
Abstract
Recent studies have shown that an increase in the population size of sika deer (Cervus nippon) decreases the abundance of rodents but increases that of several invertebrates (Scarabaeidae, Rhaphidophoridae, and earthworms) outside a deep-proof fence in Oku-Nikko, Japan. Therefore, the diet of red foxes (Vulpes vulpes) feeding on these animals is expected to vary according to changes in prey abundance. As red foxes also scavenge deer carcasses, an increase in the number of deer is expected to lead to changes in the red fox diet. In this study, the diet and sightings of red foxes inside and outside the fence were compared. Generalized linear mixed models indicate that the sika deer remains, ground-dwelling insects (Scarabaeidae and Rhaphidophoridae as a group, and Carabidae), and earthworms increased in the fox diet outside the fence, while the ingestion of voles decreased. The difference in prey consumption by red foxes was probably caused by differences in prey availability inside and outside the fence or the presence of available alternative food. These results suggest that red foxes demonstrate dietary plasticity in areas where habitat structure has been substantially modified by deer grazing. Although the abundance of voles, the primary food of red foxes, decreased outside the fence, the sighting rates of red foxes obtained by spotlight count were not significantly different between inside and outside the fence, suggesting that the presence of the others increased or available alternative food may have compensated for the negative effects of decreasing vole abundance.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Interspecific interactions influence species abundance and community composition (Morin 1999). Changes in the abundance of some species or the introduction of exotic species influence changes in species abundance and community composition (Schmitz et al. 2000; Courchamp et al. 2003; Kagata and Ohgushi 2006).
Deer (Cervidae) have recently expanded their range and increased in abundance worldwide, and their impact on natural ecosystems has been dramatic (Côté et al. 2004). Heavy foraging by deer contributes to the modification of vegetation, which also affects animals in various taxonomic groups (Flowerdew and Ellwood 2001; Russell et al. 2001; Rooney and Waller 2003; Côté et al. 2004; Ripple and Beschta 2006; Gill and Fuller 2007; Takatsuki 2009; Ripple and Beschta 2012). In addition, deer affect medium- to large-sized mammals belonging to higher trophic levels in several ways. For example, some carnivores take advantage of scavenging deer carcasses (Jędrzejewski et al. 1989; Selva et al. 2005). Conversely, the black bear (Ursus americanus) population on Anticosti Island may decline because of over browsing of berries by white-tailed deer (Odocoileus virgininianus) (Côté 2005). Interspecific interactions among overabundant deer and other species (particularly mammals at higher trophic levels which often occur at lower densities) can inform effective management practices to preclude potential causes of a decline in these species.
The red fox (Vulpes vulpes), which is distributed across the northern hemisphere, has the widest geographical range of any member of the order Carnivora (Macdonald and Reynolds 2004). Although red foxes are widely distributed in Japan, several populations of the species have been designated as endangered, vulnerable, or near threatened on the Red List at the regional level due to anthropogenic habitat modification (Tsukada 2016). It has also been reported that the number of red foxes decreased due to sarcoptic mange in some regions of Japan (Tsukada et al. 1999; Uraguchi 2008). Furthermore, increasing populations of sika deer (Cervus nippon) may have effects on red foxes in Japan. Small rodent populations, which are important food for red foxes (Koganezawa and Kurokawa 1983; Yamamoto 1994), have declined due to the decrease in understory vegetation and leaf litter caused by sika deer grazing (Suda et al. 2003; Tanaka et al. 2006). Nishio et al. (2013) pointed out that deer density could negatively affect the population of red foxes through a decline in the prey abundance of foxes caused by the modification of habitat structure by deer grazing. Understanding how red foxes respond to changes in food availability and habitat structure due to deer grazing would contribute to effective management plans, which are essential for the conservation of threatened red fox populations in Japan.
In Oku-Nikko, the population of sika deer has been increasing since 1984 and habitat structure has been substantially modified by deer grazing (Koganezawa and Satake 1996; Hasegawa 2008), and a large deer-proof fence was constructed in 2001 to protect vegetation. Similar to other areas, the number of rodents in Oku-Nikko was reported to decrease outside the fence (Koganezawa et al. 2013). However, Seki and Koganezawa (2010, 2013) demonstrated an increasing abundance of earthworms and insects (such as dung beetles) outside of the fence, probably due to the modification of understory vegetation and/or deposition of dung from the increasing sika deer population. As these invertebrates and rodents are primary food resources for red foxes in Japan (Uraguchi 2009), we would expect that the red fox diet would vary accordingly. Furthermore, since red foxes scavenge the carcasses of sika deer (Koganezawa and Kurokawa 1983; Yamamoto 1994; Tsukada and Nonaka 1996; Inagaki et al. 2020), increasing this food source should increase the consumption of deer carcasses by red foxes.
In this study, it was hypothesized that red foxes outside a deer-proof fence would feed more on sika deer, earthworms, and ground-dwelling insects, but less on rodents than inside the fence. To test this hypothesis, the diet of red foxes inside and outside the fence was compared using fecal analysis. In addition, to assess the influence of differences in resource abundance on the population of red foxes, we compared the relative densities of red foxes computed via spotlight counts inside and outside the fence.
Study area
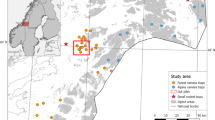
The study was conducted in Oku-Nikko (36° 45′ N, 139° 25′ E) in Nikko National Park, central Honshu, Japan, which is located in a cool temperate zone. The altitude in the study area ranged from 1270 to 1668 m. The density of sika deer, estimated using the block count method during the summer, ranged from 0.0 to 0.4/km2 in 1981–1982 (Mizuno et al. 1982) and 11.5–37.4/km2 in 1995 (Koganezawa and Satake 1996). To protect vegetation, the 17-km deer-proof fence was constructed by the Ministry of the Environment of the Government of Japan in 2001, enclosing an area of 9.8 km2. The mean ± standard deviation (SD) sika deer density, estimated by using the block count method during the study period (2006–2011), was 8.8 ± 5.3/km2 during the summer outside the fence (Tochigi 2014) and 2.6 ± 2.4/km2 during the autumn inside the fence (computed from the data of the Ministry of the Environment of the Government of Japan (2014)). Although the mean density of sika deer outside the fence did not change significantly from 2006 (11.04/km2) to 2011 (10.17/km2), the density inside the fence decreased from 7.2 to 1.2/km2 during the same period (Ministry of the Environment of the Government of Japan 2014; Tochigi 2014).
Although the trapping sites of rodents were different between years, the trapping rates (the number of trapped individuals/trapping nights) of Japanese field voles (Microtus montebelli) during summer inside and outside the fence respectively were 0.138 (124/900) and 0.011 (4/360) in 2009 and 0.088 (57/650) and 0.003 (4/1300) in 2011; those of Japanese field mice (large Japanese field mice, Apodemus speciosus, and small Japanese field mice, A. argenteus) were 0.114 (103/900) and 0.086 (31/360) in 2009 and 0.102 (66/650) and 0.058 (75/1300) in 2011 (Suda and Morita 2014; KO and YS, personal observations).
