
Abstract
Vocalizations are a vital form of communication. Call structure and use may change depending on emotional arousal, behavioral context, sex, or social complexity. Pithecia chrysocephala (golden-faced sakis) are a little-studied Neotropical species. We aimed to determine the vocal repertoire of P. chrysocephala and the influence of context on call structure. We collected data June–August 2018 in an urban secondary forest fragment in Manaus, Amazonian Brazil. We took continuous vocal recordings in 10-min blocks with 5-min breaks during daily follows of two groups. We recorded scan samples of group behavior at the start and end of blocks and used ad libitum behavioral recording during blocks. We collected 70 h of data and analyzed 1500 calls. Lowest frequencies ranged 690.1–5879 Hz in adults/subadults and 5393.6–9497.8Hz in the only juvenile sampled. We identified eight calls, three of which were juvenile specific. We found that, while repertoire size was similar to that of other New World monkeys of similar group size and structure, it also resembled those with larger group sizes and different social structures. The durations of Chuck calls were shorter for feeding contexts compared to hostile, but frequencies were higher than predicted if call structure reflects motivation. This finding may be due to the higher arousal involved in hostile situations, or because P. chrysocephala use Chuck calls in appeasement, similar to behavior seen in other primates. Call structures did not differ between sexes, potentially linked to the limited size dimorphism in this species. Our findings provide a foundation for further investigation of Pithecia vocal behavior and phylogeny, as well as applications for both captive welfare (stress relief) and field research (playbacks for surveys).
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The ability to vocalize is an important aspect of animal behavior, allowing individuals to convey information to others concerning their social relationships, status, age, sex, the location of resources, and the presence of potential threats (e.g., Strier 2007). This information, in turn, permits conspecifics and heterospecifics to respond appropriately (e.g., fleeing, heading toward a food source) (Bradbury and Vehrencamp 1998). Phylogenetic relationships play a role in the development of primate vocalizations. For example, the usage and structural properties of alarm calls in Cercobeus torquatus atys is similar to that of the closely related Cecrocebus galeritus (Range and Fischer 2004; Waser 1982). In addition, vocal repertoire sizes of Gorilla gorilla and Gorilla beringei beringei are similar (Salmi et al., 2013). A correlation also exists between the structural similarities within the loud calls of surilis, Presbytis spp., and data on their genetic distance (Meyer et al. 2012).
A positive association between vocal and social complexity exists in a variety of primate species (e.g., Bouchet et al. 2013; McComb and Semple 2005). This association support the social complexity hypothesis, which proposes that with increasingly well-developed social structure (e.g., larger group size, distinct social roles, stable social network) there is selection for increasingly well-developed vocal behavior (e.g., larger vocal repertoire, individual distinctiveness, diverse use across contexts) (Bradbury and Verhencamp 1998; Freeberg et al. 2012). Such vocal behavior adaptations provide flexibility that allows individuals to cope with the challenges of more complicated social arrangements, by allowing more detailed communication. For example, much larger repertoires are present in Cebus capuchinus (27 calls, group size 16–21) and Saimiri sciureus (52 calls, group size 15–75) than in Aotus spp. (4–6 calls, group size 2–6) (Andrew 1963; Campbell et al. 2011; Gros-Louis et al. 2008; Kantha et al. 2009; Moynihan 1964; Schott 1975).
A variety of external and internal factors can alter the spectrotemporal features of vocalizations, including age, context, and sex. For example, juveniles have specific calls in multiple animal groups such as primates, bats, and rodents (Moss et al. 1997; Sokoloff and Blumberg 2004; Zimmerman 1995), which they typically use to request help from caregivers (Redican 1975), although some use calls to coordinate play behavior (Burke et al. 2018). Infant calls often have higher frequencies than adult ones, which is likely due to their smaller body size (e.g., Fischer et al. 2002; Inoue 1988) and is an effective indicator of age. In addition, juvenile calls may be shorter (e.g., Hradec et al. 2017). Owing to differences in anatomical features such as lung capacity, vocal tract length, and vocal fold size and thickness, smaller animals should produce calls that are shorter and with a higher fundamental frequency (Ey et al. 2007).
