
Abstract
Forests exhibit strategies to cope with climate change; however, the rate of the changes on forests can be slower than the actual changes in environmental conditions. Forest management policies, such as assisted migration, may help forests to adapt their species distribution to changing climate conditions. Nonetheless, it certainly requires a better knowledge of climate influences on trees to ensure the success of specific management actions. In this study, we apply dendroclimatological methods to investigate the growth response of the main forest species present in Moncayo Natural Park to climate to assess their current relationship and to model these responses over the potential distribution of each species across the study area. Our results revealed large differences in the response of beech, pine and Pyrenean oak to prevailing climate factors and indicated species-specific patterns of climate sensitivity. The general importance of summer conditions for tree growth was confirmed. In addition, we found directional trends in correlation with specific climate factors along spatial gradients; these results are consistent with the autoecology of the studied species. Based on these findings, we present a new model approach that can serve as a key tool for forest managers to design forest communities that are more stable during climatic change.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Climate is one of the main abiotic factors that defines and limits the natural distribution of tree species (Mott 2010). Therefore, it is expected that predicted climate change (IPCC 2013) will have a significant impact on the distribution of species (Pearson and Dawson 2003).
Tree species often exhibit physiological adaptations along climatic gradients, adapting to the specific climatic and site conditions (Sáenz-Romero et al. 2016). Alternatively, species can migrate in response to climate changes. However, both functional adaptation and migration are slow processes for long-lived organisms, such as trees, and likely do not keep pace with the actual change in environmental conditions (Sáenz-Romero et al. 2016).
Management of natural protected areas and commercial forests requires long-term strategies and practical guidelines to facilitate management decisions in response to climate change (Nabuurs et al. 2017; Walentowski et al. 2017; Loran et al. 2018). Assisted migration is an important tool to adjust species distributions to changing climate conditions and mitigate its effects. However, implementing assisted migration requires knowledge of climate–growth relationships (CGRs) within a species’ range to ensure the success of the reforestation plan.
Tree-ring research is a powerful tool to assess the dynamics of climate–growth relationships, including reactions to extreme climate events (Fritts 1972) throughout long time periods. Using a tree-centered approach opens the possibility to understand tree function under specific climate and site conditions (Sass-Klaassen et al. 2016). This comes with the limitation that the gained information represents individual tree populations growing under specific site conditions.
There are numerous studies demonstrating that CGRs can strongly vary along altitudinal (Ponocná et al. 2016; Kharal et al. 2017) or environmental gradients (Čufar et al. 2014; Kraus et al. 2016; Martínez del Castillo et al. 2018b). This suggests a different ability of species to cope with climate variability and change across their distribution area. De Luis et al. (2013) presented an extensive tree-ring network for Aleppo pine (Pinus halepensis Mill.) that was used to assess CGRs and trends in climate sensitivity across the distribution area. Climate–growth relationships can be systematically assessed for species along gradients through the distribution area of the species to systematically extend this site-specific information.
It is difficult to obtain strong directional trends in climate response across the distribution range because of the diversity in environments encountered across the distribution area of a given species. As a consequence, there is a lack of information about how tree species respond to climate conditions outside of their distribution range and within the range (i.e., in areas of recent expansion, mixed stands or juvenile trees). Interestingly, De Luis et al. (2013) demonstrated that such variation in CGRs are related to prevailing climate conditions which allow for extrapolation of estimated trends across the species distribution and also into areas where the species is currently not present. Such information is useful for foresters that seek to perform assisted-migration actions, specifically into sensitive areas, i.e., at the edge of the current distribution area.
Moncayo Natural Park (MNP) is a mountain ecotone in a transitional zone between the Eurosiberian and Mediterranean biogeographical region (Longares Aladrén 2004) and forms a geographical boundary for several tree species. The ecotone areas are transitional areas between two neighboring biomes that contain different vegetation types and generally are more sensitive to environmental disturbances (Risser 1995). MNP provides a relevant sample area with a number of different sites that span different climatic types. Therefore, this area is a suitable location to apply new analysis techniques aimed to spatially test the influence of climate on forest species.
European beech (Fagus sylvatica L.), Scots pine (Pinus sylvestris L.), and Mountain pine (Pinus uncinata Ram.) are widespread European tree species with high economic and ecological value. The forests stands in Moncayo Natural Park represent one of the southern limits for all these species. Pyrenean oak (Quercus pyrenaica Willd.) distribution in Europe is limited to the Atlantic climate areas of western France and the Iberian Peninsula (Nieto Quintano et al. 2016); MNP is the northern extent of its Mediterranean range. The current distribution of these species in this Natural Park are influenced by historical land use, such as sheep and goat grazing and pine introduction via reforestation (Arrechea 2002). Graphical records (e.g., old photographs) and aerial photography indicate that the currently forested area was a meadow with isolated patches of beech and Pyrenean oak trees about one hundred years ago. The closed-canopy forest is relatively young, and the current situation does not reflect the potential spatial distribution of the species across this site. Recently, several changes in forest size and structure have been observed due to the abandonment of traditional uses triggered by protection of the region as a natural park (Martínez del Castillo et al. 2015). Specifically, beech and Pyrenean oak trees are protected by the Habitats Directive (Council Directive 92/43/EEC).
