
Abstract
Numerous studies have demonstrated alternative regimes in shallow lake ecosystems around the world, with one state dominated by submerged macrophytes and the other by phytoplankton. However, the stability of each regime, and thresholds at which lakes shift to the alternative regime, are poorly known. We used a cross-sectional analysis of 72 shallow lakes located in prairie and parkland areas of Minnesota, USA, during 2005 and 2006 to assess the occurrence of alternative regimes and shifts between them. Cluster analysis revealed two distinct groups of lakes characterized not only by different macrophyte abundance and chlorophyll a levels but also by different total phosphorus–chlorophyll a relationships. Thirty-nine lakes were macrophyte- and 23 lakes phytoplankton-dominated in both years, whereas 10 sites shifted sharply between those regimes. We failed to detect a universal shifting threshold in terms of chlorophyll a or total phosphorus. However, 95% of the lakes with chlorophyll a concentrations less than 22 μg l−1 were in a clear-water regime, whereas 95% of the lakes with chlorophyll a higher than 31 μg l−1 were in a turbid regime. Total phosphorus less than 62 μg l−1 was an accurate predictor of lakes in a stable clear-water regime, whereas a large change in biomass of planktivores and benthivores between years was the only variable weakly related to regime shifts. Our results support the theoretical prediction that regime thresholds vary among lakes. We recommend that lake managers focus on improving resilience of clear regimes in shallow lakes by reducing nutrient loading, rather than attempting to identify and manage complex triggers of regime shifts.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Alternative regimes have been documented in ecosystems as diverse as deserts, woodlands, and coral reefs (Scheffer and others 2001). Shallow lakes were one of the first systems for which alternative regimes were described, with one regime consisting of clear water dominated by submerged macrophytes, and the other turbid water dominated by phytoplankton (Scheffer and others 1993). Recent work has expanded the concept for shallow lakes to include regimes dominated by floating-leaf macrophytes and specific types of phytoplankton (Scheffer and van Nes 2007), but most shallow lake research continues to focus on the dichotomy between submerged macrophytes (hereafter macrophytes) and phytoplankton.
Key areas of research on shallow lakes include perturbations inducing shifts from one regime to the other (Hanson and Butler 1994a), influence of ambient nutrient levels on regime stability and frequency of occurrence (Bayley and Prather 2003), and stability of regimes through historic time periods (Karst and Smol 2004). Many recent studies on shallow lakes have used longitudinal approaches, studying single lakes through time (sensu Skov and others 2002). Additionally, most of these efforts focused on stability and regime shifts in response to human-induced perturbation such as biomanipulation (sensu Hansel-Welch and others 2003). These are powerful approaches and have shed considerable insight on shallow lake dynamics, but they do not assess mechanisms responsible for natural regime shifts, or the frequency of natural shifts at a landscape scale. Natural shifting mechanisms could include changes in lake depth (Blindow and others 1993), waterfowl grazing on macrophytes (van Donk and Gulati 1995), and changes in fish abundance via winterkill or fish colonization (Zimmer and others 2001). Thus, research on present-day regime shifts under natural conditions across multiple lakes would complement work done on individual lakes and on human-induced shifts.
There is also limited information on “thresholds,” points of macrophyte and phytoplankton abundance that, when crossed, induce shifts to the alternative regime. Competitive interactions between macrophytes and phytoplankton for light and nutrients are believed to be core mechanisms of alternative regimes in shallow lakes (reviewed by Scheffer 2001). These interactions, along with positive feedback mechanisms, lead to opposite attractors, pulling lakes toward dominance by either macrophytes or phytoplankton. Thus, at moderate lake depth, high abundance of both macrophytes and phytoplankton is believed to be unstable, and once one decreases below an abundance threshold, the other will increase. This is a simplistic view, as the macrophyte–phytoplankton relationship will also depend on nonalgal sources of turbidity (Jackson 2003), ambient nutrients (Bayley and others 2007), periphyton (Jones and Sayer 2003), water depth (Bayley and Prather 2003), and floating-leaf macrophytes, along with other lake properties (Scheffer and van Nes 2007). Given the complex interactions among these factors, it is unclear whether specific thresholds for the phytoplankton–macrophyte interaction can be estimated, but even approximate threshold points or ranges remain poorly defined.
One approach to estimating phytoplankton–macrophyte thresholds under natural perturbations would be to study a single or limited number of lakes through time. Shortcomings of this approach include low power to assess mechanisms inducing shifts and limited ability to measure variance in thresholds among lakes. Alternatively, one could use a “cross-sectional” approach, where many lakes are studied during a shorter time period. This strategy addresses shortcomings of the longitudinal study, but is less useful for assessing temporal variability within individual lakes.
