
Abstract
Chemical insecticides often are not efficient in reducing Alphitobius diaperinus populations in poultry farms and have induced insect resistance. Entomopathogenic nematodes (EPNs) appear as an alternative to pest biocontrol agents; however, their efficiency on rice hull litter is still unknown. The aim of this study was to evaluate the potential of Steinernema rarum (CUL isolate), Heterorhabditis bacteriophora (SMC isolate) and their symbiotic bacteria as biocontrol agents of A. diaperinus larvae and adults in rice hull litter. Mortality of insect adults and fifth- and seventh-instar larvae was determined in Petri dishes containing filter paper or rice hull. The effect of Xenorhabdus szentirmaii and Photorhabdus luminescens on L7 and adults in rice hull substrate was also evaluated. Moreover, the effectiveness of the EPNs was assessed in bags in an environment simulating the prevailing conditions in a broiler chicken farm. Under laboratory conditions, insect mortality was significantly affected by EPN species, substrate, insect stage and time after application. Mortality of L5 and L7 caused by EPNs in rice hulls reached a peak of 64 and 57.3 %, respectively, whereas symbiotic bacteria applied on rice hull showed low mortality rate at both insect stages. In the simulated poultry farm conditions, S. rarum and H. bacteriophora caused up to 40.8 and 50.4 % mortality of A. diaperinus larvae, respectively, but with adult mortality being low for both species. We conclude that infective juveniles from S. rarum CUL and H. bacteriophora SMC isolates can be used as biological control agents of A. diaperinus larvae in chicken litter containing rice hulls.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Key message
-
Alphitobius diaperinus is the principal pest affecting poultry facilities worldwide.
-
Chemical insecticide resistance development in Alphitobius diaperinus indicates an urgent need to develop alternative control methods.
-
Entomopathogenic nematodes are potential biological control agents of Alphitobius diaperinus.
-
This is the first report evaluating the efficacy of entomopathogenic nematode application on rice hull litter.
-
Steinernema rarum CUL and Heterorhabditis bacteriophora SMC isolates had promising control efficacy against Alphitobius diaperinus larvae in chicken litter containing rice hulls.
Introduction
Alphitobius diaperinus (Coleoptera: Tenebrionidae) is the principal pest affecting the poultry industry worldwide. Temperature and humidity conditions used in intensive broiler breeding systems are suitable for A. diaperinus development and ensure permanent food availability (Chernaki and Almeida 2001). These insects occur in chicken litter, a mixture of source material (rice hulls, wood shavings, etc.), excreta, feathers and other avian detritus, wasted feed and water deposited on the floor of a breeding facility (Runge et al. 2007). A. diaperinus feeds on chicken feed and even on weakened birds or carrion (Japp et al. 2010).
The presence of A. diaperinus in intensive poultry breeding systems affects both avian yield and animal and human health. This beetle transmits several viral, fungal and bacterial diseases (such as Fowlpox, Newcastle Disease, Aspergillus spp., Escherichia spp., Salmonella spp. and Campylobacter spp.), which can cause poultry weight loss and even death (Bates et al. 2004; Chernaki-Leffer et al. 2002; Hilbert et al. 2012; Leffer et al. 2010; Skov et al. 2004; Wales et al. 2010). Moreover, the digestive tract of young birds consuming this insect pest is damaged by the hardened elytra that hinder nutrient absorption or favor disease transmission (Despins and Axtell 1995). The etiological agents can multiply in the insect guts and be transmitted to poultry (in the same or different flocks); when chicken are slaughtered, contaminated meat reaches the consumer. Moreover, direct contact with larvae and adults can be detrimental, especially for people directly related to poultry farming. Schroeckenstein et al. (1988) demonstrated that A. diaperinus can cause severe allergic reactions, rhinitis and asthma in humans.
The primary strategy for A. diaperinus control in poultry houses involves the use of pyrethroid or organophosphate chemical products on chicken litter during the short periods when the facilities are empty. These chemical treatments proved to be barely efficient in reducing pest population because they are applied in a medium with a high amount of organic matter and dust (Despins et al. 1991; Japp et al. 2010). In addition, inappropriate use of chemicals and unsuitable control management practices often generate insecticide-resistant A. diaperinus populations (Chernaki-Leffer et al. 2011; Lambkin 2005). Thus, there is a need for exploring new pest control strategies, such as the use of natural antagonists.
Entomopathogenic nematodes (EPNs) of the genera Steinernema (Nematoda: Steinernematidae) and Heterorhabditis (Nematoda: Heterorhabditidae) are insect parasites with potential for use as biological control agents of insect pests (Hussaini 2014). Infective juveniles (IJs) locate the insect host and penetrate into the haemocoel through natural openings or the cuticle. Once inside the host, they release symbiotic bacteria carried in their digestive tract which kill the insect by septicemia about 48 h after infection (Forst and Clarke 2002). The genus Steinernema is related to bacteria of the genus Xenorhabdus, whereas the genus Heterorhabditis is related to Photorhabdus spp. EPNs then feed on bacteria and digested tissues and complete one to three generations inside the host, until IJs emerge and move through the soil (Poinar 1990).
The potential of EPNs for suppressing A. diaperinus is well documented (Alves et al. 2005, 2012; Geden et al. 1985, 1987; Szalanski et al. 2004). However, the diversity of materials used to prepare chicken litter in poultry systems worldwide and the heterogeneity of microenvironments generated in poultry facilities show the need for further research (Benabdeljelil and Ayachi 1996; Garcia et al. 2012; Runge et al. 2007).
