
Abstract
Wolf packs perform group vocalizations called chorus howls. These acoustic signals have a complex structure and could be involved in functions such as strengthening of social bonds, territory advertisement, or spacing between packs. We analyzed video recordings of 46 chorus howls emitted by 10 packs of wolves held in captivity, in order to investigate whether sex, age, social status, pack, or individual influence the way wolves participate in a chorus. We found that, during a chorus, wolves vocalized 63% of the time, with the howl being the most common vocalization (36% of the chorus duration), followed by woa (13.5%), other vocalizations (11.8%), and bark (1.7%). The main factor affecting the vocal behavior of wolves was age, since young wolves vocalized less and uttered shorter acoustic signals than adults. The discriminant analysis carried out with the wolves of Cañada Real pack assigned 89.3% of the cases to the correct individual, which is much better than the assignment expected by chance, suggesting that individuals could have a unique vocal usage during a chorus howl, mainly due to the use of howls and woa-woa howls. Based on our results, we propose that in the context of a chorus the woa-woa howl is important, although further research is needed to address this issue properly.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Social animals have complex vocal repertoires comprised of multiple call types that are used to communicate with each other in different contexts for different purposes, like location of group members, group reunion, territory advertisement, and coordinate their daily activity (Baker-Médard et al. 2013; Baker 2004; Bradbury and Vehrencamp 2011; Fichtel and Manser 2010; Lehner 1982; Mech 1970). Furthermore, the group members of some species emit vocalizations simultaneously, constituting group vocalizations or choruses (Foote et al. 2011; Harrington and Asa 2003). Group signals are complex, conspicuous, and are in average longer than individual vocalizations (Aubin and Mathevon 2020; Passilongo et al. 2017). Functions attributed to group signals are related to intra-group (e.g., to strengthen social bonds) and inter-group (territorial signals) communication (Bradbury and Vehrencamp 2011; Brumm 2013). Consequently, choruses are expected to encode important information that should be essential for “the group’s survival” (Caro 2005).
The wolf (Canis lupus) is a social canid that lives in groups known as packs (Mech 1970). Essentially, the pack size ranges between 2 and 11 individuals (Fuller et al. 2003), and comprises a breeding pair and their offspring, which function as a family unit during most of the year (Mech and Boitani 2003). Acoustic communication plays an important role in wolves’ behavior. Wolf packs perform group vocalizations called chorus howls that can last 30–120 s, depending on group size (Harrington 1989; Joslin 1967). Chorus howls have been described as a vocalization in which one wolf begins howling, with other members joining in until several or all members of a pack are howling together (Joslin 1967). Howls in the context of a chorus vary more than those produced by lone individuals (Harrington 1989; McCarley 1978). Choruses can be discordant, resulting in a change in the overall structure of the chorus, and this is often linked to the presence of various group mates possibly originated by a change in physiological arousal and/or to foster group cohesion (Harrington and Asa 2003; Schassburger 1993). Chorus howls are more complex than several wolves howling simultaneously. Besides howls, other vocalizations often occur in a chorus, such as growls, barks, and squeaks (Fragnaud 2014; Harrington and Asa 2003; Holt 1998; Palacios 2015; Passilongo et al. 2017). Some authors mention a unique vocal type that is performed only in chorus contexts, called the woa-woa howl (Fragnaud 2014; Holt 1998; Palacios 2015). Traditionally, the woa-woa howl has been considered a howl variation, but differences found related to its acoustic structure suggests that it could be a different vocal type (Fragnaud 2014).
It has been suggested that chorus howls could be involved in several functions regarding intra-pack communication, for example, to reunite pack members that are temporarily distant or scattered, and to strengthen their social bonds (Harrington and Asa 2003). Chorus howls also play an important role during the breeding season. As described by Coscia et al. (1991) and Mech (1970) wolf pups start vocalizing (whines and moans) a few days after birth and start to howl at the age of 1 month. Furthermore, pups frequently join a chorus howl (Palacios et al. 2016). Reproductive females often return to the den during the breeding season and might stimulate the pups with their calls to locate them (unpublished data). The presence of pups (e.g., reproduction success) can also be detected based on their acoustic energy distribution in a chorus (Palacios et al. 2016), suggesting that wolf chorus howls could encode information regarding age composition. Wolf choruses are also used as a warning for intruders, supporting the theory of spacing between packs (Harrington and Mech 1979). For instance, wolves can signal their presence over an area ranging from at least 7.8 to nearly 129.5 km2 (Joslin 1967).
