
Abstract
Global climate change impacts the functioning and productivity of forest ecosystems at various spatiotemporal scales across a wide range of biomes. Although summer temperatures are considered the main driver of boreal tree growth, the importance of soil moisture availability is likely to rise with decreasing latitude and increasing warming. Here, we combine dendrochronological measurements with evidence from tree growth modeling and remote sensing to quantify the effect of climate on phenology and productivity of Scots pines (Pinus sylvestris L.) in southern Siberia. Between 1960 and 2017, pine ring widths along a latitudinal transect from 53° to 56°N were mainly controlled by the availability of summer soil moisture. This finding challenges the common belief that summer temperatures are the predominant growth control in boreal forests. Moreover, we show that earlier growing season onsets can compensate for warming-induced drought stress. Despite the phenotypic plasticity of Scots pines to adapt to warmer and drier conditions, we speculate that predicted climate change will likely exceed the species’ physiological tolerance in much of Eurasia’s forest-steppe by the end of the twenty-first century.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The Russian boreal forest represents the world’s largest continuous forest biome, accounting for circa 50% of all boreal forests (Krankina et al. 1997) and storing an estimated 46–67 petagrams of carbon in its living biomass (Houghton et al. 2007; Krankina et al. 1996). However, predicted climate change would probably affect these forest ecosystems (Allen et al. 2015, 2010; Choat et al. 2012; Kharuk et al. 2017) by modifying the phenology and productivity of trees, compromising the forests’ capacity to sequester carbon from the atmosphere (Bradley et al. 1999; Trumbore et al. 2015), and thus affecting the global carbon cycle negatively (Bradshaw and Warkentin 2015; Giradin et al. 2016).
The significant warming trend documented across Siberia during the last decades (Groisman et al. 2012) affects different forest types’ geographical distribution and growth (Tchebakova et al. 2016). For example, the risk of climate change impacts on forest ecosystems is considerable in southern Siberia, where tree growth during the short but hot summers tends to be more constrained by soil moisture availability (Arzac et al. 2018, 2021; Tabakova et al. 2020). Therefore, the predicted warming and temperature-induced drought (Anisimov et al. 2008; Shvidenko et al. 2012) will influence the phenology and physiology of trees, affecting forest health (Frank et al. 2015; Tchebakova et al. 2016).
Plant phenological patterns are sensitive to climate variability (Menzel et al. 2020; Piao et al. 2019). Current warming trends will likely result in earlier starts and delayed ends of the growing seasons (Jeong et al. 2011; Zhang et al. 2004). An extensive literature body reports increasing growing season length of boreal forests in recent time (Fu et al. 2015; He et al. 2018a, 2018b; Menzel et al. 2006; Spinoni et al. 2015; Yang et al. 2017; Zhou et al. 2003). Hence, geographical regions experiencing above-average warming, as Russian boreal forests and forest-steppe, are of high interest for understanding the response of main forest tree species to changing climate conditions.
Long-term xylem phenology studies are time-consuming and usually restricted in space and temporal scale. Therefore, process-based models and remote sensing tools might facilitate plant phenology research (He et al. 2018a; Yang et al. 2017). For example, tree radial growth (wood formation) models, such as the Vaganov–Shashkin model (VS-model; Vaganov et al. 2006), can simulate tree growth and phenology in response to daily variability in both air temperature and soil moisture (Anchukaitis et al. 2020; He et al. 2018a; Shishov et al. 2016; Tychkov et al. 2019; Vaganov et al. 2006). At the same time, vegetation indices like the EVI (Enhanced Vegetation Index; Huete et al. 1999), based on photosynthetic activity changes, leaf biomass and canopy structure (Duchemin et al. 1999; Reed et al. 1994) are used to estimate intra-annual changes in canopy greenness and annual forest productivity and phenology at high spatiotemporal resolution. Nevertheless, remote sensing datasets (e.g., EVI) are constrained to temporal series no earlier than the year 2000, whereas tree growth simulations could be extended over longer time periods.
We combined evidence from dendrochronological measurements, tree growth modeling and remote sensing at three sites with different water balance levels to increase our understanding of Scots pine xylem phenology and growth variability in southern Siberia. More specifically, this study aims to: (i) compare VS-model phenological simulations against MODIS (Moderate Resolution Imaging Spectroradiometer) satellite observations, (ii) evaluate inter-annual changes in pine secondary growth (xylem phenology) via the VS-model over the 1960–2017 period, and (iii) link the timing of phenological phases with spring temperatures as the trigger of the growing season onset. We hypothesized that (i) water availability will be the predominant climatic factor constraining pine growth even in the wettest site, and (ii) warmer springs conditions and the plastic responses of pine cambial activity will compensate the expected ring width reductions due to summer drought stress by shifting pine phenology to earlier onsets after snowmelt at the driest site.
Materials and methods
Study area
Three Scots pine (Pinus sylvestris L.) sites with different water balance levels were sampled along a latitudinal transect (56–53°N) in southern Siberia (Krasnoyarsk Krai, Russia; Fig. 1). The northern site was located in the experimental plot “Pogorelsky bor” within the northern forest-steppe (hereafter moderate site; 56° 22′ N, 92° 57′ E). The central site in the National Park “Krasnoyarsk Stolby” in the mid-mountain taiga (hereafter wet site; 55° 54′ N, 92° 43′ E). The southern site in the National Park “Shushensky bor” in the transition from the southern taiga into the forest-steppe (hereafter dry site; 53° 18′ N, 91° 58′ E). The moderate and dry sites are mono-specific P. sylvestris forest stands. In contrast, pine trees co-occur with Larix sibirica Ledeb., Betula pendula Roth, and Abies sibirica Ledeb at the wet site.

Study area. Stars represent the location of the sampling sites. M, moderate (Pogorelsky bor); W, wet (Stolby); D, dry (Shushensky bor)
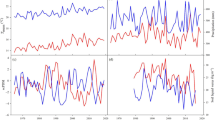
The transect is characterized by a cold continental climate (Kottek et al. 2006), with mean annual temperatures ranging from 0.1 °C at the wet site to 1.5 °C at the dry site, and the total annual precipitation ranging from 308 mm at the dry site to 668 mm at the wet site (Table 1). Daily climate data (1960–2017) were obtained from the nearest meteorological station to the moderate and dry sites (www.meteo.ru) and from a local station in the National Park “Krasnoyarsk Stolby” at the wet site. During the studied period, mean annual temperature has increased at an average rate of 0.3 ºC per decade (R2 = 0.25; P < 0.001), warming faster at the moderate and wet sites in recent time (Fig. 2a). In recent times, the total annual precipitation has decreased at the wet site without significant trends at the moderate and wet sites (Fig. 2b). Water balance (WB = P – PET), where P is the precipitation, and PET is the potential evapotranspiration calculated by the Thornthwaite equation (Thornthwaite 1948), was estimated for the 1960–2017 period. Lower WB values correspond to the dry site, but only significant decreasing trends were found at the wet site (Fig. 2c).

