
Abstract
We considered the limnological literature for an overview of biomanipulation methods that were implemented to avoid or reduce cyanobacterial bloom development in ponds and lakes. For this purpose, we reviewed 48 publications representing 34 whole-lake and large-scale case studies of different biomanipulation approaches clearly mentioning the extent of a cyanobacteria bloom problem and the cyanobacteria taxa involved. This delivered complementary information to the suite of review papers already providing elaborated syntheses on biomanipulation and associated ecotechnological measures as a restoration tool for overall eutrophication reduction and control. We considered nature-based solutions such as fish removal and associated water drawdown, addition of piscivorous fish, filter-feeding planktivorous fish, Daphnia or bivalves, re-introduction of macrophytes and a combination of accompanying restoration methods. Reasons for success or failure to control cyanobacterial blooms of especially Anabaena, Pseudanabaena, Aphanizomenon, Aphanocapsa, Limnothrix, Microcystis, Oscillatoria or Spirulina spp. could be explained through bottlenecks encountered with fish removal, stocking densities, cascading effects, associated zooplankton grazing, diet shifts away from cyanobacteria, macrophyte recovery, nutrient or pH status. Threshold values to avoid failures are synthesized from experiments or monitoring studies and presented in a conceptual scheme about cyanobacteria reduction through (1) direct abatement of existing blooms and forcing/maximization of biotic key interactions (2) reducing risk of blooms and improving lake or pond multi-functionality and (3) avoiding blooms, balancing biotic communities and enhancing existing ecosystem services. More information will be required on temporal dynamics and abundances of cyanobacteria taxa in whole-lake pre- and post-biomanipulation conditions to better evaluate the applicability and effectiveness of such nature-based solutions.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Nature-based solutions are measures which are supported by nature and jointly benefit the environment, society and the economy. Because of these multiple benefits, they are more efficient and cost-effective than traditional methods. A principal goal that can be addressed by nature-based solutions is the restoration of degraded ecosystems (European Union 2015). As a nature-based solution to restore lake and pond ecosystems, enabling them to deliver vital ecosystem services, biomanipulation can be a relatively cost-effective, albeit mostly ephemeral substitute for large-scale nutrient reductions (Søndergaard et al. 2008; Jeppesen et al. 2012). Biomanipulation, described as the intervention in an ecosystem by manipulating key components of the ecological community, has been proposed as a means to restrain freshwater primary production boosted by eutrophication (Shapiro et al. 1975). In productive, shallow water bodies, the nature of alternative stable states may prevent a systematic improvement of water clarity in spite of efforts reducing eutrophication (Scheffer et al. 1993, 2001; Ibelings et al. 2007). Similarly, larger and deeper lakes may benefit from biomanipulation in order to shift the balance in favor of a stable state characterized by non-cyanobacterial phytoplankton dominance in the pelagic zone (Horppila et al. 1998; Peckham et al. 2006), although significant fish removal in very large lakes is less likely to be cost-efficient (Jeppesen et al. 2007). Problematic and recurrent cyanobacterial blooming justified managers to resort to biomanipulation, mainly by modifying the fish fauna. The induced interactions affecting cyanobacterial abundance can be either direct (by increasing filtration capacity of filter feeders) or indirect (through a trophic cascade, changes in nutrient dynamics or macrophyte-associated mechanisms).
We did not intend to repeat the overall effects of biomanipulation on reducing phytoplankton biomass through pelagic cascading effects because these have been comprehensively reviewed (e.g., Hansson et al. 1998; Drenner and Hambright 1999; Gulati and van Donk 2002; Mehner et al. 2002; Søndergaard et al. 2007; Gulati et al. 2008; Sierp et al. 2009; Jeppesen et al. 2012). This review will focus on selected literature, only concerning whole-lake studies or large-scale studies, clearly reporting on cyanobacteria reduction, their taxa and abatement as an essential target of biomanipulation efforts. We only highlight those measures involving manipulation of aquatic organisms as well as water drawdown, which is used to assist fish removal and therefore relates to biomanipulation. Acute chemical or physical treatment of blooms of cyanobacteria neither is considered, nor are methods involved in external or internal nutrient reduction.
Most studies about the effects of biomanipulation on water clarity do not mention the effects on cyanobacteria biomass or blooms but only refer to the effects on phytoplankton biomass or Chl a concentration. Moreover, reports of case studies involving biomanipulation vary in the use of different units to describe cyanobacterial abundance (cell density, biovolume, biomass or Chl a concentration). Therefore, the degree of change after biomanipulation is seldom comparable.
In total, nine case studies and meta-analyses describing effects on cyanobacterial prominence or parameters were reviewed in which restoration involved only fish removal, or in which effects of fish removal could be clearly separated from other management actions (Table 1). In order to assess the consequences of biomanipulation through fish removal, changes in the fish assemblage, establishment of macrophytes and nutrient or pH dynamics are treated separately. We considered five studies dealing with the effect of piscivore stocking on cyanobacteria (Table 1) and six studies mentioning the effects of stocking filter-feeding planktivorous fish on cyanobacteria (Table 1). Eight studies combined fish removal with piscivore stocking, whereas six studies combined multiple restoration techniques, providing a clear description of effects on cyanobacteria (Table 1). The majority of the 34 reviewed case studies dealt with water bodies that were (highly) eutrophic comprised an area between 0.002 and 110 km2 with a depth ranging from 0.5 to 48 m and included temperate as well as tropical water bodies. A brief description of the characteristics of the considered water bodies as well as their usage is given in Table 2.
We aim to review the effect on cyanobacteria reduction in three main biomanipulation methods (fish removal, piscivorous fish stocking and stocking with filter-feeding fish) as reported in representative case studies. Additionally, we aim to unravel the effects of those case studies that combined different restoration techniques. Our objective further was to discuss bottlenecks and threshold values related to the biomanipulation methods to control cyanobacteria, including the potential ecological impacts of stocking with alien species. We then conceptually emphasize the use of biomanipulation strategies as nature-based solutions.
Overview of biomanipulation methods and proposed working mechanisms
Fish removal
Removal of benthivorous and zooplanktivorous fish probably is the most popular management tool to manipulate the lacustrine food web. As primary production in aquatic systems is strongly nutrient-driven, biomanipulation often can be thought of as a short-lived surrogate for a more thorough remediation of excess internal and external nutrient supply (Søndergaard et al. 2008; Hobbs et al. 2012). When nutrient levels are not dealt with, a return of elevated phytoplankton turbidity or cyanobacterial blooms often is inevitable. The different processes involved in classical biomanipulation and useful for cyanobacterial reduction are described hereafter.
Catch effort
The minimal fish catch effort needed lies within the order of either a 75–80 % reduction within a few years (Meijer et al. 1999; Søndergaard et al. 2000) or a removal of >200 kg ha−1 over a three-year period (Olin et al. 2006; Søndergaard et al. 2008). A mathematical relation for temperate shallow lakes was described by Jeppesen and Sammalkorpi (2002) as: required annual catch (kg ha−1) = 6.9 TP0.52 (total phosphorus in µg P L−1). In deeper lakes, the necessary catch effort could be lower (Jeppesen and Sammalkorpi 2002).
Establishment of a clear-water phase
Fish removal can increase transparency in a number of ways, including bottom-up and top-down effects. In shallow lakes and ponds, decreased bioturbation by benthivorous fish reduces concentration of suspended particles and nutrient availability in the water column (Søndergaard et al. 2008) (Fig. 1). The reduced sediment resuspension by large bream, carp, tench or roach to a great extent can be responsible for creating a window of opportunity for submerged macrophyte colonization in spring, and tends to outlive effects of biomanipulation on phytoplankton turbidity (Beklioglu and Tan 2008; Søndergaard et al. 2008). However, turbid water bodies might benefit primarily from an increased grazing pressure of large Cladocera, released from predation by zooplanktivorous fish (Fig. 1).

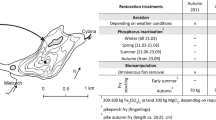
Biomanipulation tools to control cyanobacteria. Biomanipulation-mediated biotic interactions preventing cyanobacterial growth are given in black. Undesired side effects of biomanipulation are shown in gray (see box for explanation). Conventional arrows represent facilitating effects, while–—| interactions indicate negative influences. Dotted arrow: nutrient recycling by fish; YOY fish: young-of-the-year fish
Filter-feeding Cladocera, especially Daphnia spp., are able to create periods of clear water, thereby stimulating macrophyte growth. Nevertheless, filtration of both Daphnia and relatively small-bodied Bosmina could potentially alter the composition of phytoplankton in favor of large, difficult-to-process filamentous cyanobacteria or Microcystis colonies (Hanson and Butler 1994; Dawidowicz et al. 2002), thus actually increasing the risk of toxic bloom formation after biomanipulation. Although taxon identity as well as respective densities influences the interaction between daphnids and cyanobacteria (Asselman et al. 2014; Jiang et al. 2014), large-sized Daphnia are able to reduce even theoretically inedible cyanobacteria and tolerate high toxin concentrations under certain conditions (Sarnelle 2007; Chislock et al. 2013; Ekvall et al. 2014). Strong decrease in filamentous cyanobacteria following fish removal has also been attributed to peaks of Bosmina (ter Heerdt and Hootsmans 2007). Furthermore, isotopic analyses in the context of biomanipulation have suggested a generalistic feeding behavior of zooplankton toward available food sources, with consumers incorporating more carbon of prokaryotic origin when cyanobacteria dominate (Bontes et al. 2006).
High cyanobacterial densities (>50,000 cells mL−1) prior to biomanipulation were related to a limited chance of achieving lake bottom view in shallow lakes of the Netherlands, although this pattern could be explained by an insufficient fish reduction (Meijer et al. 1999). Nevertheless, an initial strong dominance of cyanobacteria appears to be no obstacle for successful restoration (Olin et al. 2006; Søndergaard et al. 2008).
Apart from interacting in the trophic food chain, fish removal might weaken cyanobacterial dominance when growth of colonies or filaments is stimulated by gut passage. Following excretion, growth of ingested Microcystis spp. and Oscillatoria agardhii can be facilitated, at least partly as a result of nutrient uptake in the alimentary canal (Kolmakov and Gladyshev 2003; Lewin et al. 2003; Zeng et al. 2014). In a Siberian reservoir, removal of crucian carp (Carassius auratus) has been shown to control blooms dominated by Microcystis, likely aided by decreased gut passage since no daphnid grazing was observed (Gladyshev et al. 2003; Kolmakov and Gladyshev 2003; Prokopkin et al. 2006).