The understory inside the fence was dominated by dwarf bamboo (Sasa nipponica), an important forage plant for sika deer (Takatsuki 1983, 1986; Yokoyama and Shibata 1998). Outside the fence, the understory was dominated by plants unpalatable to sika deer (e.g., Aster ageratoides ssp. leiophyllus) and bare ground (Koganezawa and Satake 1996; Hasegawa 2008). The vegetative canopy was dominated by deciduous broad-leaved trees and conifers both inside and outside the fence (Koganezawa and Satake 1996). According to data collected between 1981 and 2010 by the Nikko Weather Station (1292 m), the mean annual temperature was 6.9 °C, ranging from a mean of − 4.1 °C in January to a mean of 18.7 °C in August, and the mean annual precipitation was 2176 mm. The ground usually was covered with snow from mid-December until mid-April.
Methods
Collection of fecal samples
A 15.6-km route inside the fence and an 11.0-km route outside the fence were patrolled for fox fecal samples at least once a month from June 2009 to November 2010 and from May to September 2011. Fresh red fox fecal samples found during the sampling of raccoon dog feces from June 2006 to June 2010 (Seki 2011) were also included. The feces of red foxes were discriminated against from those of other carnivores by the size, shape, color, and smell (Yamamoto 1994; Koike et al. 2012).
The area was not completely fenced, with seven non-fenced points at the road and river crossings. Although a section of the sampling route was located within 500 m of non-fenced points, red fox fecal samples were not collected within this range to obviate the possibility that foxes fed inside and then defecated outside the fence or vice versa. The mean ± SD distance from fecal collection points to the non-fenced points was 1015.1 ± 307.2 m (n = 129) inside the fence and 1979.7 ± 1009.5 m (n = 124) outside the fence (50 fecal samples found from May to September 2011 were excluded because the locations of fecal samples were recorded as only inside or outside the fences). The mean core area of the red fox home range in Honshu, Japan (the same subspecies as in the current study), was 1.27 ± 1.02 km2 (SD) in Mt. Nyugasa and 1.40 ± 0.90 km2 in Mt. Ashio (Takeuchi and Koganezawa 1992; Yamamoto 1996); assuming the shape of their core area was circular, the mean radii of the core area were presumed to be 594.6 ± 232.0 m (SD) and 667.9 ± 241.3 m, respectively, which were lower than the mean distances between the fecal sample points and non-fenced points in this study. The 50 excluded fecal samples were > 500 m from the non-fenced points, and the possibility that red foxes had fed inside the fence and defecated outside the fence, and vice versa, was considered to be low.
Fecal analysis
The fecal analysis was done as previously described (Yamamoto 1994; Kaneko et al. 2009; Seki et al. 2014), which involved a procedure for detecting the presence of earthworms. The fecal samples were stored in a freezer at − 20 °C until analysis. Defrosted fecal samples were washed through a 0.5-mm mesh sieve with 1000 mL of water, and materials remaining on the sieve were preserved in 70% ethyl alcohol for identification. Food items were identified to the fullest taxonomic level possible. In addition, the rinsing water was collected into a container (height approximately 85 mm; length 130 mm; and width 140 mm) and left undisturbed for ≥ 15 min. Five milliliters of the sediment was withdrawn using a pipette, diluted with 10 mL of water, and examined for the presence of earthworm chaetae.
Data analysis
Fecal collection dates were divided into winter (January–March), spring (April–June), summer (July–September), and autumn (October–December). The frequency of occurrence (FO) was calculated for each food item in the feces: FO = 100 × number of fecal samples containing a specific food item/total number of fecal samples. Of food items occurring in red fox fecal samples, we targeted Japanese field voles, mice (including large Japanese field mice and/or small Japanese field mice), insects (Scarabaeidae, Carabidae, and Rhaphidophoridae), and earthworms, which were analyzed by Koganezawa et al. (2013) and Seki and Koganezawa (2013), and sika deer.
To assess the effect of location (inside or outside the fence) on the occurrence of each food item, a generalized linear mixed model (GLMM) was used, assuming a binomial error distribution and using a logit link function. Separate models for each type of food item (voles, mice, deer, Scarabaeidae, Carabidae, Rhaphidophoridae, and earthworms) were created. We also created a model of increased insects, including Scarabaeidae and Rhaphidophoridae, which were reported to increase outside the fence in the study area (Seki and Koganezawa 2013). These included the presence or absence of each food item in the feces as a response variable, location (inside or outside the fence) as a fixed effect, and month and year as random effects to correct for repeated sampling. Analyses were conducted in R version 3.2.0 (R Core Team 2015) using the “glmer” function of the “lme4” package.
To assess the fixed effects, the best model was selected by comparing Akaike’s information criterion (AIC) between models with and without location (full and null models, respectively). If the difference in AIC values between the full and null models was less than 2, we considered the two models as competing models (Burnham and Anderson 2002). In addition, odds ratios and their associated 95% confidence intervals (CIs) were estimated in the full models to assess the extent of the fixed effects. Here, odds are represented as the ratio of the probability that the event of interest occurs to the probability that it does not. Odds ratios are the ratios between the odds of two groups; the odds ratio is 1 when there is no relationship (Bland and Altman 2000; Kasuya 2012). For example, an odds ratio > 1 indicates an increased occurrence of food outside the fence.
In this study, the sample sizes in each season were quite different among years. It is well documented that voles can have very dynamic population cycles in Hokkaido, northern Japan (Saito et al. 1998; Stenseth et al. 2003). We also observed some differences in the trapping rates of both voles and mice between years (see the Study area section). However, the ratios of the number of their scat samples to sample sizes during each season were not significantly different among years (Fisher’s exact test, all seasons for voles and mice, P > 0.268; winter could not be analyzed due to small sample sizes). Thus, there could be few biases in the FOs of rodents caused by yearly differences in the sample sizes.
We also analyzed data from spotlight counts (n = 176) conducted between May and November during 2006–2011 by the Laboratory of Wildlife Management of Utsunomiya University to compare the relative densities of red foxes inside and outside the fence. The spotlight surveys were conducted over a fixed 9.0-km route (3.4 km inside the fence and 5.6 km outside the fence; see more details for the method in Seki et al. 2014). To minimize the effects of differences in vegetation on the probability of detecting red foxes between inside and outside the fence, we compared the number of red foxes observed on the road per 10 km (sighting rates) in each survey. As the perpendicular distance of red foxes from the road was not recorded two times (for three individuals) inside the fence and three times (for four individuals) outside the fence, we excluded these data from the analysis. A significant difference, which was set at P < 0.05, was determined between inside and outside the fence using the Wilcoxon signed-rank test in R version 3.2.0 (R Core Team 2015).