Climate conditions (temperature (a), precipitation (b) and water balance (c)) during April-September over the period 1960 − 2017. Water balance (WB) was calculated as WB = P-PET, where P represents the precipitation and PET the potential evapotranspiration. Trends are shown for the full period (ful., solid lines) and the early (ear., short-dashed lines) and recent (rec., long-dashed lines) periods. Regression coefficients (b1) in bold type are significant at P < 0.001, coefficients underlined are significant at P < 0.01 and coefficients in italic are significant at P < 0.05 (non-significant coefficients are not shown). W, wet site; M, moderate site; D, dry site
Sampling design and tree-ring measurements
Twenty mature dominant or co-dominant trees were sampled at each site during autumn 2018. Two wood cores per tree were taken at breast height using a 5-mm diameter increment borer. Cores were polished using a belt sanding machine with successive sandpaper (up to 600 grit) until a smooth and flat surface was obtained.
Wood cores were scanned at 3200 dpi with an Epson Perfection V800 scanner (Epson, Japan). Tree-ring width (RW) was measured on the scanned cores using CooRecorder version 9.3 (Cybis Elektronik & Data AB, Sweden). The accuracy of cross-dating was verified using COFECHA (Grissino-Mayer 2001), and each raw RW series was standardized using ARSTAN (Cook and Holmes 1996). Spline functions with a 50% frequency cut-off at 32 years were fitted individually to the raw series. Residual chronologies were computed using the bi-weight robust average. The approach is flexible enough to reduce any of the non-climatic lower frequency variances while at the same time preserving the high-frequency, inter-annual climate information (Cook and Peters 1981), which we consider relevant for the further assessment of the past five decades.
Simulations of tree growth and phenological dynamics
Tree growth and phenological key dates were simulated by the VS-Oscilloscope (VS-O) Ver. 1.32 (Shishov et al. 2016; Tychkov et al. 2019), the visual parametrization interface of the environmental block of tree-ring growth of the Vaganov–Shashkin model available at http://vs-genn.ru/. The model simulates daily tree growth rates (Gr) based on two pre-defined growth-limiting functions, given the temperature and soil moisture conditions occurring every day of the year. Thus, daily tree growth is defined by the limitation of the growth rates dependent on temperature or soil moisture and weighted by the photoperiod determined by the site’s latitude (Tychkov et al 2019; Vaganov et al. 2006). Consequently, the Gr = min (GrW, GrT) · GrE, where GrW, GrT and GrE are the daily partial growth rates based on soil moisture, temperature and solar irradiation, respectively. The GrE depends on the latitude (Gates, 2012), whereas GrW and GrT are nonlinear functions scaled between 0 and 1 (Fig. A1).
To assess the effect of local conditions on growth, the model requires a calibrated parametrization by fitting the simulated RW chronology against the observed one. Therefore, the evaluation of the VS-model is based on the values of Pearson’s correlation coefficient (r) and the Gleichläufigkeit synchrony (glk) between the simulated and actual RW chronologies (Shishov et al. 2016). As the output of the model, values of daily growth rates (GrW, GrT and Gr), simulated RW indices and annual estimations of the date for the start (SoS), end (EoS) and length (LoS) of the growing season are provided. By definition, the SoS occurs when the daily temperature is greater than or equal to the value of the minimum temperature required for tree growth (Tmin), and the temperature sum for the previous ten days reaches some critical level (Tbeg). The EoS is defined as the last day within a year when the growth rate's value is greater than the critical growth rate (Vcr) and the temperature sum is no longer higher or equal Tbeg (Vaganov et al. 2006). Finally, the LoS is estimated as the number of days between SoS and EoS.
In this study, the VS-model was parameterized for each site over the 1960–2017 period. Residual RW chronologies and daily climate data were used as input data. The calibration against the residual RW chronologies was performed from 1960 to 1988 and verification from 1989 to 2017.
Validation of phenology simulations against remote sensing
Simulated phenological series were compared to derived data from the Moderate Resolution Imaging Spectroradiometer (MODIS) for a 14-year period (2001–2014) at each site. Annual estimations of vegetation phenology were obtained from the MODIS Land Cover Dynamics data product MCD12Q2 (Ver. 005; Ganguly et al. 2010). The data set includes the timing for the onset of four phenological phases (greenup, maturity, senescence and dormancy) with 500 m-ground resolution. The phenophase transition dates were based on logistic models (Zhang et al. 2006), by the estimation of the curvature-change rate of the Enhanced Vegetation Index (EVI), computed from composed eight-day MODIS Nadir Bidirectional Reflectance Distribution Function (BRDF; Schaaf et al. 2002) and the Adjusted Land Surface Reflectance (NBAR-EVI) data fitted with a sigmoidal function. The seasonal onset of continuous EVI increment corresponds to annual vegetation greenup and the seasonal onset of EVI minima corresponds to vegetation dormancy dates, associated with SoS and EoS, respectively (Zhang et al. 2006).
To test differences in mean simulated phenological dates (SoS, EoS and LoS) compared to derived MODIS datasets (and due to gaps in the MODIS dataset), we applied a one-way ANOVA (data source as a fixed factor). In addition, to compare inter-annual patterns, Pearson’s correlations and linear regressions were performed.
Pine growth and phenology response to climate
Pine growth response to climate was assessed using Pearson's correlations between monthly climatic data (temperature, precipitation and water balance) and RW residual chronologies for the 1960–2017 period. Correlation coefficients were calculated with a temporal window comprising from the previous year July to September of the current growing season. In addition, we grouped the climatic variables in climatic seasons for the current growing year: winter (December-February), spring (March–May), summer (June–August) and autumn (September–November), as well during critical months for growth (May–July). Finally, climate correlations were re-assessed by splitting the study period into two equally long 29-year intervals, 1960–1988 and 1989–2017.