In this study, we investigated the growth response of beech, Scots pine, Mountain pine and Pyrenean oak to climate across an altitudinal-climatic gradient, and we modeled these responses over the potential distribution of each species inside the Natural Park. The following were our hypotheses: (i) the species exhibit different climate–growth relationships, which vary along the climatic gradient; (ii) variation in climate–growth relationships for each species across the altitudinal gradient is associated with the specific climatic variability; and (iii) climate–growth relationships models can be applied beyond the current distribution of each species to allocate potential migration areas (either natural or artificial via reforestation). Our final aim was to predict CGRs across the potential species distribution in MNP to investigate the use of this novel approach for the design of forest management activities geared to adapt species distributions to changing climate conditions. The obtained empirical model is based on the specific local conditions at MNP and hence may not be widely applicable. However, this new modeling approach illustrates the value of building models for specific areas that can be applied by foresters to manage the distribution of species. Assuming that tree secondary growth can be used as an appropriate indicator of tree performance related with the environmental conditions, the interpretation of this results can provide insights to design management actions geared to create stable forest communities that are able to cope with stress in changing (and more severe) climatic conditions.
Methods
Study area
Moncayo Natural Park is a mountain area located in the Iberian range, in the northeast of the Iberian Peninsula (Fig. 1). This Natural Park covers 11,226 ha2, and the elevation ranges from 850 to 2314 m above sea level. It is located in a transitional climate area between the Mediterranean and Eurosiberian biogeographical regions. This mountain area is characterized as a hotspot of biodiversity, with numerous tree species and types of vegetation within a relatively small area (Longares Aladrén 2004). The main forests of the park are formed by European beech (Fagus sylvatica L.), Scots pine (Pinus sylvestris L.) and Pyrenean oak (Quercus pyrenaica Willd.), but there are other tree species present, such as Holm oak (Quercus ilex L.), Sessile oak (Quercus petraea (Matt.) Liebl.), Mountain pine (Pinus uncinata Ram.), European black pine (Pinus nigra Arnold.) and birch (Betula pendula Roth.). The closed-canopy forests are located in the northern half of the park; the southern area is dominated by shrubs, open-canopy Quercus ilex forests and small stands of other species.

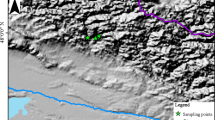
Study area with current distributions (polygons) of study species and sample sites (circles) from which tree-ring chronologies were sampled inside and outside of Moncayo Natural Park
Climate
The Natural Park is located in the eastern side of the Moncayo massif. Its geographical position represents an orographic barrier where the western and northwestern fronts prevent moisture from reaching the mid Ebro Valley. This situation produces a disruption of rising moist air flow that contributes to relatively frequent convective activity and results in higher amounts of precipitation at high elevations with a progressive decrease toward the lower elevations, whereas temperature follows the inverse path, with lower values at high elevations and higher values at low elevations.
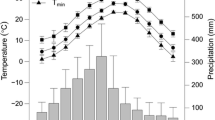
With the aim to analyze the climate of the Moncayo Natural Park, three high-resolution gridded datasets of daily precipitation and maximum and minimum temperature were created using the reddPrec R package (Serrano-Notivoli et al. 2017b) based on the reconstructed stations of Serrano-Notivoli et al. (2017a). The grids were based on a set of 2801 points regularly distributed over the study area with a spatial resolution of 200 m. Precipitation and temperature were computed daily for the period from 1950 to 2012. The annual averages for both precipitation (Fig. 2b) and temperature (Fig. 2c, d) show a clear altitudinal spatial distribution. The annual amounts of precipitation reach 1500 mm at high elevations and decrease until values drop below 400 mm at lower elevations in the eastern and southern margins (Fig. 2b). The temperatures widely vary from the mean annual minimum below 0 °C at the summit to the mean maximum over 16 °C at the lower areas (Fig. 3). Annual temperature values show a similar pattern along the altitudinal bands with a change of − 0.517 and − 0.523 °C/100 m increase in altitude for minimum and maximum temperatures, respectively, whereas annual precipitation increased by 119.89 mm/100 m increase in altitude.

Geographic and climatic gradients across study area for a elevation, b mean annual precipitation, c mean annual minimum and d mean annual maximum temperature

Climatic diagrams at Moncayo Natural Park showing monthly precipitation (PCP), maximum temperatures (T.Max) and minimum temperatures (T.min) for a lowest elevation 850 m.a.s.l. and b highest elevation 2314 m.a.s.l
Tree-ring chronologies
A total of 40 sites were selected and sampled between 2010 and 2014 along an altitudinal gradient from 950 to 1900 m.a.s.l. (Table 1, Table S1 in Supplementary Materials). Between five and 25 healthy and dominant trees were sampled at each site with an increment borer at breast height, extracting two cores per tree. Tree cores were mounted on wooden supports, dried, sanded and scanned in the laboratory. The tree rings were measured with the TSAP-Win program and LINTAB™ 5 measuring device (Rinntech, Heidelberg, Germany) with 0.01 mm precision. Crossdating was done using CooRecorder v8.0 software (Cybis Elektronik & Data AB. Saltsjöbaden, Sweden).
Ring-width measurements were detrended to remove biological trends using a negative exponential curve. A second detrending was then applied using a 50-year cubic smoothing spline with a 50% frequency response to filter out low frequency variation that reflects potential non-climatic disturbances. To obtain the detrended residual chronology to perform the analysis, two processes were done: an autoregressive modeling of the residuals and a bi-weight robust estimation of the mean. The detrending process and chronology computation was carried out using the dplR package (Bunn 2008).
Statistical analysis
Correlation function coefficients (CFCs) were calculated between all 40 residual chronologies through seasonal 3-month means for respective maximum and minimum temperatures and precipitation sums from the previous summer (June, July and August) and the current summer. Correlation analysis was performed separately for each chronology with the associated climate calculated for each location.