Rice hull, a by-product of the rice industry, is one of the main litter materials used as bedding in poultry facilities worldwide (Monira et al. 2003). Its primary function is to avoid direct contact of poultry with the floor, maintain the temperature balance in the facilities and absorb the moisture generated by the poultry feces. Despite the wide use of this material as chicken litter, to date no research studies have been conducted to determine the behavior of EPNs or their symbiotic bacteria in this basic material.
The aim of this work was to evaluate the potential of S. rarum (CUL isolate), Heterorhabditis bacteriophora (SMC isolate) and their symbiotic bacteria as biocontrol agents of A. diaperinus larvae and adults in broiler chicken farms. We hypothesize that the application of these EPNs in rice hull can control A. diaperinus.
In the field of biological control with EPNs, H. bacteriophora stands out for its efficacy in controlling several insect pest species (Grewal et al. 2005). Steinernema rarum was ecologically characterized, and was found to be pathogenic to a large number of insect families (de Doucet et al. 1999; Koppenhöfer and Kaya 1999). We used these species because their pathogenicity against A. diaperinus was found to be acceptable in previous assays.
Materials and methods
Study material
The isolates S. rarum CUL (Cululú, Santa Fe, Argentina) and H. bacteriophora SMC (Saladero Mariano Cabal, Santa Fe, Argentina) were multiplied on last-instar larvae of Galleria mellonella (Lepidoptera: Pyralidae), following the procedure of Kaya and Stock (1997). IJs were collected using modified White traps and stored at 16 °C for 7–14 days, until the start of the experiments.
Larvae and adults of A. diaperinus were collected from commercial broiler chicken poultry farms from Esperanza (Santa Fe, Argentina), where no insecticides had been applied for the previous 6 months. A. diaperinus were reared in plastic boxes containing chicken feed (Avigan®) at 25 °C at the Entomology Laboratory of the Facultad de Ciencias Agrarias (Universidad Nacional del Litoral). Fifth- and seventh-instar larvae (L5 and L7, respectively) were morphologically differentiated following the procedures described by Francisco and do Prado (2001). At the time of the experiments, L5, L7 and adults were approximately 26, 34 and 40 days old, respectively.
Rice hull material was obtained from Santa Fe province and had not been treated with chemicals before the experiment. Rice hull used as chicken litter was considered dry or wet if it contained 6 or 12 % moisture, respectively.
Pathogenicity of IJs to A. diaperinus
Pathogenicity of S. rarum CUL and H. bacteriophora SMC IJs to A. diaperinus adults, L5 and L7 was tested using Petri dishes (50 mm) either lined with filter paper (Whatman No. 1) or containing 2 g of dry or wet rice hull. Mortality of A. diaperinus was determined in both treatments.
Ten larvae or adults of the insect were put in each Petri dish; then, 500 IJs (Del Valle et al. 2014) contained in 0.5 ml of aqueous solution were surface applied to the test arena (insects were not directly sprayed because once the insects were introduced in the dish, they immediately moved downwards). Control treatments consisted of the addition of an equal volume of distilled water on each substrate and tested insect stage. Petri dishes were maintained in an incubator at 25 °C, 60 % RH under a 14L:10D photoperiod. Insect mortality was evaluated 48 and 96 h after nematode application. Cadavers were further incubated for 4 days and then dissected to verify the presence of nematodes. Five replications per treatment were performed and the experiment was repeated three times.
Isolation, identification and quantification of symbiotic bacteria
Symbiotic bacteria were isolated from last-instar larvae of G. mellonella infected by S. rarum CUL and H. bacteriophora SMC. The insect cadaver was surface-disinfected with 70 % alcohol 30 h after infection and dissected aseptically under a laminar flow hood. A small amount of hemolymph was taken with a wire loop and streaked on the surface of the NBTA medium. Petri dishes were incubated at 28 °C under aerobiosis conditions for 48 h. Then, a colony showing the morphological characteristics of phase I was selected for isolation (Akhurst 1980).
DNA was extracted using a commercial kit (Wizard® Genomic DNA Purification, Promega) to identify the symbiotic bacteria of each isolate. The 16S rDNA gene was amplified using the universal primers 27F (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1492R (5′-GGYTACCTTGTTACGACTT-3′). Each PCR tube (50 μl) contained 10 μl of a 5X reaction mixture (Promega), 1.5 mM MgCl2, 200 μM of each dNTP (Promega), 0.4 μM of each primer, 1 U GoTaq Polymerase (Promega) and 5 μl of template DNA. An MJ Research thermocycler was used; the cycling program was as follows: 94 °C for 5 min; 30 cycles of 94 °C for 1 min, 55 °C for 1 min and 72 °C for 1 min; and a final extension of 72 °C for 7 min. PCR products were visualized using 1 % p/v agarose gel electrophoresis, Gelred® staining and observation under UV light (DyNA Light UV Transilluminator, LabNet, UV light source wave length 302 nm). PCR products were purified using a commercial kit (Wizard PCR SV Gel and PCR Clean-Up System kit, Promega) and sequenced (Macrogen, South Korea). The sequences were compared with other sequences deposited at GenBank using BLAST algorithm (http://www.ncbi.nlm.nih.gov/BLAST/). The bacterial isolate obtained from S. rarum CUL was identified as Xenorhabdus szentirmaii DSPV001N and the one obtained from H. bacteriophora SMC was identified as P. luminiscens subsp. laumondii DSPV002N.