A pack of wolves consists of individuals of different social status, age, and sex. The unique composition of each pack of wolves could cause subtle variations in the structure of a chorus howl. For instance, wolves are known to be highly territorial, and group vocalizations can serve as a defensive strategy (Harrington 1989; Harrington and Asa 2003; Harrington and Mech 1979). Although the whole group can join a chorus, adults should be especially committed in the territorial defense strategies. Moreover, the physical condition of an adult animal should play in its favor to produce longer sounds, since production of sound is energy consuming (Maynard-Smith and Harper 2004). We hypothesize that adult wolves spent more time vocalizing, and emit longer vocalizations than younger individuals, especially those involved in territory advertising. Since chorus howls are also used during the breeding season to communicate with pups, we expect sex- and age-related differences in the vocal behavior of pack members during a chorus. Finally, similar to the vocal cues encoded in howls, we predict that the vocal usage of each individual should be unique and distinctive (Palacios et al. 2007; Watson et al. 2018). To test these hypotheses, we studied how different pack members contribute to a chorus howl, taking advantage of a known scenario with wolves held in captivity, with the information about gender, age, social status, and individual given by the wolf centers and wolf curators. Our research aimed to (1) describe different wolf chorus behaviors of the pack members and its relationship to group composition and (2) identify if factors like sex, age, social status, or individual have a specific effect on the time wolves spend using different vocalizations in a chorus.
Furthermore, wolf populations are often monitored through the use of howl surveys (Joslin 1967; Harrington and Mech 1983; Fuller and Sampson 1988; Thiel et al. 2009), mainly to locate packs (Harrington and Mech 1983; Fuller and Sampson 1988), detect successful reproduction (presence of pups, Palacios et al. 2016), and estimate the average number of wolves that participate in a chorus (Palacios et al. 2017; Passilongo et al. 2015). We will discuss how our results could be applied to improve the reliability of wolf surveys based on acoustic signals.
Methodology
We analyzed video recordings of chorus howls emitted by wolves held in captivity at four locations: Cañada Real (Madrid, Spain), Centro de Recuperação do Lobo Ibérico (Malveira, Portugal), Lobo Park (Málaga, Spain), and the International Wolf Centre (MN, USA). Recordings were obtained during March 2005, July–August of 2007 and 2008, and March 2017, using a digital video camera Sony Handycam DCR-HC54. The researcher simulated howls to encourage wolves to respond. Taking in consideration their highest activity point, simulations were performed during dawn and dusk. Howls were conducted from at least 5 to 30 m, often in full sight of the animals since wolves were habituated to human presence. To increase the sample size on pups we also included good quality recordings of wolves in the facilities of Bays Mountain Park (Kingsport, TN), and the Wolf Park (Battle Ground, IN) available on the internet, under the author’s permission. We analyzed a total of 46 chorus howls emitted by 10 packs comprising 30 wolves of different condition (sex, status, and age composition) from five different subspecies. The major part (80%) of the packs in our study had a parental pair in it, and the packs without one were exclusively composed by pups due to rules of the wolf center (suppl. material).
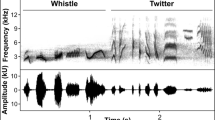
To analyze behaviors from the video recordings, we used Solomon software (Péter 2015). We conducted a focal-animal sampling (one individual is the focus of observations; Altmann 1974), analyzing every individual participating in a chorus. Previous inspection of video recordings and the repertoire of vocalizations described for the wolf (Fragnaud 2014; Holt 1998; Palacios 2015; Schassburger 1993) were used to define four sound categories (Fig. 1): howl, woa (hereafter woa-woa howls), bark (bark-like vocalizations), and other vocalizations (in our recordings, these vocalizations were mainly whines, whistles, and short harmonic sounds that could not be identified by the researcher). Solomon software allows the visual inspection of the spectrogram of the sound while watching the video images. We examined the image, audio, and spectrogram of the recordings to determine that a wolf was emitting one of the four vocal types. Sometimes it was difficult to identify the type of vocalization emitted, especially when howls and woa-woa howls were combined as a continuous vocalization.