Linear regression functions were applied to analyze the link between phenological dynamics and spring temperatures over the 1960–2017 period. Linear regressions were also used to evaluate trends in the simulated pine phenological series (SoS and EoS) over the full and the early/late periods. Finally, since the SoS is defined by temperature in the model, we evaluated how spring temperature increments might affect the start of the growing season in the future. We forecasted potential changes in SoS by extrapolating current temperature to possible increments of 4 to 5 ºC as predicted by the end of the century in southern Siberia (Groisman et al. 2012).
Results
Climate response of radial growth
Pine climate sensitivity was consistent at all the sites during the growing season (Fig. 3a). Secondary growth was favored by late spring and early summer precipitation, with earlier signal (May, r = 0.47; P < 0.001) at the dry site and later signals (June) at the moderate and wet sites (r = 0.32; P < 0.01 and r = 0.26; P < 0.05, moderate and wet sites, respectively). The aggregated effect of spring (March–May) precipitation showed a significant positive impact on growth at the dry site. Precipitation from previous growing season (August and November) also favored tree growth at the dry site (r = 0.33; P < 0.01 and r = 0.25; P < 0.05, respectively). Whereas winter precipitation (previous December) showed a negative effect at the wet site (r = − 0.36; P < 0.01). Warm April and previous October temperatures enhanced tree growth at the moderate and wet sites (r = 0.32 and r = 0.36; P < 0.01, respectively). On the other hand, May and June temperatures exerted a negative effect at the dry site (r = − 0.35; P < 0.01 and r = − 0.23; P < 0.05), with the aggregated effect of May–July temperatures amplifying the negative effect (r = − 0.41; P < 0.001).

Correlations (Pearson’s coefficient) between ring width (RW) residual chronologies of Pinus sylvestris with total monthly precipitation (P), mean monthly temperature (T) and water balance (WB) for the 1960–2017 period (a), early time 1960–1988 (b) and recent time 1989–2017 (c). Correlations were calculated from July of previous year (uppercase letters) to September of current growth year (lowercase letters), and by climatic seasons. W, wet site; M, moderate site; D, dry site; mjj, (May–July); win (winter); spr (spring); sum (summer); aut (autumn). Vertical-dashed lines separate monthly and seasonal responses
Pearson’s correlations between RW residual chronologies and water balance (P-PET) suggested a dependence on water availability at different temporal windows among the studied sites. Thus, the moderate and wet sites responded to a shorter water availability window (May, r = 0.34; P < 0.01 and r = 0.22; P < 0.05, respectively). Whereas the dry site responded for a longer temporal window (late spring and mid-late summer), with the highest signal in August (r = 0.37; P < 0.01). Previous December signals mimic the response to precipitation at the moderate and wet sites.
Pine climate sensitivity decreased in the recent period (1989–2017) compared to the early period (1960–1988). As a result, temperature and precipitation signals from late spring and summer declined and became not significant, except at the dry site where the negative effect of May–July and summer temperatures remained (Fig. 3b & c). Interestingly, the impact of water availability negatively increased, with the response to water balance becoming significant during July–August at the dry site. Moreover, the aggregated effect of water balance consecutive months showed a shift from the spring to the summer (and extended to the autumn) from the early to the recent period, with a maximal signal reached during the summer (0.52; P < 0.01).
Tree growth model simulations
Tree growth simulations were significantly correlated (P < 0.05) with the residual RW chronologies at all the sites for both the calibration and the verification period (Fig. A2, see also Table A1 for model parameters). However, at the wet site, the performance of the model was suboptimal. Correlation coefficients for the calibration period (1960–1988) ranged from r = 0.31 at the wet site (suboptimal performance of the model) to r = 0.66 at the moderate site, and the synchrony (glk, Gleichläufigkeit) between series from 66% at the wet site to 69% at the moderate and dry sites. Correlations for the verification period (1989–2017) ranged from r = 0.37 at the wet site to r = 0.51 at the moderate site, and a glk between 64% at the wet site to 75% at the dry site.
According to VS-model simulations, the growing season length averaged from 132 ± 14 (mean ± SD) at the wet site to 155 ± 12 days at the dry site over the 1960–2017 period (Table 2). The SoS was earlier at the dry site (DOY 121 ± 9) and later at the wet site (DOY 142 ± 12). The model highlighted a growth limitation by soil moisture from early summer (DOY 152 ± 9) to early autumn (DOY 254 ± 11), whereas the importance of temperature goes from autumn to mid-spring (Fig. A3).
Validation of phenology simulations against remote sensing
MODIS-derived phenological dataset showed a longer growing season length at the dry site (161 ± 5; mean ± SD) and shorter at the moderate site (136 ± 12) for the 2001–2014 period (Table 3). During the same period, the VS-model simulated a longer growing season length at the dry site (162 ± 11) and shorter at the wet site (118 ± 22). One-way ANOVA did not reveal significant differences in mean SoS at any site (P > 0.05). However, mean EoS was significantly different at the moderate and wet sites (F = 25.9; P < 0.05 and F = 5.3; P < 0.05, respectively) and LoS at the moderate site (F = 1.9; P < 0.05). Pearson’s correlations between simulated and MODIS-derived SoS were positive at all the sites but only significant at the wet site (r = 0.46; P < 0.05). Whereas EoS correlations coefficient were significant only at the dry site (r = − 0.75; P < 0.001; Fig. A4).
Pinus sylvestris phenological dynamics and response to climate
Tree growth simulations revealed a trend of earlier SoS at all the sites over the 1960–2017 period (Fig. 4). Linear regressions suggested the advance of SoS at a rate from 2.3 days/decade at the dry site (R2 = 0.18; P < 0.01) to 2.6 days/decade at the moderate site (R2 = 0.15; P < 0.05). The dry site is the only showing a significant delay of EoS (R2 = 0.10; P < 0.05) at a rate of 1.4 days/decade. No significant trends were found at the wet site. When phenological trends were evaluated in the early and recent periods, the SoS advance increased to 5.6 days/decade from 1989 to 2017 at the dry site (Fig. 4).