Here we use a cross-sectional analysis of 72 shallow lakes sampled for two consecutive years in north-central North America to address the following objectives. First, characterize lakes as turbid, clear, or shifting based on concentrations of chlorophyll a and abundance of macrophytes. Second, assess the influence of ambient nutrient levels on the proportion of lakes in turbid, clear, and shifting regimes. Third, determine if we can estimate an approximate threshold of phytoplankton abundance associated with regime shifts. Finally, attempt to identify variables associated with stable clear-water regimes, stable turbid-water regimes, or shifting regimes between years.
Methods
Our study was conducted along the eastern margin of the Prairie Pothole Region (PPR) of North America (Minnesota, USA). The PPR is a grassland landscape stretching from Alberta to Iowa, covers approximately 715,000 km2, and is dotted with thousands of shallow lakes and wetlands (Euliss and others 1999). Previous work in this region has shown alternative regimes in shallow water bodies ranging from 0.03 to 16 km2 in size, and revealed that fish can play major roles in inducing shifts between regimes (Hanson and Butler 1994a; Zimmer and others 2003). Fish presence, abundance, and community composition are highly variable both spatially and temporally (Zimmer and others 2000, 2001). These dynamic fish communities are driven by harsh abiotic conditions and winter hypoxia (Peterka 1989) and variable colonization rates resulting from climate variability and anthropogenic alteration of the landscape via ditches and culverts (Hanson and others 2005).
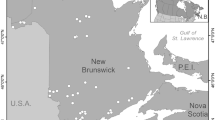
We selected study lakes from two areas of Minnesota, one in the PPR (hereafter “prairie lakes”) and the other in the PPR–deciduous forest transition area (hereafter “parkland lakes”) (Figure 1). Our study sites were dispersed across 1,292 km2 in the parkland and 1,435 km2 in the prairie. We studied lakes in both areas because ambient nitrogen (N) and phosphorus (P) levels are generally higher in lakes in the prairie (Heiskary and others 1987; Table 1).

Location of the study areas containing the parkland and prairie lakes in Minnesota, USA.
We selected 72 study sites by identifying all Type IV (semipermanent hydroperiod) and V wetlands (permanent hydroperiod) (following the classification scheme of Stewart and Kantrud [1971]) in each area using the Minnesota Department of Natural Resources’ National Wetlands Inventory (NWI) GIS database. Our sites are referenced as wetlands in the NWI database, but here we refer to them as shallow lakes. We stratified sites in each study area by assignment to 27 bins based on combinations of the following features: (1) lake size (small, medium, large, range 0.02–0.5 km2), (2) distance to nearest permanent stream, wetland, or lake (short, medium, long, range 0–1,825 m), and (3) percent agriculture within a 500 m buffer surrounding the lake (small, medium, large, range 0–97%). We then randomly selected one study site from each of the resulting bins in each area, plus 8 additional parkland lakes and 10 additional prairie lakes, with a maximum of two lakes per bin. If we were unable to obtain access to a lake, we randomly selected a new site from that bin.
Aerial photographs (2003 Farm Service Agency color digital orthophoto quadrangles) and GIS (ArcView 3.3 and ArcGIS 9.2, Environmental Systems Research Institute Inc. 2007) were used to estimate surface area of each lake in 2005. Maximum depth of the study sites was also determined during the 2005 field season by measuring depths along parallel transects through the open water zone.
Lakes were sampled for 2 years (2005 and 2006), and fish species composition and relative abundances were estimated in July of each year using two types of gear. Three mini-fyke nets (6.5 mm bar mesh with 4 hoops, 1 throat, 7.62 m lead, and a 0.69 m × 0.99 m rectangular frame opening into the trap) were set overnight in the littoral zone of each lake. One experimental gill net (61.0 m multifilament net with 19, 25, 32, 38, and 51-mm bar meshes) was set overnight along the maximum depth contour in lakes less than 2 m deep, or along a 2-m contour in lakes with sufficient depth. These methods have been shown to be effective in sampling fish assemblages in small lakes from other regions (Tonn and Magnuson 1982; Jackson and Harvey 1989; Robinson and Tonn 1989), and enabled us to capture fish of different sizes, species, and from all major trophic guilds. We summed the biomass of each species captured across the gill net and three fyke nets set in each site in each year. We then determined the total mass of planktivore and benthivore trophic guilds for each lake by summing mass of all benthivorous and planktivorous fish captured. Fish species were classified as benthivores or planktivores based on Becker (1983) and Verant and others (2007).