Spectrogram frames of the vocal types of this study: a howl, b woa, c bark, and d other. Spectrogram: 1024 FFT (Fast Fourier Transform) points, Hann window
To measure the time spent emitting both vocal types, we considered, besides the sound, changes in the body positioning that were performed by the animals during the vocalization of those two sound types (video in suppl. material). In this study we considered that a wolf was (1) howling: when emitting a long, continuous acoustic signal, maintaining a static position, with the muzzle in a fix position and the canine teeth clearly visible, and (2) emitting woa-woa howls, when emitting loud, long, continuous acoustic signals, higher in pitch and frequency modulation than howls (by aural estimate), with the mouth wide open and the lips retracted during the whole woa-woa howl sound extend. Sometimes woa-woa howls were emitted as a series of discontinuous sounds. More interactions between group members occur throughout the woa-woa howl vocalization than during howling; hence, individuals change their body position frequently.
For each individual participating in a chorus, we measured five variables: (1) time spend vocalizing (vocal), (2) time emitting howls (howl), (3) time emitting woa-woa howls (woa), (4) time barking (bark), and (5) time doing other sound types (other). Wolves were at times beyond the visual range of observations; therefore, only data with animals in full sight at least 50% of the time were considered. A random sample of video recordings were analyzed by two researchers with experience in wolf vocalizations to verify that the behaviors measured were clearly distinguishable by different observers. Information of sex, age, and social status was obtained from the animal handlers. All individuals were categorized into three age classes (adult: ≥ 2 years old, yearling: 1 year old, pups: < 1 year old), but due to the sample size, pups and yearlings were grouped into subadults. For the social status category, we considered two social rank categories: breeding pair (parental pair, PP) and other pack members.
Statistical analyses
We used Linear Mixed Models (LMM) to assess which factors determine the type of vocalizations that a wolf emits during a chorus. We built models for vocal, howl, woa, bark, and other, as dependent variables, with a Gaussian distribution. We included age, status, and their interaction with sex as fixed factors, and chorus ID and individual ID nested to pack as random factors. To further explore our dataset, we grouped together short vocalizations (bark and other), and long calls (howl and woa), and built the same models for long and short vocalizations (including chorus and individual ID as random factors). In preliminary analyses we saw that the inclusion of subspecies as random effect did not vary the results; hence, we decided not to include this factor in the final models. The significance of terms was determined by a Wald statistic (p < 0.05). We estimated the variance explained by the model calculating marginal R2 (variance explained by fixed factors) and conditional R2 (variance explained by both fixed and random factors; Nakagawa and Schielzeth 2013). We also calculated the adjusted Intraclass Correlation Coefficient to estimate the proportion of the variance explained by the grouping structure in the population to get a better idea of individual variation. LMMs were conducted in R (R Development Core Team 2013), using the lmer function (in the lme4 library).
Additionally, we performed a Flexible Discriminant Analysis (FDA; Hastie et al. 1994) on the Cañada Real pack data to see if it is possible to classify individuals according to their call usage, which would support the hypothesis of the existence of individually unique vocal behaviors in a chorus. We only used the Cañada Real pack (four individuals) since we had a greater collection, and less variation in the number of recorded choruses for each individual compared to the data of the other packs in the study. We standardized the predictors included in the FDA and used the coefficients of the canonical functions as a measure of variable importance to discriminate individuals.
Results
The chorus howls analyzed lasted a minimum of 33 s, and a maximum of 185 s, with a time length of 73.95 ± 32.47 s (mean ± sd). During a chorus, wolves vocalized 63% of the time. Howling was the most performed vocalization (57.1% of the time vocalizing wolves emitted howls), followed by woa-woa howls (21.5%), other vocalizations (18.7%), and barks (2.7%). During a chorus howl, adult wolves vocalized 73.3% of the time (46.7% howling, 18.7% emitting woa, 2.3% barking, and 5.6% emitting other type of vocalizations; Fig. 2); meanwhile, subadult wolves vocalized 42.7% of the time (20.7% howling, 12.2% emitting woa, 0.8% barking, and 9% emitting other type of vocalizations; Fig. 2).