Context can also have a major influence on the structure and use of vocalizations. For example, the motivational-structural rule proposes that calls that have a hostile or friendly intention differ in form, with hostile calls being harsh and low frequency to suggest size to an opponent, while friendly or appeasement calls are more tonal and higher in pitch (Morton 1977). This is hypothesized to have evolved to avoid physical conflict, thus saving energy and reducing the risk of physical harm (Morton 1977). This relationship between call structure and context may also reflect the caller’s emotional state. In times of high emotional arousal, respiration, salivation, and muscular tension change, resulting in changes to fundamental frequencies, durations, and rates of vocalizations (Pollermann and Archinard 2002; Titze 1994).
Sex differences in vocalizations occur in many species. For example, sex differences exist in the lowest frequency, peak frequency, and end frequency of phee calls in Callithrix kuhlii (Smith et al. 2009). Similarly, male alarm barks in Papio ursinus are of a lower frequency and longer than those made by females (Fischer et al. 2002), while female juvenile Macaca mulatta emit more complex vocalizations and longer calls than male juveniles (Tomaszycki et al. 2001). Similar to the structural differences expected from adults and juveniles due to size differences, we can predict that species showing sexual dimorphism should display differences in call structure. Evidence to support this prediction exists across various primate species (Ey et al. 2007). However, an exception to this trend exists in Leontopithecus rosalia, which show no external sexual dimorphism but display structural differences in calls between sexes (Benz et al. 1990). These differences may result from differences in the medial ventral laryngeal sac, which is larger in males than in females (Hershkovitz 1977).
We have limited information on the ecology and behavior of Pithecia species, likely due to the difficulties involved in conducting studies of wild populations, such as their often cryptic behavior and coloration, resistance to habituation, and low population densities (Ayres 1986; Mittermeier and Roosmalen 1981; Norconk 2006; Pinto et al. 2013; Vie et al. 2001). Pithecia occupy home ranges of 10.3–200 ha and live in groups of 2–12 individuals (Izawa and Yonada 1981; Lehman et al. 2001; Norconk and Setz, 2013). These groups may include multiple breeding males and females or consist only of a breeding pair and young (e.g., Lehman et al. 2001; Setz and Gaspar 1997). They are territorial, with some studies suggesting they use long calls to defend their home ranges (DiFiore et al. 2007; Norconk 2006, 2011). The IUCN currently consider Pithecia spp. as Least Concern (Mittermeier et al. 2015). However, all Pithecia species are likely impacted by hunting, habitat fragmentation, and anthropogenic disturbance (Marsh 2014; Norconk and Setz, 2013). The recent revision of the taxonomy of the genus Pithecia describes 16 full species where there were previously 5, creating a need for further research on these species, including their vocalizations (Bezerra et al. 2017; Marsh 2014). Previous studies of vocal behavior in Pithecia have described 7–18 call types in Pithecia pithecia and Pithecia aequatorialis (Buchanan 1978; Henline 2007; Keiran 2012).
Although vocal repertoires exist for Pithecia species, there are no published studies examining vocal communication in this group. Accordingly, here, we aimed to add to the current understanding of Neotropical primate communication by investigating the behavior of Pithecia chrysocephala (golden-faced sakis) living in a forest fragment within the city of Manaus, Amazonian Brazil. Pithecia species can survive in isolated forest fragments and such areas are useful because it is easier to find and follow groups (Boyle 2016; Ferrari et al. 2003; Pinto et al. 2013), although the primates may show behaviors that differ from those of their counterparts in continuous forests (Schwitzer et al. 2011).
Our goals were to determine the vocal repertoire of Pithecia chrysocephala, investigate the behavioral contexts in which they use calls, and examine the association between behavioral context and call structure. We tested predictions derived from five hypotheses:
- 1)
If congeneric species have similar vocal repertoires (see Henline 2007 for Pithecia), then the vocal repertoire of P. chrysocephala will consist of 7–18 calls.
- 2)
If the P. chrysocephala vocal repertoire fits the Social Complexity Hypothesis (Freeberg et al. 2012), then vocal repertoire size will be comparable to that of other New World species with similar group sizes and family structure, such as Aotus spp. and Callicebinae.
- 3)
If age affects call structure (e.g., Zimmermann 1995), then P. chrysocephala will have juvenile-specific calls that are structurally different from those of adult males and females.
- 4)
If call structure reflects motivation (Morton 1977), then calls used in hostile contexts will have lower frequencies than those used in nonhostile contexts.
- 5)
If calls reflect the physical size of an animal (Ey et al. (2007), then males will produce calls with lower fundamental frequencies and longer durations than females.