Symbiotic bacteria were multiplied in TSB (Oxoid, England) at 28 °C for 48 h under aerobiosis and agitation at 200 rpm. Serial dilutions from the stock culture were performed to determine the bacterial concentrations to be used in the experiments. Optical density of this culture and its dilutions was determined at 600 nm (Jenway® spectrophotometer) and, at the same time, bacterial plate count was made. The bacterial strains were quantified via a regression analysis performed with both parameters.
Pathogenicity of symbiotic bacteria
Mortality produced by X. szentirmaii and Photorhabdus luminescens in A. diaperinus L7 and adults was evaluated. Petri dishes (50 mm) lined with filter paper (Whatman No. 1) containing 2 g of dry or wet rice hull were used. Ten insect larvae or adults, depending on the treatment, were put in each Petri dish and 500 µl of bacterial suspension (1 × 108 CFU/ml; 3 % Tween-80) were then surface applied (not directly on the insects).
Control treatment consisted of an equal volume of distilled water and Tween-80 (3 %) applied on each substrate and insect stage evaluated. Petri dishes were maintained at 25 °C in an incubation chamber and insect mortality was evaluated 48 and 96 h after bacterium application. Five replications per treatment were performed and the experiment was repeated three times.
Reproduction of IJs on A. diaperinus
Ten L7 or adults of A. diaperinus contained in each Petri dish (50 mm diameter) lined with Whatman No. 1 filter paper were exposed to 500 IJs of S. rarum or H. bacteriophora contained in 0.5 ml aqueous suspension and incubated at 25 °C for 48 h. Then, the cadavers were put individually in White traps (Kaya and Stock 1997) and maintained in the dark at 25 °C for 14 days to allow multiplication and emergence of all IJs. Emerged IJs were collected daily and kept at 16 °C. IJs collected from each insect were suspended in 100 ml of aqueous suspension and the number of IJs from each cadaver was calculated by counting 3 1-ml aliquots. Five replications were performed for each insect stage and ENP isolate. The experiment was repeated three times.
Experiments in bags containing rice hull
The effectiveness of S. rarum CUL and H. bacteriophora SMC was evaluated in a growth chamber in an environment simulating the conditions prevailing in a broiler chicken farm (25 °C and 60 % RH). The simulated environment consisted of bags made of nylon mesh (mesh pore size: 0.1 mm) of 3 l of volume capacity, containing 200 g of wet rice hull and 50 g of balanced feed for poultry (Avigan®). Both L7 (200) and adults (50) were introduced into each bag, and IJs (20000) of S. rarum or H. bacteriophora, depending on the treatment, contained in 10 ml of distilled water were then applied (not directly on the insects). The bag was immediately closed to prevent the escape of insects. Bags were put individually in boxes (50 length × 30 width × 20 cm height) filled with chicken litter (with approximately 17 % moisture) obtained from a commercial broiler breeding farm; litter had been used for 3 weeks. The control treatment received an equal amount of distilled water without EPNs. The boxes containing the treated bags were kept in a growth chamber at 25 °C. The assay consisted of a completely randomized design, with each treatment being repeated six times. Seven days after the start of the experiments, each bag was carefully examined to determine mortality of A. diaperinus L7 and adults. This experiment was repeated three times.
Statistical analysis
The results were analyzed using an analysis of variance (ANOVA) in INFOSTAT statistical software (Di Rienzo et al. 2010). A factorial analysis was performed to determine the effects of treatments, chicken litter, insect stage and time after application. The data were obtained as percentages and were square root arcsine transformed before statistical analysis (non-transformed means are presented in tables and figures) to meet the requirements of normality and homogeneity of variances. Differences between means were compared using the Fisher’s LSD test with a probability of 5 %.
Results
Pathogenicity of IJs to A. diaperinus
Insect mortality was significantly affected by treatments (F = 551.4; df = 2,756; P < 0.0001), substrates (F = 104.63; df = 2,756; P < 0.0001), insect stage (F = 112.78; df = 2,756; P < 0.0001) and time after application (F = 962.2; df = 1,756; P < 0.0001). Significant interactions were detected between: treatments and substrates (F = 27.12; df = 4,756; P < 0.0001), treatments and insect stage (F = 45.35; df = 4,756; P < 0.0001), and treatments and time after application (F = 207.96; df = 2,756; P < 0.0001).
The isolates of S. rarum and H. bacteriophora placed on filter paper caused higher mortality of A. diaperinus L5 and L7 than the corresponding controls. S. rarum produced higher larval mortality on day 2 and 4 at L7 stage than H. bacteriophora (F = 25.48; df = 2,42; P < 0.0001 and F = 107.45; df = 2,42; P < 0.0001, respectively). No statistical differences in L5 mortality caused by either EPN species were observed on days 2 (F = 36.86; df = 2,42; P < 0.0001) or 4 (F = 62.83; df = 2,42; P < 0.0001) after inoculation. In the experiments using filter paper, mortality of adults was 60 % 4 days after inoculation with H. bacteriophora, with statistical differences from S. rarum (F = 8.62; df = 2,42; P = 0.0007 and F = 129.73; df = 2,42; P < 0.0001 for days 2 and 4, respectively) (Table 1).
The experiments that involved dry rice hull did not show significant differences between treatments on day 2 after nematode inoculation (F = 1.07; df = 2,42; P = 0.3527, F = 1.00; df = 2,42; P = 0.3765 and F = 1.07; df = 2,42; P = 0.3509 for L5, L7 and adults, respectively). On day 4, S. rarum and H. bacteriophora caused mortality in 54 and 63.3 % of L5 and in 42.7 and 46.7 % of L7, respectively (F = 164.81; df = 2,42; P < 0.0001 and F = 90.39; df = 2,42; P < 0.0001; respectively). Mortality of adults treated in rice hull did not exceed 2.7 %, except for the treatment involving H. bacteriophora IJs on day 4 after inoculation (F = 8.15; df = 2,42; P < 0.0001) (Table 1).