Chlorophyll a was used as an index of phytoplankton abundance, and samples were collected concurrently with fish in July. Three water samples were taken from the open-water area of each lake, placed on ice, filtered through GF/F filters within 8 h of collection, and immediately frozen. Chlorophyll a was determined via alkaline acetone extraction and fluorometry. We used the average chlorophyll a value across the three samples for each lake for each year. Total nitrogen (TN) and total phosphorus (TP) were estimated each year from a water sample collected concurrently with chlorophyll a. Samples were placed on ice, frozen, and later analyzed using procedures described in APHA (1994). Abundance of submerged aquatic macrophytes was assessed in early August (within 1–2 weeks of measuring chlorophyll a) using methods modified from Deppe and Lathrop (1992). Macrophytes were sampled in each lake at 20 stations located equidistant along four transects running the width of the lake. We made one throw of a weighted plant rake at each station, and dragged the rake along 3 m of lake bottom. We then determined total mass of macrophytes collected on each throw, and used average mass across the 20 throws for all of our analyses. Chlorophyll a and TP data were log10 transformed for our linear regression to normalize residuals and increase homogeneity of variance. Physical and biological features of our study sites are summarized in Table 1.
Our first objective was to characterize lakes as turbid, clear, or shifting regimes based on concentrations of chlorophyll a and abundance of macrophytes, and to estimate the proportion of lakes shifting regimes under natural conditions. We used k-means cluster analysis (McCune and Grace 2002) to group lakes in each year into the two most homogeneous groups possible based on abundance of macrophytes and chlorophyll a concentrations, with the two groups comprising lakes in turbid- and clear-water regimes (sensu Jackson 2003; Bayley and others 2007). Data for 2005 and 2006 were analyzed separately, allowing us to classify each lake as a stable clear-water regime both years, a stable turbid regime both years, or shifting regimes between years (those classified turbid one year and clear the other).
We then assessed whether our two groups identified with cluster analysis might represent alternative regimes. Theory predicts that lakes exhibiting alternative regimes should display dual relationships to a control factor (Scheffer and Carpenter 2003). Thus, we regressed chlorophyll a concentrations on the controlling factor TP for both years of data. We then used likelihood ratio tests to determine the most parsimonious model for the TP–chlorophyll a relationship (Genmod procedure of SAS 9.1, SAS Institute Inc. 2003). Variance explained typically increases as models become more complex with greater numbers of parameters. However, the likelihood ratio test accounts for this in assessing whether the more complex among competing, nested models provide significantly better fit relative to simpler models with fewer parameters (Hilborn and Mangel 1997). Our competing models in order of increasing complexity were (1) a common slope and intercept for both turbid and clear lakes, (2) a common slope but separate intercepts for clear and turbid lakes, and (3) separate slopes and intercepts for clear and turbid lakes. A dualistic relationship (models 2 or 3) between TP and chlorophyll a is not proof of alternative regimes in our lakes, but does support their existence (Scheffer and Carpenter 2003).
Our second objective was to test whether ambient nutrient levels influenced the proportion of clear, turbid, and shifting lakes. TN and TP concentrations were substantially higher in the prairie lakes compared to parkland lakes during the course of this study (Table 1). We used Fisher exact tests to assess whether the proportion of lakes clear, turbid, and shifting differed between prairie and parkland study areas. We also hypothesized that the probability of lakes being clear, shifting, or turbid would show a significant relationship with TP, regardless of study area. Theory predicts that the probability of lakes being clear, shifting, or turbid are highest at low, moderate, and high levels of TP, respectively (Moss and others 1997; Scheffer 1998). Thus, we used ordinal logistic regression (O’Connell 2006) to test whether TP concentrations in 2005 had a significant relationship with the probability of lakes being clear both years, shifting regimes, or being turbid both years (Logistic procedure of SAS 9.1, SAS Institute Inc. 2003).
Our third objective was to estimate thresholds of phytoplankton abundance between turbid- and clear-water regimes. This was done by examining the proportion of lakes classified as turbid and clear along the observed gradient of chlorophyll a concentrations. An abrupt shift in proportions of clear and shifting lakes would indicate a somewhat universal threshold between regimes across lakes, whereas a more gradual change in proportions of regimes would indicate that thresholds vary among individual lakes. We focused our examination of thresholds on phytoplankton abundance because chlorophyll a is widely used to estimate algal abundance, making our results widely applicable. Our methods for macrophyte mass were less standard, thus reducing their comparability to other studies.
Our fourth objective was to identify variables associated with stable clear- and turbid-water regimes, and lakes shifting regimes between years. We used classification and regression trees (CART) (Breiman and others 1984; Urban 2002) to construct predictive models for these three types of lakes. CART is a recursive technique that partitions data into homogeneous subsets based on values of predictive variables. In our case, CART uses predictor variables to partition lakes into categories of clear, turbid, and shifting status. Our predictor variables included maximum lake depth, lake surface area, study area (prairie vs. parkland), TP values in 2005, mean TP between years, change in TP between years, proportional change in TP between years, average biomass of planktivores (P), benthivores (B), and planktivores + benthivores (P + B) between years, change in biomass of P, B, and P + B between years, and proportional change in biomass of P, B, and P + B between years. Proportional change in both fish mass and TP was defined as the change between years divided by the highest value observed in the two years of data. As described above, we assessed the relationship between TP and lake regime using ordinal logistic regression, but we also included TP in our CART analysis because this technique allows us to compare predictive ability of TP relative to other predictor variables.