Phenological trends simulated by the VS-model for the starting of the growing season (circles) and ending of the growing season (squares) and linear regressions for the full period (1960–2017, black-dashed lines), early period (1960–1988, blue-dashed lines) and recent period (1989–2017, red-dashed lines). Regression coefficients (b1) and significance are shown in the figures. Wet site (a), moderate site (b), dry site (c)
Linear regression between simulated SoS and spring temperature over the full period (Fig. 5) suggested a strong response to April temperature at the moderate (R2 = 0.23; P < 0.001) and the dry (R2 = 0.49; P < 0.001) sites, whereas the wet site responded on May temperature (R2 = 0.39; P < 0.001). In this sense, a rise in spring temperature supposes the advance of SoS on an average rate of 3.1 days/ºC along the studied transect. The VS-model also forecasted the lengthening of P. sylvestris growing season linked to rising temperatures by the end of the current century. Temperature increments between 4 and 5 ºC might suppose an advance of SoS from 12 to 16 days at the moderate site and 16 to 17 days at the dry site.

Linear regression between the simulated starting of the growing season (SoS) with mean April (moderate and dry sites) and May (wet site) temperature for the period 1960–2017. Trends (gray-dashed lines) and regression coefficients (b1) and significance are shown in the figure
Discussion
Pinus sylvestris growth and climate response
Contrary to the common assumption that temperature is the primary driver of boreal forest growth (Nemani et al. 2003), pine secondary growth was mainly limited by water availability at the three sites. Precipitation and water balance signals echo previous findings in the region (Antonova and Stasova 1993; Arzac et al. 2018, 2021; Babushkina et al. 2019; Fonti and Babushkina 2016; Shah et al. 2015; Tabakova et al. 2020), highlighting the relevance of water availability for tree growth even at the coldest and wettest site. The shift in the timing and strength of climate sensitivity among the sites reflects an alleviation of drought stress with increasing latitude. In contrast, the positive response to water balance and precipitation (linked to a negative response to temperature) at the dry site suggests a strong limitation by temperature-induced drought (Arzac et al. 2021), being an adequate soil moisture level necessary for cambial reactivation and tree growth in drought-prone areas (Zeng et al. 2020). Our results also showed a decrease in the climate sensitivity to temperature and precipitation during recent years (1989–2017), in agreement with previous observations in the region (Arzac et al. 2018). However, the response to water balance increased at the dry site during the summer months in recent times, suggesting favorable conditions at the beginning of the growing season (spring) but harshened later due to summer drought. This condition may be exacerbated in future due to climate change, limiting tree growth. Differences among the sites might also be influenced by photoperiod (Körner and Basler 2010) and increasing radiation limitation due to cloudiness. Therefore, higher temperatures and lower radiation limitations (by lower cloudiness) might induce maximal growth at the driest site, despite the robust climatic control.
Pinus sylvestris phenology and climate
The advance of SoS during the last five decades is in line with previous observations and simulations in northern latitudes (Jeong et al. 2011; Piao et al. 2015; Schwartz et al. 2006; Shen et al. 2015). Moreover, the average rate of 2.5 days/decade detected over the studied sites mimics the magnitude reported by previous studies based on model simulations (e.g., Yang et al. 2017) and remote sensing (e.g., Myneni et al. 1997) in boreal regions. In this sense, warmer springs induce a faster snow melting, increasing water and nutrients availability and promoting an earlier onset of the cambial activity (Dye and Tucker 2003; Kirdyanov et al. 2003; Peng et al. 2010; Zeng et al. 2020). Despite the robust climatic control at the dry site, it also showed a longer growing season and tree growth, with clear dependence of SoS to spring warming (April). Therefore, our results might suggest that earlier growing seasons start at the dry site, due to warm springs, compensate for much of the expected growth reduction by low water availability levels later in the growing season (e.g., Shiklomanov et al. 2013).
VS-model simulations based on increasing spring temperatures projected earlier growing seasons start by the end of the century. Since the model defines the SoS based on temperature, it is possible to forecast potential realistic changes in the SoS. However, the limitation in daily climate forecast for the region difficult the prediction of growth rates and EoS dependent on temperature and soil moisture (as well as increasing drought limitation). Nevertheless, phenology changes linked to increasing temperature might substantially affect forest functioning and global carbon dynamics in future (Keenan et al. 2014; Piao et al.2019). Longer vegetation seasons in global ecosystems (Zhu et al. 2016) could trigger photosynthetic activity and carbon assimilation (Keenan et al. 2014; Zhang et al. 2020), increasing forest productivity (Nemani et al. 2003; Schwartz et al. 2006; Zhang et al. 2020).
Pinus sylvestris has shown high physiological adaptability to contrasting environmental conditions. However, the warming-induced drought might also lead to future water deficits affecting forest health in the area (Arzac et al. 2018; Kharuk et al. 2013a, 2013b; Tabakova et al. 2020). Therefore, if the induced drought exceeds pine tolerance, the Russian boreal forest structure might be affected by forest-steppe expansion over taiga (Soja et al. 2007; Tchebakova et al. 2016). Thus, leading to a decrease in forest cover and potentially reducing the productivity and carbon assimilation capacity of the taiga ecosystem (Gustafson et al. 2011; Shvidenko et al. 2012). Therefore, precipitation and snowmelt/snowpack will be determining factors to keep the soil moisture balance for tree growth and phenology in future, as is evidenced by the increasing response to water balance at the dry site in recent time (Arzac et al. 2018, 2019; Babushkina et al. 2015; Grippa et al. 2005; Shen et al. 2015).
VS-model simulations and remote sensing
Some uncertainties might arise from the use of model simulations in combination with remote sensing observations. For example, the available MODIS dataset (2001–2014) might not be robust enough to evaluate temporal trends (Yang et al. 2017). In addition, the presence of gaps in the data set because of cloudiness might also induce errors when comparing both approaches. Observed differences in EoS at the moderate and the wet sites could respond to EVI's limited capacity to reproduce growing dynamics in the evergreen needleleaf forest (Wu et al. 2014). A limitation exacerbated in mixed forests as at the wet site where the fraction of photosynthetically active radiation from deciduous species (L. sibirica and B. pendula) might overlap with P. sylvestris. In addition, a suboptimal performance of the model at the same site might respond to non-climatic factors, like the impact of insect outbreaks in the area (Safonova et al. 2019). However, their impact has been detected only in the last ten years.