Submerged macrophyte colonization
Once a clear-water phase has been established, submerged macrophytes may gradually colonize shallow regions of the water body (Fig. 1). In shallow lakes and ponds, aquatic vegetation has the potential to occupy the whole surface area, a feature with far-reaching implications for the suppression of planktonic blooms. A suite of macrophyte-mediated interactions enhancing the stability of the clear-water state have been identified, among others nutrient competition with phytoplankton, provision of shelter for horizontally migrating zooplankton and allelopathy (Burks et al. 2002; van Donk and van de Bund 2002). In the end, it becomes unrealistic to unravel mechanisms of biomanipulation once large beds of submerged macrophytes have developed.
In temperate regions, those macrophytes experiencing winter die-off rely on a yearly return of a sufficiently long clear-water phase in spring in order to establish stands large enough to inhibit phytoplankton growth in summer. Unfortunately, following biomanipulation, the likelihood of an ideal light climate for macrophyte recruitment early in the growing season diminishes year after year (Søndergaard et al. 2008; De Backer et al. 2012). A long-term stability after biomanipulation often becomes compromised by an unfavorable nutrient state or the impossibility to achieve a balanced fish community (Meijer et al. 1999). Regime shifts in shallow lakes can be distinct and do not necessarily occur consecutively during multiple years (Zimmer et al. 2003).
Addition of piscivores
Addition of piscivorous fish as a biomanipulation tool has certain advantages compared to fish removal (Berg et al. 1997), but usually is less successful than traditional fish removal (reviewed in Drenner and Hambright 1999; Søndergaard et al. 2007; Jeppesen et al. 2012) in reducing phytoplankton biomass.
Addition of piscivores is frequently used to enhance water transparency through a trophic cascade (Drenner and Hambright 1999; Seda et al. 2000; Scharf 2007; Ha et al. 2013) (Fig. 1) allowing large zooplankton (mainly Daphnia species) to increase grazing pressure on phytoplankton (Carpenter et al. 1985) (Fig. 2).

Overview of different biomanipulation strategies for cyanobacterial control at different levels of the cascading food web and the benthic compartment according to their application in different water bodies and different categories that maximize either selected key services (productivity, clear-water conditions) or the number of ecosystem services (eutrophication control, functional biodiversity). A selection of various thresholds or indicative values is presented after (*1) Ke et al. (2008), (*2) De Backer et al. (2014), (*3) Peretyatko et al. (2010), (*4) De Backer et al. (2012), (*5) Søndergaard et al. (2000), (*6) Meijer et al. (1999), (*7) Ha et al. (2013), (*8) Søndergaard et al. (2007), (*9) Hosper and Jagtman (1990), (*10) Skov and Nilsson (2007), (*11) Jeppesen et al. (1990), (*12) Zimmer et al. (2009)
Several studies in the European temperate zone mainly used pike (Esox lucius) and pikeperch (Sander lucioperca) to control planktivorous fish for biomanipulation (Meijer and Hosper 1997; Drenner and Hambright 1999; Seda et al. 2000; Skov et al. 2002; Jacobsen et al. 2004; Søndergaard et al. 2007; De Backer 2011). Repeated stocking of piscivorous eels (Anguilla anguilla) and trout (Salmo trutta lacustris, Onchorhynchus mykiss) was frequently used in reservoirs in Germany occasionally combined with catch restrictions for piscivores (Dörner and Benndorf 2003; Kasprzak et al. 2007; Scharf 2007). Other piscivorous species used to generate top-down control included European catfish (Silurus glanis) and asp (Aspius aspius) in Czech and German reservoirs (Drenner and Hambright 1999; Seda et al. 2000), walleye (Sander vitreus) and largemouth bass (Micropterus salmoides) in the USA (Potthoff et al. 2008; Deboom and Wahl 2014) and perch (Perca fluviatilis) in the Netherlands (Drenner and Hambright 1999; Gulati and van Donk 2002). In Australia, Australian bass (Macquaria novemaculeata) was used (Pinto et al. 2005), while Sierp et al. 2009 discussed the potential of using Murray cod (Maccullochella peelii peelii) as a predator.
Addition of filter-feeding planktivorous fish
This method aims at direct cyanobacteria abatement through consumption by omni-planktivorous filter-feeding fish species (Figs. 1, 2). When conditions do not allow neither effective planktivorous fish removal nor external and internal phosphorus reduction, or a submerged macrophyte re-establishment, then the cascading concept of biotic manipulations through increased zooplankton grazing rate as a tool for controlling nuisance phytoplankton becomes invalid but ultimately can be replaced by stocking phytoplanktivorous fish (Zhang et al. 2008). This restoration method implements productivity through enforcing key biotic interactions and envisages to a much lesser extent the restoration of biodiversity and ecosystem services (Fig. 2). Piscivorous fish, submerged macrophyte beds and large zooplankton usually are absent or non-effective biotic components in highly productive (sub)tropical lakes. This food-web manipulation method aims to improve water quality in highly productive (sub)tropical lakes, where grazing pressure from large zooplankton is decoupled from the pelagic food web because cyanobacterial blooms flourish to form scum (Xie and Liu 2001).
In China, large-bodied planktivorous cyprinids, silver carp (Hypophthalmichthys molitrix) and bighead carp (Aristichthys nobilis) are intensively cultured for their use in controlling especially Microcystis blooms (Tang et al. 2002; Ke et al. 2007; 2008; Xiao et al. 2010). In general, bighead carp is considered an opportunist, feeding more on zooplankton than on phytoplankton, whereas silver carp mainly feeds on phytoplankton (Cremer and Smitherman 1980; Chen 1990). These filter-feeding carps can be used to control algal blooms (Xie and Liu 2001) despite lacking a specific, stable ecological niche (Spataru and Gophen 1985) and exhibiting a diet choice largely depending on their stocking density (Wilson and Turelli 1986; Rowland et al. 2006). In addition to filter-feeding carps, filter-feeding tilapias (Oreochromis niloticus) have been used in China (Lu et al. 2006) and in tropical semiarid regions of Brazil (Menezes et al. 2010).
Combined restoration methods and macrophyte inoculation
Many of the reviewed biomanipulation studies were accompanied by supplemental restoration measures (i.e., nutrient reduction, drawdown, dredging, macrophyte plantation or Daphnia addition) or combined fish removal with piscivore addition which makes it difficult to interpret various biomanipulation outcomes with regard to cyanobacterial abundance.
Fish removal with piscivore addition
Examples of successful cyanobacterial control after fish removal with the addition of piscivorous fish (Reinertsen et al. 1990; Tátrai et al. 2003) show the potential of an increased grazing pressure of zooplankton (and concomitant macrophyte establishment) or reduced nutrient cycling in controlling cyanobacterial blooms. Nevertheless, the added value of piscivore addition has been contested in some of the studies where both measures were combined (Shapiro and Wright 1984; Meijer et al. 1999; Kozak and Goldyn 2004; Søndergaard et al. 2008; De Backer et al. 2011). Because generally the stocking of piscivores shows limited or no success in the absence of other (eco)technological measures (see Sect. 3.2), it seems that active fish removal constitutes the key component controlling cyanobacteria when both methods are combined. In the Feldberger Haussee Reservoir in Germany (Kasprzak et al. 2003, 2007), the combination of fish removal and piscivore stocking only had a limited success probably due to insufficient removal of planktivorous fish plus a low yield of piscivorous fish. In an Australian wetland system, invasive carp removal during multiple years combined with stocking Australian bass Macquaria novemaculeata could significantly reduce cyanobacterial dominance (Pinto et al. 2005).
Piscivore and Daphnia addition
We found one study that combined piscivore stocking (Oncorhynchus mykiss) with Daphnia stocking (Ha et al. 2013) (Fig. 1). The authors concluded that biomanipulation using rainbow trout and Daphnia stocking succeeded in improving water quality of Lake Shirakaba (Japan) by reducing algal abundance and a disappearance of Microcystis blooms.
Water drawdown with fish removal
In flow-through systems that can be hydrologically controlled, biomanipulation was accompanied by water drawdown to facilitate fish removal (van de Bund and van Donk 2002; Van Wichelen et al. 2007; Peretyatko et al. 2009). Oxygenation, mineralization and compaction of exposed sediments may act in combination with the down-sizing of fish populations to impair algal and cyanobacterial blooms after refilling (Cooke et al. 2005; Van Wichelen et al. 2007). The return of a desired ecological status might, however, not be plausible as long as sediments have not been dredged, for instance if oxidation of sulfides leads to strong acidification of the water (Van Wichelen et al. 2007).
A possible, but speculative, advantage of water drawdown could be the destruction of cyanobacterial resting stages accumulated on the sediment during years of bloom formation. Since massive emergence under favorable conditions can lead to cyanobacterial blooms (Microcystis: Ståhl-Delbanco et al. 2003) and relationships exist between numbers of settled resting stages and ensuing water column concentrations (Anabaena flos-aquae: Kravchuk et al. 2011), the termination of in situ recruitment could decrease the competitive abilities of cyanobacteria within the plankton community and prolong the positive effect of biomanipulation. However, survival of a small fraction of settled akinetes or colonies might be enough to generate noxious blooms in summer (Cirés et al. 2013).
Direct macrophyte stimulation
In shallow lakes and ponds, macrophyte recovery can be directly assisted by introducing whole plant shoots or thalli or by translocating either propagules or propagule-rich sediments (Smart et al. 1998; Søndergaard et al. 2000; Hilt et al. 2006) (Fig. 1). This form of biomanipulation alone is not likely to promote a sudden shift from very turbid, cyanobacteria-infested water to a clear state, but could accelerate lake-wide macrophyte establishment in combination with other restoration measures (Donabaum et al. 1999). Inoculating a very shallow (e.g., <0.50 m), sufficiently large zone with macrophytes also could locally lower cyanobacterial abundance (as shown in a microcosm experiment; Wang et al. 2012) and increase water transparency in the immediate surroundings of the macrophyte stands (van den Berg et al. 1998), thus permitting a gradual expansion over multiple years (van den Berg et al. 2001). This could eventually disrupt the dominance of cyanobacteria in the whole water body, provided that macrophyte expansion is not significantly hindered.
