induce feedback effects on forage productivity in water-limiting environment. However, information relating to organically managed biculture forage systems in the temperate climate transition areas remains scanty.
Considering the extremely limited information on biomass production, forage nutritive value, and soil carbon status associated with certified organic forage systems in the temperate region, the main objectives of this research were to investigate the responses of different parameter groups across five grass-legume biculture organic forage systems without external nutrient sources during a two-year period. The parameter groups include: (1) productivity-related parameters, including biomass, botanical composition, radiation use efficiency (RUE); (2) quality-related parameter-dynamics of forage nutritive value of the mixed swards throughout the growing season; (3) soil-related parameters, including changes in soil moisture content and total carbon; (4) economic responses-economic benefits, and (5) quantity-quality interactions indicated as correlation between monthly cutting and nutritive value indices. Particularly, four perennial grass-legume biculture systems and one annual biculture rotation system were selected in this study. It was hypothesized that annual systems should provide the most forage yield and weed suppression capacity but with limited soil carbon contribution and economic return due to greater labor input and dependency on tillage. Perennial systems are expected to provide less forage yield compared to annual systems due to slower germination and longer time for establishment, but their economic profitability and nutritive value might outcompete annual systems. Soil moisture and total C content should respond differently towards different forage species, mainly driven by root architecture and length differences of dominating forage groups. Common linear correlations, such as monthly biomass production vs. protein content, have been identified in many forage systems (Philipp et al. 2005; Cui et al. 2013, 2014), but other potential correlations (e.g. protein vs. fiber content; digestibility vs. sugar content) and corresponding levels of significance remain unclear.
Materials and methods
Site description
The study was conducted at the Middle Tennessee State University Experiential Research and Teaching Farm Laboratory at Lascassas, TN (35° 53′ N and 86° 16′ W). The research site at MTSU is located on a Hillwood gravelly silt loam (25% clay, 42% of sand, and 33% of silt on average). The middle Tennessee region (the Koppen System Type C-moist, mid-latitude climates with moist hot summers, mild winters, and greater precipitations in spring and fall) represents a typical climate transition zone-type weather with average long-term precipitation and daily temperature of 1397 mm year−1 and 15 °C, respectively (Li et al. 2021). The entire organic field was measured as approximately 0.61 ha (surrounded by a 20-m wide buffering zone) with the geometric center located at the latitude of 35° 53′ 2.45′′ N and the longitude of 86° 16′ 23.55′′ W, which is a part of a large hay field that was originally established in 2008 and mainly dedicated for cool-season forage hay production without application of chemical fertilizers, pesticides, or herbicides. The entire organic site (Entity ID#: 2990G) was certified in 2018 under the US National Organic Program 7 CFR Part 205 and had remained certified status since then. Weather conditions are continuously monitored and recorded using a scientific weather station system consisting of an HMP60 probe (Campbell Scientific, Logan, UT 84321) for measuring relative humidity and air temperature, a CS655 multiparameter smart sensor (Campbell Scientific, Logan, UT 84321) installed at 5-cm depth for measuring soil moisture and temperature, a 014A anemometer (Met One Instrument, Grants Pass, OR 97526) for wind speed, a TE525 tipping bucket rain gauge (Texas Electronics, Dallas, TX 75237) for daily precipitation, and a CR-1000 data logger (Campbell Scientific, Logan, UT 84321) for equipment control, data retrieval and storage purposes.
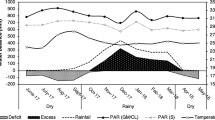
The evolution of average daily air and soil temperature is depicted in Suppl. Fig. S1a. Daily cumulative precipitation pattern and average soil moisture content are indicated in Suppl. Fig. S1b. Successful plant establishment was observed across all plots after fall planting (Suppl. Fig. S1c). The average daily air and soil temperatures were 14.9 and 15.2 °C in 2018, and 15.6 and 16.0 °C in 2019; respectively. Cumulative precipitation was 1488 mm in 2018 and 1631 mm in 2019. Soil moisture level agreed well with the precipitation pattern across two years.
Crop management and experimental design
This experimental layout was a randomized complete block design with four blocks and repeated measures. Treatments included five organic forage systems with one annual forage crop rotation system (cool-season and warm-season grass-legume annual bicultures) and four perennial systems (cool-season and warm-season grass-legume bicultures). The annual forage crop rotation system was composed by double cropping (simultaneously planting) winter wheat (Triticum aestivum L.) and Austrian winter pea (Pisum sativum L.) in the fall followed by double cropping sorghum-sudangrass hybrid [Sorghum bicolor × S. bicolor var. sudanense (Piper) Stapf.] and cowpea in the summer within the same plot. The four forage perennial systems include: bermudagrass [Cynodon dactylon (L.) Pers.]-alfalfa, tall fescue (Festuca arundinacea L.)-white clover (Trifolium repens L.), old world bluestem [Bothriochloa bladhii (Retz) S.T. Blake]-sainfoin (Onobrychis viciifolia Scop.), and orchardgrass (Dactylis glomerata L.)-alfalfa bicultures. Each grass-legume biculture was designed to help address unique advantages and challenges in an integrated forage-livestock or hay production system. For example, the superior biomass production of the selected annual rotation was previously documented (Inwood et al. 2015), however, the nutritive value dynamics and economic benefit under organic production have not been studied yet. Bermudagrass was used as a common warm-season forage species for gap-filling the summer slump of cool-season forage production, however, its great dependency on inorganic N inputs and vulnerability towards insect attack make its performance questionable while managed organically. Meanwhile, bermudagrass-alfala mixtures had indicated great persistency and N credit in the mixed sward under our climate conditions (Quinby et al. 2020) and thus were selected for this study. Tall fescue and white clover both have great forage productivity during cool-weather conditions and excellent tolerance towards continuous grazing, but summer production could be greatly compromised by heat stress, particularly under organic management. Old world bluestem-sainfoin biculture was found to enhance soil carbon and adequate nutritive value in a semiarid environment (Cui et al. 2013, 2014). However, their adaptation in the temperate environment has rarely been documented. Finally, orchardgrass and alfalfa both provide superb forage quality, but both are not well suited for frequent defoliation. Thus, the changes of their productivity and nutritive value (e.g. protein and fiber concentration) of the sward from monthly biomass production were of great interest particularly under organic systems.
A total of twenty individual plots (3.6 × 10 m2) were established. The entire field was disk-tilled on Oct. 13th of 2017 using a Disk Tiller (Kodiak 5800GR-72, Kodiak Manufacturing Inc. Charleston, TN 37310, USA) to a depth of 10–15 cm. Before seeding, all seeds of the legume species were pre-inoculated using corresponding certified organic Rhizobia spp. sources. A specially designed forage drill seeder (Eco-Drill KED-72, Kasco Manufacturing CO. Shelbyville, IN 46176, USA) was used for planting. All cool-season grass perennial systems were planted on 20 Oct. 2017 at a seeding rate of 16.8, 4.5, 13.4, and 16.8 kg ha−1 for tall fescue (cultivar KY-31, untreated), white clover (cultivar Alice, certified organic), orchardgrass (cultivar Echelon, untreated) and alfalfa (cultivar WL358LH, certified organic); respectively. Warm-season grass perennial systems were planted on 22 Mar. 2018 at a seeding rate of 11.2, 16.8, 11.2, and 16.8 kg ha−1 for bermudagrass (untreated variety), alfalfa, old world bluestem (cultivar WW-B Dahl, untreated), and sainfoin (cultivar Remont, untreated); respectively. For annual systems, winter wheat (cultivar L334, certified organic) and Austrian winter pea (untreated variety) were planted on 31 Oct. 2017 and 17 Oct. 2018 at a seeding rate of 134.4 and 132.3 kg ha−1, respectively; and sorghum-sudangrass hybrid (cultivar AS6501, untreated) and cowpea (cultivar Iron & Clay, untreated) were planted on 30 May 2018 and 29 May 2019 at a seeding rate of 28.0 and 16.8 kg ha−1, respectively. All annual system plots were tilled before planting to a depth of 10–15 cm. All seeds were either certified organic or untreated, and cultivars were chosen based on regional adaptation and seed availability.
The whole experiment was designed as a conservative system with minimum tillage and no external organic inputs. The whole system solely depends on legumes for soil N enhancement. No irrigation water was used due to the ample precipitation in the local environment. No herbicides or pesticides were used during the entire experimental period. All organic crop production management was conducted according to the guidelines and standards established by the USDA Organic Regulations and the National Organic Standards Boards (https://www.ams.usda.gov/rules-regulations/organic/nosb).