Although the comparison of inter-annual patterns between modeled and remote sensing phenological series by Pearson’s correlation was poor, mean SoS values were comparable, highlighting the model's potential to reconstruct phenological dynamics over long spatiotemporal scales (e.g., Jevšenak et al. 2020), particularly for the determination of the SoS (Tumajer et al. 2021). Nevertheless, the reconstruction of EoS and LoS dynamics based on the model might be uncertain, limiting the capacity of the model to reconstruct phenological dynamics beyond SoS. The VS-O has shown limitations determining the EoS compared to direct phenological observations (e.g., Butto et al. 2020; Jevsenak et al. 2020; Tumajer et al. 2021). A potential source of the model’s poor performance may be the overly simple statistical approach to estimate the EoS, based on values of integral growth rate and a mechanistic parameter (Vcr and Tbeg) which are constant for every year of simulation, and not considering non-climatic factors as hormones concentration and non-structural carbohydrates dynamics (Hartmann et al. 2017; Furze et al. 2019).
Conclusions
The combination of multiple data sources allowed us to disclose the effect of climate on Pinus sylvestris growth and fill gaps in xylem phenology (SoS dynamics) studies over extended spatiotemporal scales. Our findings suggest that P. sylvestris secondary growth is plastic enough to adapt to a certain threshold of harsh environmental conditions, adjusting its growth strategy to favorable climatic windows. The release of summer drought stress by earlier SoS, triggered by spring warming, might compensate for tree growth limitation by low water levels at the dry site later in the growing season. However, predicted rising temperatures and drought events in the southern Siberian forest-steppe might exceed the species’ physiological tolerance, compromising tree growth and phenology. Altogether ultimately affecting the vitality, dynamics and distribution of Siberian forests, with critical ecological consequences on the global scale.
References
Allen CD, Macalady AK, Chenchouni H, Bachelet D, McDowell N, Vennetier M, Kitzberger T, Rigling A, Breshears DD, Hogg (Ted) EH, Gonzalez P, Fensham R, Zhang Z, Castro J, Demidova N, Lim JH, Allard G, Running SW, Semerci A, Cobb N (2010) A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For Ecol Manag 259:660–684. https://doi.org/10.1016/j.foreco.2009.09.001
Allen C, Breshears D, McDowell N (2015) On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 6:1–55. https://doi.org/10.1890/ES15-00203.1
Anchukaitis KJ, Evans MN, Hughes MK, Vaganov EA (2020) An interpreted language implementation of the Vaganov–Shashkin tree-ring proxy system model. Dendrochronologia 60:125677. https://doi.org/10.1016/j.dendro.2020.125677
Anisimov O, Anokhin Y, Boltneva L, Vaganov E, Gruza G, Zaitsev A, Zolotokrylin A, Izrael Y, Insarov G, Karol I, Kattsov V, Kobysheva N, Kostianoy A, Krenke A, Mescherskaya A, Mirvis V, Oganesyan V, Pchelkin A, Revich B, Reshetnikov A, Semenov V, Sirotenko O, Sporyshev P, Terziev F, Frolov I, Khon V, Tsyban A, Sherstyukov B, Shiklomanov I, Yasuk Anisimov V (2008) Assessment report on climate change and its consequences in Russian Federation. General Summary. Federal Service for Hydrometeorology and Environmental Monitoring (Roshydroment), Moscow, p. 24.
Antonova GF, Stasova VV (1993) Effects of environmental factors on wood formation in Scots pine stems. Trees 7:214–219. https://doi.org/10.1007/BF00202076
Arzac A, Babushkina EA, Fonti P, Slobodchikova V, Sviderskaya IV, Vaganov EA (2018) Evidences of wider latewood in Pinus sylvestris from a forest-steppe of Southern Siberia. Dendrochronologia 49:1–8. https://doi.org/10.1016/j.dendro.2018.02.007
Arzac A, Popkova M, Anarbekova A, Olano JM, Gutiérrez E, Nikolaev A, Shishov V (2019) Increasing radial and latewood growth rates of Larix cajanderi Mayr. and Pinus sylvestris L. in the continuous permafrost zone in Central Yakutia (Russia). Ann for Sci. https://doi.org/10.1007/s13595-019-0881-4
Arzac A, Tabakova MA, Khotcisnkaia K, Koteneva A, Kirdyanov AV, Olano JM (2021) Linking tree growth and intra-annual density fluctuations to climate in suppressed and dominant Pinus sylvestris L. trees in the forest-steppe of Southern Siberia. Dendrochronologia. https://doi.org/10.1016/j.dendro.2021.125842
Babushkina EA, Vaganov EA, Belokopytova LV, Shishov VV, Grachev AM (2015) Competitive strength effect in the climate response of scots pine radial growth in south-central Siberia forest-steppe. Tree-Ring Res 71:106–117. https://doi.org/10.3959/1536-1098-71.2.106
Babushkina EA, Zhirnova DF, Belokopytova LV, Tychkov II, Vaganov EA, Krutovsky KV (2019) Response of four tree species to changing climate in a moisture-limited area of South Siberia. Forests. https://doi.org/10.3390/f10110999
Bradley NL, Leopold AC, Ross J, Huffaker W (1999) Phenological changes reflect climate change in Wisconsin. Proc Natl Acad Sci USA 96:9701–9704. https://doi.org/10.1073/pnas.96.17.9701
Bradshaw CJA, Warkentin IG (2015) Global estimates of boreal forest carbon stocks and flux. Glob Planet Change 128:24–30. https://doi.org/10.1016/j.gloplacha.2015.02.004
ButtòV SV, Tychkov I, Popkova M, He M, Rossi S, Deslauriers A, Morin H (2020) Comparing the cell dynamics of tree-ring formation observed in microcores and as Predicted by the Vaganov–Shashkin model. Front Plant Sci 11:1–16. https://doi.org/10.3389/fpls.2020.01268