Methods
Study Site and Groups
We conducted the study in a natural forest fragment located on the Instituto Nacional de Pesquisas da Amazonas (INPA) Campus-II in the central-western area of Manaus, Amazonian Brazil (3′′05′S, 59′′59′W) (Fig. 1). The 20-ha fragment is composed of secondary terra firme forest, bordered by tarmacked roads. Part of the area (13 ha) is open to the public, with a main pathway laid through the fragment, while several INPA offices occupy the remainder. This tropical region experiences a dry season (June–November) and a wet season (December–May), with a mean annual rainfall of 2420 mm and mean annual temperature of 26.7°C (Alvares et al. 2013).

Map of the study site, a forest fragment in INPA Campus II, Manaus, Amazonian Brazil.
Two groups of Pithecia chrysocephala lived at the study site: group A with eight individuals (two adult females, two adult males, three subadults, and one female juvenile ca. 1 yr old) and group B with 10 (two adult females, three adult males, and five subadults). We defined adults as individuals >6 yr old, subadults as 4–5 yr old, juveniles as 1–3 yr old, and infants as 0–1 yr old (M. Take pers. comm.). Group A occupies a home range of 7.68 ha and group B 8.6 ha (Take 2017). Both groups consist of a breeding pair and their offspring. They are habituated to human presence as a result of general human visitation and previous primatological work (Take 2017).
Data Collection
JM collected data during the early dry season, from June 15 to August 1, 2018, using daily follows 06.00–11.00 h and 11.00–16.00 h, alternating each day. On 06.00–11.00 h shifts, we first attempted to locate the sleeping tree of the group studied on the 11.00–16.00 h shift the previous day. Once we located the group, we conducted observations in 10-min blocks, with 5-min breaks between blocks. For 11.00–16.00 h shifts we followed the groups until they had settled at a sleeping site and remained there for 30 min at rest. We conducted 18 shifts (ca. 90 h), equally distributed between morning and afternoon shifts. We noted the location of the sleeping site and used this as a starting point the next day. We alternated the groups followed each week (group A for 34.8 h, group B for 31.2 h). During observation blocks we recorded vocalizations continuously in WAV format using a ZOOM H4n recorder (16-bit res, sampling rate of 44 Hz) with a Yoga HT-81 microphone for unidirectional recording (frequency response 1–16 kHz) (Martin and Bateson 2007). During recordings we directed the microphone toward where most (>50%) of the group were located. We maintained a distance of 2–10 m from the subjects to avoid overloading the microphone or disturbing them.
At the beginning and end of each observation block, we took a scan sample of the behavior of all visible members of the group to determine the group’s main activity (i.e., the behavior that 50% or more of the group were performing; Altmann 1974). We used the categories of general (rest, locomotion, feeding/foraging, alert), social (affiliative, play, grooming), and conflict (food conflict and intragroup conflict) based on an ethogram in Buchanan (1978). We did not take scan samples if fewer than half the group were visible. We also noted any calls that could be attributed to a specific individual in the group, detailing their age group (adult/juvenile), sex, and the behavioral context in which the call was performed ad libitum during the blocks. Additional categories used to describe juvenile behavior included parent–offspring interactions, which included nursing and requesting (in which the juvenile called repeatedly until a member of the group attended to her, often by feeding or carrying). It was not always possible to associate a call with an individual or note its age and sex.
We did not conduct tests of intraobserver reliability, but JM collected all data within a short period.
Potential Limitations
Our results must be viewed with caution for four reasons. First, the adult vocalization data are pseudo-replicates, because we could not always distinguish individuals when recording data and we do not know how much each individual contributed to the sample. Second, as only one juvenile was present, this individual was the sole source for juvenile vocal data and so results for this aspect are preliminary. Third, we could not distinguish subadults from adults owing to similarities in body size and pelage color, so we describe them as adult/subadult. Finally, the study area is in an urban fragment, so we may not have observed full behavioral or vocal repertoires (e.g., owing to lack of predation or threats; Barnett et al. 2017).
Data Manipulation
Using the spectrograms, we chose only calls of high quality for measurement after removing those with excessive background interference. We also excluded overlapping calls. When we used sequences of calls, we considered each element as an individual call. We took measurements of duration, frequency of maximum energy (FME), lowest frequency, start frequency, and end frequency for each call type using Praat (V 6.0.5) (Boersma 2001) (Fig. 2). We used the fast Fourier transform method to generate spectrograms with a Gaussian window shape and a sampling frequency of 44,100 Hz. We used a view range of 7520,000 Hz. We tested these variables for normality using a Shapiro–Wilk test, and found that they had nonparametric distributions. We calculated the median and interquartile range (Q1–Q3) for each measurement across each call type and for each behavioral context in which a call occurred (e.g., median call duration during locomotion).