Plant sampling and processing
Forage biomass sampling schedule was organized based on the following rationale: since all forage treatment systems involved perennial or annual bicultures or biculture rotations and grass component clearly dominated the mixtures, most samples were taken at the boot stages of grasses; which was usually between the early-bud to bloom stages of companion legumes, commonly adopted by forage producers in the local region (Inwood et al. 2015; Mitchell and Nelson 2017). Particularly, winter wheat and Austrian winter pea bicultures were sampled once per year (22 Apr. 2018 and 25 Apr. 2019). Sorghum-sudangrass hybrid and cowpea bicultures were sampled three times per year (7 Jul., 10 Aug., 15 Sep., 2018; 12 Jul., 15 Aug., and 20 Sep., 2019). All perennial bicultures were sampled monthly from May to Sep. in each year (21 May, 10 Jun., 7 Jul., 10 Aug., and 15 Sep., 2018; 19 May, 11 June, 12 Jul., 15 Aug., and 20 Sep., 2019) to mimic the common hay harvest interval adopted in our area (29–31 days). At each sampling, above-ground forage biomass were taken from two, 1-m2 quadrats at a 5-cm cutting height within each treatment replicate. Considering the fact that weeds encroachment could be severe in organic systems, all biculture samples were hand separated and sorted in the field into three groups, including sown grasses, sown legumes, and non-sown weeds; and were later placed in separate paper bags. Additionally, to better evaluate weed encroachment, visual estimation of main weed species was also performed from within each quadrat at each sampling. Immediately after the completion of each sampling, the entire sampled plot was harvested for hay to induce biomass production (a one-time cut was imposed at the beginning of each month with biomass regrowth evaluated in the following month). All samples were dried in a research convection oven at 60 °C for 48 h, then recorded for forage biomass production on a dry matter basis. Annual cumulative forage biomass production was used for calculating total forage yield. Specifically, for annual systems, forage yield was calculated based on the sum of Apr., Jul., Aug., and Sep. forage biomass samples. For perennial systems, it was calculated based on monthly biomass samples collected from May to Sep. No weeds were included in the yield calculation.
All dried forage samples (excluding weed samples) were recombined (grasses plus legumes) according to plot number and ground using a Wiley mill (Comeau Technique Ltd., Vandreuil-Dorion, Quebec, Canada) to pass a 1-mm screen before forage nutritive value analysis. Ground samples were then scanned on a Unity SpectraStar US-2600-XTR (Milford, MA) near-Infrared Spectrometer (NIRS) using the 2018 Grass Hay calibration model developed by the NIRS Feed and Forage Consortium (Hillsboro, WI). The Global H statistical test compared the samples against the model and other samples within the database for accurate results, where all forage samples fit the equation with H < 3.0 and are reported accordingly (Murray and Cowe 2004). A selected subset of samples was used for calibration by conventional wet chemistry procedures (Van Soest 1963; Goering and Van Soest 1970). A coefficient of determination value R2 = 0.90 was obtained for neutral detergent fiber (NDF) calibration and R2 = 0.94 was obtained for acid detergent fiber (ADF). The in vitro dry matter digestibility (IVDMD) was calculated based on ADF and NDF (Van Soest 1963). Concentrations of Ca, P, K, and Mg were determined using an inductively coupled plasma atomic emission spectrophotometer following chemical digestion using the 3:1 HNO3:HCLO4. Accuracy was ensured by analyzing the National Bureau of Standard Samples (Apple Leaves No. 1515 and Tomato Leaves No. 1573A) during mineral analysis.
Radiation use efficiency
Incoming solar radiation was measured using an LI-190R Quantum Sensor (Li-Cor Inc., NE, USA) with a CR-1000 data logger (Campbell Scientific, Logan, UT 84321) powered by a 12-V battery every two weeks. Six measurements (three above and three below the canopy) were taken during the growing season of each forage system at the noon during the selected sunny days. The sensor placement protocol for forage biculture systems closely followed the techniques described by Coll et al (2012). Accumulated intercepted photosynthetic active radiation (PAR) was calculated for all treatments on the basis of daily values (Coll et al. 2012), assuming the north–south row orientation and symmetrical west–east canopy layout around the zenith (Tsubo and Walker 2002). Particularly, the fraction of intercepted PAR was calculated as 1 – I/I0 three hours before the solar zenith (Rzenith-3 h) and at the solar zenith (Rzenith), where I0 and I represent the incident PAR above the plant canopy and immediately above the soil horizon. Four measurements were taken from each plot and averaged. Daily weighted fraction of intercepted PAR was calculated using 2Rzenith-3 h × I0 at zenith-3 h + Rzenith × I0 at zenith. The fraction of intercepted PAR between measurements were calculated using linear interpolation. Cumulative PAR was calculated between two adjacent cuttings. Finally, RUE was calculated as the ratio between above-ground forage dry matter production and cumulative intercepted PAR.
Soil sampling and processing
Within-season soil samples were taken twice per year (late summer and late fall) during sunny days from each treatment plot to evaluate the soil total carbon status. At each sampling, three soil cores (1.75-cm internal diameter) were collected from two different sampling depths (0–5 and 5–15 cm). The baseline soil samples were composites of ten soil cores collected at two different depth from the entire hay field before imposing any treatment layout in Oct. of 2017. Soil inorganic N (ammonium-N and nitrate–N) was determined colorimetrically on a microplate spectrophotometer (Sims et al. 1995) and major plant available nutrients were determined after Mehlich-1 extraction on an Inductively Coupled Plasma-Optical Emission Spectroscopy. The soil was found to be acidic, with limiting inorganic N, low P, high K, and non-limiting Ca, Mg, Mn, and Zn for supporting general forage production (Suppl. Table S1). All soil samples were air dried at room temperature and then passed through a 2-mm sieve to remove large fresh organic plant debris. To measure total soil organic carbon concentration, all samples were pulverized and analyzed using combustion method using a Vario MAX cube (Elementar, Langenselbold, Germany). Final concentration of C was calculated based on exact weight of analyzed sample weight. Meanwhile, a different set of samples were taken at two depth ranges (0–50 cm and 50–100 cm) to measure soil gravimetric water content (GWC) using a 6.35-cm diameter and 121-cm long soil auger. The wet soil weight was measured using a digital scale immediately after sampling and dry weight was recorded after drying at 105 °C for 48 h.
Statistical analysis and modeling
The MIXED procedure in SAS release 9.4 (SAS Institute 2018) was used to analyze the treatment effects based on a randomized complete block design with repeated measures. Particularly, a linear mixed effect model with different response variables (yield, monthly production, nutritive value indices, RUE, TOC, and Soil GWC) affected by treatment (forage systems) with year and season as fixed effect and block as random effect; because different weather conditions and plant physiological conditions (e.g. maturity and senescence) within each season were of interest and were expected to greatly affect forage biomass production and nutritive value. Additionally, annual precipitation and distribution pattern tend to vary greatly in the climate-transition zones, and this weather pattern could have significant impacts on the growth and interactions between grasses and legumes. Thus, year was analyzed as a fixed effect. Total forage yield was analyzed on a yearly basis, therefore, no seasonal factor was included. The REPEATED statement in the MIXED procedure was used to control for autocorrelation of observations over time. Particularly, the first-order autoregressive AR(1) covariance structure was selected as variance–covariance structure model with block factor corrected by season nested within each year. Mean separation was produced based on Tukey’s HSD method using the PDMIX800 macro, which separate and label the means using letter groupings based on pair-wise comparison created by the PDIFF option with the LSMEANS statement with Tukey Adjustment (Saxton 1998).
The structural equation modeling (SEM), which integrates factor and path analysis techniques based on univariate/multivariate statistical analysis was used for evaluating the key direct and indirect effects of different production (RUE, grass-to-legume ratio) and environmental (SWC) variables on biomass yield. In particular, the “lavaan 0.6-7” version package based on R 3.2.2 programming language was used for this modeling task (Rosseel 2012). Model performance was evaluated based on Chi2 statistics, Akaike Information Criterion (AIC) values, and the Standard Root Mean Square Residual (SRMSR) using the maximum-likelihood estimator embedded in the “Lavaan” Package.