Results
A total of 303 fecal samples (159 samples inside the fence and 144 outside the fence) were analyzed (Table 1). Animal remains (mammals, birds, reptiles, fish, insects, Acari, and earthworms), plant remains (seeds, grasses, Musci, and small pieces of leaves, stems, and roots), and artificial remains (wrapping foil and elastic) were found (Table 2). In June 2011, a hoof of a sika deer fawn was found in one fecal sample. Based on the mean FOs of the four seasons, the most frequently occurring animals were Japanese field voles (63%) followed by Rhaphidophoridae (10%), sika deer (10%), Carabidae (8%), and birds (8%) inside the fence, and Japanese field voles (45%), sika deer (29%), Rhaphidophoridae (14%), Carabidae (13%), earthworms (13%), birds (8%), and Scarabaeidae (8%) outside the fence (Table 2). In addition, other frequently occurring food materials inside and outside the fence (respectively) were fruits such as Actinidia spp. (12% and 12%), Vitis coignetiae (8% and 7%), and Malus toringo (7% and 13%), grasses (11% and 5%), and artificial materials (12% and 9%) (Table 2). Although FOs of other small plant materials were relatively high (22% and 6%), these may have been accidentally ingested by red foxes while feeding. The FO of sika deer was relatively higher in spring than in other seasons; the FO for each month during spring was 60% (3/5) in April, 43% (6/14) in May, and 63% (10/16) in June.
The FOs of target food items in each season are shown in Fig. 1. The AIC values for all full models except one mouse model were lower than those for null models (Table 3). The 95% CIs of odds ratios were > 1 for deer (6.68–64.10), increased insect (1.14–3.87), Carabidae (1.09–4.31), and earthworm (1.99–13.67) models (Fig. 2), indicating that their occurrence tended to be higher outside than inside the fence (Fig. 3). However, for the Scarabaeidae and Rhaphidophoridae models, ∆AIC was less than 2 between full and null models and the 95% CIs of odds ratios included 1, while the proportions of values > 1 were relatively large (0.81–4.65 and 0.87–3.48, in Scarabaeidae and Rhaphidophoridae models, respectively; Fig. 2). On the other hand, the 95% CIs of odds ratios for the vole model were lower than 1 (0.38–0.96; Fig. 2), indicating that their occurrence was lower outside than inside the fence (Fig. 3).

Seasonal changes in the frequency of occurrence of the primary food resources of red foxes inside and outside a deer-proof fence in Oku-Nikko, Japan. Fecal samples were collected during the period June 2006 to September 2011. Figures in parentheses give the number of years of fecal sample collecting. Error bars represent the standard error of the mean

Odds ratios and 95% confidence interval for estimation of a fixed effect (location outside a deer-proof fence) in generalized linear mixed models. Increased insects include Scarabaeidae and Rhaphidophoridae

Estimation of the frequency of occurrence (FO) of the primary food resources of red foxes and their 95% confidence interval in generalized linear mixed models inside and outside a deer-proof fence in Oku-Nikko, Japan. The FO values were back-transformed to the 0–100 scale. Increased insects include Scarabaeidae and Rhaphidophoridae
The observed numbers of red foxes on the road were 5 inside the fence and 20 outside the fence. Sighting rates, the mean of which was 0.084 ± 0.037 (the standard error of the mean) inside the fence and 0.203 ± 0.047 outside the fence, were not significantly different between inside and outside the fence (Wilcoxon signed-rank test, V = 95, P = 0.178).
Discussion
Red fox diet
Red foxes are adaptable and opportunistic omnivores, with a diet ranging from invertebrates to mammals, birds, and fruit (Macdonald and Reynolds 2004), with small mammals as their main food resource (e.g., Uraguchi 2009; Murdoch et al. 2010; Días-Ruiz et al. 2013; Spencer et al. 2014; Tsukada et al. 2014). We found that red foxes ate small mammals, sika deer, insects, earthworms, and fruits in Oku-Nikko (Table 2). Red foxes mainly consumed Japanese field voles, whereas mice were not fed on frequently (Table 2). In Oku-Nikko, the trapping rates of mice were not lower compared to those of the voles (see the Study area section), which does not necessarily mean that there were few mice in the study area, but that red foxes prefer Japanese field voles. This preference for voles is reported not only in Japan but in other countries too (Abe 1975; Macdonald 1977; Misawa 1979; Yoneda 1979; Kondo 1980; Koganezawa and Kurokawa 1983; Doncaster et al. 1990; Jȩdrzejewski and Jȩdrzejewska 1992; Yamamoto 1994; Sidorovich et al. 2006; Panzacchi et al. 2008b; Kondo and Shiraki 2012, 2013; Meisner et al. 2014; Lanszki et al. 2020). The low consumption of mice by red foxes, even in areas with abundant mice, has been shown in other areas, including Japan (Macdonald 1977; Misawa 1979; Yoneda 1979; Doncaster et al. 1990; Panzacchi et al. 2008b; Kondo and Shiraki 2012, 2013).
Red foxes also frequently consumed sika deer outside the fence especially between winter and spring (Table 2, Fig. 2). Although the FOs of sika deer during winter seemed to be almost the same between inside and outside the fence, this could be due to the small sample size or temporal distribution of the samples. All samples collected inside the fence were derived from one year (2010), whereas samples outside the fence were from 4 years (2006–2010) (Table 1); sika deer remains were found in one fecal sample in 2007 and in two samples from 2010 outside the fence. According to the Nikko Weather Station, snowfall during winter was 247 cm in 2010, which was greater than that in 2007–2009 (117 cm, 228 cm, and 108 cm, respectively). Thus, we inferred that deer mortality could have been higher in 2010 with severe weather conditions, which contributed to higher FO inside the fence during winter. The mortality of sika deer older than 1 year is highest during winter and spring (Minami et al. 2009). The consumption of deer carcasses by red foxes (also observed by Seki 2017) was probably opportunistic. Such seasonal patterns for the consumption of ungulate carcasses by red foxes have been reported in Japan and in other countries (Koganezawa and Kurokawa 1983; Cavallini and Volpi 1996; Padial et al. 2002; Cagnacci et al. 2003; Sidorovich et al. 2006; Giuliano et al. 2019). The red fox is known to be the major predator of roe deer (Capreolus capreolus) fawns (Borg 1962; Aanes and Andersen 1996; Kjellander and Nordström 2003; Jarnemo et al. 2004; Jarnemo and Liberg 2005; Panzacchi et al. 2008a, b; Melis et al. 2013). The roe deer is quite small as compared to sika deer: the adult weight of roe deer is ≤ 35 kg (Macdonald and Barrett 1993), although that of the subspecies of sika deer (C. n. centralis) ranges from 37.6 to 100.0 kg (Jiang 2015). Thus, it is considered to be more difficult for red foxes to predate on sika deer fawns. However, red fox predation on sika deer fawns is believed to occur quite often. During spring, sika deer remained in a higher proportion each month. As only a few sika deer older than 1 year die during the period between May and September (Minami et al. 2009), the availability of their carcasses was probably low in May–June. Thus, we inferred that the higher consumption of sika deer by red foxes during the period was not derived from the carcasses but from predation on deer fawns since the calving season of sika deer in the study area was estimated to be from May to August, with a peak in the middle of May (Iwamoto et al. 2009). In this study, a deer fawn hoof was found in a fecal sample in June. Tsukada and Nonaka (1996) also reported that red foxes fed on sika deer fawns in June. These results indicate that sika deer fawns may be an important food resource for red foxes. Chasing a sika deer under a year old (presumably approximately 10 months) by a red fox was also observed on March 27, 2007, in the study area (Y.S., personal observation). Red fox predation on fawns of white-tailed deer, similar sized to sika deer (Macdonald and Barrett 1993), was also observed in South Carolina (Epstein et al. 1983).