Choat B, Jansen S, Brodribb TJ, Cochard H, Delzon S, Bhaskar R, Bucci SJ, Feild TS, Gleason SM, Hacke UG, Jacobsen AL, Lens F, Maherali H, Martínez-Vilalta J, Mayr S, Mencuccini M, Mitchell PJ, Nardini A, Pittermann J, Pratt RB, Sperry JS, Westoby M, Wright IJ, Zanne AE (2012) Global convergence in the vulnerability of forests to drought. Nature 491:752–755. https://doi.org/10.1038/nature11688
Cook ER, Holmes R (1996) Guide for computer program ARSTAN,in: Grissino-Mayer HD, Holmes RL, Fritts HC (Eds.), The International Tree-Ring Data Bank Program Library Version 2.0 User’s Manual. Laboratory of Tree-Ring Research , University of Arizona, Tucson, USA, pp. 75–87
Cook ER, Peters K (1981) The smoothing spline: a new approach to standardizing forest interior tree-ring width series for dendroclimatic studies. Tree-Ring Bull 41:45–53
Duchemin B, Goubier J, Courrier G (1999) Monitoring phenological key stages and cycle duration of temperate deciduous forest ecosystems with NOAA/AVHRR Data. Remote Sens Environ 67(1):68–82
Dye DG, Tucker CJ (2003) Seasonality and trends of snow-cover, vegetation index, and temperature in northern Eurasia. Geophys Res Lett 30:3–6. https://doi.org/10.1029/2002GL016384
Fonti P, Babushkina EA (2016) Tracheid anatomical responses to climate in a forest-steppe in Southern Siberia. Dendrochronologia 39:32–41. https://doi.org/10.1016/j.dendro.2015.09.002
Frank DC, Reichstein M, Bahn M, Thonicke K, David F, Mahecha MD, Smith P, van der Velde M, Vicca S, Babst F, Beer C, Buchmann N, Canadell JG, Ciais P, Cramer W, Ibrom A, Miglietta F, Poulter B, Rammig A, Seneviratne SI, Walz A, Wattenbach M, Zavala MA, Zscheischler J (2015) Effects of climate extremes on the terrestrial carbon cycle: Concepts, processes and potential future impacts. Glob Chang Biol 21:2861–2880. https://doi.org/10.1111/gcb.12916
Fu YH, Zhao H, Piao S, Peaucelle M, Peng S, Zhou G, Ciais P, Song Y, Vitasse Y, Zeng Z, Janssens IA, Huang M, Menzel A, Pen J (2015) Declining global warming effects on the phenology of spring leaf unfolding. Nature 526:104–107. https://doi.org/10.1038/nature15402
Furze ME, Huggett BA, Aubrecht DM, Stolz CD, Carbone MS, Richardson AD (2019) Whole-tree nonstructural carbohydrate storage and seasonal dynamics in five temperate species. New Phytol 221:1466–1477. https://doi.org/10.1111/nph.15462
Gates DM (2012) Biophysical ecology. Cour Corp
Ganguly S, Friedl MA, Tan B, Zhang X, Verma M (2010) Land surface phenology from MODIS: characterization of the collection 5 global land cover dynamics product. Remote Sens Environ 114:1805–1816. https://doi.org/10.1016/j.rse.2010.04.005
Girardin MP, Bouriaud O, Hogg EH, Kurz W, Zimmermann NE, Metsaranta JM, de Jong R, Frank DC, Esper J, Büntgen U, Guo XJ, Bhatti J (2016) No growth stimulation of Canada’s boreal forest under half-century of combined warming and CO2 fertilization. Proc Natl Acad Sci USA. https://doi.org/10.1073/pnas.1610156113
Grippa M, Kergoat L, Le Toan T, Mognard NM, Delbart N, L’Hermitte J, Vicente-Serrano SM (2005) The impact of snow depth and snowmelt on the vegetation variability over central Siberia. Geophys Res Lett 32:1–4. https://doi.org/10.1029/2005GL024286
Grissino-Mayer HD (2001) Evaluating crossdating accuracy: a manual and tutorial for the computer program COFECHA. Tree-Ring Res 57:205–221
Groisman PY, Blyakharchuk TA, Chernokulsky AV, Arzhanov MM, Marchesini LB, Bogdanova G, Borzenkova II, Bulygina ON, Karpenko AA, Karpenko L, Knight R, Khoin VC, Korovin G, Meshcherskaya A, Mokhov I, Parfeonva E, Razuvaev V, Speranskaya N, Tchebakova N, Vygodskaya N (2012) Climate Changes in Siberia. In: Groisman P, Gutman G (Eds) Regional environmental changes in siberia and their global consequences. Springer Science + Business Media, pp 57–109. https://doi.org/10.1007/978-94-007-4569-8
Gustafson EJ, Shvidenko AZ, Scheller RM (2011) Effectiveness of forest management strategies to mitigate effects of global change in south-central Siberia. Can J for Res 41:1405–1421. https://doi.org/10.1139/x11-065
Hartmann FP, Rathgeber CBK, Fournier M, Moulia B (2017) Modelling wood formation and structure: power and limits of a morphogenetic gradient in controlling xylem cell proliferation and growth. Ann for Sci 74:14. https://doi.org/10.1007/s13595-016-0613-y
He M, Yang B, Shishov V, Rossi S, Bräuning A (2018a) Relationships between wood formation and cambium phenology on the Tibetan Plateau during 1960–2014. Forests. https://doi.org/10.3390/f9020086
He M, Yang B, Shishov V, Rossi S, Bräuning A (2018b) Projections for the changes in growing season length of tree-ring formation on the Tibetan Plateau based on CMIP5 model simulations. Int J Biometeorol 62:631–641
Houghton RA, Butman D, Bunn AG, Krankina ON, Schlesinger P, Stone TA (2007) Mapping Russian forest biomass with data from satellites and forest inventories. Environ Res Lett. https://doi.org/10.1088/1748-9326/2/4/045032
Huete A, Justice C, van Leeuwen W (1999) Modis vegetation index (MOD13): algorithm theoretical basis document, Version 3
Jeong SJ, Ho CH, Gim HJ, Brown ME (2011) Phenology shifts at start versus end of growing season in temperate vegetation over the Northern Hemisphere for the period 1982–2008. Glob Change Biol 17:2385–2399. https://doi.org/10.1111/j.1365-2486.2011.02397.x
Jevšenak J, Tychkov I, Gričar J, Levanič T, Tumajer J, Prislan P, Arnič D, Popkova M, Shishov VV (2020) Growth-limiting factors and climate response variability in Norway spruce (Picea abies L.) along an elevation and precipitation gradients in Slovenia. Int J Biometeorol. https://doi.org/10.1007/s00484-020-02033-5