Stocking with alien species
Although stocking with alien species is not the rule, sometimes commercially important piscivore fish species (Salmo trutta lacustris, Oncorhynchus mykiss) have been introduced in a water body as a restoration tool (Scharf 2007; Ha et al. 2013) without fully knowing the impact they have on local native biodiversity. Gulati et al. (2008) discussed in detail the use of invasive zebra mussel (Dreissena polymorpha) to control cyanobacteria (Fig. 1). Apparently, there are some promising results showing that zebra mussels filter and ingest Microcystis under laboratory conditions (Baker et al. 1998; Dionisio Pires et al. 2005). Large-scale enclosure experiments, in systems where the zebra mussel is already present, investigating the effect of zebra mussels on both native biodiversity or cyanobacteria blooms have not, to our knowledge, been conducted yet in Europe. On the other hand in the USA, where they combined field surveys with large-scale experiments, a higher biomass of M. aeruginosa and Anabeana spp. was found in the presence of D. polymorpha (Raikow et al. 2004; Knoll et al. 2008).
It is striking that most of the studies dealing with D. polymorpha as a biomanipulation tool neglect largely the potential ecological impacts of introducing invasive mussels. Dreissena polymorpha is an invasive species in many European countries (Karatayev et al. 2007; Hallstan et al. 2010; Oscoz et al. 2010; Sousa et al. 2011) and recognized as an “ecosystem engineer” (Sousa et al. 2009). Vilà et al. (2010) even placed the zebra mussel in the top 10 of invasive species with detrimental impacts on a wide range of ecosystem services. Established zebra mussels usually compete with native bivalves and other benthic invertebrates, alter the nutrient cycle considerably and disturb the benthic structure and fish community (Strayer 2009; Higgins and Vander Zanden 2010). Many of these effects encountered in the Great Lakes of the USA have hardly been investigated in Europe. Dionisio Pires et al. (2005) found no significant differences in the clearance rates on cyanobacteria between Daphnia galeata and Dreissena polymorpha. Moreover, Bontes et al. (2007) and Dionisio Pires et al. (2007) showed that native unionid mussels are also able to filter and ingest Microcystis and Oscillatoria. Recently, He et al. (2014) showed a positive association between the development of macrophyte beds of Vallisneria natans and the presence of the native bivalve Hyriopsis cumingii. Given the abovementioned ecological impacts, the introduction of invasive mussels is not recommended. The practical application of using native mussels for biomanipulation of cyanobacteria presents some considerable limitations (substrate suitability, culturing…), but their use is considered worthy of further investigation.
Causes of success or failure to control cyanobacteria
Bottlenecks of fish removal
The shock effect of a sufficiently large reduction in cyprinid fish mostly inhibits cyanobacterial abundance on short notice, especially early in the growth season. Through time, increased dominance and reoccurrence of summer blooms appear to be inevitable in many cases of fish removal (Meijer et al. 1999; van de Bund and van Donk 2002; Beklioglu and Tan 2008; Søndergaard et al. 2008). Because a principle goal of biomanipulation is to lower the risk of cyanobacterial blooms, a return to turbid conditions but without dominance of cyanobacteria and related health risks could still be considered as a partial temporary success.
Fish: scale of reduction, in-lake recruitment and recolonization
A primary cause for biomanipulation failure or lack of prolonged effects is insufficient fish removal, or a quick recovery of zooplanktivorous populations via recruitment or dispersal. Cyanobacterial abundance is not affected when the minimal catch has not been reached (Olin et al. 2006; Søndergaard et al. 2008). A complete removal of fish often results in a trophic cascade powerful enough to allow cyanobacteria to be controlled, even in the absence of macrophytes (van de Bund and van Donk 2002; Peretyatko et al. 2012a), but in some instances, the uncontrolled development of invertebrate zooplanktivores (especially Chaoborus) may induce a collapse of Daphnia populations (Benndorf 1995). Whether or not the filtering action of Daphnia and its limitations increase the probability of cyanobacterial blooms, might depend on taxon identity and the size of the daphnid species (see Sect. 2.1.2). Some studies clearly demonstrate the ability of Daphnia to reduce cyanobacteria after biomanipulation (Benndorf 1995; Annadotter et al. 1999; Peretyatko et al. 2012a; Ekvall et al. 2014), even under high toxicity, while others found Daphnia to be incapable of or insufficiently abundant for cyanobacterial control (Hanson and Butler 1994; Horppila et al. 1998; Dawidowicz et al. 2002; Pedusaar et al. 2010).
In a study conducted by Olin et al. (2006), cyanobacterial biomass decreased in European boreal lakes (Finland) in which a considerable portion of larger cyprinid fish had been removed over a number of years. Only when the catch exceeded >200 kg ha−1 within 3 years, chances of reduced cyanobacterial occurrence were high, and blooms of cyanobacteria were delayed toward the end of the growth season while duration shortened.
In the large Enonselkä basin of Lake Vesijärvi (Finland), biomanipulation was conducted in order to tackle massive blooms of Aphanizomenon flos-aquae and Microcystis spp. (Horppila et al. 1998). Cyanobacterial blooms were successfully inhibited, and summer average biomass of cyanobacteria decreased from 1.4 mg L−1 to values below 0.4 mg L−1. There was no evidence for a strong trophic cascade, and consequently, it was hypothesized that biomanipulation decreased resuspension by bottom-dwelling fish as well as translocation of P from the littoral to the pelagic zone via migrating roach (Horppila et al. 1998; Keto and Tallberg 2000). Alternatively, decreased pH values following biomanipulation might have inhibited cyanobacterial growth (Keto and Tallberg 2000), analogous to the observed influence of pH on cyanobacterial bloom development in hypereutrophic ponds (Peretyatko et al. 2012b).
At similar latitude, attempts to restore Lake Ülemiste in Estonia by fish removal and reduced external loading led to a reduction in phytoplankton biomass, although only spring Secchi depth improved (Pedusaar et al. 2010). The role of Daphnia filtration proved to be negligible. The lake had been dominated by Limnothrix redekei, but biomanipulation induced a shift in favor of chroococcal species (Microcystis, Aphanocapsa and others). Those taxa gained a bigger share within the cyanobacterial assemblage and co-dominated with filamentous species in subsequent years. Only in spring did eukaryotic taxa dominate the phytoplankton. In Lake Ringsjön (Sweden), continued fish removal during 8 years has gradually improved the ecological situation by decreasing cyanobacterial biomass, as shown by Cronberg 1999 and more recently by Ekvall et al. (2014). Biomass of cyanobacteria in June significantly decreased below 2 mg L−1 resulting from an increasingly effective top-down control. Microcystin levels for June dropped compared to the concentration before fish reduction. Data for July and August showed no amelioration through time, however, either because of succession of cyanobacterial species or predation by young-of-the-year fish.
Efficacy of biomanipulation in moderately interconnected, (hyper)eutrophic ponds has been tested in peri-urban parks and forested zones at the Brussels-Capital Region, Belgium (Peretyatko et al. 2009; De Backer et al. 2012). Prior to biomanipulation, ponds frequently endured toxic cyanobacterial blooms. Complete draining removed all fish. After biomanipulation, Daphnia controlled phytoplankton biomass and turbidity, in most cases enabling extensive growth of submerged macrophytes (Peretyatko et al. 2009). Where submerged vegetation was not restored, a clear-water state was maintained by intense cladoceran grazing. The high filtration rates did not lead to a shift from dominance of edible, eukaryotic phytoplankton toward colonial or filamentous cyanobacteria, suggesting that zooplankton alone could prevent cyanobacterial bloom formation in eutrophic, fishless ponds (Peretyatko et al. 2009, 2012a). In one pond, still lacking submerged macrophytes several years after biomanipulation, a bloom of Aphanizomenon occurred as soon as fish reappeared (Peretyatko et al. 2012a).
When biomanipulation only briefly restored a clear-water phase, a generally lowered cyanobacterial biovolume was still apparent even after build-up of total phytoplankton biovolume to pre-management conditions (Peretyatko et al. 2012a). However, in these situations, cyanobacterial peaks were higher than those observed in successful cases and periods, which indicates that a collapse of the clear-water state increases the risk of blooms of cyanobacteria.
Macrophytes: extent of cover and stability
The importance of submerged macrophytes for the stability of a clear-water state has been emphasized by various authors (Scheffer et al. 1993; Søndergaard et al. 2008). Results of fish removal in shallow lakes in the Netherlands showed an increased resilience above 25 % surface cover by macrophytes (Meijer et al. 1999), a value similar to the threshold of 30 % observed for Danish lakes (Jeppesen et al. 1990) and peri-urban ponds in Belgium (De Backer et al. 2012) (Fig. 2). In a meta-analysis of Danish shallow lakes, drastic fish removal significantly decreased cyanobacterial biomass up till 5 years (Søndergaard et al. 2008). In many of these lakes, macrophytes quickly colonized large parts. A 5-year span of cyanobacterial bloom prevention resulting from macrophyte reappearance has also been found in a Mediterranean, semiarid lake (Lake Eymir, Turkey; Beklioglu and Tan 2008).
In the Netherlands, Lake Zwemlust was successfully biomanipulated by replacing the original fish community with rudd and small pike (van de Bund and van Donk 2002). Prior to biomanipulation, blooms of Microcystis aeruginosa intervened with recreational activities. After biomanipulation, an extended clear-water phase characterized by high Daphnia grazing allowed submerged macrophytes to suppress cyanobacterial dominance in summer. This situation lasted for several years, until succession led to a transition from Elodea nuttallii to Potamogeton berchtoldii dominance, which proved sensitive to decline. The loss of macrophytes initiated, at first, shifts between clear and turbid water within the same growing season, and finally, reoccurrence of cyanobacterial blooms. In the sixth summer after biomanipulation, a peak of Anabaena and Pseudanabaena signaled the initial intervention had reached its limits (Romo et al. 1996).