Invertebrates such as insects and earthworms were also eaten by red foxes inside and outside the fence (Table 2). Red foxes ate Coleoptera (Scarabaeidae and Carabidae) from spring to summer and Orthoptera (Rhaphidophoridae) from summer to autumn (Table 2, Fig. 1). In Japan, red foxes eat insects (Misawa 1979; Yoneda 1979; Koganezawa and Kurokawa 1983; Ueuma et al. 2005). These studies did not identify insects to order and family levels, but where this was done, foxes were reported to mainly eat Coleoptera and Orthoptera (Yamamoto 1994; Koike et al. 2012; Kondo and Shiraki 2013). A similarly high occurrence of Coleoptera or Orthoptera among insects in diets of red foxes has also been observed in many other countries (Doncaster et al. 1990; Serafini and Lovari 1993; Lucherini and Crema 1994; Cagnacci et al. 2003; Baker et al. 2006; Lanszki et al. 2007; Panzacchi et al. 2008b; Murdoch et al. 2010; Remonti et al. 2012; Drygala et al. 2013; Giuliano et al. 2019; O’Connor et al. 2020), indicating that these insect groups are the primary food resource from among insects in the fox diet. Earthworms are also frequently hunted by red foxes in some areas (Macdonald 1980). In England, for example, earthworms constituted up to 38% of the red fox diet in Wiltshire (Baker et al. 2006) and up to 45% in Oxford (Doncaster et al. 1990). One possible explanation for the higher consumption of earthworms by red foxes is prey availability, because foraging for earthworms by red foxes has been shown to be correlated with earthworm abundance (Macdonald 1980). In Finland and Germany, however, the consumption of earthworms by red foxes was low, whereas other carnivores frequently consumed earthworms in the same area (Kauhala et al. 1998; Drygala et al. 2013). Such differences in the consumption of earthworms by red foxes are probably caused by differences in the abundance of their primary prey, that is, small mammals, as Weber (1996) has shown that red foxes consumed a higher proportion of earthworms in the years when voles became scarce.
Red foxes also frequently ate plant materials such as fruits, particularly Actinidia spp., Vitis coignetiae, and Malus toringo during autumn (Table 2). The higher fruit consumption of red foxes has also been reported in other countries (Calisti et al. 1990; Cagnacci et al. 2003; Remonti et al. 2012; Bakaloudis et al. 2015; Lanszki et al. 2019; O’Connor et al. 2020). In Japan, the higher FO of Actinidia spp. has also been reported in other areas (Misawa 1979; Yamamoto 1994; Tsukada and Nonaka 1996; Ueuma et al. 2005; Kondo and Shiraki 2013); V. coignetiae was also found in red fox fecal samples (Misawa 1979; Yamamoto 1994; Ueuma et al. 2005), but the FO of fruit was not high. It has been suggested that the red fox prefers A. arguta over V. coignetiae (Kondo and Shiraki 2012, 2013). In Hungary, however, red foxes frequently consumed V. vinifera (Lanszki et al. 2019), suggesting that the fruit selection of the red fox can differ even though the fruits are from the same genus. Furthermore, frequently occurring fruits in other areas of Japan were different, i.e., Rubus vernus in Mt. Hakusan (Ueuma et al. 2005) and Akebia spp. in Mt. Nyugasa (Yamamoto 1994), indicating that fruit consumption by red foxes may depend on the availability of fruit (Tsukada and Nonaka 1996). Grasses were found with relatively high FOs but are presumably to improve occasional intestinal disorders which occur in carnivores (Imaizumi 2007).
Effects of sika deer density on red fox diet and population
Voles, sika deer, insects, and earthworms were the principal food resources of red foxes in the current study. The FOs of voles were higher inside than outside the fence, with an odds ratio < 1, indicating that red foxes fed more on Japanese field voles inside the fence (Table 3, Figs. 2–3). For sika deer, Carabidae, increased insects (including Scarabaeidae and Rhaphidophoridae), and earthworms, the FOs were lower inside the fence and the odds ratio were > 1, indicating that these food resources were eaten more outside the fence than inside (Table 3, Figs. 2–3). We propose several possible explanations for these results, which are not necessarily mutually exclusive.
The densities of sika deer were different inside and outside the fences (see Study area section). Populations of Japanese field voles and Japanese field mice decreased and insect (Scarabaeidae and Rhaphidophoridae) and earthworm populations increased following modification of the understory vegetation and/or defecation by sika deer in the study area (Seki and Koganezawa 2010, 2013; Koganezawa et al. 2013). Consequently, differences in the feeding pattern of red foxes inside and outside the fence were probably motivated by differences in resource abundance (Abe 1975; Jȩdrzejewski and Jȩdrzejewska 1992). Although the abundance of Japanese field mice was higher inside than outside the fence (Koganezawa et al. 2013; see also the Study area section), the current study did not show greater consumption of mice by red foxes inside the fence, which was probably due to their low proclivity for mice. Carabidae abundance was not significantly different inside and outside the fence (Seki and Koganezawa 2013), but the insects were consumed more outside the fence by red foxes, indicating that other factors other than insect abundance were driving the consumption of the insect by red foxes.
Consumption of certain prey by red foxes can be driven by foraging opportunity. In general, greater vegetative cover can provide increased protection from predators of small-bodied animals (Birney et al. 1976; Peles and Barrett 1996). It has been reported that the consumption of voles tended to decrease with increasing forest cover (Yoneda 1983; Kidawa and Kowalczyk 2011). The mean herbaceous layer height and coverage were reported to be higher inside than outside the fence in the present study area (Okuda et al. 2012; Okuda 2013). Lower understory coverage outside the fence is expected to increase fox foraging opportunities, which could contribute to the higher consumption of invertebrates outside the fence. However, even though there is higher coverage inside the fence, the consumption of voles by red foxes was still higher inside the fence. This indicates that the consumption of voles by red foxes is more likely to be affected by changes in prey abundance than changes in understory coverage.
Another possible factor that could cause a higher consumption of certain prey by red foxes outside the fence is a dietary shift towards alternative prey species with a decline in their main prey, that is, voles. Red fox predation is a plausible mechanism determining the dynamics of the small vertebrate community, and their predation pressure on alternative prey species varies in relation to the prey abundance and consumption of voles (Angelstam et al. 1985; Lindström et al. 1994). Dietary shifts in red foxes have been shown not only towards vertebrates such as grouse, hare, deer fawn, and mammalian carrion (Angelstam et al. 1985; Lindström et al. 1994; Kjellander and Nordström 2003; Sidorovich et al. 2006) but also for invertebrates, wild fruits, and rubbish (Ferrari and Weber 1995; Weber 1996). Thus, lower vole availability outside the fence could force red foxes to feed mainly on more deer carcasses in winter, deer fawn and invertebrates in spring, invertebrates in summer, and fruits and invertebrates in autumn (Table 2). If this concept is correct, Carabidae is presumably the main alternative prey among insects because there is no significant difference in their abundance between inside and outside the fence (Seki and Koganezawa 2013).