As a second step, the variability and patterns of distribution of obtained seasonal CFCs for temperature and precipitation were explored. Generalized additive modeling (GAM) was used to detect relationships between the distribution patterns of obtained CFCs (dependent variable) and the mean annual values for precipitation and maximum and minimum temperature calculated for each study site (independent variables). This study is species-specific, but P. sylvestris and P. uncinata were grouped together due to similarities in climate–growth relationships found in this area between this species when compared with the other analyzed species. However, this similar behavior should not be extrapolated to other locations where both species are present.
The GAMs were constructed using the ‘gam’ package in R environment (Hastie and Tibshirani 1986). The quasibinomial family was used in order to describe the error distribution (Eq. 1). CFC values can theoretically vary from − 1 to 1; therefore, observed values were re-scaled (CFCsb) for model construction (Eq. 2).
The accuracy of the models was evaluated using a likelihood ratio test by comparing the obtained models (full models) with restricted models where the explanatory variables of interest were omitted and only the intercept term was included (null models). The p values for the likelihood ratio tests that compared the full and reduced models were calculated using the Chi square distribution.
Finally, the obtained models were applied to the current species distribution in the MNP and also for the area of the Natural Park designated by a specific altitudinal range for each species. The boundaries of this altitudinal range are limited by the highest and lowest presence of a given species inside the Natural Park (from 1100 to 1750 m for beech; from 950 to 2100 m for pines and from 900 to 1500 m for Pyrenean oak). These areas are considered as potential distribution areas for the study species as they contain the specific environmental conditions required for their survival. The climatic conditions inside the different altitudinal gradients defined for each species are gathered in the sampled zones; therefore, the application of the models to these specific climatic conditions is not an extrapolation outside the values domain on which the models are constructed. This approach is similar to the fundamental niche concept, where a combination of environmental variables defines the limits for survival without taking into account biotic competition (Reed and Clark 1978).
Results
Chronologies
In total 427 trees were sampled in the area, covering an altitudinal range from 950 to 1900 m.a.s.l. (Table S1 in supplementary material). The youngest and oldest trees sampled for each species were 71 and 215 years for beech, 41 and 95 years for pine, and 37 and 271 years for Pyrenean oak. In total 40 chronologies where constructed, 15 for beech, 11 for pines and 14 for Pyrenean oak. The statistical analysis indicated a high quality of all chronologies based on the four commonly used statistical indicators in dendrochronology: the mean correlation between ring-width series (Rbar), the expressed population signal (EPS), the signal to noise ratio (SNR) and the mean sensitivity (SENS) (Table S1). Rbar values ranged from 0.21 to 0.55, with a mean of 0.42. SNR ranged from 1.33 to 28.20, with a mean of 12.41. SENS ranged from 0.2 to 0.46, with a mean value of 0.3. All EPS values reached the threshold of 0.85, indicating that all chronologies reflect a high common environmental signal of the sampled tree populations.
Climate–growth analysis
The results of the correlation analyses between the 40 chronologies and the climate variables (precipitation, maximum and minimum temperature grouped by seasons) by species are summarized in Fig. 4. The boxplots represent the variability of the correlation coefficients of all sampled sites of each species, therefore, across the studied populations (Fig. 4). If the box is completely beyond the dashed line (e.g., CFC for beech regarding T.max in previous summer), it implies that in all chronologies existed a significant relationship with the climate variable (in this case negative). If the box is completely inside the range described by the dashed line (e.g., CFC for beech regarding T.max in previous autumn), it implies that all chronologies do not showed a significant relationship with that climatic variable. If the box cross the dashed line (e.g., CFC for beech regarding T.max in winter), it implies that at least one chronology presented a significant relationship (in this case, one positive and another negative, but in most of the cases the significance threshold was not reached). Lastly, the dashed line indicates statistical significance (P < 0.05) beyond 0.25 or − 0.25, threshold given by the number of years included in the analysis (i.e., 62).

Climate–growth relationships for chronologies of beech (n = 15), pines (n = 11) and Pyrenean oak (n = 14); dashed horizontal lines indicate statistical significance (P < 0.05) for 62 years of record; PCP = seasonal precipitation, TMAX, TMIN = seasonal maximum and minimum temperature, respectively. pSUM = previous summer, pAUT = previous autumn, WIN = winter, SPR = spring, SUM = summer. The central lines of boxplots indicate the median value, vertical hinges indicate first and third quartiles, error bars indicate the 95% confidence interval of the median and dots indicate outlines, values beyond the 95% confidence interval threshold
Beech growth was significantly related to wet and cold conditions during the summer preceding the current growing season. Precipitation had a positive and significant effect in all but one case, meanwhile the maximum temperature affected growth negatively (CFCs from − 0.26 to − 0.47). Large variation in correlation coefficients for current summer (precipitation and maximum temperature) and especially winter conditions (precipitation and minimum temperature) determine different responses among the 15 beech populations.
The growth of the pine populations was mainly driven by precipitation and maximum temperature during summer in the current growing season with above-average rainfall and below-average maximum temperature significantly favoring pine growth, whereas during spring, high temperatures had a significant positive influence at the majority of the pine sites. Large variation between pine populations occurred in correlations with spring precipitation and maximum temperature during the previous summer.
Pyrenean oaks were especially sensitive to precipitation. There was a positive influence of precipitation during all growing periods, even from the previous autumn. There was a general negative influence of summer maximum temperatures, significant at some sites, and a positive relationship between previous autumn minimum temperatures and Pyrenean oak growth. In all cases, minimum temperatures had less influence on growth than precipitation or maximum temperatures. Generally, the Pyrenean oak populations vary more in their climate response than those of beech and pine as indicated by the large size of the boxplots, although only the median correlation with winter precipitation was significant.