We developed our CART model using a training data set comprising 70% of our study sites, with the remaining 30% reserved for model validation. Because we were attempting to predict three types of lakes, our model required a minimum of two partitions of the data. We minimized over fitting and determined optimum model size in the training data set by selecting the number of partitions that yielded the smallest error rate in k-fold cross-validation (k = 25) (De’ath and Fabricius 2000). Generality of the optimal training-set model was evaluated by testing its ability to correctly predict turbid, clear, and shifting lakes in the validation data set. We used JMP (SAS Institute Inc. 2002) to perform our CART analysis.
Results
Cluster analysis identified two homogenous groups of lakes characterized by large differences in the abundance of both macrophytes and phytoplankton between groups (Figure 2). These results were then used to classify each lake as in a stable clear-water regime, a stable turbid-water regime, or shifting regimes between years. Lakes classified as clear both years averaged 9 μg l−1 chlorophyll a and 1,360 g sample−1 of macrophytes, compared to 81 μg l−1 chlorophyll a and 91 g sample−1 of macrophytes in lakes classified as turbid both years. Lakes classified as shifting regimes changed sharply between years, averaging 15 μg l−1 chlorophyll a and 1,061 g sample−1 macrophytes while in the clear regime, and 93 μg l−1 chlorophyll a and 165 g sample−1 of macrophytes in the turbid regime. Thus, shifting lakes exhibited phytoplankton and macrophyte abundance similar to clear lakes in one year and turbid lakes the other year. We did not find a universal threshold level of chlorophyll a that separated clear from turbid lakes. However, across both years of the study, 95% of the lakes with chlorophyll a less than 22 g l−1 were classified as clear, whereas 95% of the lakes with chlorophyll a greater than 31 μg l−1 were classified as turbid (Figure 2).

Concentrations of chlorophyll a and macrophyte biomass in lakes classified as clear and turbid in 2005 (top) and 2006 (bottom). Classification of lakes was based on k-means cluster analysis. The dashed vertical lines represent 22 and 31 μg l−1 chlorophyll a. Across both years of the study, 95% of lakes below 22 μg l−1 were classified as clear and 95% of lakes above 31 μg l−1 were classified as turbid.
Our results indicated that the alternative regimes identified with cluster analysis exhibited dualistic relationships with TP, as the optimal model describing the chlorophyll a and TP relationship included a common slope but separate intercepts for turbid and clear lakes (Figure 3). Log likelihood ratio tests indicated that the most complex model, fitting separate slopes and separate intercepts (2005: r 2 = 0.81, 2006: r 2 = 0.76), was not a significant improvement (2005: P = 0.872, 2006: P = 0.553) over the moderate complexity model fitting separate intercepts and a common slope (2005: r 2 = 0.81, 2006: r 2 = 0.76). However, the moderate complexity model was a significant improvement (2005: P < 0.001, 2006: P < 0.001) over the simplest model fitting a common slope and intercept (2005: r 2 = 0.65, 2006: r 2 = 0.64). This dualistic relationship supports that cluster analysis classified lakes into clear versus turbid alternative regimes, and Figure 3 suggests either regime was possible when chlorophyll a fell between approximately 13 and 38 μg l−1. Similarly, either regime was possible over a TP range of approximately 40–500 μg l−1, although one lake was classified as turbid with TP levels of just 16 μg l−1.

Relationship between concentrations of TP and chlorophyll a in turbid and clear lakes in 2005 (top) and 2006 (bottom). Log likelihood ratio tests indicated the most parsimonious model in both years consisted of separate intercepts but the same slope for turbid and clear lakes.
Fifty-four percent of the lakes were clear both years, 32% were turbid, and 14% shifted regimes between years. Of the 10 lakes shifting regimes, 4 lakes shifted clear to turbid from 2005 to 2006, whereas 6 switched from turbid to clear. The proportion of lakes in stable clear regimes was significantly higher in low-nutrient parkland lakes (80%) compared to high-nutrient prairie lakes (30%) (P < 0.001), and the proportion of stable turbid lakes was lower in parkland lakes (11%) relative to prairie lakes (51%) (P < 0.001). However, no difference was observed in the proportion of lakes shifting regimes (parkland 9%, prairie 19%, P = 0.309). Ordinal logistic regression indicated a significant relationship between TP in 2005 and the probability that lakes remained clear both years, shifted regimes between years, or remained turbid both years (P < 0.001). Results supported theoretical predictions for shallow lakes, as the probabilities of being clear both years, shifting between years, and being turbid both years were highest at low, moderate, and high levels of TP, respectively (Figure 4). A score test for the proportional odds assumption was not significant (P < 0.245), supporting our use of ordinal regression on these data.