Días-Ruiz et al. (2013) showed a latitudinal pattern in the diet of the red fox. The current results suggest that the diet of red foxes varies even on a relatively small spatial scale, supporting that red foxes can plastically vary their diet in the area with substantially modified habitat structure by sika deer grazing. The influence of these changes in diet on the population of red foxes was not significant during the study period. The most probable reason for this is that the effect of vole decline caused by sika deer on the fox population has been offset by the increased availability of other prey such as insects, earthworms, and carcasses and fawns of sika deer outside the fence. Such compensation has also been reported in other areas. Weber et al. (1999) reported that the increase in the consumption of anthropogenic food probably prevented a decrease in red fox breeding success. In areas with thick snow cover and thus low availability of small rodents, increased ungulate carcass availability may compensate for the negative effects of severe winters (Selås and Vik 2006). Although the vole abundance decreased outside the fence, the overall effects of sika deer on the red fox population may be non-significant, at least during the study period, due to the presence of other increased or available alternative foods.
In contrast, populations of other carnivores such as raccoon dogs (Nyctereutes procyonoides) and Japanese badgers (Meles anakuma) probably increase when their food (insects and earthworms) increases due to habitat modification by deer (Seki and Koganezawa 2013; Seki et al. 2014). Such differences could be caused by differences in their dietary plasticity. The principal food of Japanese badgers and raccoon dogs is invertebrates and fruits in Japan (Kaneko 2009; Saeki 2009). The proportion of rodent consumption by Japanese badges and raccoon dogs was reported to be very low, whereas the consumption by red foxes was quite high in the same area (Yamamoto 1994). The ability of Japanese badgers and raccoon dogs to hunt rodents may be lower than that of the red fox because the legs of the former carnivores are relatively shorter than those of the fox. Only a few Japanese badgers and raccoon dogs were observed inside the fence (Seki and Koganezawa 2013; Seki et al. 2014), which also suggests that their plasticity against changes in the food abundance caused by sika deer is lower than red fox plasticity.
As the carcasses and fawns of several ungulates are primary food resources for the red fox, as previously mentioned, recent increases in deer abundance worldwide have had a positive direct effect on the red fox population. Red fox predation also plays a key role in contributing to the high mortality of roe deer fawns (Aanes and Andersen 1996; Jarnemo et al. 2004; Jarnemo and Liberg 2005; Panzacchi et al. 2008a). Our results also suggest the possibility of red fox predation on sika deer fawn. If the higher occurrence of sika deer in the scats of red foxes in spring was largely derived from red fox predation on sika deer fawn, conservation of red foxes may potentially contribute to decreasing the population of sika deer. In addition, the effect of deer abundance on populations of small mammals, ground-dwelling insects, and earthworms has been demonstrated in several areas (Gardner et al. 1997; Stewart 2001; Kanda et al. 2005; Karberg and Lilleskov 2009; Buesching et al. 2011), which could also influence the population of the red fox. Interactions between deer and red fox food sources will improve our understanding of the plasticity of red foxes with habitat modification by deer and will facilitate the design and implementation of deer management practices.
Availability of data and material
The datasets analyzed during the current study are available from the corresponding author on reasonable request.
Code availability
Not applicable.
References
Aanes R, Andersen R (1996) The effects of sex, time of birth, and habitat on the vulnerability of roe deer fawns to red fox predation. Can J Zool 74:1857–1865
Abe H (1975) Winter food of the red fox, Vulpes vulpes schrencki Kishida (Carnivora : Canidae), in Hokkaido, with special reference to vole populations. Appl Entomol Zool 10:40–51
Angelstam P, Lindstörm E, Widén P (1985) Sychronous short-term population fluctuations of some birds and mammals in Fennoscandia—occurrence and distribution. Holarct Ecol 8:285–298
Bakaloudis DE, Bontzorlos VA, Vlachos CG, Papakosta MA, Chatzinikos EN, Braziotis SG, Kontsiotis VJ (2015) Factors affecting the diet of the red fox (Vulpes vulpes) in a heterogeneous Mediterranean landscape. Turk J Zool 39:1151–1159
Baker P, Furlong M, Southern S, Harris S (2006) The potential impact of red fox Vulpes vulpes predation in agricultural landscapes in lowland Britain. Wildl Biol 12:39–50
Birney EC, Grant WE, Baird DD (1976) Importance of vegetative cover to cycles of Microtus populations. Ecology 10:40–51
Bland JM, Altman DG (2000) The odds ratio. BMJ 320:1468
Borg K (1962) Predation on roe deer in Sweden. J Wildl Manag 26:133–136
Buesching CD, Newman C, Jones JT, Macdonald DW (2011) Testing the effects of deer grazing on two woodland rodents, bankvoles and woodmice. Basic Appl Ecol 12:207–214
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach, 2nd edn. Springer Verlag, New York
Cagnacci F, Lovari S, Meriggi A (2003) Carrion dependence and food habits of the red fox in an Alpine area. Ital J Zool 70:31–38
Calisti M, Ciampalini B, Lovari S, Lucherini M (1990) Food habits and trophic niche variation of the red fox Vulpes vulpes in a Mediterranean coastal area. Rev Ecol (Terre Vie) 45:309–320
Cavallini P, Volpi T (1996) Variation in the diet of the red fox in a Mediterranean area. Rev Ecol (Terre Vie) 51:171–189
Côté SD (2005) Extirpation of a large black bear population by introduced white-tailed deer. Conserv Biol 19:1668–1671
Côté SD, Rooney TP, Tremblay JP, Dussault C, Waller DM (2004) Ecological impacts of deer overabundance. Annu Rev Ecol Evol Syst 35:113–147
Courchamp F, Ghapuis J, Pasgal M (2003) Mammal invaders on islands: impact, control and control impact. Biol Rev 78:347–383
Días-Ruiz F, Delibes-Mateos M, García-Moreno JL, López-Martín JM, Ferreira C, Ferreras P (2013) Biogeographical patterns in the diet of an opportunistic predator: the red fox Vulpes vulpes in the Iberian Peninsula. Mammal Rev 43:59–70