Measurements used to describe Pithecia chrysocephala vocalizations in Manaus, Brazil, between June and August 2018.
We grouped contexts into friendly (e.g., play, feeding, locomotion) and hostile (e.g., intragroup conflict, food conflict) intention for statistical comparison.
Statistical Analyses
We recorded 70 h of vocal data and used 1500 calls for analysis. We could distinguish sex for 70 calls (34 female calls, 36 male).
JM performed all statistical analyses using SPSS v.25. We grouped calls according to their structural differences in spectrograms, their duration, frequency of maximum energy, lowest frequency, start frequency, end frequency, and by ear. When naming calls, we used Henline’s (2007) categories for Pithecia pithecia calls where appropriate; otherwise we chose new names onomatopoetically. We used a cross-validated discriminant function analysis (DFA) to assess the reliability of the recorded vocal repertoires.
To determine the effect of behavioral context on call structure, we compared chuck call parameters across feeding/conflict contexts using a cross-validated DFA. We chose this call because it was the most evenly spread between contexts. We did not test juvenile calls for contextual differences, as sample sizes between contexts were not large enough for valid analysis. We used Mann–Whitney U tests to determine the effect of sex on call structures of whistles, chucks, and trills. We chose these calls because they were the only calls in which we identified both male and female callers.
Ethical Note
We obtained ethical clearance from Oxford Brookes University before beginning data collection. We also obtained additional permission from INPA to study Pithecia chrysocephala in the area, and all research complied with Brazilian law. While conducting this study we took care to adhere to the guidelines for the treatment of animals in research outlined by the Association for the Study of Animal Behaviour (Buchanan et al. 2012). We did not trap or handle animals, and as such the potential for zoonotic disease transmission was minimal. During daily follows we maintained minimum distance of 2 m from individuals to minimize stress for the animals, and we retreated if they showed excessive stress (e.g., emitting alarm calls while looking at the researcher). The authors declare that they have no conflict of interest.
Data Availability
The data used for this study are available from the authors upon request.
Results
Adult/Subadult Vocal Repertoire
We identified five structurally different adult/subadult call types (Fig. 3, Table I).

Spectrograms of calls by adult/subadult Pithecia chrysocephala in Manaus, Brazil, June–August 2018, including whistle(a), chuck(b), trill(c), soft growl(d), and multiple throat rattle(e) calls. We rarely heard soft growl calls and could not obtain clearer spectrograms.
Whistles were simple, unmodulated sounds.
Chucks were very short, sharp, and unmodulated calls.
Trills were long and often slightly modulated.
We heard the soft growl only four times over the data collection period and only in males. Soft growls were the least variable and lowest frequency calls in the adult/subadult repertoire.
The throat rattle was a low-frequency call, and the longest in the adult/subadult repertoire.
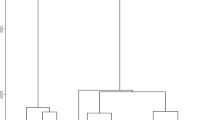
Calls were correctly classified 85% of the time by DFA with leave-one-out cross-validation (N = 5 call types and N = 1343 calls). Each call type was correctly classified >80% of the time (Table II, Fig. 4). All call parameters contributed significantly to the DFA (Duration: Wilk’s λ = 0.332, P < 0.001; FME: Wilk’s λ = 0.933, P < 0.001, lowest frequency: Wilk’s λ = 0.528, P < 0.001; start frequency: Wilk’s λ = 0.506, P <0.001; end frequency: Wilk’s λ = 0.674, P < 0.001).

Distribution of discriminant scores for adult/subadult Pithecia chrysocephala call types along two canonical discriminant functions in Manaus, Brazil, June–August 2018.
Juvenile Vocal Repertoire
We identified six calls emitted by the juvenile (Fig. 5).

Spectrogram of calls by one juvenile Pithecia chrysocephala in Manaus, Brazil, June–August 2018, including whistle(a), chuck(b), trill(c), trew(d), peep(e), and cry(f) calls. We rarely heard trew calls, and so could not obtain clearer spectrograms.
Juvenile whistles and chucks were similar in structure to those of adults/subadults but always at a higher frequency (Table III).