Estimated relationship between TP concentrations in 2005 and the probability of lakes being clear in both 2005 and 2006, shifting regimes between years, or being turbid in both years. Estimates are from ordinal logistic regression.
Results for our CART analysis on the training data set indicated an optimal model with two predictor variables (Figure 5). The optimal model predicted lakes with TP less than 61.5 μg l−1 would be in a clear-water regime. Lakes with TP greater than 61.5 μg l−1 were further partitioned based on proportional change in summed biomass of planktivores and benthivores, with lakes having greater than 67% change in fish mass predicted to be shifting regimes, whereas lakes with less than 67% change in fish mass were predicted to be turbid. For the training data set, the model correctly predicted 89% of the clear lakes and 94% of the turbid lakes, but only 57% of shifting lakes (Table 2). The correct prediction percentage was also high for predicting clear lakes in the validation data set (82% correct), but dropped substantially for turbid (50% correct) and shifting (33% correct) lakes. Thus, the CART model showed reasonable accuracy for predicting clear regimes based on TP, but predicting turbid versus shifting regimes using change in fish biomass was less accurate. We captured 28 species of fish, but black bullhead (Ameiurus melas) and common carp (Cyprinus carpio) represented 79 and 15% of total benthivore mass captured, respectively. Planktivores were dominated by fathead minnow (Pimephales promelas) and yellow perch (Perca flavescens), accounting for 53 and 10% of total planktivore mass, respectively.

Classification tree predicting lake regime (clear both years, turbid both years, or shifting regimes between years) based on CART analysis of the 52 lakes in the training data set during 2005 and 2006. The analysis assessed 13 potential predictor variables, but this optimal model was based on mean concentration of TP and the proportional change in summed biomass of planktivores and benthivores between years. The resulting model predicts a clear-water regime in lakes with low TP, shifting regimes in lakes with high phosphorus and large change in fish biomass, and a turbid regime in lakes with high phosphorus and small change in fish biomass. The splitting value for the predictive variable and the number of lakes corresponding to each split are given for each “branch” of the tree, and percentages in the terminal nodes (boxes) indicate the response rate for each type of lake.
Discussion
Our results indicate shallow prairie and parkland lakes exhibit alternative regimes. However, the proportion of clear and turbid sites differs between the two study areas, reflecting low and high ambient nutrient levels in parkland and prairie sites, respectively. Our results also suggest lakes may shift regimes over a relatively wide range of phytoplankton abundance, as opposed to a distinct, universal threshold marking a transition point from one regime to the other. We also observed a strong relationship between low TP levels and clear-water regimes, but failed to find strong predictors of regime shifts. Variable thresholds of phytoplankton abundance, coupled with unpredictable triggers for regime shifts, supports the suggestion that lake managers should focus on increasing resilience of clear-water regimes, as opposed to trying to identify and manage complex triggers of regime shifts (Scheffer and others 2001).
Our cross-sectional study did not assess whether lakes remain stable during multiple years. It is plausible that shifting lakes are inherently unstable and phytoplankton and macrophyte abundance in these systems fluctuates dramatically most years. However, ancillary data collected in 2004 and 2007 from some of our shifting lakes do not support yearly shifts. Chlorophyll a concentrations (μg l−1) from 2004 through 2006 were 186, 158, and 27 in the first lake, 13, 20, and 149 in the second, and 8, 28, and 178 in the third, whereas a fourth lake was 289, 243, 3, 7, and 3 μg l−1 during 2004–2008. Additionally, Zimmer and others (2003) observed no regime shifts in 19 PPR shallow lakes during five consecutive years of study. These data suggest that shifting patterns observed between 2005 and 2006 were distinct shifts and not part of high interannual dynamics, as observed for shallow lakes of boreal Canada (Bayley and others 2007).