The correlation analyses indicate large differences between the average response of beech, pines and Pyrenean oak to prevailing climatic factors and, moreover, contrasting responses within species to climate conditions across the environmental gradient of the Moncayo Natural Park.
Model application
To explore the variability of the sets of correlation coefficients yielded for the study species along the climate gradients for mean annual precipitation and the mean of the annual maximum and minimum temperatures within the study area, a total of 45 GAMs were constructed (5 seasons * 3 climate variables * 3 genera) and applied to the current species distribution in the MNP and also for the area of the Natural park designated by a specific altitudinal range for each species (considered as potential distribution areas). Predicted correlation coefficients for all analyzed seasonal climatic factors are shown in Figs. 5 (beech), 6 (pines) and 7 (oak). The reliability and validity of the models were evaluated based on the deviance and the significance level (Table 2). Of the 45 models, 22 were statistically significant.

Beech predicted correlation coefficients for the previous summer (pSUM) and current summer (SUM) precipitation (PCP) and maximum temperature (T.MAX) across the MNP potential distribution area

Pine (P. sylvestris and P. uncinata): predicted correlation coefficients for spring (SPR) and summer (SUM) precipitation (PCP) and maximum temperature (T.MAX) across the MNP potential distribution area

Oak: predicted correlation coefficients for winter (WIN), spring (SPR) and summer (SUM) precipitation (PCP) and summer maximum temperature (T.MAX) across the MNP potential distribution area
In beech, previous summer precipitation influence was positive and the maximum temperatures played a negative role across the potential distribution area (Fig. 5). The variations of CFCs associated with these climatic variables are not explained by the existing climatic gradients in the territory; therefore, the models are not significant (Fig. 2). Current summer conditions had a distinct influence across the territory, shifting from positive to negative depending on altitude. The effect of precipitation for the current summer was positive in warm, dry, low-altitude areas, whereas it was negative at high altitudes where conditions were colder and wetter. High temperatures during summer affected growth positively at high altitudes and negatively at lower altitudes. Variations in CFCs during summer can be explained by climatic variability across the study areas as shown in Fig. 2.
Pine growth was influenced mostly by spring and summer conditions during the current growing season (Fig. 6). Precipitation in spring increased growth in low elevation zones, whereas in summer it was important all over the territory, especially in low elevation zones which are dryer. Maximum temperatures in spring increased growth in high elevation zones but in summer it became a growth-limiting factor, particularly in lower zones. In this case, CFCs significantly varied across climate gradients (Table 2).
The amount of precipitation boosted oak growth during the entire year of the growing season, although with different intensity (Fig. 7). In general, the relationship between precipitation and growth was stronger in low elevation zones during spring and summer (Table 2). The occurrence of high temperatures during summer limited tree growth generally, with small variations across the study area.
Discussion
Climatic influence on tree growth
In Moncayo Natural Park, the studied trees (beech, pines and oaks) showed different patterns of climate sensitivity. Similar observations in climate–growth relationships at different elevations in mountain areas have also been detected in other studies (i.e., Hartl-Meier et al. 2014; Kharal et al. 2017; Ponocná et al. 2016; Wang et al. 2017). Moreover, there are some common elements for all the species, such as the general importance of summer conditions for tree growth and the variation in correlation with specific climate factors along spatial gradients.
Beech growth has proven to be sensitive to drought at numerous sites (Weemstra et al. 2013; Tegel et al. 2014; Cavin and Jump 2016; Farahat and Linderholm 2018) and also in southern marginal stands in Spain (Rozas et al. 2015; Dorado-Liñán et al. 2017). In this study we found that wet and cold conditions during the previous summer are favorable for beech growth whereas the influence of current summer conditions varied significantly along the spatial climatic gradient. At lower elevation sites, beech growth is negatively influenced by high temperatures and low precipitation during summer, which supports results of other studies (Chen et al. 2015; Zimmermann et al. 2015) and can be explained by the species’ strategy of rapid stomata closure to prevent the risk of embolism and inactivation of the water transporting system (Aranda et al. 2000; Granier et al. 2007). Conversely, at higher elevations, growth is enhanced when summer conditions are warmer. Trees may benefit from higher radiation on summer, that promotes photosynthesis in higher and colder conditions, that compensates water stress and embolism on beech trees. Čufar et al. (2008) described a negative influence of August maximum temperatures on beech growth at higher elevation sites in Slovenia. This might be due to a prolonged growing season for the Slovenian beeches (until August) as reported by Prislan et al. (2013) whereas at our study site the growing period is rather short, lasting for 67 days at lower elevations and 80 days at higher elevations, starting in May or early June and ending by late-July or early-August (Martínez del Castillo et al. 2016). Therefore, beech may already be close to dormancy during the late summer. The influence of winter precipitation on beech growth differs along the altitudinal gradient; it is negative in high elevations and positive at low elevation sites.
The growth of pines was mainly favored by wet and cold conditions during the current summer. In cold environments, high maximum temperatures during spring promoted an early start to the growing season which leads to an extended growing season that favored tree growth. Earlier cambium re-activation in pines is shown by heating experiments (Gričar et al. 2007). In summer, this relationship changed and pine growth was limited by high maximum temperature, which supports the findings of other studies (e.g., Martínez del Castillo et al. 2018b; Cailleret and Davi 2011; Di Filippo et al. 2007) and likely related to increased drought stress caused by high evaporation rates. Pines are known to exhibit a plastic growth behavior which is indicated by their ability to anticipate favorable spring conditions and start growing early and also by re-activation of their growth after cessation from dry conditions during summer (de Luis et al. 2007; Camarero et al. 2010; De Luis et al. 2011; Novak et al. 2016; Martínez del Castillo et al. 2018a). Furthermore, at southern and/or low elevation sites with mild temperatures during winter, pines may not enter in a real dormancy period (Gričar et al. 2016; Prislan et al. 2016).