Strong positive effects of combined fish and wave reduction were observed by Bontes et al. (2006) in an exclosure experiment in a turbid lake dominated by Limnothrix, Oscillatoria and Microcystis (Terra Nova, the Netherlands). In the biomanipulated compartment, water was clear and filamentous algae and submerged macrophytes grew extensively. Cyanobacteria were negatively affected (mean 16,298 cells mL−1) compared to the in-lake reference site (mean 86,070 cells mL−1). Whole-scale removal of benthivorous and planktivorous fish in Lake Terra Nova resulted in high cladoceran densities and allowed macrophyte recolonization, and significantly inhibited filamentous cyanobacteria (ter Heerdt and Hootsmans 2007).
In biomanipulated peri-urban ponds (Brussels, Belgium), the most essential biotic component stabilizing the clear-water state turned out to be the extent of submerged macrophyte cover. Below 30 % surface cover, ponds tended to shift back to turbid conditions after fish recolonization (De Backer et al. 2012). In general, the presence of fish did not significantly increase phytoplankton biovolume if more than 30 % of the surface area was covered by submerged macrophytes, although in the absence of large Cladocera at TP concentrations exceeding 0.300 mg L−1, submerged macrophytes could maintain a clear-water state only when cover reached at least 82 % at some point during the growth season (De Backer et al. 2014) (Fig. 2).
Dynamics of nutrient status and pH
Ultimately, avoidance of cyanobacterial blooms and the stability of the clear-water state rely on favorable nutrient dynamics. Positive effects of biomanipulation will be prolonged if nutrient concentrations have been restricted prior to management of the food web. Pot and ter Heerdt (2014) analyzed successful recolonization of aquatic vegetation in a biomanipulated shallow lake dominated by Oscillatoria agardhii following gradual reductions of external nutrient loading. On the contrary, Pedusaar et al. (2010) found limited effects of biomanipulation on water clarity, although nutrient levels had been reduced prior to, as well as because of, fish removal. Additional to anthropogenically induced nutrient changes, stochastic events like droughts and associated nitrogen fluxes could trigger reoccurrence of cyanobacteria after biomanipulation (Beklioglu and Tan 2008).
Teissier et al. (2012) found a weakened relationship between TP and Chl a concentrations in the water column of urban and peri-urban ponds (Brussels, Belgium) with an extensive submerged macrophyte cover (>30 %) and/or efficient zooplankton grazers (mean large cladoceran length >1 mm) compared to cyanobacteria-infested ponds before biomanipulation and those having only sparse vegetation (<30 % cover). De Backer et al. (2014) identified the TP threshold level, namely a summer average of 0.300 mg P L−1, above which maintenance of the clear-water state is highly unlikely, even after an initial success (Fig. 2). This TP threshold is markedly higher than those suggested for shallow lakes (0.062 mg P L−1: Zimmer et al. 2009; 0.080–0.150 mg P L−1: Jeppesen et al. 1990; 0.100 mg P L−1: Hosper and Jagtman 1990), most likely because of a relatively higher vegetation cover and cladoceran grazing pressure in the peri-urban ponds (De Backer et al. 2014).
The potential impact of acidification following biomanipulation was demonstrated in Lake Kraenepoel, Belgium. Complete drawdown with fish removal and drying of the sediment layer for an extended period of time broke the dominance of large-sized cyanobacteria and euglenoids (Van Wichelen et al. 2007). Initially after refilling, high grazing rates controlled phytoplankton, but oxidation of sulfides within the organically rich sediments during drawdown dramatically acidified the water column later in the year, with pH values as low as 3.2. Although this resulted in clear water during the remaining study period, the low pH prevented successful recovery of the submerged macrophyte vegetation. At the same time, the northern basin was successfully restored because organic sediments were removed prior to refilling.
There is a substantial overlap in factors that favor cyanobacteria in (hyper)eutrophic ponds. Shallowest and smallest urban ponds warm up fast and develop substantial phytoplankton biomass. Because of their richness in phosphorus and nitrogen, intense phytoplankton growth eventually leads to depletion of free CO2 and associated pH increase. Persisting high pH conditions give a competitive advantage to bloom-forming cyanobacteria that are well adapted to such conditions (Shapiro 1973, 1997). The combination of phytoplankton biomass and pH appeared to be the best predictor for cyanobacterial blooms in (peri-)urban ponds, showing that most cyanobacterial blooms are confined to Secchi depth below 0.6 m and, more importantly, pH above 8 (Peretyatko et al. 2010, 2012b).
Some very shallow, peri-urban ponds (Brussels) at high pH have developed elevated cyanobacterial biomass despite biomanipulation and removal of nearly all fish. Mesocosms in such biomanipulated ponds, previously affected by severe blooms, were used to test the hypothesis of lowered pH-reducing cyanobacteria development (Teissier et al. 2011). Two out of three mesocosms at pH > 8.5 showed a strong response during their experiment that lasted 38 days. An artificial experimental lowering of pH around 7 through addition of CO2 in the water column kept the cyanobacterial biomass low (not the eukaryotic phytoplankton) in nine mesocosms.
Although it is difficult to conclude with certainty that lowered pH also could be a management tool for abatement of already built-up cyanobacterial blooms, its capacity for prevention of blooms seems sufficiently important (Fig. 2). Although large mesocosms of 1 m3 were considered, the pH management still must be achieved at the whole pond scale. Such results in very shallow urban ponds clearly indicate that elevated in situ pH conditions following biomanipulation may jeopardize the effectiveness of fish removal on the cyanobacterial bloom formations. Liming, a common practice among pond managers and owners to increase pH, therefore should be avoided and discouraged in the light of these experimental field observations.
Bottlenecks of piscivore addition
Limited success rate
We found few studies that reported a successful decline of cyanobacteria after piscivore stocking as the only restoration measure. In Lake Gjersjoen, Norway, the dominance of Oscillatoria agardhii decreased after stocking with pike-perch (Lyche 1990). A strong top-down control on phytoplankton biomass was also found in Lake Lyng, Denmark, after repeated stocking with pike (Søndergaard et al. 1997). Phytoplankton biomass was dominated by cyanobacteria before biomanipulation (mainly Anabaena spiroides and A. flos-aquae). After biomanipulation, a switch occurred in the phytoplankton community toward a dominance of cryptophytes and chrysophytes. Cyanobacteria contribution to the total phytoplankton biomass decreased from 61 % before biomanipulation until 14 %, 5 years after biomanipulation (Søndergaard et al. 1997). The authors argued however that the effect of stocking is only successful in the year that the biomanipulation took place and that a sufficient macrophyte cover is essential to maintain a permanent reduction in phytoplankton.
Failure of top-down control
Drenner and Hambright (2002) reviewed 17 studies examining the effects of introducing piscivores on phytoplankton biomass and found that for the majority of the studies, the biomanipulation measure had no effect on reducing the algal abundance. For example, Drenner et al. (2002) evaluated the stocking of largemouth bass in a eutrophic reservoir in Texas, USA. Although the effects of bass cascaded down through the food web to the phytoplankton, densities of cyanobacteria did not change after stocking of largemouth bass. On the contrary, they observed a shift from relatively edible phytoplankton to large cyanobacteria. In deep Lake Mutek (Poland), piscivore stocking lowered densities of zooplanktivorous fish, but success was only partial since an increase in small-bodied cladocerans in the epilimnion was associated with a phytoplankton community shift in favor of filamentous cyanobacteria (Dawidowicz et al. 2002).
The hypereutrophic Bautzen reservoir in Germany has a long history of piscivore biomanipulation starting in 1981. The reservoir (surface area 5.33 km2) has a mean depth of 7.4 m and showed regular blooms of cyanobacteria caused by high nutrient loading. The reservoir was regularly stocked with pike-perch, pike, European catfish and eel supplemented with catch restrictions for anglers. The effects varied during the 18 year post-biomanipulation period (Kasprzak et al. 2007). After biomanipulation had begun (period 1981–1993; piscivore percentage of 10–40 %) and phosphorus loading had risen, there was a decline of the planktivorous fish population and an increase in the relative abundance of Daphnia galeata but inedible phytoplankton significantly increased due to Microcystis blooms. The period from 1994 to 1999 (>40 % piscivores) was associated with a drop in inedible phytoplankton biomass after reduced phosphorus loading, due to the use of a water treatment technology (Deppe et al. 1999), but Daphnia biomass declined again to values similar to the pre-biomanipulation period. Thus, the biomanipulation was only partly successful and primarily controlled by nutrient loading.
A first potential mechanism involved in the lack of support for the trophic cascade hypothesis is the gape limitation of piscivore fish (Hambright 1994). The presence of a predatory fish can alter the composition and size distribution of the prey fish community (Nowlin et al. 2006). If larger-bodied, less vulnerable species are dominating the fish community, this can prevent the effects of piscivore addition cascading down to the phytoplankton. In particular, pike have been used to control the young-of-the-year (YOY) planktivorous fish such as roach Rutilus rutilus and/or bream Abramis brama (Søndergaard et al. 1997; Skov et al. 2002; De Backer 2011). Both post-stocking mortality due to cannibalism and lack of piscivory among the stocked pike due to their small size (Skov et al. 2002) are important factors for the failure of pike introductions. In order to have an effect on the plankton level, high densities (0.1 individuals m−2 year−1, Søndergaard et al. 2000) are required (Fig. 2), stocking time and body length (larger than the conventional of <30 cm) of pike will be essential (Skov and Nilsson 2007) and the restoration measure needs to be repeated regularly to achieve stability (Søndergaard et al. 2000).
Benndorf et al. (2000) showed that strong piscivory can have the same effects on suppressing large zooplankton and water transparency as planktivorous fish, a process called “over-biomanipulation.” In systems where almost all planktivorous fish are eliminated, uncontrolled development of invertebrate predators such as Chaoborus sp. can exert a strong predation pressure on large zooplankton compensating for the effects of the piscivore addition (Fig. 1). Even if piscivore addition resulted in an increase in large zooplankton, as was the case in the Bautzen reservoir (Kasprzak et al. 2007), high nutrient loading can prevent the top-down control. Further, the intense grazing of large zooplankton, enhanced by the piscivore addition, can shift the phytoplankton assemblage toward harmful cyanobacteria (Drenner et al. 2002) that can have toxic effects on zooplankton survival (Olvera-Ramirez et al. 2010). An inhibition of large Cladocera because of their susceptibility to toxins would limit feeding rates (Paerl et al. 2001). Some authors however suggested that the effect of zooplankton on consuming cyanobacteria is negligible (Boon et al. 1994; Degans and De Meester 2002), while more recent evidence suggests that zooplankton are able to control different cyanobacteria species (Gloeotrichia echinulata, Cylindrospermopsis raciborskii, Anabaena solitaria and A. flos-aquae) and can lower microcystin concentrations, both in the (sub)tropics as in temperate regions (Fey et al. 2010; Kâ et al. 2012; Ekvall et al. 2014).