Results
Forage biomass production, botanical composition, and forage yield
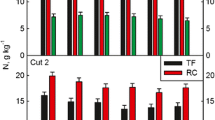
Yearly forage yield was calculated as the accumulation of aboveground forage biomass production over each year (Fig. 1). It was significantly affected by forage systems (P < 0.001), years (P < 0.001), but no two-way interactions were found (P = 0.23; Table 1). Therefore, forage yield data was later presented by the two-year averages. Forage biomass production at each sampling date included sown grasses and legumes. It was significantly affected by forages system (P < 0.001) but not affected by year (P = 0.13) or year-treatment interaction (P = 0.46). Date of sampling (monthly cutting) had significant effect on biomass production and interacted with forage system (P < 0.05). No year-by-month (P = 0.41) or three-way (P = 0.31) interactions were detected, thus, data were later presented by month but averaged over years. Averaged over the years, annual systems had greater forage yield than perennial systems (36%). Mean April forage biomass production for winter wheat-Austrian winter pea biculture was around 4.0 Mg ha−1 with 83.0, 12.4, and 4.5% of winter wheat, Austrian winter pea, and unsown species; respectively (Fig. 2; Table 2). For the May cutting, no differences were detected among forage systems, and the average biomass production across all biculture systems was around 1.2 Mg ha−1 with 65.5, 10.9, and 23.5% of average sown grasses, sown legumes, and weeds (Table 2); respectively. Likewise, average June production was around 1.5 Mg ha−1 with 66.7, 8.9, and 24.2% of average sown grasses, sown legumes, and weeds; respectively. Sorghum-sudangrass hybrid-cowpea biculture production started in July and was significantly greater than both tall fescue-white clover (1.0 Mg ha−1 more) and orchardgrass-alfalfa bicultures (0.9 Mg ha−1 more). In August, bermudagrass-alfalfa biculture yielded more biomass than sorghum-sudangrass hybrid-cowpea (1.1 Mg ha−1 more) or tall fescue-white clover biculture (1.3 Mg ha−1 more). From July to September, annual systems as well as tall fescue-white clover bicultures consistently had low percentage of weeds (8.6 and 8.5%, respectively). Bermudagrass-alfalfa biculture had high weed percentages in July (31.9%) and September (11.4%) and low percentage in August (14.1%). Old world bluestem-sainfoin biculture indicated great weed suppressing capacity in July (5.2%) and August (4.0%), but decreased in September (24.2%). The most dominant weed species were evaluated according to each treatment-year averaging over all sampling months. Annual mean weed species diversity and top-three ranking dominating species estimated using ground coverage from each quadrat area are shown in Table 3.

Average annual forage dry matter yield as affected by different forage systems at Lascassas, TN, reported as average values across 2018 and 2019 growing seasons. Means without a common letter differ at a P = 0.05 significance level. Standard errors (SE) are reported based on pooled estimates of population standard deviation used by the Tukey’s HSD procedure

Average monthly initiation of forage biomass and botanical composition (grass, legumes, or weeds) affected by different forage systems at Lascassas, TN. in 2018 and 2019 growing seasons. Means without a common letter differ at a P = 0.05 significance level. Standard errors (SE) are reported based on pooled estimates of population standard deviation used by the Tukey’s HSD procedure
Forage nutritive value
Forage nutritive value was analyzed using the identical statistical models as for forage biomass production, because all biomass samples (except for weeds) were later processed for the NIRS analysis following dry weight recording. The results were presented as two-year average values due to the lack of year-treatment interactions (P > 0.05). Mean forage CP concentration of annual bicultures was close to 90.8 g kg−1 in April (winter wheat-Austrian winter pea mixture), gradually increased along the growing season after transitioning to sorghum-sudangrass hybrid-cowpea biculture, and peaked at the August production to 131 g kg−1 (Table 4). Tall fescue-white clover biculture generally had greater CP concentration than other bicultures (except for the September production, when tall fescue-bicultures and annual systems showed no differences). Annual systems consistently had similar CP concentration to other perennial bicultures, except tall fescue-white clover mixtures. Annual systems had greater ADF concentration than bermudagrass-alfalfa and tall fescue-white clover bicultures in July, and both old world bluestem-sainfoin and orchardgrass-alfalfa bicultures had greater ADF concentration than tall fescue-white clover biculture. For NDF concentration, bermudagrass-alfalfa and old world bluestem-sainfoin bicultures were greater than tall fescue-white clover and orchardgrass-alfalfa bicultures. The IVDMD results indicated that the sward of tall fescue-white clover biculture was more digestible than orchardgrass-alfalfa biculture in both May and June; and was more digestible than any other forage system in July, but comparable to annual system in September and both the annual system and orchardgrass-alfalfa biculture in August. Tall fescue-white clover biculture had greater fructan concentration than all other perennial bicultures in May and June. Lignin concentration follows the trend of NDF and ADF pattern and the detailed results are depicted in Table 4.
The correlation analysis between different nutritive value indices and biomass production are presented by forage systems as depicted in Suppl. Fig. S2 to S6. Briefly speaking, for annual systems, significant correlation (P < 0.05 and R2 ≈ 0.5) was found between ADF and NDF, ADF and IVDMD, ADF and fructan, ADF and lignin, NDF and IVDMD, NDF and fructan, as well as IVDMD and lignin (Suppl. Fig. S2). The data histogram indicated complex distribution patterns. For bermudagrass-alfalfa bicultures, except for those variable pairs including fructan, all linearly fitted models indicated statistical significance (Suppl. Fig. S3) and the distribution pattern for each variable was generally following the Gaussian distribution curve. For tall fescue-white clover biculture, only CP-NDF model was deemed to be significant with adequate amount of variation explained. All nutritive value and production indices were normally distributed (Suppl. Fig. S4). For old world bluestem-sainfoin biculture, biomass production measured as monthly cutting correlated strongly with ADF and IVDMD (P < 0.05 and R2 ≈ 0.5), and ADF was strongly correlated with IVDMD; with all nutritive value indices normally distributed (Suppl. Fig. S5). Finally, for orchardgrass-alfalfa biculture, CP was negatively correlated with both ADF and NDF, ADF was positively correlated with NDF but negatively correlated with IVDMD, and both lignin and NDF were negatively correlated with IVDMD (P < 0.05 and R2 ≈ 0.5, Suppl. Fig. S6).
Forage mineral concentration
Concentrations of Ca, P, K, Mg, as well as Ca:P ratio were significantly affected by treatments when evaluated at P = 0.05 significance level. Interactions between harvesting month and treatment were significant on P, K, Mg concentration and Ca:P ratio, except for Ca concentration (P = 0.23). Year effect slightly interacted with treatment on P concentration and C:P ratio, but was caused mainly by magnitude differences across month. No year interactions (P > 0.05) were found in other treatments, thus, data are presented by averaged values. As indicated in Table 5, early-season annual systems had lower Ca concentration than both fescue-white clover and orchardgrass alfalfa bicultures. Likewise, annual systems indicated lower P concentration than orchardgrass alfalfa biculture in late season (September). For K, perennial systems consistently indicated greater concentration than the annual systems in early season, but this effect was reversed at the late harvesting season. The concentration of Mg in forage biomass was significantly different between treatments only in May and September, with the annual system having the lowest and orchardgrass-alfalfa the greatest Mg concentration. For the late harvesting season, orchardgrass alfalfa bicultures had greater Mg concentration than bermudagrass and alfalfa biculture. For Ca:P ratio, perennial systems consistently had greater values than the annual systems, except for the orchardgrass and alfalfa biculture from May to July, as well as bermudagrass and alfalfa biculture from May to August. The advantage of maintaining high Ca:P ratio from orchardgrass alfalfa bicultures was observed through the mid-to-late growing season.
Radiation use efficiency
RUE was significantly affected by treatment (P < 0.05) and month (P < 0.05). Additionally, a two-way interaction was detected between month and treatment on RUE (P < 0.05), thus, RUE were presented by month but averaged over years. System-wise RUE depended greatly on month (Table 1). Throughout the entire growing season, bermudagrass-alfalfa biculture consistently had greater RUE than any cool-season grass legume biculture except for the September month (Fig. 3). Likewise, old world bluestem-sainfoin biculture always had greater RUE than any other forage system except in May (not different from bermudagrass-alfalfa biculture) and September (only greater than orchardgrass-alfalfa biculture) months. Annual systems had very similar RUE patterns compared to both warm-season grass legume bicultures (similar to bermudagrass-alfalfa biculture in July and August; and similar to old world bluestem-sainfoin biculture in September).