The treatment involving wet rice hull exhibited low insect larval and adult mortality on day 2, with statistical differences between control and EPNs only at L5 stage (F = 16.73; df = 2,42; P < 0.0001). On day 4 after inoculation in wet rice hull, IJs of S. rarum caused higher mortality in A. diaperinus larvae than H. bacteriophora IJs (F = 112.59; df = 2,42; P < 0.0001 and F = 74.14; df = 2,42; P < 0.0001 for L5 and L7, respectively). Mortality of adult insects caused by H. bacteriophora IJs on day 4 was significantly higher than S. rarum (F = 21.92; df = 2,42; P < 0.0001) (Table 1).
The interaction between time after application and insect stage was significant. Statistical differences were observed between control and insect stage at day 4 after application, with mortality rate being higher at L5 than at L7 (F = 3.53; df = 2,130; P = 0.0322).
Mortality caused by S. rarum varied significantly with the different substrates (F = 62.61; df = 2,130; P < 0.0001 and F = 27.08; df = 2,130; P < 0.0001 for 2 and 4 days, respectively). Insect mortality caused by S. rarum was highest on filter paper and lowest on dry rice hulls. Mortality inflicted by H. bacteriophora was higher in experiments using filter paper than those using rice hulls, without significant differences between dry or wet rice hull substrate (F = 21.13; df = 2,130; P < 0.0001 and F = 14.07; df = 2,130; P < 0.0001 for 2 and 4 days, respectively).
On day 2 after application, mortality caused by S. rarum differed significantly between insect stages, being highest in L5 and lowest in adults (F = 31.42; df = 2,130; P < 0.0001). However, on day 4, there were no differences between L5 and L7, but there were differences between these larval stages and the adult stage, which showed the lowest mortality (F = 104.67; df = 2,130; P < 0.0001).
Mortality caused by H. bacteriophora was lower in adults and L7 than in L5 on day 2 after application (F = 9.92; df = 2,130; P < 0.0001); on day 4, mortality caused by this isolate showed significant differences among the three insect stages, being highest in L5 and lowest in adults (F = 12.73; df = 2,130; P < 0.0001).
Pathogenicity of symbiotic bacteria to A. diaperinus
A significant effect of treatments (F = 85.52; df = 2,504; P < 0.0001), insect stage (F = 16.93; df = 1,504; P < 0.0001) and substrate (F = 87.42; df = 2,504; P < 0.0001) was observed on mortality of A. diaperinus in this experiment. Time after application did not have a significant effect (F = 2.12; df = 1,504; P = 0.1460). The only significant interaction was detected between treatments and substrates (F = 75.64; df = 4,504; P < 0.0001).
Photorhabdus luminescens DSPV 002N caused mortality of above 18 and 28 % of A. diaperinus larvae and adults in the filter paper experiments on days 2 (F = 19.39; df = 2,42; P < 0.0001 and F = 29.0; df = 2,42; P < 0.0001, respectively) and 4 (F = 34.88; df = 2,42; P < 0.0001 and F = 21.99; df = 2,42; P < 0.0001, respectively) after inoculation. The use of X. szentirmaii DSPV 001N was not effective against the insect (Table 2). The experiments involving dry and wet rice hull showed low or nil mortality rate in both insect stages, without significant differences between treatments (Table 2).
The substrates did not affect mortality rates obtained in control treatments (F = 0.16; df = 2,86; P = 0.8505 and F = 0.35; df = 2,86; P = 0.7654 for 2 and 4 days, respectively) and treatments involving the application of S. rarum (F = 0.09; df = 2,86; P = 0.9175 and F = 0.58; df = 2,86; P = 0.5648 for 2 and 4 days, respectively). However, insect mortality rate was higher after the application of H. bacteriophora on filter paper than rice hulls (F = 44.17; df = 2,86; P < 0.0001 and F = 55.81; df = 2,86; P < 0.0001 for 2 and 4 days, respectively).
No significant differences between insect stages were observed in mortality rates of larvae and adults of A. diaperinus in the experiments using H. bacteriophora (F = 1.98; df = 1,86; P = 0.1635 and F = 1.69; df = 1,86; P = 0.1966 for 2 and 4 days, respectively). The effect of insect stage was significant in control treatments (F = 7.95; df = 1,86; P = 0.006 and F = 8.46; df = 1,86; P = 0.0046 for 2 and 4 days, respectively) and in experiments involving S. rarum (F = 4.22; df = 1,86; P = 0.0429 and F = 7.73; df = 1,86; P = 0.0067 for 2 and 4 days, respectively).
Reproduction of IJs in A. diaperinus
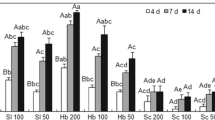
The S. rarum isolate exhibited a greater reproductive potential than H. bacteriophora had on A. diaperinus larvae (F = 46.57; df = 1,28; P < 0.0001). However, a higher number of IJs of H. bacteriophora than S. rarum was generated in adult insects (F = 12.10; df = 1,28; P = 0.0017) (Fig. 1). Both EPNs exhibited higher reproduction potential of IJs in insect larvae (F = 201.09; df = 1,28; P < 0.0001 and F = 9.10; df = 1,28; P = 0.0054 for S. rarum and H. bacteriophora, respectively) than in adults.