Boxplots with datapoints of each vocal type (howl, woa, bark, and other), time spent vocalizing, and doing long vocalizations (howl and woa) by individuals, according to age classes (adults and subadults)
Linear Mixed Models built for the different vocal types showed an important effect due to the random nested factor (chorus and individual within a pack) especially for the time vocalizing and emitting other vocalizations (vocal: R2c − R2m = 0.27, other: R2c − R2m = 0.24; Table 1). Among the fixed factors, age had the biggest effect on the time emitting different vocal types (Table 1). Subadults vocalized significantly less than adults (mixed model’s estimate: − 0.405, p-value: 0.001; Table 1) and emitted other vocalizations more than any type of sound. The results of the model we built for long vocalizations confirmed the differences found between age classes (subadults) with an estimate of − 0.55 and a p-value of 1.35e−05 (Table 1). Our models showed an interaction between social status and sex, in which parental females vocalized more time than parental males (estimate: 0.364, p: 0.021), spending more time emitting long vocalizations (estimate: 0.316, p: 0.048), mainly woa-woa howls (estimate: 0.412, p: 0.06, marginally significant). The FDA carried out with the wolves of Cañada Real pack (15 chorus howls), correctly assigned 89.3% of the cases to the right individual (ind1: 87%, ind2: 100%, ind3: 87%, and ind4: 87%), which is much better than the assignment expected by chance (25%; Fig. 3). The coefficients of the canonical functions showed that the usage of howls and woas were the most important variables to discriminate individuals (coefficients: howl = − 2.65, woa = − 2.23, bark = − 0.63, other = − 0.57).

Scatter plot illustrating the DFA of the vocal behavior of each individual in a chorus (data of the Cañada Real pack). Number of individuals: 1, 2, 3, and 4
Discussion
We described the vocal behaviors of the members of a pack of wolves during a chorus, and its relation to group composition. The choruses analyzed had a mean duration of 73.95 ± 32.47 s. In other studies the overall duration of a chorus measured throughout different wolf populations seems to be consistent with our results (e.g., North American wolves, Canis lupus lycaon: 59.9 ± 18.1 − mean ± SD, Harrington 1989; Iberian wolves, Canis lupus signatus: 66 ± 27 − mean ± SD, Palacios et al. 2016; Polish wolves, Canis lupus lupus: 67 ± 5 − mean ± SE, Nowak et al. 2007), suggesting that the chorus howl should be considered a group vocalization with a similar duration across wolf subspecies. Passilongo et al. (2017) identified barks, whines, growls, whimpers, squeaks, and yelps as part of this group vocalization. Previously, other authors have described squeaks, barks, growls, and miscellaneous vocalizations in a chorus (Holt 1998). Our study adds important information about the usage of calls in a wolf chorus. For instance, our findings indicate that howl was the most frequent vocal type emitted in a chorus, as reported previously by Harrington (1989), Holt (1998), and Passilongo et al. (2017). Although, so far, hardly any studies offer a proper distinction between howls and woa-woa howls (but see Fragnaud 2014), our results suggest that the woa-woa howl is a relevant vocal type in the context of a chorus (wolves spent more than 20% of the time they vocalized in a chorus emitting woa-woa howls). Woa-woa howls were first reported by Tooze (1987), and briefly described by Holt (1998), although both authors failed to clearly make a distinction between howls and woa-woa howls. More recently, Fragnaud (2014) found significant differences between woa-woa howls and howls in the mean duration, frequency range, and frequency modulation, and suggested this sound type should be categorized separately. However, we still lack a published detailed description of the woa-woa howl.