Seasonal radiation use efficiency (RUE) of different forage systems at Lascassas, TN, reported as average values across 2018 and 2019 growing seasons. Means without a common letter differ at a P = 0.05 significance level. Standard errors (SE) are reported based on pooled estimates of population standard deviation used by the Tukey’s HSD procedure
Soil total carbon and gravimetric water content
Soil total carbon was not affected by forage system (P = 0.27), but by year (P = 0.04) and their two-way interactions (P = 0.03); a significant depth effect (0 < 0.001) was found, as well as its two-way interaction with forage system (P < 0.05) and three-way interaction with forage system and year (P < 0.01). Thus, soil total carbon data was later presented by year and sampling depth (Fig. 4). In 2018 fall season, annual system had greater soil total carbon at 0–5 cm than the 5–15 cm level, and the 0–5 cm depths under perennial forage systems. Additionally, both bermudagrass-alfalfa and orchardgrass-alfalfa bicultures had greater soil total carbon at the 0–5 cm than bermudagrass-alfalfa biculture at the 5–15 cm level. In 2019 spring season, orchardgrass-alfalfa biculture had greater soil total carbon than any other forage system at the 5–15 cm level. Additionally, the 0–5 cm soil total carbon under the annual systems was greater than the 5–15 cm level of any forage system except for tall fescue-white clover biculture. Total 0–5 cm soil carbon of tall fescue-white clover biculture was greater than that of bermudagrass-alfalfa, orchardgrass-alfalfa, and its own at the 5–15 cm level. All 0–5 cm soil total carbon concentrations across all forage systems were greater than those from the 5–15 cm depth. We also investigated the year effects on soil total carbon separated by different forage systems. As indicated, old world bluestem-sainfoin biculture had significantly increased soil total carbon from 2018 to 2019 (Fig. 5). No treatment-by-depth interaction (P = 0.18) was found on soil GWC, but data were presented by depth to better indicate moisture change at different depths. No significant treatment-year interactions were detected from soil GWC, thus, cross-year average data were presented (Fig. 6). Consistently greater soil GWC was observed from the annual systems, tall fescue and white clover, or old world bluestem-sainfoin biculture than bermudagrass-alfalfa or orchardgrass-alfalfa biculture.

Soil total carbon as affected by different forage systems at Lascassas, TN, reported as average values across 2018 and 2019 growing seasons. Means without a common letter differ at a P = 0.05 significance level. Standard errors (SE) are reported based on pooled estimates of population standard deviation used by the Tukey’s HSD procedure

Soil total carbon as affected by years within different forage systems at Lascassas, TN, reported as average values across 2018 and 2019 growing seasons. Means without a common letter differ at a P = 0.05 significance level. Standard errors (SE) are reported based on pooled estimates of population standard deviation used by the Tukey’s HSD procedure

Average soil gravimetric water content (GWC) measured at 0–50 cm and 50–100 cm as affected by different forage systems at Lascassas, TN, Means without a common letter differ at a P = 0.05 significance level. Error bars indicate the standard deviation of the mean. The top and the bottom edge of each box indicate the 75th and 25th percentile of the data, respectively. The center bar indicates the mean median, and the height indicates the interquartile range
Influence of different production and environmental variables on yield
The overall structural model yielded acceptable performance value (Χ2 = 39.1, P = 0.13, AIC = 81.3, SRMSR = 0.04); which explained 25 and 40% of the variation in yield and grass-to-legume ratio, respectively (Fig. 7). Moreover, the direct path coefficients of RUE on yield and grass-to-legume ratio on RUE were both significant with a value of 0.8 and 13.1, respectively. A significant covariance was also detected between soil GWC and grass-to-legume ratio.

Proposed path analysis network indicating the hypothesized inter-relationships between different production (Radiation Use Efficiency, RUE; Grass to Legume ratio, G2L) and average soil Gravimetric Water Content (GWC, 0–100 cm) on yield. Red arrows indicate significant standardized path coefficients (values on the arrow, P < 0.05). Black arrows indicate insignificant standardized path coefficients (values on the arrow, P > 0.05). Blue double arrows indicate significant correlation (covariance values on the arrow, P < 0.05). Number of stars indicates level of significance of each path model ((*P < 0.05, **P < 0.01, ***P < 0.001)
Discussion
Annual systems provided greater yield and old world bluestem-based biculture indicated great adaptability
Higher forage yield from annual systems (Fig. 1) was anticipated and can be partially explained by their more rapid establishment and fast growth compared with perennial systems. Perennial forages allocate significant portion of energy in the first two or three years developing root systems rather than above-ground biomass production (Inwood et al., 2015). Additionally, the mild temperature and ample precipitation in the winter season and hot summer in the local environment greatly favored the production of each annual rotation phase, including C3 species combination (e.g., winter wheat-Austrian winter pea biculture) and C4 dominated (e.g., sorghum-sudangrass hybrid) systems, respectively (Fribourg 1995; MacAdam and Nelson 2018). Our average forage yield from annual systems (12.7 Mg ha−1) is very similar to the yield (12 Mg ha−1) of the annual system of wheat/crimson clover (Trifolium incarnatum L.) and sorghum-sudangrass hybrid reported from a study conducted at Eastern Tennessee (Inwood et al. 2015). We used the same annual species selection/cultivation methods as Inwood et al. (2015) but without poultry litter application. Therefore, this yield level is impressive considering no external nutrients were added in our annual systems throughout the entire duration of the experiment. The inclusion of cowpea, a warm-season subtropic/tropic legume with enriched protein and biomass production potential (Saidi et al. 2010; Nave et al. 2020), had greatly improved summer forage biomass production and overall nutritive value of the biculture. In general, productivity of annual systems out-perform perennial systems, as indicated in our results (except for old world bluestem-sainfoin biculture). However, it is also worth mentioning that perennial crops typically take more than two or three years to reach their peak production (Kering et al. 2012), thus, the results from this two-year study might underestimate the yield potential of some of these selected perennial systems. To our knowledge, information relating to the production of old world bluestem in the tropical/subtropical environment is extremely limited, with published data focusing on organic production even non-existent. As a primary group of introduced C4 perennial grass species in the semiarid region, old world bluestems had indicated excellent drought tolerant capacity and economic return in water-limited environment (Philipp et al. 2005, 2007; Cui et al. 2013, 2014). As indicated in this study, it also performed very well in the humid subtropical environment while managed organically. It had comparable yield compared with annual forage biculture rotation, and had significantly greater yield than common grass-legume combinations (e.g., tall fescue-white clover biculture; Fig. 1). Sainfoin biomass remains low throughout the entire growing season (Table 2). This could be caused by that fact that this non-bloating (Jones and Lyttleton 1971), slow-to-establish (Cui et al. 2014) forage is better adaptable to cooler spring and high soil pH conditions (Bolger and Marches 1990). Thus, we speculate that old world bluestem in conjunction with other legumes, such as alfalfa or red clover (Trifolium pretense L.), might serve as better alternative in warm-season perennial systems compared to old world bluestem-sainfoin biculture or traditional bermudagrass-based monoculture or biculture systems. Finally, annual systems indicated poor economic benefit and warm-season perennial systems yielded great economic profit calculated based on production inputs and output values (details in supplement materials, Table S2).
Different grass-legume bicultures had different botanical composition dynamics throughout the growing season
Due to the lack of inorganic chemical inputs (e.g., herbicides), organic cropping systems are well known for weeds evasion and pest infestation (Francis and Van Wart 2009; Liebman and Davis 2009). We investigated botanical composition of unsown species (weeds) in this study (Table 2). As indicated, annual systems typically had less intensity of weeds throughout the entire growing season (average weeds composition ≈ 7.5%), which was similar to tall fescue-white clover biculture (≈ 9.7%), but much less than other perennial biculture systems (bermudagrass-alfalfa biculture, 25.5%; old world bluestem-sainfoin biculture, 17.0%; and orchardgrass-alfalfa biculture, 31.8%). This was largely contributed by the greater forage biomass yield and moderate tillage implemented before each planting. Severe weed encroachment was observed in both orchardgrass-alfalfa and bermudagrass-alfalfa bicultures. The main weed species included curly dock (Rumex crispus L.), creeping buttercup (Ranunculus repens L.), common plantain (Plantago major L.) and henbit (Lamium amplexicaule L.). Orchardgrass is a productive and highly nutritious cool-season grass widely distributed in North America and Europe (Xue et al. 2020). However, persistence of orchardgrass is often challenged by selective grazing, heat stress, soil fertility, insects, diseases, and harvesting frequency (Jones and Tracy 2015). In this study, we suspect that the limited establishment performance and severe early-season curly dock outbreak in those plots greatly compromised the percentage of orchardgrass. The high percentage of weed in bermudagrass were anticipated, as late-spring seed planting of warm-season grasses in the temperate zones is often interfered by severe spring weed outbreak (Inwood et al. 2015), thus asexual propagation of bermudagrasses such as sprigging is usually preferred due to its rapid establishment and ease of operation (de Barreda et al. 2013). This issue could be exacerbated by the fact that, certified organic/untreated sprigs for bermudagrass were extremely difficult to locate and even unavailable in many regions. Both tall fescue-white clover and old world bluestem-sainfoin bicultures indicated satisfactory capacity in controlling weed encroachment (< 20%). Again, information relating to these forage systems based on organic management practices is extremely limited, thus, there are no existing data to compare with. The excellent performance of tall fescue and white clover could be attributed to the fact that both species are well-adapted to this region and the endophytic toxins associated with tall fescue (Sleper and Buckner 1995) greatly enhanced its persistency as noted in many previous studies (Chamblee and Lovvorn 1953; Sheaffer 1989).