In the case of oak, water availability is largely determined by the amount of precipitation, which triggers tree growth in lower and dryer areas. Although oaks are known to be able to recover water potential due to a deeper and extensive root system when compared with beech (Aranda et al. 2000), a severe and prolonged water deficit leads to an irreversible cavitation of earlywood vessels with tylose formation (Pérez-de-Lis et al. 2018). Several studies identified the positive effect of spring/summer precipitation on the growth of ring-porous species (García-Suárez et al. 2009; García-González and Souto-Herrero 2017). Similarly, high maximum temperatures during summer limit oak growth across sites in Central Europe (Čufar et al. 2014; Mérian et al. 2011).
Spatial distribution of correlations between climate and growth
As hypothesized, distribution patterns for correlations between growth of tree species and prevailing climate factors are at least partially, related to the existing climate gradients. The significance of the models proved that these influences of the seasonal climatic variables are explicable by the climate variability over the territory. This fact is most noticeable in beech, with 11 significant models out of 15, somewhat in pines, with 7 significant models out of 15 and less evident in oak, with only 4 significant models out of 15. Climate–growth relationships for the three species were tested with GAMs to create models which allow projection of the influence of climate on the growth of selected species along climate gradients within the study area.
Although the variance of some variables was largely unexplained and statistical significance was lacking (i.e., previous summer precipitation in beech), information obtained from the GAMs is still valuable. On one hand, significance models suggest the existence of a plastic climate–growth relationship for the species across the studied distribution area. Such plasticity indicates that the influence of a given climate factor is climatically determined and would probably be modified if changes in climate occur. This is key for the development of accurate predictions of species behavior across a study area under different climate change scenarios. Another fact is that the lack of significant models suggests that the influence of a given seasonal climate factor is constant across the study area (or at least not affected by prevailing climate conditions). The influence of such a seasonal climate factor is then expected to be more stable if changes in climate occur (at least if such changes do not exceed the actual climate range within the study site).
Applicability of the model approach in Moncayo Natural Park
Over the last several decades, forest management actions performed in MNP aimed at conserving and improving forested habitats (Martínez del Castillo et al. 2015). Specifically, the objectives for beech and Pyrenean oak forests are to improve their natural regeneration and to improve the quality and status of conservation, since these species are protected by the Habitats Directive. Conversely, pines were introduced in the park as reforestations and they are not protected by the Directive.
Or results showed that beech forest may be favored in the middle of the actual altitudinal range (1400–1500 m.a.s.l), and progressively extend to southern locations in the center of the Park where the species is currently not present. The climatic conditions of this new area are similar to the climatic conditions where beech forest is currently growing, and our models forecasted similar climate–growth relationships for the most important climatic variables influencing beech growth. However, it could not spread in the lowlands of this mountain due to higher temperatures and water scarcity.
The specific management actions in MNP for pines are aimed to naturalize the structure of the forest and reduce density by low-impact logging. The pine forest models showed that forest could be extended to high zones in the center part of the Park, which is partially unforested, given the similar climatic conditions of that area and the models results. By spreading pine forest cover to other areas, with logging actions the current pine forest at medium altitudes could be progressively replaced by protected species, in this case beech or Pyrenean oak, avoiding at the same time pine loss at MNP.
Pyrenean oak forests are affected by drought especially in low zones; therefore, logging actions to reduce tree density during dry years could compensate for growth reduction by decreasing competition. In addition, Pyrenean oak forests in Mediterranean areas largely have over-aged coppice stands where growth is reduced and trees are more sensitive to summer drought leading to oak-decline (Corcuera et al. 2006). Therefore, management actions to create uneven aged stands, as the proposed logging, may become necessary. Our results showed a positive reaction to water availability, consequently, these actions may not be necessary in rainy years, but could be very helpful during dry years.
Finally, the models suggest that the lower elevations in the southern part of the Park, in general, are not suitable for these tree species due to slightly different climate conditions.
Potential use of the model approach as a forest management tool
The climate–growth models, as presented in this study, provide a novel and complementary approach to forest-growth models or species distribution models, which are important tools for forest management (Porté and Bartelink 2002; Wullschleger et al. 2014). However, those models are based on data mining or regression techniques and hence largely lack ecophysiological explanations (Walentowski et al. 2017). Although the present and future distribution of tree species is a complex combination of multiple biotic and abiotic factors, to date, models based on climate provide the best available guide for policy making (Pearson et al. 2004; Booth 2017). Moreover, our models are applied to a simple version of the fundamental niche for each species, unlike most of the species distribution models, that are based on current distribution without taking into account information on species climatic requirements beyond their potential distributions (Booth 2017).
The models are empirical, and therefore, the forecasted CFCs are applicable to the environmental domain from where the samples were taken. In this sense, the model interpolates the results within the environmental gradient (climatic-altitudinal gradient in this case) of the sampled area. In addition, both CFCs analysis and the modeling were carried out in a specific range of climatic variability; therefore, the results of our models are not able to be extrapolated outside this range. The extrapolation of the results outside the range (i.e., other locations of this forests species over Europe) would include unsolvable uncertainties that would produce unreliable results (National Research Council 2012). Nevertheless, the applicability of our new methodological approach is universal and may be applicable to any other environment. However, application of the approach requires a systematic sampling strategy to ensure that the application of the models do not exceed the range of values used to construct the models.