Bottlenecks of stocking with filter-feeding fish
Stocking densities
Increased stocking of subtropical lakes with both filter-feeding planktivorous silver carp and bighead carp could at least suppress Microcystis blooms directly by grazing in Lake Donghu near the Yangtze River (China) (Xie and Liu 2001; Zhang et al. 2008). Although high fish stocking levels lowered total phytoplankton biomass, Microcystis biomass and microcystin concentrations, when compared to surrounding lake water, were statistically insignificant (Ke et al. 2007). Silver carp appeared to feed mainly on phytoplankton, whereas bighead carp mainly on zooplankton (>60 %). Both carp species however preyed upon more zooplankton because of the abundant food resource (Ke et al. 2007). When silver and bighead carps are used to control cyanobacterial blooms, a sufficiently high stocking density is very important for a successful practice. At too low stocking densities, silver and bighead carps would experience less competition and shift their feeding behavior toward zooplankton, thereby decreasing the overall grazing efficiency of cyanobacterial bloom reduction (Ke et al. 2008) (Fig. 1). Silver and bighead carp should be kept at a stocking threshold of about 50 g m−3 (Fig. 2) because a decline in filter-feeding grazing would make a water body again vulnerable to cyanobacterial bloom (Xie and Liu 2001).
An interesting in situ experiment was conducted by Ke et al. (2008) in a bay of the shallow hypereutrophic subtropical Lake Taihu in China with severe Microcystis blooms that persisted over decades. Meiliang Bay (northern part of Lake Taihu) has a surface area of 100 km2 and an average depth of 2 m (ranging from 1.8 to 2.3 m). Surface water temperatures ranged seasonally from 12.7 (November) to 33.6 °C (August). The main purpose was to provide evidence for controlling cyanobacterial blooms by using silver carp (70 %) and bighead carps (30 %) at different stocking densities under natural changing environmental conditions during 2 years (2004 and 2005). Ke et al. (2008) observed that both carp species in fact grazed more on zooplankton than on phytoplankton. Chlorophyta were a main food in spring, whereas cyanobacteria (Microcystis) reached highest diet percentages in summer months. Only in the warmest months, Microcystis temporarily contributed 80–100 % of the phytoplankton in the gut contents of silver and bighead carp. Silver and bighead carps maintained their feeding intensity during blooms (Ke et al. 2008).
Tilapia can ingest and digest a large quantity (58–78 %) of Microcystis with ingestion rate increasing with fish weight (Lu et al. 2006). Initial cyanobacterial (Microcystis aeruginosa, Oscillatoria princes and Spirulina maxima) biomass of 70 % (relative to the total phytoplankton biomass) was reduced to much lower levels of 11–22 % in Lake Yuehu. The order of magnitudes however remained high and reached hundreds of 106 cells L−1 with Secchi depths of mostly <1 m after treatment (Lu et al. 2006). A thick surface scum of Microcystis aeruginosa and Anabaena spiroides disappeared in a small pond, about 20 days after tilapia fingerlings were stocked (Lu et al. 2006). Stocking densities of 3–15 g m−3 were reported to control blooms. Whole-lake stocking strongly reduced cyanobacteria (Oscillatoria princes and Microcystis aeruginosa), thereby increasing Secchi depth but also nutrient levels (TP, TN, COD) and changing phytoplankton composition toward a dominance of Cryptophyta and Bacillariophyta. Among filter feeders, tilapia appears to be better in controlling blooms (Lu et al. 2006).
Outside Eastern Asia, few successful attempts to control cyanobacteria with filter-feeding fish have been reported. Microcystis aeruginosa and Botryococcus braunii (Chlorophyta) ingestion by silver carp was reported for a tropical urban lake in Brazil (Starling et al. 1998). Reduction in tilapia biomass to 40 g m−3 resulted in significant water quality improvements, and Microcystis abatement by stocking silver carp at moderate fish stocking densities of 40–50 g m−3 induced significant decreases in cyanobacteria density (40–44 %), phytoplankton biomass (22–38 %) and TP (21–31 %). Reducing tilapia and adding silver carp were found to act independent, and the combination of both filter-feeding fish reduced TP by 38 %, cyanobacteria density by 75 % and phytoplankton biomass by 60 % (Starling 1998). Likewise, in Israel, a reduction in cyanobacteria was found due to stocking with silver carp (Leventer and Teltsch 1990). Overall, this method of stocking fairly large amounts of filter-feeding fish is considered as an effective management technique in eutrophic water bodies that substantially lack macrozooplankton (Zhang et al. 2008).
Persistence of Microcystis under fish pressure
High stocking rates of tilapia (>100 g m−3) can promote blooms of Microcystis and decrease water clarity by supplying phytoplankton with additional nutrients, mainly phosphorus (Starling 1998). Microcystis and Aphanizomenon spp. also were shown to survive gut passage in fish and may use the internally supplied phosphorus. A mucilage layer around Microcystis protects cells from digestion (Lewin et al. 2003). Silver carp could effectively ingest toxic Microcystis cells up to 84.4 % of total consumed phytoplankton (Chen et al. 2006). In vitro experiments demonstrated that feces of tilapia, silver and bighead carp contained Microcystis cells (Datta and Jana 1998). Such feces containing small colonies or single cells will sink to the bottom where recruitment of nutrient-enriched Microcystis may occur. It might be hypothesized that the desired grazing effect of silver carp on Microcystis does not outweigh the overall effect of additional nutrient recycling induced by the stocked filter-feeding fish. Biomanipulation should have a minimum effect of ichthyo-eutrophication, by applying appropriate stocking (Ke et al. 2007).
Attempts to use silver carp in temperate regions
Addition of silver carp in temperate water bodies, mesocosm attempts in France and repeated stocking in the Saidenbach Reservoir in Germany, drastically reduced the large cladoceran biomass because zooplankton appeared to be the main source of food. Small-sized phytoplankton (<20 µm) is not grazed down effectively by silver carp, and more importantly, silver carp did not effectively reduce cyanobacteria even at higher stock levels (Domaizon and Devaux 1999; Horn 2003).
Biomanipulation strategies as nature-based solutions to minimize adverse effects of cyanobacteria and to maximize ecosystem services
The different methods of biomanipulation emphasize interventions at different food-web levels and trophic degrees (Fig. 2). The initial conditions determine the biomanipulation strategy for cyanobacterial control. In heavily infested (sub-)tropical water bodies, abatement is aimed at the addition of filter-feeding fish (level 3), partially removing a rather persistent bloom of Microcystis (level 1) as forced biotic key interactions. This direct cyanobacterial bloom abatement through enhancement of productivity at trophic level 3 requires minimal management or engineering intervention to maximize the selected key service (Fig. 3). Rearing, caging, adding and controlling filter-feeding fish populations contribute minimally to ecosystem services of such (hyper)eutrophic water bodies. Stocking water bodies with phytoplanktivorous fish therefore can be considered as a method to temporarily control visible nuisance algal blooms to a certain extent in shallow hypereutrophic (sub)tropical lakes where zooplankton cannot effectively control phytoplankton production.

Conceptual scheme of nature-based solutions at different levels of the cascading food web and the benthic compartment according to maximization of either key services or number of ecosystem services (subdivision of water body types is similar to Fig. 2)
Small-sized man-made urban ponds that are annually infested during long periods by various cyanobacterial taxa can be managed easily and cheaply through level 3 removal of (nearly) all plankti-, benthi- and molluscivorous fish to obtain clear-water conditions (Fig. 2). Such fishless water bodies can be managed at highest TP levels when compared to all other biomanipulation strategies and only occasionally may show again phytoplankton build-up after collapse of the benthic submerged macrophyte beds or the return of young fish (through incomplete removal or deliberately stocked by fishermen) grazing on large zooplankton. The main aim of this nature-based solution is forcing the biotic key interactions between level 1 and level 2 thereby physically reducing internal P-loading through lowered bioturbidity of fish and positive feedback of submerged aquatic vegetation (Fig. 3). A (repeatedly) complete drawdown and fish removal strongly reduce the risk of cyanobacterial bloom events, thereby improving the multi-functionality (e.g., water retention, nature conservation, recreation, fishing activities) and number of ecosystem services of the urban ponds. Accompanying management or engineering interventions could be the water-level adjustment to lower the negative effects of bird herbivory. Liming of man-made ponds should be discouraged because pH levels above 8 most likely favor cyanobacterial blooms (Fig. 3).
The biomanipulation strategies to avoid cyanobacterial blooms in shallow lakes and large ponds are multiple and aim at first eutrophication control and secondly at the enhancement of functional biodiversity in each compartment (Fig. 2). Stabilized communities at each trophic level are envisaged through partial removal of plankti-, benthi- and molluscivorous fish, selective addition of piscivores and spontaneous recovery of balanced submerged aquatic vegetation (Fig. 3). Accompanying management or engineering interventions to maximize the number of ecosystem services may consist of further reducing external and internal TP to mesotrophic levels (catchment land use changes, TP fixation/oxygenation), sediment removal, creation of sediment islands (Gulati et al. 2008), fishing regulations (restrictions or even prohibition) or even addition of large Cladocera (Fig. 3). The latter suite of nature-based solutions is the preferred strategy, not solely to avoid cyanobacterial blooms but also to stabilize various biotic interactions between each level of the cascading food web.
Future studies on biomanipulation to control cyanobacteria should consider more detailed information about the cyanobacteria, their ratio within the phytoplankton community and specifications on colonial versus filamentous, edible versus non-edible, N2-fixing versus non-fixing, vacuolated versus non-vacuolated strains. This will provide more information on the dynamics and abundance of cyanobacteria taxa before and after a combination of whole-lake biomanipulation and associated restoration methods to better evaluate the overall applicability and effectiveness of these nature-based solutions.