Doncaster CP, Dickman CR, Macdonald DW (1990) Feeding ecology of red foxes (Vulpes vulpes) in the city of Oxford, England. J Mammal 71:188–194
Drygala F, Werner U, Zoller H (2013) Diet composition of the invasive raccoon dog (Nyctereutes procyonoides) and the native red fox (Vulpes vulpes) in north-east Germany. Hystrix Ital J Mamm 24:190–194
Epstein MB, Feldhamer GA, Joyner RL (1983) Predation on white-tailed deer fawns by bobcat, foxes, and alligators: predator assessment. Proc Annu Conf Southeast Assoc Fish Wildl Agencies 37:161–172
Ferrari N, Weber J-M (1995) Influence of the abundance of food resources on the feeding habits of the red fox, Vulpes vulpes, in western Switzerland. J Zool (Lond) 236:117–129
Flowerdew JR, Ellwood SA (2001) Impacts of woodland deer on small mammal ecology. Forestry 74:277–287
Gardner SM, Hartley SE, Davies A, Palmer SCF (1997) Carabid communities on heather moorlands in northeast Scotland: the consequences of grazing pressure for community diversity. Biol Conserv 81:275–286
Gill RMA, Fuller RJ (2007) The effects of deer browsing on woodland structure and songbirds in lowland Britain. IBIS 149:119–127
Giuliano D, Battisti A, Bassano B (2019) Feeding ecology of the red fox in the Soana Valley, Gran Paradiso National Park. J Mountain Ecol 12:1–18
Hasegawa J (2008) Changes in Nature of Tochigi Prefecture. Self Publication, Tochigi (in Japanese)
Imaizumi T (2007) Wild dogs. Data House, Tokyo (in Japanese)
Inagaki A, Allen ML, Maruyama T, Yamazaki K, Tochigi K, Naganuma T, Koike S (2020) Vertebrate scavenger guild composition and utilization of carrion in an East Asian temperate forest. Ecol Evol 10:1223–1232
Iwamoto C, Matsuda N, Maruyama T, Koganezawa M (2009) Estimates of calving season of sika deer in Oku-Nikko. Bull on Wildl Tochigi Pref 36:18–21 (in Japanese)
Jarnemo A, Liberg O (2005) Red fox removal and roe deer fawn survival—a 14-year study. J Wildl Manag 69:1090–1098
Jarnemo A, Liberg O, Lockowandt S, Olsson A, Wahlström K (2004) Predation by red fox on European roe deer fawns in relation to age, sex, and birth date. Can J Zool 82:416–422
Jȩdrzejewski W, Jȩdrzejewska B (1992) Foraging and diet of the red fox Vulpes vulpes in relation to variable food resources in Biatowieza National Park, Poland. Ecography 15:212–220
Jędrzejewski W, Jȩdrzejewska B, Szymura A (1989) Food niche overlaps in a winter community of predators in the Białowieża Primeval Forest, Poland. Acta Theriol 34:487–496
Jiang Z (2015) Sika deer. In: Seki Y, Enari H, Kodera Y, Tsuji Y (eds) Field survey methods for wildlife management: from the identification of field signs of mammals to data analysis. Kyoto University Press, Kyoto, pp 39–52 (in Japanese)
Kagata H, Ohgushi T (2006) Bottom-up trophic cascades and material transfer in terrestrial food webs. Ecol Res 21:26–34
Kanda N, Yokota T, Shibata E, Sato H (2005) Diversity of dung-beetle community in declining Japanese subalpine forest caused by an increasing sika deer population. Ecol Res 20:135–141
Kaneko Y (2009) Meles anakuma Temminck, 1844. In: Ohdachi SD, Ishibashi Y, Iwasa M, Saitoh T (eds) The wild mammals of Japan. Shoukadoh, Kyoto, pp 258–260
Kaneko Y, Shibuya M, Yamaguchi N, Fujii T, Okumura T, Matsubayashi K, Hioki Y (2009) Diet of Japanese weasels (Mustela itatsi) in a sub-urban landscape: implication for year-round persistence of local populations. Mamm Study 34:97–105
Karberg NJ, Lilleskov EA (2009) White-tailed deer (Odocoileus virginianus) fecal pellet decomposition is accelerated by the invasive earthworm Lumbricus terrestris. Biol Invasions 11:761–767
Kasuya E (2012) Generalized linear models. Kyoritsu Publisher, Tokyo (in Japanese)
Kauhala K, Laukkanen P, von Rége I (1998) Summer food composition and food niche overlap of the raccoon dog, red fox and badger in Finland. Ecography 21:457–463
Kidawa D, Kowalczyk R (2011) The effects of sex, age, season and habitat on diet of the red fox Vulpes vulpes in northeastern Poland. Acta Theriol 56:209–218
Kjellander P, Nordström J (2003) Cyslic voles, prey switching in red fox, and roe deer dynamics—a test of the alternative prey hypothesis. Oikos 101:338–344
Koganezawa M, Kurokawa M (1983) Altitudinal distribution of middle-sized mammals in Nikko area, Tochigi Prefecture (II) – focused on the winter distribution and the food habits of the red fox (Vulpes vulpes). Memoirs of Tochigi Pref Mus 1:67–82 (in Japanese with English abstract)
Koganezawa M, Satake C (1996) Effects of grazing by sika-deer on the vegetation of Oku-Nikko and their management. Trans Nat Found Proj 5:57–66 (in Japanese with English abstract)
Koganezawa M, Seki Y, Okuda K, Fujitsu A, Ito M (2013) Effects of high deer density on small mammals and their predators in Oku-Nikko, Tochigi prefecture, Japan. Trans Nat Found Proj 21:77–84 (in Japanese with English abstract)
Koike S, Morimoto H, Goto Y, Kozakai C, Yamazaki K (2012) Insectivory by five sympatric carnivores in cool-temperate deciduous forests. Mamm Study 37:73–83
Kondo T (1980) Food habits of the red fox (Vulpes vulpes japonica) and the Japanese marten (Martes melampus melampus). Memoirs of Osaka Kyoiku University, Ser. 3 1:19–23 (in Japanese with English abstract)
Kondo A, Shiraki S (2012) Preference for specific food species of the red fox Vulpes vulpes in Abashiri, eastern Hokkaido. Mamm Study 37:43–46
Kondo A, Shiraki S (2013) Relation between food habits and food resource of the red fox Vulpes vulpes schrencki in Abashiri, eastern Hokkaido. Bull Shiretoko Mus 35:47–58 (in Japanese with English abstract)
Lanszki J, Zalewski A, Horváth G (2007) Comparison of red fox Vulpes vulpes and pine marten Martes martes food habits in a deciduous forest in Hungary. Wildl Biol 13:258–271
Lanszki Z, Purger JJ, Bocz R, Szép D, Lanszki J (2019) The stone marten and the red fox consumed predominantly fruits all year round: a case study. Acta Zool Acad Sci Hung 65:45–62
Lanszki Z, Horváth GF, Bende Z, Lanszki J (2020) Differences in the diet and trophic niche of three sympatric carnivores in a marshland. Mamm Res 65:93–104
Lindström ER, Andrén H, Angelstam O, Cederlund G, Hörnfeldt B, Jäderberg L, Lemnell P-A, Martinsson B, Sköld K, Swenson JE (1994) Disease reveals the predator: sarcoptic mange, red fox predation, and prey populations. Ecology 75:1042–1049
Lucherini M, Crema G (1994) Seasonal variation in diet and trophic niche of the red fox in an Alpine habitat. Z Säugetierkd 59:1–8