The same methodological approach can be applied to predict changes in climate–growth relationships under future climate change scenarios. This requires input of a tree-ring network and high-resolution climate data, in addition to high-resolution future climate projections. Future climate conditions can exceed the range of the current climate variability; therefore, the models would have to extrapolate the results which might not be realistic. A methodological effort should be done in the future to address these topics, due to the importance of forest performance in future climates.
Forecasting future distribution of species is a complex task. The plastic response of species to climate variability is a key factor but it is not sufficiently incorporated into current modeling approaches. Complementary approaches, as presented here, can improve these models, but dense tree-ring networks are required.
Conclusions
The study on the growth response of beech, Scots pine, Mountain pine and Pyrenean oak in Moncayo Natural Park yielded information on spatial changes in climate–growth relationships across the altitudinal gradient. This was mainly related to different climatic conditions; therefore, the altitude of the sampling needs to be considered in dendroclimatological studies performed in the edge distribution limit of the species or in areas with large elevation gradient.
The study presents the current relationship between tree species and climate within their realized elevation range as well as the design and application of tree-ring based models to predict performance of the species across the potential distribution areas in the Park. The model results indicate that the total forested area could be expanded with additional management actions, mainly to higher altitudes and in a central zone of the Park which is currently unforested.
Forest management policy needs better tools to manage the distribution of tree species on afforested areas according to current and expected changing climatic conditions. The newly developed modeling approach can form a key tool to support concrete forestry decisions about species selection and estimation of species performance in specific areas for which it is calibrated. Apart from their direct application, the models showed a potential for direct integration into decision-making systems in a sustainable multifunctional forest management environment. The application of the actions derived from the models would enhance sustainable forest management and could help to mitigate climate change effects.
References
Aranda I, Gil L, Pardos JA (2000) Water relations and gas exchange in Fagus sylvatica L. and Quercus petraea (Mattuschka) Liebl. in a mixed stand at their southern limit of distribution in Europe. Trees Struct Funct 14:344–352. https://doi.org/10.1007/s004680050229
Arrechea E (2002) La gestión forestal en los espacios naturales protegidos: el ejemplo del Parque Natural del Moncayo. Rev Ecosistemas 11(2)
Booth TH (2017) Assessing species climatic requirements beyond the realized niche: some lessons mainly from tree species distribution modelling. Clim Change 145:1–13. https://doi.org/10.1007/s10584-017-2107-9
Bunn AG (2008) A dendrochronology program library in R (dplR). Dendrochronologia 26:115–124. https://doi.org/10.1016/j.dendro.2008.01.002
Cailleret M, Davi H (2011) Effects of climate on diameter growth of co-occurring Fagus sylvatica and Abies alba along an altitudinal gradient. Trees Struct Funct 25:265–276. https://doi.org/10.1007/s00468-010-0503-0
Camarero JJ, Olano JM, Parras A (2010) Plastic bimodal xylogenesis in conifers from continental Mediterranean climates. New Phytol 185:471–480. https://doi.org/10.1111/j.1469-8137.2009.03073.x
Cavin L, Jump AS (2016) Highest drought sensitivity and lowest resistance to growth suppression are found in the range core of the tree Fagus sylvatica L. not the equatorial range edge. Glob Change Biol 23:1–18. https://doi.org/10.1111/gcb.13366
Chen K, Dorado-Liñán I, Akhmetzyanov L et al (2015) Influence of climate drivers and the North Atlantic Oscillation on beech growth at marginal sites across the Mediterranean. Clim Res 66:229–242. https://doi.org/10.3354/cr01345
Corcuera L, Camarero JJ, Sisó S, Gil-Pelegrín E (2006) Radial-growth and wood-anatomical changes in overaged Quercus pyrenaica coppice stands: functional responses in a new Mediterranean landscape. Trees Struct Funct 20:91–98. https://doi.org/10.1007/s00468-005-0016-4
Čufar K, Prislan P, De Luis M, Gričar J (2008) Tree-ring variation, wood formation and phenology of beech (Fagus sylvatica) from a representative site in Slovenia, SE Central Europe. Trees Struct Funct 22:749–758. https://doi.org/10.1007/s00468-008-0235-6
Čufar K, Grabner M, Morgós A et al (2014) Common climatic signals affecting oak tree-ring growth in SE Central Europe. Trees Struct Funct 28:1–11. https://doi.org/10.1007/s00468-013-0972-z
de Luis M, Gričar J, Čufar K, Raventós J (2007) Seasonal dynamics of wood formation in Pinus halepensis from drya and semi-arid ecosystems in Spain. IAWA J 28:389–404. https://doi.org/10.1007/s10342-007-0199-x
De Luis M, Novak K, Raventós J et al (2011) Climate factors promoting intra-annual density fluctuations in Aleppo pine (Pinus halepensis) from semiarid sites. Dendrochronologia 29:163–169. https://doi.org/10.1016/j.dendro.2011.01.005
De Luis M, Čufar K, Di Filippo A et al (2013) Plasticity in dendroclimatic response across the distribution range of Aleppo pine (Pinus halepensis). PLoS ONE 8:e83550. https://doi.org/10.1371/journal.pone.0083550