One of the goals of our study was to address the way individuals of a pack of wolves participated in a chorus howl. Previous research has focused on the vocal repertoire of the chorus howl, but to our knowledge, did not measure the contribution of each individual’s unique vocal behavior (Fragnaud 2014; Harrington 1989; Holt 1998; Passilongo et al. 2017). The most apparent finding to emerge from the analysis is that individuals within a pack have unique ways to use vocalizations in a chorus howl. A fair amount of research has been carried out searching for individual differences in specific acoustic signals across a variety of species, namely, regarding the acoustic structure of vocalizations. For instance, individual distinction in calls has been reported for meerkats (Suricatta suricatta, Townsend et al. 2010), wild agile gibbon (Hylobates agilis, Oyakawa et al. 2007), horned guan (Oreophasis derbianus, González-García et al. 2017), and coyotes (Canis latrans, Mitchell et al., 2006). In the case of wolves, Tooze et al. (1987), Palacios et al. (2007), and Root-Gutteridge et al. (2014) concluded that howls uttered by wolves have individual differences. Although not all differences found between individuals encode information for the receiver (Townsend et al. 2010), investigation of individual vocal recognition has been demonstrated in the call types of a few species, for example, elephants (McComb et al. 2000), zebra finches (Taeniopygia guttata, Elie and Theunissen 2018), and African penguins (Spheniscus demersus, Favaro et al. 2015). Our work might offer a novel approach on the chorus howl, since regardless of the acoustic features of vocal types, it highlights the fact that wolves have individually unique vocal behaviors when they participate in a chorus howl. Recent publications on the behaviors of group-living animals describe that stable societies benefit form learning and recalling the identities of their group mates, a characteristic that comes hand in hand with social complexity (Ward and Webster 2016). Wolves are a good example of group coordination and cohesion, and some activities like hunting or collaboration to ensure reproductive success could greatly benefit from recognizing your group members (Mech and Boitani 2003). Researchers have found that the acoustic behavior of wolves increases as the pups grow, suggesting that the group could profit from it for location of offspring and food (Mech and Boitani 2003). To which extent the differences in the use of vocal types we have found in our study are used for wolves to recognize and communicate with other pack mates needs further research. We believe our work proves a great potential for individual distinctiveness concerning their vocal behaviors in a chorus, given that the analysis made on this variable (Individual, DFA, and LMM’s) explained most of the data.
In line with numerous works that found differences between immature and full-grown individuals in their vocalizations, our research suggests that age class affects the different use of vocalizations emitted during a chorus (Blumstein and Daniel 2004; Mathevon et al. 2010; Volodin et al. 2016). Our results show that adults vocalize consistently during a chorus howl and emit longer sounds than younger ones. As far as we know, the adults of a pack of wolves vocalize frequently to maintain their territory free from intruders. Considering our results, and assuming that neighboring packs might perceive the degree of threat in the group vocalizations of rivals, it would make sense that adults should emit longer vocalizations than pups. Furthermore, the scarcer, shorter, and more chaotic uttering of sounds by younger animals could be explained by changes in body size, limited experience, naïve behavior, and contextual sound learning (Janik and Slater 2000). Hence, the mechanics of sound production predicts that larger, older, or bigger animals emit longer calls, with lower and less modulated fundamental frequency, than smaller ones (Ey et al. 2007). Since learning is a gradual process (Payne 1985), we would expect young individuals to mimic the sounds of their surrounding adults over time, till they gain experience, and learn to use the same signals in different contexts (Coscia et al. 1991; Janik and Slater 2000). Another plausible cause for the differences found between juveniles and adults is the caller’s competitive ability (e.g., displays of size, condition, or fighting ability) that should be greater in adults (Fitch and Hauser 2003; Reby and McComb 2003). Since younger individuals are still in development phase (weaker and smaller than adults) we could also argue that the differences found between age classes in our data goes in line with the idea that the production of some type of calls is of higher cost for individuals in poor condition than for those in good condition (Searcy and Nowicki 2005).
The presence of young individuals in a pack is the evidence of recent reproduction, which from an applied point of view is the key to monitor the status of wolf populations. The analysis of recordings of chorus howls obtained during summer-autumn is useful to determine the presence of pups (Palacios et al. 2016). However, it is not always possible to record chorus howls since the aural estimates of the presence of pups in a chorus are not always reliable (Palacios et al. 2017). Our research indicates that chorus howls that include higher proportions of short vocalizations (e.g., whines and squeaks) could reveal recent successful reproduction, which could be crucial in areas of recent wolf recolonization. Hence, our results may provide useful clues for wolf monitoring programs about the presence of young individuals in a pack.
With respect to other factors tested in this study, we found an interaction between sex and status. Parental females vocalized more and emitted more woa-woa howls than parental males. We are not sure to which extent this result could be due to a sample size artifact, an effect of the wolves’ conditions of captivity, or a confirmation of the relevance of woa-woa howls in the acoustic communication between pack members. Further research is needed to address the role of the woa-woa howls in a chorus howl.