Macdonald DW (1977) On food preference in the red fox. Mammal Rev 7:7–23
Macdonald DW (1980) The red fox, Vulpes vulpes, as a predator upon earthworms, Lumbricus terrestris. Z Tierpsychol 52:171–200
Macdonald DW, Barrett P (1993) Mammals of Europe. Princeton University Press, Princeton, Oxford
Macdonald DW, Reynolds JC (2004) Red fox Vulpes vulpes Linnaeus, 1758. In: Sillero-Zubiri C, Hoffmann M, Macdonald DW (eds) Canids: foxes, wolves, jackals and dogs. Status survey and conservation action plan. IUCN/SSC Canid Specialist Group, Gland, Switzerland, Cambridge, pp 129–136
Meisner K, Sunde P, Clausen KK, Clausen P, Fælled CC, Hoelgaard M (2014) Foraging ecology and spatial behavior of the red fox (Vulpes vulpes) in a wet grassland ecosystem. Acta Theriol 59:377–389
Melis C, Nilsen EB, Panzacchi M, Linnell JDC, Odden J (2013) Roe deer face competing risks between predators along a gradient in abundance. Ecosphere 4:1–12
Minami M, Ohnishi N, Takatsuki S (2009) Survival patterns of male and female sika deer on Kinkazan Island, northern Japan. In: McCullough DR, Takatsuki S, Kaji K (eds) Sika deer – biology and management of native and introduced populations. Springer, Tokyo, pp 375–384
Ministry of the Environment of the Government of Japan (2014) Deer Management in Senjougahara in Oku-Nikko. http://www.env.go.jp/park/nikko/effort/deer.html. Accessed May 2014
Misawa E (1979) Changes in the food habits of the red fox, Vulpes vulpes schrencki KISHIDA according to habitat conditions. J Mamm Soc Jpn 7:311–320 (in Japanese with English abstract)
Mizuno A, Ishida K, Tanida K, Ueuma Y, Torii H, Azuma S, Maruyama N, Furubayashi K, Itoh T, Takatsuki S, Ootaishi N (1982) Dynamics of larger mammals (bear and deer) and natural environment in Japanese forest. In: Agency of Environment (ed) Comprehensive research on the development of basic technologies for management and conservation of natural environment research report in 1981; comprehensive research on the development of the methodology of urban environmental planning research report in 1981. Agency of Environment, Tokyo, pp. 117-1–117-22 (in Japanese with English abstract)
Morin PJ (1999) Community Ecology. Blackwell, Oxford
Murdoch JD, Munkhzul T, Buyandelger S, Reading RP, Sillero-Zubiri C (2010) Seasonal food habits of corsac and red foxes in Mongolia and the potential for competition. Mamm Biol 75:36–44
Nishio T, Takada MB, Uno H, Sato Y, Yanagawa H (2013) Factors influencing road-kill frequency of red fox Vulpes vulpes with attention to the effects of landscape structure and harvest of sika deer Cervus nippon yesoensis for nuisance control and hunting in eastern Hokkaido, Japan. Mamm Sci 53:301–310 (in Japanese with English abstract)
O’Connor JM, Srivastava SK, Tindale NW, Burnett SE (2020) From carrion to Christmas beetles: the broad dietary niche of the red fox in a hybrid ecosystem in south-eastern Queensland. Aust J Zool 67:82–93
Okuda K (2013) Effects of vegetation modification due to high deer density on breeding bird communities in Oku-Nikko, Japan. PhD thesis. Tokyo University of Agriculture and Technology, Tokyo, Japan (in Japanese)
Okuda K, Seki Y, Koganezawa M (2012) Effects of vegetation modification due to high deer density on breeding bird communities in Oku-Nikko, Japan. J Jpn For Soc 94:236–242 (in Japanese with English abstract)
Padial JM, Avila E, Gil-Sánchez JM (2002) Feeding habits and overlap among red fox (Vulpes vulpes) and stone marten (Martes foina) in two Mediterranean mountain habitats. Mamm Biol 67:137–146
Panzacchi M, Linnell JDC, Odden J, Odden M, Andersen R (2008a) When a generalist becomes a specialist: patterns of red fox predation on roe deer fawns under contrasting conditions. Can J Zool 86:116–126
Panzacchi M, Linnell JDC, Serrao G, Eie S, Odden M, Odden J, Andersen R (2008b) Evaluation of the importance of roe deer fawns in the spring–summer diet of red foxes in southeastern Norway. Ecol Res 23:889–896
Peles JD, Barrett G (1996) Effects of vegetative cover on the population dynamics of meadow voles. J Mammal 77:857–869
R Core Team (2015) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Remonti L, Balestrieri A, Ruiz-González A, Gómez-Moliner BJ, Capelli E, Prigioni C (2012) Intraguild dietary overlap and its possible relationship to the coexistence of mesocarnivores in intensive agricultural habitats. Popul Ecol 54:521–532
Ripple WJ, Beschta RL (2006) Linking a cougar decline, trophic cascade, and catastrophic regime shift in Zion National Park. Biol Conserv 133:397–408
Ripple WJ, Beschta RL (2012) Trophic cascades in Yellowstone: the first 15 years after wolf reintroduction. Biol Conserv 145:205–213
Rooney TP, Waller DW (2003) Direct and indirect effects of white-tailed deer in forest ecosystems. For Ecol Manag 181:165–176
Russell FL, Zippin DB, Fowler NL (2001) Effects of white-tailed deer (Odocoileus virginianus) on plants, plant populations and communities: a review. Am Midl Nat 146:1–26
Saeki M (2009) Nyctereutes procyonoides (Gray, 1834). In: Ohdachi SD, Ishibashi Y, Iwasa MA, Saitoh T (eds) The wild mammals of Japan. Shoukadoh, Kyoto, pp 216–217
Saito T, Stenseth NC, Bjørnstad ON (1998) The population dynamics of the vole Clethrionomys rufocanus in Hokkaido, Japan. Res Popul Ecol 40:61–76
Schmitz OJ, Hambäck PA, Beckerman AP (2000) Trophic cascades in terrestrial systems: a review of the effects of carnivore removals on plants. Am Nat 155:141–153
Seki Y (2011) Trophic interaction between the sika deer and the raccoon dog in Oku-Nikko, Japan. PhD thesis. Tokyo University of Agriculture and Technology, Tokyo, Japan (in Japanese)
Seki Y (2017) Effects of sika deer on medium- to large-sized carnivores. In: Kaji K, Iijima H (eds) Sika deer in Japan: the science and management of overabundant populations. University of Tokyo Press, Tokyo, pp 83–101 (in Japanese)
Seki Y, Koganezawa M (2010) Factors influencing the increase in earthworms outside deer-proof fences in Oku-Nikko, central Japan: the influence of the modification of understory vegetation by sika deer. J Jpn For Soc 92:241–246 (in Japanese with English abstract)
Seki Y, Koganezawa M (2013) Does sika deer overabundance exert cascading effects on the raccoon dog population? J For Res 18:121–127
Seki Y, Okuda K, Koganezawa M (2014) Indirect effects of sika deer on Japanese badgers. Mamm Study 39:201–208