Number of infective juveniles of Steinernema rarum CUL and Heterorhabditis bacteriophora SMC emerged from seventh-instar larvae and adults of Alphitobius diaperinus. Results are expressed as mean ± standard error. Different letters indicate significant differences between treatments, according to Fisher’s LSD test (P < 0.05)
Experiments in bags containing rice hull
The first replicate of the experiment, which simulated poultry farm conditions, showed that IJs of the H. bacteriophora isolate killed 35.6 % of A. diaperinus larvae, showing statistical differences from S. rarum (F = 4.37; df = 2,15; P = 0.0319) (Fig. 2a). The second and third replicates of the experiments involving both EPNs produced 38 and 50.4 % mortality of insect larvae, without differences between isolates, but with differences from the control treatment (F = 33.56; df = 2,15; P < 0.0001 and F = 34.52; df = 2,15; P < 0.0001, respectively) (Fig. 2b, c).

Mortality of seventh-instar larvae and adults of Alphitobius diaperinus caused by infective juveniles of Steinernema rarum CUL and Heterorhabditis bacteriophora SMC in nylon bags containing rice hull. Results are expressed as mean ± standard error. Uppercase and lowercase letters indicate statistical differences according to Fisher’s LSD test (P < 0.05) between treatments and adult and larvae of A. diaperinus, respectively. a, b and c correspond to the three replications of the experiment
Mortality of A. diaperinus adults did not differ statistically from the control, either in the first or the second replicate of the experiment (F = 1.25; df = 2,15; P = 0.3137 and F = 0.05; df = 2,15; P = 0.9538, respectively) (Fig. 2a, b). However, H. bacteriophora IJs caused 11.7 % mortality of adults in the third replicate of the experiment, with this value being significantly higher than in the control treatment (F = 3.84; df = 2,15; P = 0.0451) (Fig. 2c).
Discussion
The results obtained in this work confirm the hypothesis postulating that the studied EPNs applied in rice hull can be used as biological control agents of A. diaperinus. IJs of S. rarum CUL and H. bacteriophora SMC were pathogenic to A. diaperinus larvae and, to a lesser degree, to adults. By contrast, the bacteria associated with both EPNs were not efficient in suppressing the pest in rice hull.
Steinernema rarum IJs applied in rice hull caused high mortality (42–64 %) of A. diaperinus L5 and L7, indicating that IJs of this isolate can survive and remain infective in the rice hull medium. S. rarum has a host foraging strategy at an intermediate level between “ambusher” and “cruiser” (Koppenhöfer and Kaya 1999), which would allow these nematodes to remain nictating on rice hull waiting for hosts or to move in search of them. Larvae and adults of A. diaperinus show active movement in chicken litter bedding in poultry farms; therefore, the ambusher behavior can be considered an ecological adaptation that would favor A. diaperinus control. Mortality values of A. diaperinus larvae caused by S. rarum obtained in the present study are close to values reported for S. carpocapsae (Alves et al. 2005; Pezowicz et al. 2013), a species with ambusher foraging strategy (Campbell and Gaugler 1993).
The H. bacteriophora SMC isolate caused between 37 and 46 % mortality of A. diaperinus larval stages when applied in rice hull. Pezowicz et al. (2013) recorded 20 % mortality of A. diaperinus larvae caused by H. bacteriophora applied on pine sawdust, using a method very similar to the one used in the present work. In assays using Petri dishes lined with filter paper, Alves et al. (2012) reported 38 % mortality of A. diaperinus larvae caused by H. bacteriophora, a lower value than the one obtained in the present work. Mortality caused by H. bacteriophora is much lower than that caused by other species of the genus. For example, H. indica IJs produced more than 94 % larval mortality when applied on sawdust litter near poultry feeder pans (Rodrigueiro et al. 2008).
Heterorhabditis bacteriophora SMC exhibited higher virulence than S. rarum as biological control agent of adults of the studied pest. Our results show that adult mortality on rice hull was low for S. rarum and medium-to-low for H. bacteriophora. However, larvae and adults of A. diaperinus infected with EPNs can play an important role in chicken litter by generating new IJs that will be released near the hosts.
Our results indicate differences in susceptibility to the evaluated EPNs between larvae and adults, which is in agreement with findings reported by Rodrigueiro et al. (2008) and Geden et al. (1985).
Szalanski et al. (2004) reported a great variability in mortality of A. diaperinus adults caused by different isolates of S. carpocapsae and S. feltiae tested in filter paper assays. Accordingly, different isolates of the species evaluated here can vary in effectiveness as biological agents for the control of this host, indicating the need for individually evaluating each isolate to determine its biocontrol potential.
The widespread use of filter paper to evaluate EPN pathogenicity to insects should be validated with assays that indicate the behavior of EPNs at the site where they will be actually applied. Accordingly, our results clearly show that mortality was significantly lower in A. diaperinus larvae and adults in rice hull than in filter paper.
A chicken litter containing rice hull has particular physical and chemical characteristics that would influence the behavior of the present organisms. Its size, density, thermal conductivity and moisture retention capacity generate a suitable environment for poultry breeding (Scanes et al. 2004). The degree of hull moisture will affect IJ survival and movement. Moisture in chicken litter varies within a breeding facility, increasing near drinkers. IJs would find more suitable conditions to penetrate the hosts at these wetter sites, where they can move more easily and find a higher A. diaperinus population than in the remaining sites. Rodrigueiro et al. (2008) reported that applications of H. indica IBCB–N05 in chicken litter near feeders caused over 70 and 94 % mortality of A. diaperinus adults and larvae, respectively. In addition, the same work showed that EPN applications to a limited area of the facility can be efficient in reducing the pest population and significantly decreasing the amount of IJs that will be released, making the use of those EPNs economically feasible. Therefore, understanding each poultry production system and the heterogeneity of insect distribution, moisture and balanced feed should be considered in future research works that aim at optimizing IJ release in these environments.