For grass-legume mixtures, botanical composition is well known for impacting forage nutritive value, because of the significant differences in chemical composition between grass and legume leaf tissues (Collins and Newman 2017). However, the interactions and behaviors on a system level were rarely focused, particularly under organic practices. Legumes are well-known for providing N credit through symbiotic N2 fixation (Westhoff 2009), but could also cause great domination (Muthukumar et al. 2017) in mixtures causing bloat hazard while improperly managed (Jones and Lyttleton 1971). All forage systems had shown relatively high grass-to-legume ratios (> 5:1) throughout the duration of this experiment. Again, the limited sainfoin production had resulted in extremely high grass-to-legume ratios. It was expected that legume domination will increase over time, particularly without any inorganic N fertilizer input. Thus, two-year duration might not be long enough for detecting this legume domination, long-term organic forage production should be an important future research direction.
Radiation use efficiency was largely influenced by dominant forage species
Radiation use efficiency is an important ecophysiological index for crop production in the temperate regions, where ample precipitation and frequent cloudy conditions greatly limit radiation availability. Particularly, RUE of organic grass-legume biculture systems, has been largely under-investigated. Generally speaking, C3 species, such as cool-season grasses and legumes are well known for having poor RUE compared with C4 plants (Giunta et al. 2009; Sandaña et al. 2012). Although lack of year-treatment interactions, our study indicated that RUE of various forage biculture systems had a strong seasonal variability (Fig. 3), which was similar to what was reported by Ojeda et al. (2018). Additionally, as expected, C4-dominated systems generally had greater RUE than cool-season grass-legume bicultures during most of the time throughout the growing season. We observed that old world bluestem-sainfoin constantly had the greatest RUE, indicating its great adaptability and performance potential for dry summer seasons. The RUE of tall-fescue-based systems were low (ranging from 1.8 to 2.3 g DM MJ−1), but was generally in agreement with the results reported by Ojeda et al. (2018).
Forage nutritive value exhibited complex responses towards maturity and botanical composition
Forage nutritive value reflects the species composition (grasses/legumes) and also the growth and maturity level of the combined swards. Often times, the interaction among different species and the complex growth patterns of mixed swards affected by environmental conditions make it difficult to isolate the contributory effects on nutritive value, particularly in biculture systems (Barker et al. 2010; Cui et al. 2013, 2014). In this organic forage study, cool-season perennial systems constantly provided superior CP concentration than warm-season systems (old world bluestem and bermudagrass-based treatments). Again, these CP difference was primarily caused by the photosynthetic pathway and cellular structural composition differences between C3 and C4 species (Collins and Newman 2017).It was expected that inclusion of legumes could greatly help maintaining CP level in C4-based forage systems according to Cui et al. (2013), however, this N credit contribution seemed insufficient for offsetting the rapid senescence property and more pronounced vascular tissues of C4 grasses compared with C3 (Belesky et al. 1991; Nave et al. 2014). Acid detergent fiber serves as an estimator for forage dry matter digestibility (Linn and Martin 1989). Thus, based on ADF content, the digestibility of annual and warm-season perennial forage systems could become critically low in late summer and late fall, respectively; potentially causing reduction in animal performance. Meanwhile, at comparable maturity stages, inclusion of legumes in grass-based forage systems could effectively reduce NDF and consequentially increase dry matter intake by grazing herbivores (Van Soest 1965; Cui et al. 2013). This agrees with what we found under organic systems, as lower grass-to-legume ratios (Table 2) typically manifested lower NDF concentration (Table 4). The IVDMD results reflected the combined effects of forage maturity and botanical composition. Generally speaking, orchardgrass and tall fescue should provide very similar IVDMD, and the general superior animal performance of orchardgrass vs. tall fescue was largely caused by the lack of endophytic toxins (Sleper and Buckner 1995), which could not be detected using the NIRS. Therefore, the greater IVDMD of tall fescue-white clover biculture in May and June should be largely attributed to differences in percentages of legumes. Non-structural carbohydrates, particularly fructan, predominates in cool-season grasses, and are well-known for inducing equine laminitis (Longland and Byrd 2006). When managed organically, tall-fescue-based systems had greater fructan concentration than others, thus, equine producers should proceed with caution while pasture-grazing tall fescue dominated lands. Unlike CP, ADF, or NDF; legumes generally have much greater concentration of lignin than grasses regardless of maturity stage (Collins and Newman 2017). Thus, botanical composition induced much greater interactive effects with cutting month over lignin concentration as observed in this study (Table 4). Across all treatments, the observed ranges of CP, ADF, NDF, and IVDMD were comparable to those reported from several polyculture forage systems (Burns and Fisher 2008; Cui et al. 2013; Tamu et al. 2014; Gelley et al. 2016). Additionally, it was well-understood and generally expected that nutritive values differ greatly among monthly cuttings within each forage system; thus, no comparisons were conducted among cutting months, and focuses were mainly placed on treatment effects. Key mineral element concentration in all five forage systems indicated no deficiency concerns for grazing steers (National Research Council 1996). In general, orchardgrass and alfalfa biculture consistently had similar or greater mineral concentration and Ca:P ratio than other systems. Additionally, grass-legume perennial systems generally outperformed annual systems, this could be largely attributed to the deeper root profiles of perennial legumes enabling greater nutrient uptake at a deeper soil level. All Ca:P ratios are larger than the recommended 1:1 level, indicating little concern on Ca deficiency for grazing cattle.
Finally, weed encroachment is one of the most important limiting factors impacting organic crop production (Francis and Van Wart 2009; Liebman and Davis 2009). Therefore, this study investigated weed composition to provide better information for site-specific organic management purposes. However, due to the lack of estimation equation from NIRS, all weed components were removed from nutritive value analysis. As indicated in Table 2, annual systems provided excellent weed suppression capacity, and cool-season dominated forage systems are prone to warm-season weed outbreaks (e.g. crabgrass, nutsedge, and nimblewill found in fescue or orchardgrass-legume mixtures) and vice versa (e.g. curly dock and henbit found in bermudagrass or old world bluestem-legume mixtures). Due to the acceptable palatability and nutritive value of both curly dock and crabgrass, we argue that the most challenging weeds for organic forage production in the transition zone should include henbit (Lamium amplexicaule L.), nimblewill (Muhlenbergia schreberi L.), and yellow nutsedge (Cyperus esculentus L.).
Correlation analysis unveiled strong relationship between different biomass and nutritive value indices
A less stringent criterion (P < 0.05 and R2 ≈ 0.5) was used in this study to evaluate the correlation between two production/nutritive indices. This is mainly because of the limited sample sizes and compositional complexity of the biculture samples for NIRS analysis. As expected, many classic correlations were identified across many forage systems, such as those pairs between ADF and NDF, ADF and IVDMD, as well as NDF and IVDMD. The lack of biomass-nutritive index correlations of annual systems could be mainly caused by the complexity of species composition, harvest season, and rotation patterns. This was also indicated in the histogram plotting. However, the short-growing season and more uniformed management regime of annual systems led to much more homogeneous sample batch indicated by the large number of significant inter-nutritive-index correlations with large R2 values (Suppl. Fig. S2). For C4 grass-based systems, bermudagrass-alfalfa biculture had strong correlations between monthly cuttings and CP. This finding agreed well with what was reported by Gelley et al (2016). For old world bluestem-sainfoin biculture, similarly, monthly biomass production was significantly correlated CP but with a lower R2 value. Both warm-season perennial systems indicated very strong correlation between monthly biomass production and ADF, NDF, IVDMD; hinting that monthly biomass production could serve as a reliable predictor for CP, ADF, NDF, and IVDMD within certain warm-season perennial systems. For C3 grass-based systems, similar biomass-nutritive index relationships were harder to find, which agreed with the results reported by Nave et al. (2013), indicating a much more complex interplay between maturity/production and nutritive responses of C3 grasses. The significant regression models successfully captured the correlations between two variables, which can be used for developing decision-making tools that producers can rely on to estimate hard-to-measure nutritive indices (e.g., CP, ADF, NDF, IVDMD, etc.) using biomass production information.