Data availability
Authors could make the data available if requested for any questions related to the article.
Code availability
Not applicable.
References
Aubin T, Mathevon N (2020) Coding strategies in vertebrate acoustic communication, animal signals and communication. Springer International Publishing, Cham. https://doi.org/10.1007/978-3-030-39200-0
Altmann J (1974) Observational study of behavior: sampling methods. Behaviour 49:227–266. https://doi.org/10.1163/156853974X00534
Baker-Médard SA, Baker-Médard MSA, Baker MC, Logue DM (2013) Chorus song of the Indri (Indri indri: Primates, Lemuridae): group differences and analysis of within-group vocal interactions. Int J Comp Psychol 26:241–255
Baker MC (2004) The chorus song of cooperatively breeding laughing kookaburras (Coraciiformes, Halcyonidae: Dacelo novaeguineae): characterization and comparison among groups. Ethology 110:21–35. https://doi.org/10.1046/j.1439-0310.2003.00941.x
Bradbury JW, Vehrencamp SL (2011) Principles of animal communication, Second, Edi. Sinauer Associates, Inc., All, Sunderland, Massachusetts
Brumm H (2013) Animal signals and communication 2: animal communication and noise. https://doi.org/10.1007/978-3-642-41494-7
Blumstein DT, Daniel JC (2004) Yellow-bellied marmots discriminate between the alarm calls of individuals and are more responsive to calls from juveniles. Anim Behav 68:1257–1265. https://doi.org/10.1016/j.anbehav.2003.12.024
Caro T (2005) Antipredator defenses in birds and mammals. University of Chicago Press, Chicago, IL
Coscia EM, Phillips DP, Fentress JC (1991) Spectral analysis of neonatal wolf Canis lupus vocalizations. Bioacoustics 3:275–293. https://doi.org/10.1080/09524622.1991.9753190
Elie JE, Theunissen FE (2018) Zebra finches identify individuals using vocal signatures unique to each call type. Nat Commun 9:4026. https://doi.org/10.1038/s41467-018-06394-9
Ey E, Pfefferle D, Fischer J (2007) Do age- and sex-related variations reliably reflect body size in non-human primate vocalizations? A review. Primates 48:253–267. https://doi.org/10.1007/s10329-006-0033-y
Favaro L, Gamba M, Alfieri C, Pessani D, McElligott AG (2015) Vocal individuality cues in the African penguin (Spheniscus demersus): a source-filter theory approach. Sci Rep 5:17255. https://doi.org/10.1038/srep17255
Fichtel C, Manser M (2010) Vocal communication in social groups, in: Animal Behaviour: Evolution and Mechanisms. pp. 29–54. https://doi.org/10.1007/978-3-642-02624-9_2
Fitch WT, Hauser MD (2003) Unpacking honesty: vertebrate vocal production and the evolution of acoustic signals. In: Simmons AM, Popper AN, Fay RR (eds) Animal communication. Springer, Berlin, pp 65–137
Foote JR, Fitzsimmons LP, Mennill DJ, Ratcliffe LM (2011) Male black-capped chickadees begin dawn chorusing earlier in response to simulated territorial insertions. Anim Behav 81:871–877. https://doi.org/10.1016/j.anbehav.2011.01.028
Fragnaud H (2014) Vocalizations emitted by iberian wolves (Canis lupus signatus) during chorus howls. Universite Paris sud
Fuller TK, Sampson BA (1988) Evaluation of a simulated howling survey for wolves. J Wildl Manag 52:60. https://doi.org/10.2307/3801059
Fuller TK, Mech LD, Cochrane JF (2003) Wolf population dynamics. In Wolves: Behavior, Ecology, and Conservation. pp. 161–191
González-García F, Sosa-López JR, Ornelas JF, Jordano P, Rico-Gray V, Urios Moliner V (2017) Individual variation in the booming calls of captive Horned Guans (Oreophasis derbianus): an endangered Neotropical mountain bird. Bioacoustics 26:185–198. https://doi.org/10.1080/09524622.2016.1233513
Hastie T, Tibshirani R, Buja A (1994) Flexible discriminant analysis by optimal scoring. J Am Stat Assoc 89:1255–1270. https://doi.org/10.1080/01621459.1994.10476866