Keenan TF, Gray J, Friedl MA, Toomey M, Bohrer G, Hollinger DY, Munger JW, O’Keefe J, Schmid HP, Wing IS, Yang B, Richardson AD (2014) Net carbon uptake has increased through warming-induced changes in temperate forest phenology. Nat Clim Change 4:598–604. https://doi.org/10.1038/nclimate2253
Kharuk V, Im S, Oskorbin P, Petrov I, Ranson K (2013a) Siberian pine decline and mortality in southern Siberian mountains. For Ecol Manag 310:312–320. https://doi.org/10.1016/j.foreco.2013.08.042
Kharuk VI, Ranson K, Oskorbin P, Im S, Dvinskaya M (2013b) Drought-caused forest decline in the Trans-Baikal Lake area. For Ecol Manag J 289:385–392. https://doi.org/10.1016/j.foreco.2012.10.024
Kharuk VI, Im ST, Petrov IA, Golyukov AS, Ranson KJ, Yagunov MN (2017) Climate-induced mortality of Siberian pine and fir in the Lake Baikal Watershed. Siberia for Ecol Manag 384:191–199. https://doi.org/10.1016/j.foreco.2016.10.050
Kirdyanov A, Hughes M, Vaganov EA, Schweingruber F, Silkin P (2003) The importance of early summer temperature and date of snow melt for tree growth in the Siberian Subarctic. Trees 17:61–69
Körner C, Basler D (2010) Phenology under global warming. Science 327:1461–1462
Kottek M, Grieser J, Beck C, Rudolf B, Rubel F (2006) World Map of the Köppen–Geiger climate classification updated. Meteorol Zeitschrif 15:259–263. https://doi.org/10.1127/0941-2948/2006/0130
Krankina ON, Harmon ME, Winjum JK (1996) Carbon storage and sequestration in the Russian forest sector. Ambio 25:284–288. https://doi.org/10.2307/4314475
Krankina ON, Dixon RK, Kirilenko AP, Kobak KI (1997) Global climate change adaptation: examples from Russian boreal forests. Clim Change 36:197–215. https://doi.org/10.1007/978-1-4613-8471-7_31
Menzel A, Sparks T, Estrella N, Koch E, Aasa A, Aha R, Alm-KKübler K, Bissolli P, Braslavská O, Briede A, Chmielewski F, Crepinsek Z, Curnel Y, Dahl A, Defila C, Donnellly A, Filella Y, Jatczak K, Måge F, Mestre A, Nordli Ø, Peñuelas J, Pirinen P, Remišová V, Scheifinger H, Striz M, Susnik A, Van Vlie A, Wielgolaski F-E, Zus A (2006) European phenological response to climate change matches the warming pattern. Glob Change Biol 12:1969–1976. https://doi.org/10.1111/j.1365-2486.2006.01193.x
Menzel A, Yuan Y, Matiu M, Sparks T, Scheifinger H, Gehrig R, Estrella N (2020) Climate change fingerprints in recent European plant phenology. Glob Chang Biol 26:2599–2612. https://doi.org/10.1111/gcb.15000
Myneni RB, Keelingt CD, Tucker CJ, Asrar G (1997) Increased plant growth in the northern high latitudes from 1981 to 1991. Nature 386:698–702
Nemani RR, Keeling CD, Hashimoto H, Jolly WM, Piper SC, Tucker C, Myneni RB, Running SW (2003) Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science 300:1560–1563. https://doi.org/10.1126/science.1082750
Peng S, Piao S, Ciais P, Fang J, Wang X (2010) Change in winter snow depth and its impacts on vegetation in China. Glob Chang Biol 16:3004–3013. https://doi.org/10.1111/j.1365-2486.2010.02210.x
Piao S, Tan J, Chen A, Fu YH, Ciais P, Liu Q, Janssens IA, Vicca S, Zeng Z, Jeong SJ, Li Y, Myneni RB, Peng S, Shen M, Peñuelas J (2015) Leaf onset in the northern hemisphere triggered by daytime temperature. Nat Commun. https://doi.org/10.1038/ncomms7911
Piao S, Liu Q, Chen A, Janssens IA, Fu Y, Dai J, Liu L, Lian X, Shen M, Zhu X (2019) Plant phenology and global climate change: current progresses and challenges. Glob Change Biol 25(6):1922–1940. https://doi.org/10.1111/gcb.14619
Reed B, Brown J, VanderZee D, Loveland T, Merchant J, Ohlen D (1994) Measuring phenological variability from satellite imagery. J Veg Sci 5:703–714
Safonova A, Tabik S, Alcaraz-Segura D, Rubtsov A, Maglinets Y, Herrera F (2019) Detection of fir trees (Abies sibirica) damaged by the bark beetle in unmanned aerial vehicle images with deep learning. Remote Sens. https://doi.org/10.3390/rs11060643
Schaaf CB, Gao F, Strahler AH, Lucht W, Li XW, Tsang T, Strugnell NC, Zhang X, Jin Y, Muller J-P, Lewis P, Barnsley M, Hobson P, Disney M, Roberts G, Dunderdale M, Doll C, Hu DRP, B, Liang S, Privette JL, Roy D, (2002) First operational BRDF, albedo nadir reflectance products from MODIS. Remote Sens Environ 83:135–148. https://doi.org/10.1016/S0034-4257(02)00091-3
Schwartz MD, Ahas R, Aasa A (2006) Onset of spring starting earlier across the Northern Hemisphere. Glob Chang Biol 12:343–351. https://doi.org/10.1111/j.1365-2486.2005.01097.x
Shah SK, Touchan R, Babushkina E, Shishov VV, Meko DM, Abramenko OV, Belokopytova LV, Hordo M, Jevsenak J, Kedziora W, Kostyakova TV, Moskwa A, Oleksiak Z, Omurova G, Ovchinnikov S, Sadeghpour M, Saikia A, Zsewastynowicz L, Sidenko T, Strantsov A, Tamkeviciute M, Tomusiak R, Tychkov I (2015) August to July precipitation from tree rings in the forest-steppe zone of Central Siberia (Russia). Tree-Ring Res 71:37–44. https://doi.org/10.3959/1536-1098-71.1.37
Shen M, Piao S, Cong N, Zhang G, Jassens IA (2015) Precipitation impacts on vegetation spring phenology on the Tibetan Plateau. Glob Chang Biol 21:3647–3656. https://doi.org/10.1111/gcb.12961
Shiklomanov AI, Lammers RB, Lettenmaier DP, Polischuk YM, Savichev OG, Smith LC, Chernokulsky AV (2013) Hydrological changes: historical analysis, contemporary status, and future projections. In: Groisman P, Gutman G (eds) Regional environmental changes in siberia and their global consequences. Springer Environmental Science and Engineering. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-4569-8_4
Shishov VV, Tychkov II, Popkova MI, Ilyin VA, Bryukhanova MV, Kirdyanov AV (2016) VS-oscilloscope: a new tool to parameterize tree radial growth based on climate conditions. Dendrochronologia 39:42–50. https://doi.org/10.1016/j.dendro.2015.10.001