Our results indicated the probability of lakes being turbid or clear was least sensitive to changes in chlorophyll a both below 22 μg l−1 and above 31 μg l−1, with the former identifying levels where 95% of the lakes were clear and the latter levels where 95% of lakes were turbid. Between 22 and 31 μg l−1 chlorophyll a, the two regimes occurred with similar frequency. This indicates phytoplankton thresholds between regimes for most lakes fell within the 22–31 μg l−1 range, and this range suggests variability among individual lakes. Depth may be an important source of variance in threshold values, as it has a strong influence on the relationships among nutrients, phytoplankton abundance, and macrophyte abundance (Scheffer and van Nes 2007). Moreover, theoretical models suggest that forward and backward shifts between regimes in individual lakes may occur at different levels of water clarity (Scheffer 1998). Although we found no absolute threshold, our results imply that lake managers may maximize the probability of stable clear regimes by maintaining chlorophyll a levels below 22 μg l−1. Interestingly, Bayley and others (2007) estimated an 18 μg l−1 chlorophyll a threshold between regimes for shallow lakes in boreal regions of Alberta, Canada, close to the 22–31 μg l−1 range observed here.
Shallow lake theory predicts that the stability of clear and turbid regimes should be inversely related, and the stability of each is influenced by nutrient concentrations (reviewed by Scheffer 1998). Our results support these predictions, as the probability of being clear was negatively related to TP, the probability of shifting regimes peaked at moderate TP levels, whereas the probability of being turbid was positively related to TP. The importance of ambient nutrient levels for the occurrence of turbid and clear regimes can also be seen at the broader scale comparing nutrient-rich prairie lakes to nutrient-poor lakes of the parkland region. Given the relationship between nutrients and regime stability, one would expect a higher proportion of turbid lakes in areas with higher ambient nutrient levels. Our data support this hypothesis, as the turbid regime occurred more often in the high-nutrient prairie lakes, whereas the clear regime occurred more often in the low-nutrient parkland lakes. Minnesota has a natural north–south gradient of ambient nutrient levels, with median TP concentrations in the northern “lakes and forest” ecoregion being 28 μg l−1, whereas the median in the more southern “northern glaciated plains” ecoregion is 177 μg l−1 (Heiskary and others 1987). We hypothesize a continuum of alternative regimes along this gradient, where occurrence of clear regimes peaks in northern lakes with low nutrients, shifting lakes peak in mid-latitude lakes with moderate nutrient levels, and turbid lakes peak in southern lakes with highest ambient nutrient levels.
Our CART analysis also detected TP as a strong predictor of clear regimes, but failed to find strong predictors for either turbid regimes or shifts in regimes between years. The CART model predicted clear-water regimes in lakes with average TP less than 62 μg l−1, similar to the approximately 50–100 μg l−1 estimate for shallow lakes in boreal Canada (Bayley and others 2007) and the 50 μg l−1 estimate for lakes larger than 3 ha in Denmark (Jeppesen and others 1990). Despite these similarities in the stability of the clear-water regime along P gradients, results for individual lakes vary due to influences of other factors such as N concentrations (Gonzáles Sagrario and others 2005) and lake size (Jeppesen and others 1990). Thus, although 62 μg l−1 may serve as an approximate management goal for maintenance of a clear-water regime, in this region lake managers should recognize that results will vary across lakes and they should strive for maintaining the lowest P levels possible. The concept of minimizing nutrient levels is supported by our logistic analysis, suggesting that probabilities of clear regimes continue to increase through the lowest concentrations of TP observed in our lakes.
CART also indicated that proportional change in biomass of planktivores and benthivores best distinguished shifting from turbid lakes to some extent. However, the complexity and stochastic nature of regime shifts likely precluded identification of any variable consistently associated with regime shifts. Regime shifts can be induced by disturbances or changes in the stability of lakes (Scheffer 1998). However, whether disturbance induces regime shifts also depends on the intrinsic stability and resilience of individual lakes (Scheffer and others 2001), further complicating any effort to identify a specific variable responsible for regime shifts in a given region. Such predictions for lakes across multiple ecoregions will be even more challenging. The inability of TP to distinguish shifting from turbid lakes in our CART analysis may seem surprising given the results of our logistic regression. However, average concentrations of TP were rather similar between shifting (171 μg l−1) and turbid lakes (205 μg l−1) relative to clear lakes (48 μg l−1).
Although a 67% change in biomass of planktivores and benthivores was our best predictor of regime shifts, the overall relationship was relatively weak. In part, the difficulty of finding factors related to shifts in our lakes might be due to the fact that forward and reverse shifts are likely induced by different variables (Moss and others 1997). On the other hand, the difficulty in finding relationships could in part be due to the fact that we had only a small group of shifting lakes for statistical analysis.
Previous work has shown that fish have strong impacts on shallow lakes of the PPR, and changes in fish abundance can induce shifts between regimes (Hanson and Butler 1994a; Zimmer and others 2001; Potthoff and others 2008). Accordingly, fish are often the focus of management efforts (Hanson and Butler 1994b). Although fish exert strong influences on shallow lakes, in our study we failed to find strong relationships between fish biomass and turbid, clear, and shifting lakes. Thus, managers should supplement management of fish populations with strategies that increase resilience of clear-water regimes, including strategies that reduce nutrient loading.