Another aspect to be considered is that in Latin America, most of the commercial poultry breeding facilities use chicken litter on the floor. This is an advantage for nematodes, allowing them to remain active on the floor of the facilities as well, where a large proportion of pupae and larvae of A. diaperinus (Salin et al. 2000) are present.
The number of IJs generated in A. diaperinus larvae was several times lower than that found for other EPN species. For example, using an experimental design different from the one used here, Rocha Costa et al. (2007, 2008) determined that A. diaperinus larvae infected with S. carpocapsae and H. baujardi generated progenies of 7706 and 7421 IJs/larva, respectively. Szalanski et al. (2004) estimated that each adult of this pest can generate 5000 IJs of S. carpocapsae and S. feltiae, a greater progeny than that determined for H. bacteriophora SMC and S. rarum CUL. Differences in the progenies between larval and adult stages are due to differences in size, body weight and tissue composition. The external morphology of adult insects is likely to hinder invasion by IJs.
The application of P. luminescens DSPV 002N caused mortality of A. diaperinus larvae (18.7–22.7 %) and adults (28–30.7 %) in filter paper assays, but did not show any effects in rice hull. The use of X. szentirmaii DSPV 001N did not cause significant mortality in insect larvae or adults.
It is well known that Xenorhabdus spp. and Photorhabdus spp. need a nematode vector to penetrate the host and be released in the host hemolymph to further secrete toxins that will rapidly kill the insect (Forst and Clarke 2002; Rodou et al. 2010). However, Mahar et al. (2008) confirmed that applying solutions containing cells of X. nematophila or its cell-free filtrates was lethal to larvae of Spodoptera exigua and Plutella xylostella and nymphs of Schistocerca gregaria. These authors also isolated the bacteria from the dead hosts, indicating that the bacterium can enter the host in the absence of the nematode vector. Abdel-Razek (2003) reported mortality of pupae of Plutella xylostella caused by spray application of Xenorhabdus nematophilus and P. luminescens. The author attributed this result to the capacity of bacteria to penetrate the spiracular orifices (the only organ with an opening to the external environment) of P. xylostella pupal cases and survive to the host immune response. Mortality of A. diaperinus observed in our experiments involving the application of P. luminescens might be due to the fact that the bacterium managed to penetrate this insect through its natural openings (spiracles, mouth or anus). Further studies are necessary to determine the entry site/s of these bacteria into the insect.
Oral toxicity of EPN-associated bacteria have also been investigated. Gerritsen et al. (2005) indicated that Frankliniella occidentalis can take up toxins when sucking from a leaf sprayed with P. temperata, killing the insect. Shrestha et al. (2011) recorded mortality rates caused by the oral toxicity of Photorhabdus spp. of 100 and 45.3 % for G. mellonella and Lucilia caesar, respectively. The toxin complex of P. luminescens was lethal to Manduca sexta larvae by oral ingestion as well as hemocoel injection (Blackburn et al. 1998). Bacteria applied on chicken litter in poultry facilities might reach part of the balanced feed present in the litter. A. diaperinus larvae might ingest this feed as well as the bacteria, which might produce larval death or affect fecundity and/or normal development of future pest generations, as shown in other insect species by Gerritsen et al. (2005) and Shrestha et al. (2011).
The assays conducted in nylon bags containing rice hull showed promising results in terms of the control of A. diaperinus larvae with S. rarum CUL and H. bacteriophora SMC. However, the control of A. diaperinus adults was low with both isolates, especially with S. rarum CUL. Further studies about persistence and survival of IJs over time are necessary for implementing pest management programs using these EPNs.
Knowing the action of different EPN species and isolates is necessary to select the nematodes that are most adapted to a specific poultry environment, increasing the possibilities of success of future programs for the biological control of A. diaperinus. Our research work is the first in studying the efficiency of EPNs applied on rice hull and in evaluating the action of symbiotic bacteria associated with EPNs. This work is also the first one that involves an isolate of S. rarum in the control of this pest. We conclude that IJs from the S. rarum CUL and H. bacteriophora SMC isolates can be used as biological control agents of A. diaperinus larvae in chicken litter containing rice hull.
Author contribution
EEDV and LSF conceived and designed the research. EEDV, LSF, MM and MVZ conducted the experiments. EEDV, LSF, PL and MED analyzed data and wrote the manuscript. All authors read and approved the manuscript.