Harrington FH (1989) Chorus howling by wolves: acoustic structure, pack size and the beau geste effect. Bioacoustics-The Int. J Anim Sound Its Rec 2:117–136
Harrington FH, Asa CS (2003) Wolf communication. In: Mech LD, Boitani L (eds) Wolves: behavior, ecology, and conservation. University of Chicago Press, Chicago, pp 66–103
Harrington FH, Mech LD (1979) Wolf howling and its role in territory maintenance. Behaviour 68:207–249
Holt TD (1998) A structural description and reclassification of the wolf, Canis lupus, chorus howl. Dalhousie University
Janik VM, Slater PJB (2000) The different roles of social learning in vocal communication. Anim Behav 60:1–11. https://doi.org/10.1006/anbe.2000.1410
Joslin PWB (1967) Movements and home sites of timber wolves in Algonquin Park. Am Zool 7:279–288
Lehner PN (1982) Differential vocal response of coyotes to “group howl” and “group yip-howl” playbacks. J Mammal 63:675–678. https://doi.org/10.2307/1380279
Mathevon N, Koralek A, Weldele M, Glickman SE, Theunissen FE (2010) What the hyena’s laugh tells: sex, age, dominance and individual signature in the giggling call of Crocuta crocuta. BMC Ecol 10:9. https://doi.org/10.1186/1472-6785-10-9
Maynard-Smith J, Harper D (2004) Animal signals
McCarley H (1978) Vocalizations of red wolves (Canis rufus). J Mammal 59:27–35
McComb K, Moss C, Sayialel S, Baker L (2000) Unusually extensive networks of vocal recognition in African elephants. Anim Behav 59:1103–1109. https://doi.org/10.1006/anbe.2000.1406
Mech LD (1970) The wolf: the ecology and behavior of an endangered species. The Natural History Press, Garden City, New York
Mech LD, Boitani L (2003) Wolves: behavior, ecology, and conservation. University of Chicago Press
Mitchell BR, Makagon MM, Jaeger MM, Barrett RH (2006) Information content of coyote barks and howls. Bioacoustics 15:289–314. https://doi.org/10.1080/09524622.2006.9753555
Nakagawa S, Schielzeth H (2013) A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol Evol 4:133–142. https://doi.org/10.1111/j.2041-210x.2012.00261.x
Nowak S, Jędrzejewski W, Schmidt K, Theuerkauf J, Mysłajek RW, Jędrzejewska B (2007) Howling activity of free-ranging wolves (Canis lupus) in the Białowieża Primeval Forest and the Western Beskidy Mountains (Poland). J Ethol 25:231–237. https://doi.org/10.1007/s10164-006-0015-y
Oyakawa C, Koda H, Sugiura H (2007) Acoustic features contributing to the individuality of wild agile gibbon (Hylobates agilis agilis) songs. Am J Primatol 69:777–790. https://doi.org/10.1002/ajp
Palacios V, Font E, Márquez R (2007) Iberian wolf howls: acoustic structure, individual variation, and a comparison with north american populations. J Mammal 88:606–613. https://doi.org/10.1644/06-MAMM-A-151R1.1
Palacios V (2015) Vocal communication in the iberian wolf. Universidad de Valencia
Palacios V, López-Bao JV, Llaneza L, Fernández C, Font E (2016) Decoding group vocalizations: the acoustic energy distribution of chorus howls is useful to determine wolf reproduction. PLoS ONE 11:1–12. https://doi.org/10.1371/journal.pone.0153858
Palacios V, Font E, García EJ, Svensson L, Llaneza L, Frank J, López-Bao JV (2017) Reliability of human estimates of the presence of pups and the number of wolves vocalizing in chorus howls: implications for decision-making processes. Eur J Wildl Res 63:59. https://doi.org/10.1007/s10344-017-1115-4
Passilongo D, Mattioli L, Bassi E, Szabó L, Apollonio M (2015) Visualizing sound: counting wolves by using a spectral view of the chorus howling Visualizing sound : counting wolves by using a spectral view of the chorus howling. Front Zool 9. https://doi.org/10.1186/s12983-015-0114-0
Passilongo D, Marchetto M, Appolonio M (2017) Singing in a wolf chorus: structure and complexity of a multicomponent acoustic behaviour. Hystrix. Ital J Mammal 28:1–6. https://doi.org/10.4404/hystrix-28.2-12019