References
APHA. 1994. Standard methods for the examination of waste and wastewater. American Public Health Association, Washington, DC.
Bayley SE, Creed IF, Sass GZ, Wong AS. 2007. Frequent regime shifts in trophic states in shallow lakes of the Boreal Plain: alternative “unstable” states? Limnol Oceanogr 52:2002-12.
Bayley SE, Prather CM. 2003. Do wetland lakes exhibit alternative stable states? Submersed aquatic vegetation and chlorophyll in western boreal shallow lakes. Limnol Oceanogr 48:2335-45.
Becker GC. 1983. Fishes of Wisconsin. University of Wisconsin Press, Madison, Wisconsin
Blindow I, Andersson G, Hargeby A, Johansson S. 1993. Long-term patterns of alternative stable states in two shallow eutrophic lakes. Freshw Biol 30:159-67.
Breiman L, Friedman JH, Olshen RA, Stone CJ. 1984. Classification and regression trees. Chapman & Hall, New York.
De’ath G, Fabricius KE. 2000. Classification and regression tree: a powerful yet simple technique for ecological data analysis. Ecology 81:3178-92.
Deppe ER, Lathrop RC. 1992. A comparison of two rake sampling techniques for sampling aquatic macrophytes. Wisconsin Department of Natural Resources, Findings #32, PUBL-RS-732-92, Madison, Wisconsin
Environmental Systems Research Institute Inc. 2007. What’s new in ArcGIS 9.2. Environmental Systems Research Institute, Redlands, California.
Euliss NH, Jr., Wrubleski DA, Mushet DM. 1999. Wetlands of the prairie pothole region: invertebrate species composition, ecology, and management. In: Batzer, D.P., Rader, R.P., Wissinger, S.A. (Eds.), Invertebrates in freshwater wetlands of North America: ecology and management. John Wiley and Sons, New York, pp. 471-514.
Gonzáles Sagrario MA, Jeppesen E, Gomá J, Søndergaard M, Peder Jensen J, Lauridsen T, Landkildehus F. 2005. Does high nitrogen loading prevent clear-water conditions in shallow lakes at moderately high phosphorus concentrations? Freshw Biol 50:27-41.
Hansel-Welch N, Butler MG, Carlson TJ, Hanson MA. 2003. Changes in macrophyte community structure in Lake Christina (Minnesota), a large shallow lake, following biomanipulation. Aquat Bot 75:323–37.
Hanson MA, Butler MG. 1994a. Responses of plankton, turbidity, and macrophytes to biomanipulation in a shallow prairie lake. Can J Fish Aquat Sci 51:1180-8.
Hanson MA, Butler MG. 1994b. Responses to food web manipulation in a shallow waterfowl lake. Hydrobiologia 279-280:457-66.
Hanson MA, Zimmer KD, Butler MG, Tangen BA, Herwig BR, Euliss NH, Jr. 2005. Biotic interactions as determinants of ecosystem structure in prairie wetlands: an example using fish. Wetlands 25:764-75.
Heiskary SA, Wilson BC, Larsen DP. 1987. Analysis of regional patterns in lake water quality: using ecoregions for lake management in Minnesota. Lake Reserv Manage 3:337-44.
Hilborn R, Mangel M. 1997. The ecological detective: confronting models with data. Princeton University Press, Princeton, NJ.
Jackson LJ. 2003. Macrophyte-dominated and turbid states of shallow lakes: evidence from Alberta lakes. Ecosystems 6:213-23.
Jackson DA, Harvey HH. 1989. Biogeographic associations in fish assemblages: local vs. regional processes. Ecology 70:1472-84.
Jeppesen E, Jensen JP, Kristensen P, Sondergaard M, Mortensen E, Sortkjaer O, Olrik K. 1990. Fish manipulation as a lake restoration tool in shallow, eutrophic, temperate lakes 2: threshold levels, long-term stability, and conclusions. Hydrobiologia 200/201:219-27.
Jones JI, Sayer CD. 2003. Does the fish-invertebrate periphyton cascade precipitate plant loss in shallow lakes? Ecology 84:2155-67.
Karst TL, Smol JP. 2004. Paleolimnological evidence of limnetic nutrient concentration equilibrium in a shallow, macrophyte-dominated lake. Aquat Sci 62:20-38.
McCune B, Grace JB. 2002. Analysis of Ecological Communities. MJM Software Design, Gleneden Beach, Oregon.
Moss B, Madgwick J, Phillips G. 1997. A guide to the restoration of nutrient-enriched shallow lakes. W W Hawes, Norwich.
O’Connell AA. 2006. Logistic Regression Models for Ordinal Response Variables. Sage, Publications, Inc., Thousand Oaks, California.