Selås V, Vik JO (2006) Possible impact of snow depth and ungulate carcasses on red fox (Vulpes vulpes) populations in Norway, 1897–1976. J Zool 269:299–308
Selva N, Jȩdrzejewska B, Jȩdrzejewski W, Wajrak A (2005) Factors affecting carcass use by a guild of scavengers in European temperate woodland. Can J Zool 83:1590–1601
Serafini P, Lovari S (1993) Food habits and trophic niche overlap of the red fox and the stone marten in a Mediterranean rural area. Acta Theriol 38:233–244
Sidorovich VE, Sidorovich AA, Izotova IV (2006) Variations in the diet and population density of the red fox Vulpes vulpes in the mixed woodlands of northern Belarus. Mamm Biol 71:74–89
Spencer EE, Crowther MS, Dickman CR (2014) Diet and prey selectivity of three species of sympatric mammalian predators in central Australia. J Mammal 95:1278–1288
Stenseth NC, Viljugrein H, Saitoh T, Hansen TF, Kittilsen MO, Bølviken E, Glöckner F (2003) Seasonality, density dependence, and population cycles in Hokkaido voles. Proc Natl Acad Sci U S A 100:11478–11483
Stewart AJA (2001) The impact of deer on lowland woodland invertebrates: a review of the evidence and priorities for future research. Forestry 74:259–270
Suda K, Morita J (2014) Species identification for Apodemus speciosus, A. argenteus, and Microtus montebelli using the footprint recording method, and its application for relative abundance estimation. Mamm Sci 54:231–241 (in Japanese with English abstract)
Suda K, Araki R, Maruyama N (2003) Effects of sika deer on forest mice in evergreen broad-leaved forests on the Tsushima Islands, Japan. Bios Conserv 5:63–70
Takatsuki S (1983) The importance of Sasa nipponica as a forage for sika deer (Cervus nippon) in Omote-Nikko. Jpn J Ecol 33:17–25
Takatsuki S (1986) Food habits of sika deer on Mt. Goyo, northern Honshu. Ecol Res 1:119–128
Takatsuki S (2009) Effects of sika deer on vegetation in Japan: a review. Biol Conserv 142:1922–1929
Takeuchi M, Koganezawa M (1992) Home range and habitat utilisation of the red fox Vulpes vulpes in the Ashio Mountains, central Japan. J Mamm Soc Jpn 17:95–110
Tanaka M, Saito M, Ohi K, Fukuda H, Shibata E (2006) Relationship between dwarf−bamboo growth and small rodnts on Mt. Ohdaigahara, central Japan, with special reference to construction of deer-proof fences. J Jpn For Soc 88:348–353
Tochigi (2014) Conservation and management of Sika deer of 2012 in Tochigi Prefecture—report on monitoring result. Natural Environment Division of Tochigi Prefecture, Tochigi (in Japanese)
Tsukada H (2016) Population and habitat management techniques of small- to medium-sized mammals. In: Hayama S, Miura S, Kaji K, Suzuki M (eds) Wildlife Management: Theory and Techniques. Buneido, Tokyo, pp 451–465 (in Japanese)
Tsukada H, Nonaka N (1996) Foraging behavior of red foxes Vulpes Vulpes schrencki utilizing human food in the Shiretoko National Park, Hokkaido. Mamm Study 21:137–151
Tsukada H, Okada H, Yamanaka M, Nonaka N, Oku Y (1999) The spreading of sarcoptic mange among red foxes Vulpes vulpes and its impact on the local fox population in Shiretoko Peninsula, Hokkaido, Japan. Mamm Sci 39:247–256 (in Japanese with English abstract)
Tsukada H, Li W, Duo H, Guo Z, Fu Y, Peng M, Shen X, Jing J, Yuan A, Ni M, He S, Huang F, Feng K, Ishikawa K, Kobayashi I, Shinohara A, Nonaka N (2014) A comparison of the diet and fine-scale distribution of sympatric Tibetan and red foxes in Qinghai, PR China. Wildl Biol 20:356–361
Ueuma Y, Tokuno C, Tsuji M (2005) Food of red fox (Vulpes vulpes japonica), Japanese marten (Martes melanpus melanpus) and Hondo stoat (Mustela ermine nippon) analyzed dropping contents on the trails in Mt. Hakusan. Annu Rep Hakusan Nat Conserv Center 32:31–36 (in Japanese)
Uraguchi K (2008) Disease and ecology: red fox. In: Takatsuki S, Yamagiwa J (eds) Middle-, and large-sized mammals including primates (Mammalogy in Japan 2). University of Tokyo Press, Tokyo, pp 149–171 (in Japanese)
Uraguchi K (2009) Vulpes vulpes (Linnaeus, 1758). In: Ohdachi SD, Ishibashi Y, Iwasa MA, Saitoh T (eds) The wild mammals of Japan. Shoukadoh, Kyoto, pp 214–215
Weber J-M (1996) Food selection by adult red foxes Vulpes vulpes during a water vole decline. Wildl Biol 2:283–288
Weber J-M, Meia J-S, Meyer S (1999) Breeding success of the red fox Vulpes vulpes in relation to fluctuating prey in central Europe. Wildl Biol 5:241–244
Yamamoto Y (1994) Comparative analyses on food habits of Japanese marten, red fox, badger and raccoon dog in Mt. Nyugasa, Nagano Prefecture, Japan. Nat Environ Sci Res 7:45–52 (in Japanese with English abstract)
Yamamoto Y (1996) Home range and habitat utilization of the red fox Vulpes vulpes japonica in the sub-alpine belt in central Japan. Nat Environ Sci Res 9:47–61 (in Japanese with English abstract)
Yokoyama S, Shibata E (1998) Characteristics of Sasa nipponica grassland as a summer forage resource for sika deer on Mt. Ohdaigahara, central Japan. Ecol Res 13:193–198
Yoneda M (1979) Prey preference of the red fox, Vulpes vulpes schrencki KISHIDA (carnivora: canidae), on small rodents. Appl Entomol Zool 14:28–35
Yoneda M (1983) Influence of red fox predation upon a local population of small rodents—III. seasonal changes in predation pressure, prey preference and predation effect. Appl Entomol Zool 18:1–10
Acknowledgements
We are grateful to Utsunomiya University accommodation staff in Oku-Nikko for their assistance during survey periods. In this study, we used data from spotlight counts conducted between 2006 and 2011 by students from the Laboratory of Wildlife Management University. We are grateful to them for the use of their data and their important contributions to the survey. We would like to thank two anonymous referees and an associate editor for their valuable advice and constructive criticism of the manuscript and Editage (www.editage.com) for English language editing.
Author information
Authors and Affiliations
Contributions
Y.S. and K.O. formulated the idea and conceived the experiment. Y.S., K.O., and M.K. designed the experiment. Y.S. and M.I. performed the experiments and analyzed the data. Y.S. wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Seki, Y., Ito, M., Okuda, K. et al. Effects of sika deer density on the diet and population of red foxes. Eur J Wildl Res 67, 30 (2021). https://doi.org/10.1007/s10344-021-01475-5
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10344-021-01475-5