Soil total organic matter highlights the effect of the tillage during experiment establishment
The essence of organic production is to improve soil health and function on an ecosystem basis. Numerous studies have reported increased soil fertility/quality (Spargo et al. 2011; Cavigelli et al. 2013) and alteration of carbon and N cycling (Wander et al. 1994, 2007) within certified or transitional row-crop systems. However, information on certified forage systems remains limited. Evidently, the initial site preparation and tillage had induced significant amount of carbon loss across different depth as indicated in the overall soil total carbon reduction compared to the baseline data (Fig. 4). Consistent soil total carbon differences were observed between two sampling depths but little treatment effects were detected. Our soil total carbon level was generally lower than the results reported in a transitional forage cropping system study reported by Inwood et al. (2015) in Eastern Tennessee and a long-term perennial forage cropping system review by Franzluebbers et al. (2005) in the Prairie Parkland of Texas. The reasons are two folds: (1) Inwood et al. (2015) used poultry litter as an organic nutrient source in the fall season, which could significantly increase total C by more than 15% according to Franzluebbers et al. (2005) and Hernandez-Ramirez et al. (2009); (2) the synthesis study conducted by Franzluebbers (2005) focused mainly on those long-term (> 5 years) forage studies. It was expected that large amount of carbon contributed by perennial forages was in the format of fresh or partially decomposed root residues, which were generally removed from the soil laboratory analysis procedure. Finally, the frequent hay removal could significantly lengthen the stabilization phase of the passive soil organic matter pool (Rasmussen and Collins 1991), leading to a more conservative estimation of the soil total carbon contribution. The rapid rate of increase of soil total carbon detected under the old world bluestem-sainfoin biculture (Fig. 5) was unexpected. Given the performance on total yield (Fig. 1), monthly biomass production (Fig. 2), as well as the limited soil total carbon responses of annual systems; it is reasonable to attribute the over-time soil total carbon enhancement under the old world bluestem-sainfoin biculture to both increased above-ground biomass production and below-ground root biomass increase. For future study, longer-term implementation of similar forage systems and adoption of more sensitive soil carbon fractionation methods, such as particulate organic matter carbon and/or permanganate-oxidizable carbon, are warranted.
Soil gravimetric water content agreed well with botanical composition pattern
There is a broad agreement on the effects of botanical composition on soil water dynamics (Alamdarloo et al. 2018; Xu et al. 2018; Wu et al. 2018). We quantified soil GWC in addition to soil total carbon to investigate if soil moisture dynamics could be impacted under organic forage systems or not. Generally speaking, the deep tap-root systems of legumes greatly favor the utilization of deep-profile soil moisture, but could also reduce yield and hasten senescence during prolonged water deficit conditions (Alamdarloo et al. 2018). Thus, we anticipated the greater legume percentages in the biculture systems could cause greater uptake of the soil moisture particularly at a deeper soil level (> 15 cm). As indicated in our results, the lower grass-to-legume ratio in the two alfalfa-included bicultures (Table 2) had resulted in lower soil GWC levels across both sampling depths. However, we think this level of moisture difference could contribute little to productivity differences, as even the lowest GWC at the deeper soil profile was still above 10%. Additionally, ample precipitation in the temperate zones could recharge soil moisture rapidly due to the higher infiltration capacity of grasslands, particularly those legume incorporated grasslands, which were indicated to have larger-diameter root channels and enhanced preferential flow (Wu et al. 2017).
Radiation use efficiency indicated significant positive effects on yield and grass-to-legume ratio
Investigating the effects of different ecophysiological and environmental factors on yield using systematic modeling could provide important insights to better understanding the behavior of key production drivers. In this study, we found that RUE is indeed a key factor determining forage yield in the local environment. Again, unlike other semiarid environments, this indicates forage systems in the temperate regions are largely radiation limited. Additionally, grass-to-legume ratio greatly influences the overall RUE as captured by the SEM, because warm-season grasses (C4) could significantly enhance RUE compared to legumes (C3). The significant correlation between grass-to-legume ratio and soil GWC was anticipated as explained in “Soil gravimetric water content agreed well with botanical composition pattern” section, and was also captured by the path analysis model. We suspected that grass and legumes in biculture systems could behave differently towards solar radiation utilization than grass or legume monocultures alone. However, in this study, we did not investigate different components separately rather than quantification of biomass productivity, which needs to be addressed in future studies.
Conclusion
In conclusion, this study indicated that annual forage systems can provide greater biomass production/yield, however, their nutritive value and mineral concentration declined rapidly and could incur much greater production input cost with much smaller profit margin than perennial systems. Spring planting warm-season perennial forages under organic farming could be challenged by weed outbreak, and external N inputs seemed to be necessary for maintaining their nutritive value particularly during the mid-to-late growing seasons. Particularly, old world bluestems indicated great adaptation, radiation use efficiency, and soil carbon enhancement capacity in the local environment. The results from this study also indicated that monthly biomass production, an easy-to-measure index, could be used for accurately estimating forage nutritive value for grass-legume bicultures; thus, informing producers the optimal time window for hay harvesting or grazing. It is also worth mentioning that sampling procedures used in this study were based on hay sampling methods instead of grazing samples. Therefore, the nutritive results might represent an overestimation of herbage biomass production potential but an underestimation of forage nutritive value available to the grazing animals. Finally, long-term system study and better soil carbon fractionation methods are needed in the future to better evaluate the ecosystem service and long-term behavior of each forage system.
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- ADF:
-
Acid detergent fiber
- NDF:
-
Neutral detergent fiber
- CP:
-
Crude protein
- IVDMD:
-
In-vitro dry matter digestibility
- NIRS:
-
Near-infrared spectroscopy
- RUE:
-
Radiation use efficiency
- GWC:
-
Gravimetric water content
References
Alamdarloo EH, Manesh MB, Khosravi H (2018) Probability assessment of vegetation vulnerability to drought based on remote sensing data. Environ Monit Assess 190:702. https://doi.org/10.1007/s10661-018-7089-1
Archer DW, Jaradat AA, Johnson JM-F, Weyers SL, Gesch RW, Forcella F, Kludze HK (2007) Crop productivity and economics during the transition to alternative cropping systems. Agron J 99:1538–1547. https://doi.org/10.2134/agronj2006.0364
Barker DJ, Ferraro FP, Nave RLG, Sulc RM, Lopes F, Albrecht KA (2010) Analysis of herbage mass and herbage accumulation rate using Gompertz equations. Agron J 102:849–857. https://doi.org/10.2134/agronj2009.0381
Belesky DP, Perry HD, Windham WR, Mathias EL, Fedders JM (1991) Productivity and quality of bermudagrass in a cool temperate environment. Agron J 83:810–813. https://doi.org/10.2134/agronj1991.00021962008300050007x