Shvidenko AZ, Gustafson E, Mcguire AD, Kharuk VI, Schepaschenko DG, Shugart HH, Tchebakova NM, Vygodskaya NN, Onuchin AA, Hayes DJ, Mccallum I, Maksyutov S, Mukhortova LV, Soja AJ, Belelli-marchesini L, Kurbatova JA, Oltchev AV, Parfenova EI, Shuman JK (2012) Regional environmental changes in siberia and their global consequences. In: Groisman P, Gutman G (Eds) Regional environmental changes in Siberia and Their global consequences. Springer Science + Business Media, pp. 171–249. https://doi.org/10.1007/978-94-007-4569-8
Soja AJ, Tchebakova NM, French NHF, Flannigan MD, Shugar HH, Stocks BJ, Sukhinin AI, Parfenova EI, Chapin FS, Stackhouse PW (2007) Climate-induced boreal forest change: Predictions versus current observations. Glob Planet Change 56:274–296. https://doi.org/10.1016/j.gloplacha.2006.07.028
Spinoni J, Vogt J, Barbosa P (2015) European degree-day climatologies and trends for the period 1951–2011. Int J Climatol 35:25–36. https://doi.org/10.1002/joc.3959
Tabakova M, Arzac A, Martínez E, Kirdyanov A (2020) Climatic factors controlling Pinus sylvestris radial growth along a transect of increasing continentality in southern Siberia. Dendrochronologia. https://doi.org/10.1016/j.dendro.2020.125709
Tchebakova NM, Parfenova EI, Korets MA, Conard SG (2016) Potential change in forest types and stand heights in central Siberia in a warming climate. Environ Res Lett 11:35016. https://doi.org/10.1088/1748-9326/11/3/035016
Thornthwaite DW (1948) An approach toward a rational classification of climate. Geogr Rev 38:55–94. https://doi.org/10.2307/210739
Trumbore S, Brando P, Hartmann H (2015) Forest health and global change. Science 349:814–818. https://doi.org/10.1126/science.aac6759
Tumajer J, Kašpar J, Kuželová H, Shishov VV, Tychkov II, Popkova MI, Vaganov EA, Treml V (2021) Forward modeling reveals multidecadal trends in cambial kinetics and phenology at treeline. Front Platn Sci 12:32. https://doi.org/10.3389/fpls.2021.613643
Tychkov I, Sviderskaya IV, Babushkina EA, Popkova MI, Vaganov EA, Shishov VV (2019) How can the parameterization of a process-based model help us understand real tree-ring growth? Trees 33:345–357. https://doi.org/10.1007/s00468-018-1780-2
Vaganov EA, Hughes MK, Shashkin AV (2006) Growth dynamics of conifer tree rings Images of past and future environments. Springer-Verlag, Berlin Heidelberg. https://doi.org/10.1017/CBO9781107415324.004
Wu C, Gonsamo A, Gough CM, Chen JM, Xu S (2014) Modeling growing season phenology in North American forests using seasonal mean vegetation indices from MODIS. Remote Sens Environ 147:79–88. https://doi.org/10.1016/j.rse.2014.03.001
Yang B, He M, Shishov V, Tychkov I, Vaganov E, Rossi S (2017) New perspective on spring vegetation phenology and global climate change based on Tibetan Plateau tree-ring data. Proc Natl Acad Sci 114:6966–6971. https://doi.org/10.1073/pnas.1616608114
Zeng Q, Rossi S, Yang B, Qin C, Li G (2020) Environmental drivers for cambial reactivation of Qilian junipers (Juniperus przewalskii) in a semi-arid region of northwestern China. Atmpsphere 11:1–312. https://doi.org/10.3390/atmos11030232
Zhang X, Friedl MA, Schaaf CB, Strahler AH (2004) Climate controls on vegetation phenological patterns in northern mid- and high latitudes inferred from MODIS data. Glob Change Biol 10:1133–1145. https://doi.org/10.1111/j.1365-2486.2004.00784.x
Zhang X, Friedl MA, Schaaf CB (2006) Global vegetation phenology from moderate resolution imaging Spectroradiometer (MODIS): evaluation of global patterns and comparison with in situ measurements. J Geophys Res Biogeosci. https://doi.org/10.1029/2006JG000217
Zhang Y, Williams AP, Gentine P (2020) Light limitation regulates the response of autumn terrestrial carbon uptake to warming. Nat Clim Change 15:115. https://doi.org/10.1038/s41558-020-0806-0
Zhou L, Kaufmann RK, Tian Y, Myneni RB, Tucker CJ (2003) Relation between interannual variations in satellite measures of northern forest greenness and climate between 1982 and 1999. J Geophys Res D Atmos. https://doi.org/10.1029/2002jd002510
Zhu Z, Piao S, Myneni RB, Huang M, Zeng Z, Canadell JG, Ciais P, Sitch S, Friedlingstein P, Arneth A, Cao C, Cheng L, Kato E, Koven C, Li Y, Lian X, Liu Y, Liu R, Mao J, Pan Y, Peng S, Peuelas J, Poulter B, Pugh TAM, Stocker BD, Viovy N, Wang X, Wang Y, Xiao Z, Yang H, Zaehle S, Zeng N (2016) Greening of the earth and its drivers. Nat Clim Change 6:791–795. https://doi.org/10.1038/nclimate3004
Acknowledgements
This work was carried out with the support of Ministry of Science and Higher Education of the Russian Federation [FSRZ-2020-0014] and the Russian Science Foundation [Grant 18-74-10048]. We are indebted with E. Martínez for his assistance on field and samples preparation and with the two National Parks and research station for allowing access to their territory. A previous version was revised by JM Olano. We thank to the anonymous reviewers for their comments and suggestions, which significantly improved the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflicts of interest
The authors have no conflicts of interest.
Additional information
Communicated by Peter Annighoefer.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Arzac, A., Tychkov, I., Rubtsov, A. et al. Phenological shifts compensate warming-induced drought stress in southern Siberian Scots pines. Eur J Forest Res 140, 1487–1498 (2021). https://doi.org/10.1007/s10342-021-01412-w
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-021-01412-w