Peterka JJ. 1989. Fishes of northern prairie wetlands. In: Van der Valk, A. (Ed.), Northern prairie wetland. Iowa State University Press, Ames, pp. 302-15.
Potthoff AJ, Herwig BR, Hanson MA, Zimmer KD, Butler MG, Reed JR, Parsons BG, Ward MC. 2008. Cascading food-web effects of piscivore introductions in shallow lakes. J Appl Ecol 45:1170-9.
Robinson CLK, Tonn WM. 1989. Influence of environmental factors and piscivory in structuring fish assemblages of small Alberta lakes. Can J Fish Aquat Sci 46:81-9.
SAS Institute Inc. 2002. JMP User’s Guide, Version 5. SAS Institute, Inc., Cary, North Carolina
SAS Institute Inc. 2003. SAS 9.1 help and documentation. SAS Institute Inc., Cary, North Carolina
Scheffer M. 1998. Ecology of shallow lakes. Kluwer Academic Publishers, Dordrecht, the Netherlands.
Scheffer M. 2001. Alternative attractors of shallow lakes. Sci World 1:254-63.
Scheffer M, Carpenter SR. 2003. Catastrophic shifts in ecosystems: linking theory to observation. Trends Ecol Evol 18:648-56.
Scheffer M, Carpenter SR, Foley JA, Folkes C, Walker B. 2001. Catastrophic shifts in ecosystems. Nature 413:591-6.
Scheffer M, Hosper SH, Meijer M-L, Moss B, Jeppesen E. 1993. Alternative equilibria in shallow lakes. Trends Ecol Evol 8:275-9.
Scheffer M, van Nes EH. 2007. Shallow lake theory revisited: various alternative regimes driven by climate, nutrients, depth and lake size. Hydrobiologia 584:455-66.
Skov C, Perrow MR, Berg S, Skovgaard H. 2002. Changes in the fish community and water quality during seven years of stocking piscivorous fish in a shallow lake. Freshw Biol 47:2388-400.
Stewart RE, Kantrud HA. 1971. Classification of natural ponds and lakes in the glaciated prairie region. Bureau of Sport Fisheries and Wildlife Resources Publication No. 92, Washington, D.C
Tonn WM, Magnuson JJ. 1982. Patterns in the species composition and richness of fish assemblages in northern Wisconsin lakes. Ecology 63:1149-66.
Urban DL. 2002. Classification and regression trees. In: McCune, B., Grace, J.B. (Eds.), Analysis of ecological communities. MjM Software Design, Gleneden Beach, Oregon, pp. 222-32.
van Donk E, Gulati RD. 1995. Transition of a lake to turbid state six years after biomanipulation: mechanisms and pathways. Water Sci Technol 32:197-206.
Verant ML, Konsti ML, Zimmer KD, Deans CA. 2007. Factors influencing nitrogen and phosphorous excretion rates of a fish community in a shallow lake. Freshw Biol 52:1968-81.
Zimmer KD, Hanson MA, Butler MG. 2000. Factors influencing invertebrate communities in prairie wetlands: a multivariate approach. Can J Fish Aquat Sci 57:76-85.
Zimmer KD, Hanson MA, Butler MG. 2001. Effects of fathead minnow colonization and removal on a prairie wetland ecosystem. Ecosystems 4:346-57.
Zimmer KD, Hanson MA, Butler MG. 2003. Relationships among nutrients, phytoplankton, macrophytes, and fish in prairie wetlands. Can J Fish Aquat Sci 60:721-30.
Acknowledgments
This project was funded by the Minnesota Department of Natural Resources (MN DNR) and the University of St. Thomas. Robert Wright and Adam Berland assisted with site selection, David Staples assisted with ordinal regression, and we thank Jerry Younk, Mitch Haustein, Matt Gorman, Michelle Verant, Luke Schroeder, Sam Friederichs, Patti Gamboni, Rian Cleary, Timothy Houger, and Matthew Bischof for assistance with field and lab work, and Randy Elmer and the Lake Christina Gun Club for logistical support. Leah Domine, Dan McEwen, Jeff Lawrence, and Melissa Drake provided comments that improved this manuscript. We also thank the many private landowners, the United States Fish and Wildlife Service, and the MN DNR for allowing access to the study sites.
Author information
Authors and Affiliations
Corresponding author
Additional information
Author contributions KDZ, MAH, BRH, and MLK all contributed to the design of the study, performed the research, analyzed data, and helped write the article.
Rights and permissions
About this article
Cite this article
Zimmer, K.D., Hanson, M.A., Herwig, B.R. et al. Thresholds and Stability of Alternative Regimes in Shallow Prairie–Parkland Lakes of Central North America. Ecosystems 12, 843–852 (2009). https://doi.org/10.1007/s10021-009-9262-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-009-9262-4