Birkhofer K, Bezemer TM, Bloem J, Bonkowski M, Christensen S, Dubois D, Ekelund F, Fließbach A, Gunst L, Hedlund K, Mäder P, Mikola J, Robin C, Setälä H, Tatin-Froux F, Van der Putten WH, Scheu S (2008) Long-term organic farming fosters below and aboveground biota: implications for soil quality, biological control and productivity. Soil Biol Biochem 40:2297–2308. https://doi.org/10.1016/j.soilbio.2008.05.007
Bolger TP, Matches AG (1990) Water-use efficiency and yield of sainfoin and alfalfa. Crop Sci 30:143–148. https://doi.org/10.2135/cropsci1990.0011183X003000010032x
Burns JC, Fisher DS (2008) ‘Coastal’ and ‘Tifton 44’ bermudagrass availability on animal and pasture productivity. Agron J 100:1280–1288. https://doi.org/10.2134/agronj2007.0381
Cavigelli MA, Teasdale JR, Conklin AE (2008) Long-term agronomic performance of organic and conventional field crops in the mid-atlantic region. Agron J 100:785–794. https://doi.org/10.2134/agronj2006.0373
Cavigelli MA, Mirsky SB, Teasdale JR, Spargo JT, Doran J (2013) Organic grain cropping systems to enhance ecosystem services. Renew Agric Food Syst 28:145–159. https://doi.org/10.1017/S1742170512000439
Chamblee DS, Lovvorn RL (1953) The effect of rate and method of seeding on the yield and botanical composition of alfalfa-orchardgrass and alfalfa-tall fescue. Agron J 45:192–196. https://doi.org/10.2134/agronj1953.00021962004500050004x
Coll L, Cerrudo A, Rizzalli R, Monzon JP, Andrade FH (2012) Capture and use of water and radiation in summer intercrops in the south-east Pampas of Argentina. Field Crops Res 134:105–113. https://doi.org/10.1016/j.fcr.2012.05.005
Collins M, Newman YC (2017) Forage quality. In: Collins M, Nelson CJ, Moore KJ, Barnes RF (eds) Forages, an introduction to grassland agriculture, vol 1, 7th edn. Iowa State Univ. Press, Ames, pp 35–50
Cui S, Allen VG, Brown CP, Wester DB (2013) Growth and nutritive value of three old world bluestems and three legumes in semiarid Texas High Plains. Crop Sci 53:329–340. https://doi.org/10.2135/cropsci2012.02.0107
Cui S, Zilverberg CJ, Allen VG, Brown CP, Moore-Kucera J, Wester DB, Mirik M, Chaudhuri S, Phillips N (2014) Carbon and nitrogen responses of three old world bluestems to nitrogen fertilization or inclusion of a legume. Field Crops Res 164:45–53. https://doi.org/10.1016/j.fcr.2014.05.011
Dawson JC, Huggins DR, Jones SS (2008) Characterizing nitrogen use efficiency in natural and agricultural ecosystems to improve the performance of cereal crops in low-input and organic agricultural systems. Field Crops Res 107:89–101. https://doi.org/10.1016/j.fcr.2008.01.001
de Barreda DG, Reed T, Yu J, McCullough P (2013) Spring establishment of four warm-season turfgrasses after fall indaziflam applications. Weed Technol 27(3):448–453. https://doi.org/10.1614/WT-D-12-00168.1
Delate K, Cambardella CA (2004) Agroecosystem performance during transition to certified organic grain production. Agron J 96:1288–1298. https://doi.org/10.2134/agronj2004.1288
Delbridge TA, Coulter JA, King RP, Sheaffer CC, Wyse DL (2011) Economic performance of long-term organic and conventional cropping systems in Minnesota. Agron J 103:1372–1382. https://doi.org/10.2134/agronj2011.0371
Francis C, Van Wart J (2009) History of organic farming and certification. In: Francis C (ed) Organic farming: the ecological system, Agronomy Monograph, vol 54, ASA, CSSA, SSSA, Madison, WI, pp 3–17. https://doi.org/10.2134/agronmonogr54.c1
Franzluebbers AJ (2005) Soil organic carbon sequestration and agricultural greenhouse gas emissions in the southeastern USA. Soil Tillage Res 83:120–147. https://doi.org/10.1016/j.still.2005.02.012
Franzluebbers AJ (2010) Soil organic carbon in managed pastures of the southeastern
Fribourg HA (1995) Summer annual grasses. In: Barnes RF et al (ed), Forages, an introduction to grassland agriculture, vol 1. 5th edn. Iowa State Univ. Press, Ames, pp 463–472
Gelley C, Nave RLG, Bates G (2016) Forage nutritive value and herbage mass relationship of four warm-season grasses. Agron J 108:1603–1613. https://doi.org/10.2134/agronj2016.01.0018
Giunta F, Pruneddu G, Motzo R (2009) Radiation interception and biomass and nitrogen accumulation in different cereal and grain legumes species. Field Crops Res 110:76–84. https://doi.org/10.1016/j.fcr.2008.07.003
Goering HK, Van Soest PJ (1970) Forage fiber analysis (apparatus, reagents, procedures and some applications). ARS USDA Handbook No. 379. U.S. Gov. Print. Office, Washington, DC
Hernandez-Ramirez G, Brouder SM, Smith DR, Van Scoyoc GE (2009) Carbon and nitrogen dynamics in an eastern Corn Belt soil: nitrogen source and rotation. Soil Sci Soc Am J 73:128–137. https://doi.org/10.2136/sssaj2007.0381
Hinsinger P (2001) Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: a review. Plant Soil 237:173–195
Inwood SEE, Bates G, Butler D (2015) Forage performance and soil quality in forage systems under organic management in the Southern United States. Agron J 107:1641–1652. https://doi.org/10.2134/agronj14.0472
Jones WT, Lyttleton JW (1971) Bloat in cattle. A survey of legume forages that do and do not produce bloat. N Z J Agric Res 14:101–107. https://doi.org/10.1080/00288233.1971.10421306
Jones G, Tracy B (2015) Orchardgrass die-off: How harvest management and heat stress may be reducing the persistence of orchardgrass hay stands. Crops Soils 48:4–8. https://doi.org/10.2134/cs2015-48-3-1
Kering MK, Butler TJ, Biermacher JT, Guretzky JA (2012) Biomass yield and nutrient removal rates of perennial grasses under nitrogen fertilization. Bioenergy Res 5(1):61–70. https://doi.org/10.1007/s12155-011-9167-x
Li Y, Allen VG, Chen J, Hou F, Brown CP, Green P (2013) Allelopathic influence of a wheat or rye cover crop on growth and yield of no-till cotton. Agron J 105:1581–1587. https://doi.org/10.2134/agronj2013.0065
Li Z, Chen C, Nevins A, Pirtle T, Cui S (2021) Assessing and modeling ecosystem carbon exchange and water vapor flux of a pasture ecosystem in the temperate climate-transition zone. Agronomy 11:2071. https://doi.org/10.3390/agronomy11102071
Liebman M, Davis AS (2009) Managing weeds in organic farming systems: an ecological approach. In: Fracis C (ed) Organic farming: the ecological system. Agronomy Monograph, vol 54, ASA. Madison, WI, pp 173–196
Linn JG, Martin NP (1989) Forage quality tests and interpretation. Minnesota Ext. Services, Univ. Minnesota, AG-FO-2637. Univ. of Minnesota, St. Paul, MN
Longland AC, Byrd BM (2006) Pasture nonstructural carbohydrates and equine laminitis. J Nutr 136(7):2099–2102. https://doi.org/10.1093/jn/136.7.2099S
MacAdam JW, Nelson CJ (2018) Physiology of forage plants. In: Collins M, Nelson CJ, Moore KJ, Barnes RF (eds) Forages, an introduction to grassland agriculture, vol 1, 7th edn, Blackwell Publishing, Ames, IA, pp 51–70
Mahoney PR, Olson KD, Porter PM, Huggins DR, Perillo CA, Kent Crookston R (2004) Profitability of organic cropping systems in southwestern Minnesota. Renew Agric Food Syst 19(01):35–46. https://doi.org/10.1079/RAFS200361
Mitchell LE, Nelson CJ (2003) Forages and grassland in a changing world. In: Barnes RF, Nelson CJ, Collins M, Moore KJ (eds) Forages, an introduction to grassland agriculture, vol 1, 6th edn, Blackwell Publishing, Ames, IA, pp 3–24
Mitchell RB, Nelson CJ (2017) Structure and morphology of legumes and other forbs. In: Collins M, Nelson CJ, Moore KJ, Barnes RF (eds) Forages, an introduction to grassland agriculture, vol 1, 7th edn, Iowa State Univ. Press, Ames, pp 35–50
Murray I, Cowe I (2004) Sample preparation. In: Roberts CA, Workman JJ, Reeves JB (eds) Near-infrared spectroscopy in agriculture, ASA, CSSA, and SSSA, Madison, WI, pp 75–112
National Research Council (1996) Nutrient requirements of beef cattle, 7th edn, Natl. Acad press, Washington, DC
Nave RLG, Sulc RM, Barker DJ, St-Pierre N (2014) Changes in forage nutritive value among vertical strata of a cool-season grass canopy. Crop Sci 54:2837–2845. https://doi.org/10.2135/cropsci2014.01.0018
Nave RLG, Quinby MP, Griffith AP, Corbin MD, Bates GE (2020) Forage mass, nutritive value and economic viability of cowpea overseeded in tall fescue and sorghum-sudangrass swards. Crop Forage Turfgrass Mgmt 6:e20003. https://doi.org/10.1002/cft2.20003