Di Filippo A, Biondi F, Čufar K et al (2007) Bioclimatology of beech (Fagus sylvatica L.) in the Eastern Alps: spatial and altitudinal climatic signals identified through a tree-ring network. J Biogeogr 34:1873–1892. https://doi.org/10.1111/j.1365-2699.2007.01747.x
Dorado-Liñán I, Akhmetzyanov L, Menzel A (2017) Climate threats on growth of rear-edge European beech peripheral populations in Spain. Int J Biometeorol 61:2097–2110. https://doi.org/10.1007/s00484-017-1410-5
Farahat E, Linderholm HW (2018) Growth–climate relationship of European beech at its northern distribution limit. Eur J For Res 137:1–11. https://doi.org/10.1007/s10342-018-1129-9
Fritts HC (1972) Tree rings and climate. Sci Am 226:92–100. https://doi.org/10.1038/scientificamerican0572-92
García-González I, Souto-Herrero M (2017) Earlywood vessel area of Quercus pyrenaica Willd. is a powerful indicator of soil water excess at growth resumption. Eur J For Res 136:329–344. https://doi.org/10.1007/s10342-017-1035-6
García-Suárez AM, Butler CJ, Baillie MGL (2009) Climate signal in tree-ring chronologies in a temperate climate: a multi-species approach. Dendrochronologia 27:183–198. https://doi.org/10.1016/j.dendro.2009.05.003
Granier A, Reichstein M, Bréda N et al (2007) Evidence for soil water control on carbon and water dynamics in European forests during the extremely dry year: 2003. Agric For Meteorol 143:123–145. https://doi.org/10.1016/j.agrformet.2006.12.004
Gričar J, Zupančič M, Čufar K, Oven P (2007) Regular cambial activity and xylem and phloem formation in locally heated and cooled stem portions of Norway spruce. Wood Sci Technol 41:463–475. https://doi.org/10.1007/s00226-006-0109-2
Gričar J, Prislan P, De Luis M et al (2016) Lack of annual periodicity in cambial production of phloem in trees from Meditterranean areas. IAWA J 37:349–364. https://doi.org/10.1163/22941932-20160138
Hartl-Meier C, Dittmar C, Zang C, Rothe A (2014) Mountain forest growth response to climate change in the Northern Limestone Alps. Trees 28:819–829. https://doi.org/10.1007/s00468-014-0994-1
Hastie T, Tibshirani R (1986) Generalized additive models. Stat Sci 1:297–310. https://doi.org/10.1214/ss/1177013604
IPCC (2013) Fifth assessment report of the intergovernmental panel on climate change. In: Pachauri RK, Meyer LA (eds) Core Writing Team. IPCC, Geneva, Switzerland, 151 pp
Kharal DK, Thapa UK, St. George S et al (2017) Tree-climate relations along an elevational transect in Manang Valley, central Nepal. Dendrochronologia 41:57–64. https://doi.org/10.1016/j.dendro.2016.04.004
Kraus C, Zang C, Menzel A (2016) Elevational response in leaf and xylem phenology reveals different prolongation of growing period of common beech and Norway spruce under warming conditions in the Bavarian Alps. Eur J For Res 135:1011–1023. https://doi.org/10.1007/s10342-016-0990-7
Longares Aladrén LA (2004) Moncayo vegetal landscape in Aragón. In: Peña Monne JL, Longares LA, Sanchez Fabre M (eds) Physical Geography of Aragón. General and thematic subjects. Universidad de Zaragoza and Institución Fernando el Católico, Zaragoza, pp 187–197
Loran C, Kienast F, Bürgi M (2018) Change and persistence: exploring the driving forces of long-term forest cover dynamics in the Swiss lowlands. Eur J For Res 137:693–706. https://doi.org/10.1007/s10342-018-1134-z
Martínez del Castillo E, García-Martin A, Longares Aladrén LA, de Luis M (2015) Evaluation of forest cover change using remote sensing techniques and landscape metrics in Moncayo Natural Park (Spain). Appl Geogr 62:247–255. https://doi.org/10.1016/j.apgeog.2015.05.002
Martínez del Castillo E, Longares Aladrén LA, Gričar J et al (2016) Living on the edge: contrasted wood-formation dynamics in Fagus sylvatica and Pinus sylvestris under Mediterranean Conditions. Front Plant Sci 7:370. https://doi.org/10.3389/fpls.2016.00370
Martínez del Castillo E, Prislan P, Gričar J et al (2018a) Challenges for growth of beech and co-occurring conifers in a changing climate context. Dendrochronologia 52:1–10. https://doi.org/10.1016/J.DENDRO.2018.09.001
Martínez del Castillo E, Tejedor E, Serrano-Notivoli R et al (2018b) Contrasting patterns of tree growth of Mediterranean Pine Species in the Iberian Peninsula. Forests 9:416. https://doi.org/10.3390/f9070416
Mérian P, Bontemps J-D, Bergès L, Lebourgeois F (2011) Spatial variation and temporal instability in climate–growth relationships of sessile oak (Quercus petraea [Matt.] Liebl.) under temperate conditions. Plant Ecol 212:1855–1871. https://doi.org/10.1007/s11258-011-9959-2
Mott CL (2010) Environmental constraints to the geographic expansion of plant and animal species. Nat Educ Knowl 3(10):72
Nabuurs GJ, Delacote P, Ellison D et al (2017) By 2050 the mitigation effects of EU forests could nearly double through climate smart forestry. Forests 8:1–14. https://doi.org/10.3390/f8120484
National Research Council (2012) Assessing the reliability of complex models. National Academies Press, Washington, DC
Nieto Quintano P, Caudullo G, de Rigo D (2016) Quercus pyrenaica in Europe: distribution, habitat, usage and threats. In: San-Miguel-Ayanz J, de Rigo D, Caudullo G, Houston Durrant T, Mauri A (eds) European atlas of forest tree species. The Publications Office of the European Union, Luxembourg, p e01f807
Novak K, De Luis M, Gričar J et al (2016) Missing and dark rings associated with drought in Pinus halepensis. IAWA J 37:260–274. https://doi.org/10.1163/22941932-20160133