References
Annadotter H, Cronberg G, Aagren R, Lundstedt B, Nilsson P, Ströbeck S (1999) Multiple techniques for lake restoration. Hydrobiologia 395(396):77–85
Asselman J, Hochmuth JD, De Schamphelaere KAC (2014) A comparison of the sensitivities of Daphnia magna and Daphnia pulex to six different cyanobacteria. Harmful Algae 39:1–7
Baker SM, Levinton JS, Kurdziel JP, Shumway SE (1998) Selective feeding and biodeposition by zebra mussels and their relation to changes in phytoplankton composition and seston load. J Shellfish Res 17:1207–1213
Beklioglu M, Tan CA (2008) Restoration of a shallow Mediterranean lake by biomanipulation complicated by drought. Fund Appl Limnol 171:105–118
Benndorf J (1995) Possibilities and limits for controlling eutrophication by biomanipulation. Int Revue Ges Hydrobiol 80:519–534
Benndorf J, Wissel B, Sell AF, Hornig U, Ritter P, Böing W (2000) Food web manipulation by extreme enhancement of piscivory: an invertebrate predator compensates for the effects of planktivorous fish on a plankton community. Limnologica 30:235–245
Berg S, Jeppesen E, Søndergaard M (1997) Pike (Esox lucius L.) stocking as a biomanipulation tool 1. Effects on the fish population in Lake Lyng, Denmark. Hydrobiologia 342(343):311–318
Bontes BM, Pel R, Ibelings BW, Boschker HTS, Middelburg JJ, van Donk E (2006) The effects of biomanipulation on the biogeochemistry, carbon isotopic composition and pelagic food web relations of a shallow lake. Biogeosciences 3:69–83
Bontes BM, Verschoor AM, Dionisio Pires LM, van Donk E, Ibelings BW (2007) Functional response of Anodonta anatina feeding on a green alga and four strains of cyanobacteria, differing in shape, size and toxicity. Hydrobiologia 584:191–204
Boon PI, Bunn SE, Green JD, Shiel RJ (1994) Consumption of cyanobacteria by freshwater zooplankton; implications for the success of ‘top town’ control of cyanobacterial blooms in Australia. Aust J Mar Freshw Res 45:875–887
Burks RL, Lodge DM, Jeppesen E, Lauridsen TL (2002) Diel horizontal migration of zooplankton: costs and benefits of inhabiting the littoral. Freshw Biol 47:343–365
Carpenter SR, Kitchell JF, Hodgson JR (1985) Cascading trophic interactions and lake productivity. Bioscience 35:634–639
Chen SL (1990) Fish and its role on nutrient cycling in water. In: Liu JK (ed) Ecological studies of Lake Donghu (1). Science Press, Beijing, pp 292–371 (in Chinese)
Chen J, Xie P, Zhang D, Ke ZX, Yang H (2006) In situ studies on the bioaccumulation of microcystins in the phytoplanktivorous silver carp (Hypophthalmichthys molitrix) stocked in Lake Taihu with dense toxic Microcystis blooms. Aquaculture 261:1026–1038
Chislock MF, Sarnelle O, Jernigan LM, Wilson AE (2013) Do high concentrations of microcystin prevent Daphnia control of phytoplankton? Water Res 47:1961–1970
Cirés S, Wörmer L, Agha R, Quesada A (2013) Overwintering populations of Anabaena, Aphanizomenon and Microcystis as potential inocula for summer blooms. J Plankton Res 35:1254–1266
Cooke GD, Welch EB, Peterson SA, Nichols SA (2005) Restoration and management of lakes and reservoirs, 3rd edn. CRC Press, Boca Raton
Cremer MC, Smitherman RO (1980) Food habits and growth of silver and bighead carp in cages and ponds. Aquaculture 20:57–64
Cronberg G (1999) Qualitative and quantitative investigations of phytoplankton in Lake Ringsjön, Scania, Sweden. Hydrobiologia 404:27–40
Datta S, Jana BB (1998) Control of bloom in a tropical lake: grazing efficiency of some herbivorous fishes. J Fish Biol 53:12–24
Dawidowicz P, Prejs A, Engelmayer A, Martyniak A, Kozłowski J, Kufel L, Paradowska M (2002) Hypolimnetic anoxia hampers top-down food-web manipulation in a eutrophic lake. Freshw Biol 47:2401–2409
De Backer S (2011) Restoration of ecological quality in eutrophic peri-urban ponds through complete fish removal and water drawdown: biotic interactions, evaluation and implications for management. Dissertation, Vrije Universiteit Brussel
De Backer S, Teissier S, Triest L (2012) Stabilizing the clear-water state in eutrophic ponds after biomanipulation: submerged vegetation versus fish recolonization. Hydrobiologia 689:161–176
De Backer S, Teissier S, Triest L (2014) Identification of total phosphate, submerged vegetation cover and zooplankton size thresholds for success of biomanipulation in peri-urban eutrophic ponds. Hydrobiologia 737:281–296
Deboom CS, Wahl DH (2014) Piscivore enhancement effects on food webs depend on planktivore body size and species composition in replicated whole lake experiments. Hydrobiologia 736:31–49
Degans H, De Meester L (2002) Top-down control of natural phyto- and bacterioplankton prey communities by Daphnia magna and by the natural zooplankton community of the hypertrophic Lake Blankaart. Hydrobiologia 479:39–49
Deppe T, Ockenfeld K, Meybohm A, Opitz M, Benndorf J (1999) Reduction of Microcystis blooms in a hypertrophic reservoir by a combined ecotechnological strategy. Hydrobiologia 408(409):31–38
Dionisio Pires LM, Ibelings BW, Brehm M, van Donk E (2005) Comparing grazing on lake seston by Dreissena and Daphnia: lessons for biomanipulation. Microb Ecol 50(2):242–252
Dionisio Pires LM, Bontes BM, Samchyshyna L, Jong J, van Donk E, Ibelings BW (2007) Grazing on microcystin-producing and microcystin-free phytoplankters by different filter-feeders: implications for lake restoration. Aquat Sci 69:534–543
Domaizon I, Devaux J (1999) Experiment study of the impacts of silver carp on plankton communities of eutrophic Villerest reservoir (France). Aquat Ecol 33:193–204
Donabaum K, Schagerl M, Dokulil MT (1999) Integrated management to restore macrophyte domination. Hydrobiologia 395(396):87–97
Dörner H, Benndorf J (2003) Piscivory by large eels on young-of-the-year fishes: its potential as a biomanipulation tool. J Fish Biol 62:491–494
Drenner RW, Hambright KD (1999) Review: biomanipulation of fish assemblages as a lake restoration technique. Arch Hydrobiol 146:129–165
Drenner RW, Hambright KD (2002) Piscivores, trophic cascades, and lake management. Sci World J 2:284–307
Drenner RW, Baca RM, Gilroy JS, Ernst MR, Jensen DJ, Marshall DH (2002) Community responses to piscivorous largemouth bass: a biomanipulation experiment. Lake Reserv Manage 18:44–51
Ekvall MK, Urrutia-Cordero P, Hansson L (2014) Linking cascading effects of fish predation and zooplankton grazing to reduced cyanobacterial biomass and toxin levels following biomanipulation. PLoS ONE. doi:10.1371/journal.pone.0112956
European Union (2015) Towards an EU Research and Innovation policy agenda for Nature-Based Solutions & Re-Naturing Cities, Final report, p 74. doi: 10.2777/765301
Fey SB, Mayer ZA, Davis SC, Cottingham KL (2010) Zooplankton grazing of Gloeotrichia echinulata and associated life history consequences. J Plankton Res 32:1337–1347
Gladyshev MI, Chuprov SM, Kolmakov VI, Dubovskaya OP, Zadorin AA, Zuev IV, Ivanova EA, Kravchuk ES (2003) A biomanipulation bypassing the trophic cascade in a small reservoir. Dokl Biol Sci 390:235–236
Gulati RD, van Donk E (2002) Lakes in the Netherlands, their origin, eutrophication and restoration: state-of-the-art review. Hydrobiologia 478:73–106
Gulati RD, Dionisio Pires LM, van Donk E (2008) Lake restoration studies: failures, bottlenecks and prospects of new ecotechnological measures. Linmnologica 38:233–247
Ha J-Y, Saneyoshi M, Park H-D, Toda H, Kitano S, Homma T, Shiina T, Moriyama Y, Chang K-H, Hanazato T (2013) Lake restoration by biomanipulation using piscivore and Daphnia stocking; results of the biomanipulation in Japan. Limnology 14:19–30
Hallstan S, Grandin U, Goedkoop W (2010) Current and modeled potential distribution of the zebra mussel (Dreissena polymorpha) in Sweden. Biol Invasions 12:285–296
Hambright KD (1994) Morphological constraints in the piscivore-planktivore interaction: implications for the trophic cascade hypothesis. Limnol Oceanogr 39:897–912
Hansson L-A, Annadotter H, Bergman E, Hamrin S, Jeppesen E, Kairesalo T, Luokkanen E, Nilsson P-A, Søndergaard M, John Strand J (1998) Biomanipulation as an application of food-chain theory: constraints, synthesis, and recommendations for temperate lakes. Ecosystems 1:558–574
Hanson MA, Butler MG (1994) Responses of plankton, turbidity, and macrophytes to biomanipulation in a shallow prairie lake. Can J Fish Aquat Sci 51:1180–1188
He H, Liu X, Yu J, Li K, Guan B, Jeppesen E, Liu Z (2014) Effects of cyanobacterial blooms on submerged macrophytes alleviated by the native Chinese bivalve Hyriopsis cumingii: a mesocosm experiment study. Ecol Eng 71:363–367
Higgins SN, Vander Zanden MJ (2010) What a difference a species makes: a meta—analysis of dreissenid mussel impacts on freshwater ecosystems. Ecol Monograph 80:179–196
Hilt S, Gross EM, Hupfer M, Morscheid H, Mählmann J, Melzer A, Poltz J, Sandrock S, Scharf E, Schneider S, van de Weyer K (2006) Restoration of submerged vegetation in shallow eutrophic lakes—a guideline and state of the art in Germany. Limnologica 36:155–171