Payne R (1985) Large scale changes over 19 years in songs of humpback whales in Bermuda. Z Tierpsychol 68:89–114. https://doi.org/10.1111/j.1439-0310.1985.tb00118.x
Péter A (2015) Solomon Coder© A simple and free solution for behavior coding. https://solomon.andraspeter.com/
R Development Core Team 3.0.1 (2013) A Language and Environment for Statistical Computing. R Found Stat Comput
Reby D, McComb K (2003) Anatomical constraints generate honesty: acoustic cues to age and weight in the roars of red deer stags. Anim Behav 65:519–530. https://doi.org/10.1006/anbe.2003.2078
Searcy WA, Nowicki S (2005) The evolution of animal communication
Schassburger RM (1993) Vocal communication in the timber wolf, Canis lupus, Linnaeus. Adv Ethol 30
Thiel RP, Hall W, Heilhecker E, Wydeven AP (2009) An isolated wolf population in central Wisconsin, in: Recovery of Gray Wolves in the Great Lakes Region of the United States. Springer New York pp. 107–117. https://doi.org/10.1007/978-0-387-85952-1_7
Tooze ZJ (1987) Some aspects of the structure and function of long-distance vocalizations of timber wolves (Canis lupus). Dalhouise University, Halifax, Nova Scotia
Townsend SW, Hollén LI, Manser MB (2010) Meerkat close calls encode group-specific signatures, but receivers fail to discriminate. Anim Behav 80:133–138. https://doi.org/10.1016/j.anbehav.2010.04.010
Volodin IA, Sibiryakova OV, Volodina EV (2016) Sex and age-class differences in calls of Siberian wapiti Cervus elaphus sibiricus. Mamm Biol 81:10–20. https://doi.org/10.1016/j.mambio.2015.09.002
Ward A, Webster M (2016) Sociality: the behaviour of group-living animals, sociality: the behaviour of group-living animals. Springer International Publishing, Cham. https://doi.org/10.1007/978-3-319-28585-6
Watson SK, Townsend SW, Range F (2018) Wolf howls encode both sender- and context-specific information. Anim Behav 145:59–66. https://doi.org/10.1016/j.anbehav.2018.09.005
Harrington FH, Mech LD (1983) Wolf pack spacing: howling as a territory- independent spacing mechanism in a territorial population. Behav Ecol Sociobiol 12:161–168
Root-Gutteridge H, Bencsik M, Chebli M, Gentle LK, Terrell-Nield C, Bourit A, Yarnell RW (2014) Improving individual identification in captive Eastern grey wolves (Canis lupus lycaon) using the time course of howl amplitudes. Bioacoustics 23(1):39–53. https://doi.org/10.1080/09524622.2013.817318
Acknowledgements
We thank F. Petrucci-Fonseca and S. Pinho (C.R.L.I., Malveira, Portugal), J. Escudero (La Dehesa), P. España and E. Sánchez (Cañada Real, Madrid, Spain), D. Weigend (Lobo Park, Málaga, Spain), and Lori Schmidt (International Wolf Center, Minnesota), for allowing us to obtain recordings from the captive wolves under their care. We also thank Bays Mountain Park (Kingsport, TN) and Jay Huron and the Wolf Park (Battle Ground, IN) for their permission for using some of their recordings of pups howling in a chorus. The author from CITAB was supported by National Funds by FCT—Portuguese Foundation for Science and Technology, under the project UIDB/04033/2020.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Ethics approval
All appropriate ethics and other approvals were obtained for the research.
Consent for publication
The manuscript is not being considered for publication elsewhere while it is being considered for publication in this journal and it has not been previously submitted to this journal.
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary file1 (MP4 7870 KB)
Rights and permissions
About this article

Cite this article
Marti-Domken, B., Sanchez, V.P. & Monzón, A. Pack members shape the acoustic structure of a wolf chorus. acta ethol 25, 79–87 (2022). https://doi.org/10.1007/s10211-021-00388-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10211-021-00388-5