Pearson RG, Dawson TP (2003) Predicting the impacts of climate change on the distribution of species: are bioclimate envelope models useful? Glob Ecol Biogeogr 12:361–371. https://doi.org/10.1046/j.1466-822X.2003.00042.x
Pearson RG, Dawson TP, Liu C (2004) Modelling species distributions in Britain: a hierarchical integration of climate and land-cover data. Ecography (Cop) 27:285–298. https://doi.org/10.1111/j.0906-7590.2004.03740.x
Pérez-de-Lis G, Rozas V, Vázquez-Ruiz RA, García-González I (2018) Do ring-porous oaks prioritize earlywood vessel efficiency over safety? Environmental effects on vessel diameter and tyloses formation. Agric For Meteorol 248:205–214. https://doi.org/10.1016/j.agrformet.2017.09.022
Ponocná T, Spyt B, Kaczka R et al (2016) Growth trends and climate responses of Norway spruce along elevational gradients in East-Central Europe. Trees 30:1633–1646. https://doi.org/10.1007/s00468-016-1396-3
Porté A, Bartelink H (2002) Modelling mixed forest growth: a review of models for forest management. Ecol Modell 150:141–188. https://doi.org/10.1016/S0304-3800(01)00476-8
Prislan P, Gričar J, de Luis M et al (2013) Phenological variation in xylem and phloem formation in Fagus sylvatica from two contrasting sites. Agric For Meteorol 180:142–151. https://doi.org/10.1016/j.agrformet.2013.06.001
Prislan P, Gričar J, de Luis M et al (2016) Annual cambial rhythm in Pinus halepensis and Pinus sylvestris as indicator for climate adaptation. Front Plant Sci 7:1923. https://doi.org/10.3389/fpls.2016.01923
Reed KL, Clark SG (1978) The niche and forest growth. In: Edwards RL (ed) The natural behavior and response to stress of Western Coniferous Forests, IBP SYNTHE. Dowden, Hutchinson & Ross, Stroudsburg
Risser PG (1995) The status of the science examing ecotones. Bioscience 45:318–325
Rozas V, Camarero JJ, Sangüesa-Barreda G et al (2015) Summer drought and ENSO-related cloudiness distinctly drive Fagus sylvatica growth near the species rear-edge in northern Spain. Agric For Meteorol 201:153–164. https://doi.org/10.1016/j.agrformet.2014.11.012
Sáenz-Romero C, Lindig-Cisneros RA, Joyce DG et al (2016) Assisted migration of forest populations for adapting trees to climate change. Rev Chapingo Ser Ciencias For y del Ambient XXII:303–323. https://doi.org/10.5154/r.rchscfa.2014.10.052
Sass-Klaassen UGW, Fonti P, Cherubini P et al (2016) A tree-centered approach to assess impacts of extreme climatic events on forests. Front Plant Sci 7:1069. https://doi.org/10.3389/fpls.2016.01069
Serrano-Notivoli R, Beguería S, Saz Sánchez MA et al (2017a) SPREAD: a high-resolution daily gridded precipitation dataset for Spain—an extreme events frequency and intensity overview. Earth Syst Sci Data 9:721–738. https://doi.org/10.5194/essd-9-721-2017
Serrano-Notivoli R, de Luis M, Beguería S (2017b) An R package for daily precipitation climate series reconstruction. Environ Model Softw 89:190–195. https://doi.org/10.1016/j.envsoft.2016.11.005
Tegel W, Seim A, Hakelberg D et al (2014) A recent growth increase of European beech (Fagus sylvatica L.) at its Mediterranean distribution limit contradicts drought stress. Eur J For Res 133:61–71. https://doi.org/10.1007/s10342-013-0737-7
Walentowski H, Falk W, Mette T et al (2017) Assessing future suitability of tree species under climate change by multiple methods: a case study in southern Germany. Ann For Res 60:101–126. https://doi.org/10.15287/afr.2016.789
Wang X, Yu D, Wang S et al (2017) Tree height-diameter relationships in the alpine treeline ecotone compared with those in closed forests on Changbai Mountain, Northeastern China. Forests 8:1–13. https://doi.org/10.3390/f8040132
Weemstra M, Eilmann B, Sass-Klaassen UGW, Sterck FJ (2013) Summer droughts limit tree growth across 10 temperate species on a productive forest site. For Ecol Manag 306:142–149. https://doi.org/10.1016/j.foreco.2013.06.007
Wullschleger SD, Epstein HE, Box EO et al (2014) Plant functional types in Earth system models: past experiences and future directions for application of dynamic vegetation models in high-latitude ecosystems. Ann Bot 114:1–16
Zimmermann J, Hauck M, Dulamsuren C, Leuschner C (2015) Climate warming-related growth decline affects Fagus sylvatica, but not other broad-leaved tree species in Central European mixed forests. Ecosystems 18:560–572. https://doi.org/10.1007/s10021-015-9849-x
Acknowledgements
This study was supported by DGA-La Caixa (Project GA-LC-031/2010) and by the Spanish Ministry of Economy and competitiveness (Projects CGL2012-31668, CGL2015-69985). E. Martinez del Castillo benefited from a PhD Grant (No. BES-2013-064453) funded by the Spanish Ministry of Economy and competitiveness. International cooperation was supported by a short stay (No. EEBB-I-15-09810) and by the COST Action FP1106, (STReESS). The authors gratefully thank the staff of Moncayo Natural Park (Gob. Aragón) for their help in the field work.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interests.
Additional information
Communicated by David Drew.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Martínez del Castillo, E., Longares, L.A., Serrano-Notivoli, R. et al. Spatial patterns of climate–growth relationships across species distribution as a forest management tool in Moncayo Natural Park (Spain). Eur J Forest Res 138, 299–312 (2019). https://doi.org/10.1007/s10342-019-01169-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-019-01169-3