Hobbs WO, Ramstack Hobbs JM, LaFrançois T, Zimmer KD, Theissen KM, Edlund MB, Michelutti N, Butler MG, Hanson MA, Carlson TJ (2012) A 200-year perspective on alternative stable state theory and lake management from a biomanipulated shallow lake. Ecol Appl 22:1483–1496
Horn W (2003) Long-term development of the crustacean plankton in the Saidenbach Reservoir (Germany)—changes, causes, consequences. Hydrobiologia 504:185–192
Horppila J, Peltonen H, Malinen T, Luokkanen E, Kairesalo T (1998) Top-down or bottom-up effects by fish: issues of concern in biomanipulation of lakes. Restor Ecol 6:20–28
Hosper SH, Jagtman E (1990) Biomanipulation additional to nutrient control for restoration of shallow lakes in The Netherlands. Hydrobiologia 200:523–534
Ibelings BW, Portielje R, Lammens EHRR, Noordhuis R, van den Berg MS, Joosse W, Meijer M-L (2007) Resilience of alternative stable states during the recovery of shallow lakes from eutrophication: lake Veluwe as a case study. Ecosystems 10:4–16
Jacobsen L, Berg S, Skov C (2004) Management of lake fish populations and lake fisheries in Denmark: history and current status. Fish Manag Ecol 11:219–224
Jeppesen E, Sammalkorpi I (2002) Lakes. In: Perrow M, Dovy R (eds) Restoration practice. Handbook of ecological restoration, vol 2. Cambridge University Press, Cambridge, pp 297–324
Jeppesen E, Jensen JP, Kristensen P, Søndergaard M, Mortensen E, Sortkjær O, Olrik K (1990) Fish manipulation as a lake restoration tool in shallow, eutrophic, temperate lakes 2: threshold levels, long-term stability and conclusions. Hydrobiologia 200:219–227
Jeppesen E, Meerhoff M, Jacobsen BA, Hansen RS, Søndergaard M, Jensen JP, Lauridsen TL, Mazzeo N, Branco CWC (2007) Restoration of shallow lakes by nutrient control and biomanipulation—the successful strategy varies with lake size and climate. Hydrobiologia 581:269–285
Jeppesen E, Søndergaard M, Lauridsen TL, Davidson TA, Liu Z, Mazzeo N, Trochine C, Özkan K, Jensen HS, Trolle D, Starling F, Lazzaro X, Johansson LS, Bjerring R, Liboriussen L, Larsen SE, Landkildehus F, Egemose S, Meerhoff M (2012) Biomanipulation as a restoration tool to combat eutrophication: recent advances and future challenges. Adv Ecol Res 47:411–488
Jiang X, Yang W, Xiang X, Niu Y, Chen L, Zhang J (2014) Cyanobacteria alter competitive outcomes between Daphnia and Bosmina in dependence on environmental conditions. Fund Appl Limnol 184:11–22
Kâ S, Mendoza-Vera JM, Bouvy M, Champalbert G, N'Gom-Kâ R, Pagano M (2012) Can tropical freshwater zooplankton graze efficiently on cyanobacteria? Hydrobiologia 679:119–138
Karatayev AY, Padilla DK, Minchin D, Boltovskoy D, Burlakova LE (2007) Changes in global economies and trade: the potential spread of exotic freshwater bivalves. Biol Invasions 9:161–180
Kasprzak P, Koschel R, Krienitz L, Gonsiorczyk Anwald K, Laude U, Wysujack K, Brach H, Mehner T (2003) Reduction of nutrient loading, planktivore removal and piscivore stocking as tools in water quality management: the Feldberger Haussee biomanipulation project. Limnologica 33:190–204
Kasprzak P, Benndorf J, Gonsiorczyk T, Koschel R, Krienitz L, Mehner T, Hülsmann S, Schultz H, Wagner A (2007) Reduction of nutrient loading and biomanipulation as tools in water quality management: long-term observations on Bautzen Reservoir and Feldberger Haussee (Germany). Lake Reserv Manage 23:410–427
Ke ZX, Xie P, Guo LG, Liu YQ, Yang H (2007) In situ study on the control of toxic Microcystis blooms using phytoplanktivorous fish in the subtropical Lake Taihu of China: a large fish pen experiment. Aquaculture 265:127–138
Ke ZX, Xie P, Guo LG (2008) In situ study on effect of food competition on diet shifts and growth of silver and bighead carps in large biomanipulation fish pens in Meiliang Bay, Lake Taihu. J Appl Ichthyol 24:263–268
Keto J, Tallberg P (2000) The recovery of Vesijärvi, a lake in southern Finland: water quality and phytoplankton interpretations. Boreal Environ Res 5:15–26
Knoll LB, Sarnelle O, Hamilton SK, Kissman CEH, Wilson AE, Rose JB, Morgan MR (2008) Invasive zebramussels (Dreissena polymorpha) increase cyanobacterial toxin concentrations in low-nutrient lakes. Can J Fish Aquat Sci 65:448–455
Kolmakov VI, Gladyshev MI (2003) Growth and potential photosynthesis of cyanobacteria are stimulated by viable gut passage in crucian carp. Aquat Ecol 37:237–242
Kozak A, Goldyn R (2004) Zooplankton versus phyto- and bacterioplankton in the Maltanski Reservoir (Poland) during an extensive biomanipulation experiment. J Plankton Res 26:37–48
Kravchuk ES, Ivanova EA, Gladyshev MI (2011) Spatial distribution of resting stages (akinetes) of the cyanobacteria Anabaena flos-aquae in sediments and its influence on pelagic populations. Mar Freshw Res 62:450–461
Leventer H, Teltsch B (1990) The contribution of silver carp (Hypophthalmichthys molitrix) to the biological control of Netofa reservoirs. Hydrobiologia 191:47–55
Lewin CW, Kamjunke N, Mehner T (2003) Phosphorus uptake by Microcystis during passage through fish guts. Limnol Oceanogr 48:2392–2396
Lu K, Jin C, Dong S, Gu B, Bowen SH (2006) Feeding and control of blue-green algal blooms by tilapia (Oreochromis niloticus). Hydrobiologia 568:111–120
Lyche A, Faafeng BA, Brabrand A (1990) Predictability and possible mechanisms of plankton response to reduction of piscivorous fish. Hydrobiologia 200(201):251–261
Mehner T, Benndorf J, Kasprzak P, Koschel R (2002) Biomanipulation of lake ecosystems: successful applications and expanding complexity in the underlying science. Freshw Biol 47:2453–2465
Meijer M-L, Hosper H (1997) Effects of biomanipulation in the large and shallow Lake Wolderwijd, The Netherlands. Hydrobiologia 342(343):335–349
Meijer M-L, De Boois I, Scheffer M, Portielje R, Hosper H (1999) Biomanipulation in shallow lakes in the Netherlands: an evaluation of 18 case studies. Hydrobiologia 409:13–30
Menezes RF, Attayde JL, Rivera Vasconcelos F (2010) Effects of omnivorous filterfeeding fish and nutrient enrichment on the plankton community and water transparency of a tropical reservoir. Freshw Biol 55:767–779
Nowlin WH, Drenner RW, Guckenberger RW, Lauden MA, Alonso GT, Fennell JE, Smith JL (2006) Gape limitation, prey size refuges and the top–down impacts of piscivorous largemouth bass in shallow pond ecosystems. Hydrobiologia 563:357–369
Olin M, Rask M, Ruuhijärvi J, Keskitalo J, Horppila J, Tallberg P, Taponen T, Lehtovaara A, Sammalkorpi I (2006) Effects of biomanipulation on fish and plankton communities in ten eutrophic lakes of southern Finland. Hydrobiologia 553:67–88
Olvera-Ramirez R, Centeno-Ramos C, Martinez Jeronimo F (2010) Toxic effects of Pseudanabaena tenuis (Cyanobacteria) on the cladocerans Daphnia magna and Ceriodaphnia dubia. Hidrobiologica 20:203–212
Oscoz J, Tomas P, Duran C (2010) Review and new records of non-indigenous freshwater invertebrates in the ebro river basin (Northeast Spain). Aquat Invasions 5:263–284
Paerl HW, Fulton RS III, Moidander PH, Dyble J (2001) Harmful freshwater algal blooms, with an emphasis on cyanobacteria. Sci World J 1:76–113
Peckham SD, Chipman JW, Lillesand TM, Dodson SI (2006) Alternate stable states and the shape of the lake trophic distribution. Hydrobiologia 571:401–407
Pedusaar T, Sammalkorpi I, Hautala A, Salujõe J, Järvalt A, Pihlak M (2010) Shifts in water quality in a drinking water reservoir during and after the removal of cyprinids. Hydrobiologia 649:95–106
Peretyatko A, Teissier S, De Backer S, Triest L (2009) Restoration potential of biomanipulation for eutrophic peri-urban ponds: the role of zooplankton size and submerged macrophyte cover. Hydrobiologia 634:125–135
Peretyatko A, Teissier S, De Backer S, Triest L (2010) Assessment of the risk of cyanobacterial bloom occurrence in urban ponds: probabilistic approach. Ann Limnol—Int J Limnol 146:121–133
Peretyatko A, Teissier S, De Backer S, Triest L (2012a) Biomanipulation of hypereutrophic ponds: when it works and why it fails. Environ Monit Assess 184:1517–1531
Peretyatko A, Teissier S, De Backer S, Triest L (2012b) Classification trees as a tool for predicting cyanobacterial blooms. Hydrobiologia 689:131–146
Pinto L, Chandrasena N, Pera J, Hawkins P, Eccles D, Sim R (2005) Managing invasive carp (Cyprinus carpio L.) for habitat enhancement at Botany Wetlands, Australia. Aquatic Conserv: Mar Freshw Ecosyst 15:447–462
Pot R, ter Heerdt GNJ (2014) Succession dynamics of aquatic lake vegetation after restoration measures: increased stability after 6 years of development. Hydrobiologia 737:333–345
Potthoff AJ, Herwig BR, Hanson M, Zimmer KD, Butler MG, Reed JR, Parsons BG, Ward MC (2008) Cascading food-web effects of piscivore introductions in shallow lakes. J Appl Ecol 45:1170–1179
Prokopkin IG, Gubanov VG, Gladyshev MI (2006) Modelling the effect of planktivorous fish removal in a reservoir on the biomass of cyanobacteria. Ecol Model 190:419–431