References
Abdel-Razek AS (2003) Pathogenic effects of Xenorhabdus nematophilus and Photorhabdus luminescens (Enterobacteriaceae) against pupae of the Diamondback moth, Plutella xylostella (L.). J Pest Sci 76:108–111
Akhurst RJ (1980) Morphological and functional dimorphism in Xenorhabdus spp., bacteria symbiotically associated with the insect pathogenic nematodes Neoaplectana and Heterorhabditis. J Gen Microbiol 121:303–309
Alves LFA, Rohde C, Alves VS (2005) Patogenicidade de Steinernema glaseri e S. carpocapsae (Nematoda: Rhabdita) contra o cascudinho, Alphitobius diaperinus (Panzer) (Coleoptera: Tenebrionidae). Neotrop Entomol 34:139–141
Alves VS, de Neves PMJO, Alves LFA, Moino A Jr, Holz N (2012) Entomopathogenic nematodes (Rhabditida: Heterorhabditidae and Steinernematidae) screening for lesser mealworm Alphitobius diaperinus (Coleoptera: Tenebrionidae) control. Rev Colomb Entomol 38:76–80
Bates C, Hiett KL, Stern NJ (2004) Relationship of Campylobacter isolated from poultry and from darkling beetles in New Zealand. Avian Dis 48:138–147
Benabdeljelil K, Ayachi A (1996) Evaluation of alternative litter materials for poultry. J Appl Poult Res 5:203–209
Blackburn M, Golubeva E, Bowen D, Ffrench-Constant RH (1998) A novel insecticidal toxin from Photorhabdus luminescens, Toxin complex a (Tca) and its histopathological effects on the midgut of Manduca sexta. Appl Environ Microbiol 64:3036–3041
Campbell JF, Gaugler R (1993) Nictation behavior and its ecological implications in the host search strategies of entomopathogenic nematodes (Heterorhabditidae and Steinernematidae). Behaviour 126:156–169
Chernaki AM, Almeida LM (2001) Exigências térmicas, período de desenvolvimento e sobrevivência de imaturos de Alphitobius diaperinus (Panzer) (Coleoptera: Tenebrionidae). Neotrop Entomol 30:365–368
Chernaki-Leffer AM, Biesdorf SM, Almeida LM, Leffer EVB, Vigne F (2002) Isolamento de enterobacérias em Alphitobius diaperinus e na cama de aviários no oeste do Estado do Paraná, Brasil. Rev Bras Cienc Avic 4:243–247
Chernaki-Leffer AM, Sosa-Gómez DR, Almeida LM, Lopes ION (2011) Susceptibility of Alphitobius diaperinus (Panzer) (Coleoptera, Tenebrionidae) to cypermethrin, dichlorvos and triflumuron in southern Brazil. Rev Bras Entomol 55:125–128
de Doucet MMA, Bertolotti MA, Giayetto AL, Miranda M (1999) Host range, specificity, and virulence of Steinernema feltiae, Steinernema rarum, and Heterorhabditis bacteriophora (Steinernematidae and Heterorhabditidae) from Argentina. J Invertebr Pathol 73:237–242
Del Valle EE, Balbi EI, Lax P, Rondán Dueñas J, Doucet ME (2014) Ecological aspects of an isolate of Steinernema diaprepesi (Rhabditida: Steinernematidae) from Argentina. Biocontrol Sci Technol 24:690–704
Despins JL, Axtell RC (1995) Feeding behavior and growth of broiler chicks fed larvae of the darkling beetle, Alphitobius diaperinus. Poult Sci 74:331–336
Despins JL, Turner EC Jr, Pfeiffer DG (1991) Evaluation of methods to protect poultry house insulation from infestations by lesser mealworm (Coleoptera: Tenebrionidae). J Agric Entomol 8:209–217
Di Rienzo JA, Casanoves F, Balzarini MG, Gonzalez L, Tablada M, Robledo CW (2010) InfoStat, release 2010. FCA, Universidad Nacional de Córdoba, Córdoba
Forst S, Clarke D (2002) Bacteria-nematode symbiosis. In: Gaugler R (ed) Entomopathogenic nematology. CABI, Wallingford, pp 57–77
Francisco O, do Prado AP (2001) Characterization of the larval stages of Alphitobius diaperinus (Panzer, 1797) (Coleoptera: Tenebrionidae) using head capsule width. Rev Bras Biol 61:125–131
Garcia RG, Almeida Paz ICL, Caldara FR, Nääs IA, Bueno LGF, Freitas LW, Graciano JD, Simm S (2012) Litter materials and the incidence of carcass lesions in broilers chickens. Braz J Poult Sci 14:219–224
Geden CJ, Axtell RC, Brooks WM (1985) Susceptibility of the lesser mealworm, Alphitobius diaperinus (Coleoptera: Tenebrionidae) to the entomogenous nematodes Steinernema feltiae, S. glaseri (Steinernematidae) and Heterorhabditis heliothidis (Heterorhabditidae). J Entomol Sci 20:331–339
Geden CJ, Arends JJ, Axtell RC (1987) Field trails of Steinernema feltiae (Nematoda: Steinernematidae) for control of Alphitobius diaperinus (Coleoptera: Tenebrionidae) in commercial broiler and turkey houses. J Econ Entomol 80:136–141
Gerritsen LJM, Georgieva J, Wiegers GL (2005) Oral toxicity of Photorhabdus toxins against thrips species. J Invertebr Pathol 88:207–211
Grewal PS, Ehlers R-U, Shapiro-Ilan DI (2005) Nematodes as biocontrol agents. CABI, Wallingford
Hilbert F, Smulders FJM, Chopra-Dewasthaly R, Paulsen P (2012) Salmonella in the wildlife-human interface. Food Res Int 45:603–608
Hussaini SS (2014) Potential of entomopathogenic nematodes in integrated pest management. In: Abrol DP (ed) Integrated pest management: current concepts and ecological perspective. Academic Press, Waltham, pp 193–223
Japp AK, Bicho CL, Silva AVF (2010) Importância e medidas de controle de Alphitobius diaperinus em aviários. Cienc Rural 40:1668–1673