Oberholtzer L, Dimitri C, Schumacher G (2012) Linking farmers, healthy foods, and underserved consumers: exploring the impact of nutrition incentive programs on farmers and farmers’ markets. J Agric Food Syst Community Dev 2:63–77. https://doi.org/10.5304/jafscd.2012.024.002
Ojeda JJ, Caviglia OP, Agnusdei MG, Errecart PM (2018) Forage yield, water, and solar radiation-productivities of perennial pastures and annual crops sequences in the south-eastern Pampas of Argentina. Field Crops Res 221:19–31. https://doi.org/10.1016/j.fcr.2018.02.010
Philipp D, Allen VG, Mitchell RB, Brown CP, Wester DB (2005) Forage nutritive value and morphology of three old world bluestems under a range of irrigation levels. Crop Sci 45:2258–2268. https://doi.org/10.2135/cropsci2004.0669
Philipp D, Allen VG, Lascano RJ, Brown CP, Wester DB (2007) Production and water use efficiency of three old world bluestems. Crop Sci 47:787–794. https://doi.org/10.2135/cropsci06.05.0340
Quinby MC, Nave RLG, Bates GE, Mcintosh D (2020) Harvest interval effects on the persistence and productivity of alfalfa grown as a monoculture or in mixtures in the southeastern United States. Crop Forage Turfgrass Manage 6(1):e20018. https://doi.org/10.1002/cft2.20018
Rasmussen PE, Collins HP (1991) Long-term impacts of tillage, fertilizer, and crop residue on soil organic matter in temperate semiarid regions. Adv Agron 45:93–134. https://doi.org/10.1016/S0065-2113(08)60039-5
Rosseel Y (2012) lavaan: an R package for structural equation modeling. J Stat Softw 48(2):1–36
Saidi M, Itulya FM, Aguyoh J, Ngouajio M (2010) Leaf harvesting time and frequency affect vegetative and grain yield of cowpea. Agron J 102:827–833. https://doi.org/10.2134/agronj2009.0421
Sandaña P, Ramírez M, Pinochet D (2012) Radiation interception and radiation use efficiency of wheat and pea under different P availabilities. Field Crops Res 127:44–55. https://doi.org/10.1016/j.fcr.2011.11.005
Sanderson MA, Brink G, Ruth L, Stout R (2012) Grass-legume mixtures suppress weeds during establishment better than monocultures. Agron J104:36–42. https://doi.org/10.2134/agronj2011.0130
SAS Institute Inc (2018) SAS® 9.4 in-database products: user’s guide, 8th edn, SAS Institute Inc., Cary
Saxton AM (1998) A macro for converting mean separation output to letter groupings in Proc Mixed. In: Proceedings 23rd SAS users group international conference, Nashville, TN, 22–25 Mat. 1998. SAS Inst., Cary, NC, pp 1243–1246
Schoofs A, Entz MH (2000) Influence of annual forage on weed dynamics in a cropping system. Can J Plant Sci 80:187–198. https://doi.org/10.4141/P98-098
Sheaffer CC (1989) Effect of competition on legume persistence. In: Marten GC et al (ed) Persistence of forage legumes, ASA, CSSA, and SSSA, Madison, WI, pp 327–334
Sims GK, Ellsworth TR, Mulvaney RL (1995) Microscale determination of inorganic nitrogen in water and soil extracts. Commun Soil Sci Plant Anal 26:303–316. https://doi.org/10.1080/00103629509369298
Sleper DA, Buckner RC (1995) The fescues. In: Barnes RF et al. (ed) Forages, an introduction to grassland agriculture, vol 1, 5th edn, Iowa State Univ. Press, Ames, pp 345–356
Spargo JT, Cavigelli MA, Mirsky SB, Maul JE, Meisinger JJ (2011) Mineralizable soil nitrogen and labile soil organic matter in diverse long-term cropping systems. Nutr Cycling Agroecosyst 90:253–266. https://doi.org/10.1007/s10705-011-9426-4
Temu VW, Rude BJ, Baldwin BS (2014) Nutritive value response of native warm-season forage grasses to harvest intervals and durations in mixed stands. Plants 3:266–283. https://doi.org/10.3390/plants3020266
Tsubo M, Walker S (2002) A model of radiation interception and use by a maize-bean intercrop canopy. Agric for Meteorol 110:203–215. https://doi.org/10.1016/S0168-1923(01)00287-8
Tu C, Louws FJ, Creamer NG, Mueller JP, Brownie C, Fager K, Bell M, Hu S (2006) Responses of soil microbial biomass and N availability to transition strategies from conventional to organic farming systems. Agr Ecosyst Environ 113:206–215. https://doi.org/10.1016/j.agee.2005.09.013
United States of America. In: Abberton M et al (ed) Grassland carbon sequestration: management, policy and economics. Integrated Crop Management 11. FAO, Rome, Italy, pp 163–175
Van Soest PJ (1963) Use of detergents in the analysis of fibrous feeds. II. A rapid method for the determination of fiber and lignin. J Assoc off Agric Chem 46:829–835. https://doi.org/10.1093/jaoac/73.4.491
Wander MM, Traina SJ, Stinner BR, Peters SE (1994) Organic and conventional management effects on biologically-active soil organic-matter pools. Soil Sci Soc Am J 58:1130–1139. https://doi.org/10.2136/sssaj1994.03615995005800040018x
Wander MM, Yun W, Goldstein WA, Aref S, Khan SA (2007) Organic N and particulate organic matter fractions in organic and conventional farming systems with a history of manure application. Plant Soil 291:311–321. https://doi.org/10.1007/s11104-007-9198-4
Westhoff P (2009) The economics of biological nitrogen fixation in the global economy. In Emerich DW, Krishman HB (eds) Nitrogen fixation in crop production, Agronomy. Monographs 52, ASA, CSSA, SSSA, Madison, WI, pp 309–328
Willer H, Lernound J (eds) (2016) The world of Organic Agriculture. Statistics and Emerging Trends. Research Institute of Organic Agriculture (FiBL), Frick, and IFOAM—Organics International, Bonn
Wu GL, Liu Y, Yang Z, Cui Z, Deng L, Chang XF, Shi ZH (2017) Root channels to indicate the increase in soil matrix water infiltration capacity of arid reclaimed mine soils. J Hydrol (amst) 546:133–139. https://doi.org/10.1016/j.jhydrol.2016.12.047
Wu GL, Huang Z, Liu YF, Cui Z, Liu Y, Chang X, Tian FP, López-Vicente M, Shi ZH (2018) Soil water response of plant functional groups along an artificial legume grassland succession under semi-arid conditions. Agric For Meteorol 278:107670. https://doi.org/10.1016/j.agrformet.2019.107670
Xu HJ, Wang XP, Zhao CY, Yang XM (2018) Diverse responses of vegetation growth to meteorological drought across climate zones and land biomes in northern China from 1981 to 2014. Agricultural and Forest Meteorology, vol 262, pp 1–13. https://doi.org/10.1016/j.agrformet.2018.06.027
Xue Z, Liu N, Wang Y, Yang H, Wei Y, Moriel P, Palmer E, Zhang Y (2020) Combining Orchardgrass and Alfalfa: effects of forage ratios on in vitro rumen degradation and fermentation characteristics of silage compared with hay. Animals 10:59. https://doi.org/10.3390/ani10010059
Acknowledgements
We would like to acknowledge Mr. Matthew Wade (Farm Manager, Middle Tennessee State University Experiential Learning Farm Laboratory) and Mr. Danny Troup (Crop Unit Manager) for providing support for establishing and maintaining the research sites. This research was partially supported by the National Institute of Food and Agriculture (2017-51106-2703; 2018-70001-28761), Guizhou Provincial Science and Technology Projects (Qian Ke He Zhi Cheng [2022] Yi Ban106), the Special Natural Science Foundation of Guizhou University (Gui Da Te Gang He Zi [2020]08), Dushan plum science and technology courtyard of Guizhou and Guizhou Talent Base of Grassland Ecological Animal Husbandry (RCJD2018-13). The authors want to appreciate the anonymous reviewers for their diligent work and help in improving this manuscript.
Author information
Authors and Affiliations
Contributions
Conceptualization: ZL, SC. Data curation: SX. Formal analysis: ZL, SN. Funding acquisition: ZL, CC, SC. Investigation: RN, JK, GB, DM. Methodology: SJ, CC, SC. Project administration: CC, SC. Resources: SJ, SC. Supervision: NR, CC, SC. Validation: SC. Visualization: SX. Writing—original draft: ZL. Writing—review and editing: CC, SC.
Corresponding authors
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Li, Z., Xu, S., Rajan, N. et al. Productivity and nutritive value of no-input minimum tillage organic forage systems. Nutr Cycl Agroecosyst 124, 335–357 (2022). https://doi.org/10.1007/s10705-022-10235-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10705-022-10235-z