Raikow DF, Sarnelle O, Wilson AE, Hamilton SK (2004) Dominance of the noxious cyanobacterium Microcystis aeruginosa in low-nutrient lakes is associated with exotic zebra mussels. Limnol Oceanogr 49(2):482–487
Reinertsen H, Jensen A, Koksvik J, Langeland A, Olsen Y (1990) Effects of fish removal on the limnetic ecosystem of a eutrophic lake. Can J Fish Aquat Sci 47:166–173
Romo S, van Donk E, Gylstra R, Gulati R (1996) A multivariate analysis of phytoplankton and food web changes in a shallow biomanipulated lake. Freshw Biol 36:683–696
Rowland SJ, Mifsud C, Nixon M, Boyd P (2006) Effects of stocking density on the performance of the Australian freshwater silver perch (Bidyanus bidyanus) in cages. Aquaculture 253:301–308
Sarnelle O (2007) Initial conditions mediate the interaction between Daphnia and bloom-forming Cyanobacteria. Limnol Oceanogr 52:2120–2127
Scharf W (2007) Biomanipulation as a useful water quality management tool in deep stratifying reservoirs. Hydrobiologia 583:21–42
Scheffer M, Hosper SH, Meijer M-L, Moss B, Jeppesen E (1993) Alternative equilibria in shallow lakes. Trends Ecol Evol 8:275–279
Scheffer M, Carpenter S, Foley JA, Folke C, Walker B (2001) Catastrophic shifts in ecosystems. Nature 413:591–596
Seda J, Hejzlar J, Kubecka J (2000) Trophic structure of nine Czech reservoirs regularly stocked with piscivorous fish. Hydrobiologia 429:141–149
Shapiro J (1973) Blue-green algae—why they become dominant. Science 179(4071):382–384
Shapiro J (1997) The role of carbon dioxide in the initiation and maintenance of blue-green dominance in lakes. Freshw Biol 37:307–323
Shapiro J, Lamarra V, Lynch M (1975) Biomanipulation: an ecosystem approach to lake restoration. In: Brezonik PL, Fox JL (eds) Proceedings of a symposium on water quality management through biological control. University of Florida, Gainesville, pp 85–96
Shapiro J, Wright D (1984) Lake restoration by biomanipulations, round lake, Minnesota- the first two years. Freshw Biol 14:371–383
Sierp MT, Qin JG, Recknagel F (2009) Biomanipulation: a review of biological control measures in eutrophic waters and the potential for Murray cod Maccullochella peelii peelii to promote water quality in temperate Australia. Rev Fish Biol Fish 19:143–165
Skov C, Nilsson PA (2007) Evaluating stocking of YOY pike Esox lucius as a tool in the restoration of shallow lakes. Freshw Biol 52:1834–1845
Skov C, Perrow MP, Berg S, Skovgaard H (2002) Changes in the fish community and water quality during 7 years of stocking piscivorous fish in a shallow lake. Freshw Biol 47:2388–2400
Smart RM, Dick GO, Doyle RD (1998) Techniques for establishing native aquatic plants. J Aquat Plant Manag 36:44–49
Søndergaard M, Jeppesen E, Berg S (1997) Pike (Esox lucius L.) stocking as a biomanipulation tool 2. Effects on lower trophic levels in Lake Lyng, Denmark. Hydrobiologia 342(343):319–325
Søndergaard M, Jeppesen E, Jensen JP, Lauridsen T (2000) Lake restoration in Denmark. Lakes Reserv: Res Manag 5:151–159
Søndergaard M, Jeppesen E, Lauridsen TL, Skov C, Van Nes EH, Roijackers R, Lammens E, Portielje R (2007) Lake restoration in Denmark and the Netherlands: successes, failures and long-term effects. J Appl Ecol 44:1095–1105
Søndergaard M, Liboriussen L, Pedersen AR, Jeppesen E (2008) Lake restoration by fish removal: short- and long-term effects in 36 Danish lakes. Ecosystems 11:1291–1305
Sousa R, Gutiérrez JL, Aldridge DC (2009) Non-indigenous invasive bivalves as ecosystem engineers. Biol Invasions 11:2367–2385
Sousa R, Pilotto F, Aldridge DC (2011) Fouling of European freshwater bivalves (Unionidae) by the invasive zebra mussel (Dreissena polymorpha). Freshw Biol 56:867–876
Spataru P, Gophen M (1985) Feeding behavior of silver carp Hypophthalmichthys molitrix VAL. and its impact on the food web in Lake Kinneret, Israel. Hydrobiologia 120:53–61
Ståhl-Delbanco A, Hansson L, Gyllström M (2003) Recruitment of resting stages may induce blooms of Microcystis at low N:P ratios. J Plankton Res 25:1099–1106
Starling F (1998) Development of biomanipulation strategies for the remediation of eutrophication problems in a urban reservoir, Lago Paranoá—Brazil. Ph.D. Thesis. Institute of Aquaculture, University of Stirling, Scotland, p 259
Strayer DL (2009) Twenty years of zebra mussels: lessons from the mollusk that made headlines. Front Ecol Environ 7:135–141
Tang H, Xie P, Lu M, Xie L, Wang J (2002) Studies on the effects of silver carp (Hypophthalmichthys molitrix) on the phytoplankton in a shallow hypereutrophic lake through an enclosure experiment. Int Rev Hydrobiol 87:107–119
Tátrai I, Mátyás K, Korponai J, Paulovitz G, Pomogyi P, Héri J (2003) Regulation of plankton by omnivore cyprinids in a shallow lake in the Kis-Balaton Reservoir system. Hydrobiologia 504:241–250
Teissier S, Peretyatko A, De Backer S, Triest L. (2011) Contrôle des blooms cyanobactériens par manipulation de la biodisponibilité du carbone: étude en mésocosmes. Report Brussels Environmental Institute (DNEF/SDU/BIOL2009), Brussels, p 58
Teissier S, Peretyatko A, De Backer S, Triest L (2012) Strength of phytoplankton—nutrient relationship: evidence from 13 biomanipulated ponds. Hydrobiologia 689:147–159
Ter Heerdt G, Hootsmans M (2007) Why biomanipulation can be effective in peaty lakes. Hydrobiologia 584:305–316
Van de Bund WJ, van Donk E (2002) Short-term and long-term effects of zooplanktivorous fish removal in a shallow lake: a synthesis of 15 years of data from Lake Zwemlust. Freshw Biol 47:2380–2387
Van den Berg MS, Coops H, Meijer M-L, Scheffer M, Simons J (1998) Clear water associated with a dense Chara vegetation in the shallow and turbid lake Veluwemeer, The Netherlands. In: Jeppesen E, Søndergaard M, Søndergaard M, Christoffersen K (eds) The structuring role of submerged macrophytes in lakes. Ecological studies. Springer, New York, pp 339–352
Van den Berg MS, Coops H, Simons J (2001) Propagule bank buildup of Chara aspera and its significance for colonization of a shallow lake. Hydrobiologia 462:9–17
Van Donk E, van de Bund WJ (2002) Impact of submerged macrophytes including charophytes on phyto- and zooplankton communities: allelopathy versus other mechanisms. Aquat Bot 72:261–274
Van Wichelen J, Declerck S, Muylaert K, Hoste I, Geenens V, Vandekerkhove J, Michels E, De Pauw N, Hoffmann M, De Meester L, Vyverman W (2007) The importance of drawdown and sediment removal for the restoration of the eutrophied shallow Lake Kraenepoel (Belgium). Hydrobiologia 584:291–303
Vilà M, Basnou C, Pyšek P, Genovesi P, Gollasch S, Nentwig W, Olenin S, Roques A, Roy D, Hulme PE, DAISIE partners (2010) How well do we understand the impacts of alien species on ecosystem services? A pan-European, cross-taxa assessment. Front Ecol Environ 8:135–144
Wang H, Zhong G, Yan H, Liu H, Wang Y, Zhang C (2012) Growth control of cyanobacteria by three submerged macrophytes. Environ Eng Sci 29:420–425
Wilson DS, Turelli M (1986) Stable underdominance and the evolutionary invasion of empty niches. Am Nat 127:835–850
Xiao L, Ouyang H, Li H, Chen M, Lin Q, Han BP (2010) Enclosure study on phytoplankton response to stocking of silver carp (Hypophthalmichthys molitrix) in a eutrophic tropical reservoir in South China. Int Rev Hydrobiol 95:428–439
Xie P, Liu J (2001) Practical success of biomanipulation using filter-feeding fish to control cyanobacteria blooms. Sci World J 1:337–356
Zeng Q, Gu X, Mao Z, Chen X (2014) In situ growth and photosynthetic activity of Cyanobacteria and phytoplankton dynamics after passage through the gut of silver carp (Hypophthalmichthys molitrix), bighead carp (Aristichthys nobilis), and Nile tilapia (Oreochromis niloticus). Hydrobiologia 736:51–60
Zhang X, Xie P, Huang X (2008) A review of nontraditional biomanipulation. Sci World J 8:1184–1196
Zimmer K, Hanson M, Butler G (2003) Relationships among nutrients, phytoplankton, macrophytes, and fish in prairie wetlands. Can J Fish Aquat Sci 60:721–730
Zimmer K, Hanson MA, Herwig BR, Konsti ML (2009) Thresholds and stability of alternative regimes in Shallow Prairie-Parkland Lakes of Central North America. Ecosystems 12:843–852
Acknowledgments
Financial support was obtained from the Vrije Universiteit Brussel (BAS42), the Flemish Interuniversity Council for Development and Cooperation (VLIR-UOS, ICP Biology: Human Ecology) and the Brussels Institute for Environment (Brussels pond and river projects 2006–2015).
Author information
Authors and Affiliations
Corresponding author
Additional information
Guest editors: Petra M. Visser, Bas W. Ibelings, Jutta Fastner & Myriam Bormans/Cyanobacterial blooms. Ecology, prevention, mitigation and control.
Ludwig Triest, Iris Stiers, Stijn Van Onsem equally contributed as first author.
Rights and permissions
About this article

Cite this article
Triest, L., Stiers, I. & Van Onsem, S. Biomanipulation as a nature-based solution to reduce cyanobacterial blooms. Aquat Ecol 50, 461–483 (2016). https://doi.org/10.1007/s10452-015-9548-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10452-015-9548-x