Kaya HK, Stock SP (1997) Techniques in insect nematology. In: Lacey LA (ed) Manual of techniques in insect pathology, Biological techniques series. Academic Press, London, pp 281–324
Koppenhöfer AM, Kaya HK (1999) Ecological characterization of Steinernema rarum. J Invertebr Pathol 73:120–128
Lambkin TA (2005) Baseline responses of adult Alphitobius diaperinus (Panzer) (Coleoptera: Tenebrionidae) to fenitrothion, and susceptibility status of populations in Queensland and New South Wales, Australia. J Econ Entomol 98:938–942
Leffer AM, Kuttel J, Martins LM, Pedroso AC, Astolfi-Ferreira CS, Ferreira F, Ferreira AJ (2010) Vectorial competence of larvae and adults of Alphitobius diaperinus in the transmission of Salmonella enteritidis in poultry. Vector-Borne Zoonot 10:482–487
Mahar AN, Jan ND, Mahar GM, Mahar AQ (2008) Control of insects with entomopathogenic bacterium Xenorhabdus nematophila and its toxic secretions. Int J Agric Biol 10:52–56
Monira KN, Islam MA, Alam MJ, Wahid MA (2003) Effect of litter materials on broiler performance and evaluation of manureal value of used litter in late autumn. Asian Austral J Anim 16:555–557
Pezowicz E, Mazurkiewicz A, Tumialis D (2013) Sensitivity of imago and larvae of the lesser mealworm Alphitobius diaperinus (Panzer 1797) in a sawdust litter to selected species and strains of Steinernematidae and Heterorhabditidae under laboratory conditions. Ann Wars Univ of Life Sci—SGGW 52:161–166
Poinar GO (1990) Biology and taxonomy of Steinernematidae. In: Gaugler RR, Kaya HK (eds) Entomopathogenic nematodes in biological control. CRC Press, Boca Raton, pp 23–61
Rocha Costa JC, Dias RJP, Morenz MJF (2007) Determining the adaptation potential of entomopathogenic nematode multiplication of Heterorhabditis riobravus and Steinernema carpocapsae (Rhabditida: Heterorhabditidae, Steinernematidae) in larvae of Alphitobius diaperinus (Coleoptera: Tenebrionidae) and Galleria mellonella (Lepidoptera: Pyralidae). Parasitol Res 102:139–144
Rocha Costa JC, Dias RJP, Morenz MJF (2008) Determining the adaptation potential of entomopathogenic nematode multiplication of Heterorhabditis baujardi and Steinernema carpocapsae (Rhabditida: Heterorhabditidae, Steinernematidae) in larvae of Alphitobius diaperinus (Coleoptera: Tenebrionidae) and Galleria mellonella (Lepidoptera: Pyralidae). Parasitol Res 102:823
Rodou A, Ankrah DO, Stathopoulos C (2010) Toxins and secretion systems of Photorhabdus luminescens. Toxins 2:1250–1264
Rodrigueiro TSC, Ginarete CMA, Leite LG, Tavares FM, Goulart RM, Giacometti FHC (2008) Eficiência de Heterorhabditis indica IBCB-N05 (Rhabditida: Heterorhabditidae) no controle de Alphitobius diaperinus (Coleoptera: Tenebrionidae) sob comedouros de granja avícola. Arq Inst Biol 75:279–284
Runge GA, Blackall PJ, Casey KD (2007) Chicken litter. Issues associated with sourcing and use. Rural Industries Research and Development Corporation, Publication Nº 07/035. Australia
Salin C, Delettre YR, Cannavacciuolo M, Vernon P (2000) Spatial distribution of Alphitobius diaperinus (Panzer) (Coleoptera: Tenebrionidae) in the soil of a poultry house along a breeding cycle. Eur J Soil Biol 36:107–115
Scanes CG, Brant G, Ensminger ME (2004) Poultry science, 4th edn. Pearson Prentice Hall, Upper Saddle River
Schroeckenstein DC, Meier-Davis S, Graziano FM, Falomo A, Bush RK (1988) Occupational sensitivity to Alphitobius diaperinus (Panzer) (Lesser mealworm). J Allergy Clin Immun 82:1081–1088
Shrestha YK, Jang EK, Yu YS, Kwon M, Shin JH, Lee KY (2011) Oral toxicity of symbiotic bacteria Photorhabdus spp. against immature stages of insects. J Asia Pac Entomol 14:127–130
Skov MN, Spencer AG, Hald B, Petersen L, Nauerby B, Carstensen B, Madsen M (2004) The role of litter beetles as potential reservoir for Salmonella enterica and thermophilic Campylobacter spp. between broiler flocks. Avian Dis 48:9–18
Szalanski AL, Palmer TW, Mckay T, Steelman CD (2004) Infectivity of Steinernema spp. (Nematoda: Steinernematidae) to adult litter beetles, Alphitobius diaperinus (Coleoptera: Tenebrionidae) in the laboratory. Biocontrol Sci Technol 14:81–85
Wales AD, Carrique-Mas JJ, Rankin M, Bell B, Thind BB, Davies RH (2010) Review of the carriage of zoonotic bacteria by arthropods, with special reference to Salmonella in mites, flies and litter beetles. Zoonoses Public Health 57:299–314
Acknowledgments
This study was supported by the programme Curso de Acción para la Investigación y el Desarrollo, CAI+D, of the Universidad Nacional del Litoral.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by M. Traugott.
Rights and permissions
About this article

Cite this article
Del Valle, E.E., Frizzo, L.S., Malmierca, M. et al. Biological control of Alphitobius diaperinus with Steinernema rarum CUL and Heterorhabditis bacteriophora SMC and feasibility of application in rice hull. J Pest Sci 89, 161–170 (2016). https://doi.org/10.1007/s10340-015-0669-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10340-015-0669-8