The trill was similar to a lower frequency adult/subadult trill.
The trew was a short and low-frequency call.
Cries and peeps were both modulated high-frequency calls, with peeps increasing in frequency and cries increasing and curving back down on spectrograms.
Calls were correctly classified 54.8% of the time by DFA with leave-one-out cross-validation (N = 6 call types and N = 126 calls). Each call type was correctly classified >37% of the time, with the exception of trills, which were 0% correct (Table IV, Fig. 6). All call parameters contributed significantly to the DFA (Duration: Wilk’s λ = 0.754, P < 0.001; FME: Wilk’s λ = 0.401, P < 0.001, lowest frequency: Wilk’s λ = 0.449, P < 0.001; start frequency: Wilk’s λ = 0.489, P < 0.001; end frequency: Wilk’s λ = 0.394, P < 0.001).We identified six calls emitted by the juvenile (Fig. 5).

Distribution of the discriminant scores for call types in one juvenile Pithecia chrysocephala along two canonical discriminant functions in Manaus, Brazil, June–August 2018.
Context and Relationship to Call Structure
Both adult and juvenile calls varied in their context (Tables V and VI). Adult/subadult whistle calls were present in the widest variety of contexts of all adult call types and were the only calls used during grooming. Individuals used these calls alone but also in quick succession with other whistles in play or locomotion contexts, and with chucks during feeding. Adult/subadult chucks varied in their dominant frequency and context, with situations such as territorial conflicts with the other group eliciting higher dominant frequency and longer calls than in less distressing situations such as feeding. Individuals often used chuck calls in succession during play or as an alert (e.g., individuals frequently used them when researchers approached at the start of data collection). The juvenile used whistle and chuck calls in a similar way to adults/subadults, with the addition of when requesting help from adults/subadults (e.g., food or for adults to carry them) (Table V). The use of trills was much less variable across contexts when used by the juvenile compared to adults/subadults, appearing in only one context compared with five in adults/subadults. We typically heard soft growls before conflicts between groups, usually when the other group was nearby but out of the sakis’ sight. We visually confirmed the presence of the other group. Pithecia chrysocephala typically performed throat rattle calls several times in succession, decreasing in dominant frequency over time and often synchronized with other group members in combination with piloerection and branch-shaking displays. We observed soft growls and throat rattles exclusively during conflict with the other saki group. We observed juvenile trew, cry, and peep calls only in social contexts and in succession until the behavior observed ended. While peeps and cries were both used in request contexts, cries were emitted in more apparently distressing situations such as after falling from a tree or when the juvenile could not cross a gap between two trees unaided, whereas peeps were present for less apparently stressful requests, such as for food or to for adults to carry them.
The structure of feeding and conflict chuck calls varied with context (Table VII). Call context was correctly classified 66.3% of the time by DFA with a leave-one-out cross-validation (N = 2 contexts and N = 163 calls). Each call type was correctly classified >62% of the time (Table VIII, Fig. 7). Only duration contributed significantly to the DFA (Wilk’s λ = 0.791, P < 0.001).

Distribution of the discriminant scores for adult/subadult Pithecia chrysocephala chuck calls along the discriminant function (duration) in Manaus, Brazil, June–August 2018. Bars show the behavioral context, boxes show the interquartile range of call durations, and whiskers show the minimum and maximum durations.
The Relationship Between Call Structure and Sex
Call structure differed significantly for chucks in terms of duration (Mann–Whitney U test: U = 11.5, P = 0.02, N = 17) but not FME (U = 18, P = 0.09, N = 17), lowest frequency (U = 20.5, P = 0.156, N = 17), start frequency (U = 16.5, P = 0.07, N = 17), or end frequency (U = 22.5, P = 0.221, N = 17) (see Table IX for acoustic measurements). We found no significant sex differences in duration (U = 39, P = 0.487, N = 20), FME (U = 37, P = 0.396, N = 20), lowest frequency (U = 42.5, P = 0.67, N = 20), start frequency (U = 42.5, P = 0.671, N = 20), and end frequency (U = 43, P = 0.699, N = 20) in trills. Similarly, we found no significant sex differences in duration (U = 75, P = 0.643, N = 26), FME (U = 68.5, P = 0.425, N = 26), lowest frequency (U = 62, P = 0.257, N = 26), start frequency (U = 63.5, P = 0.291, N = 26), and end frequency (U = 67, P = 0.403, N = 26) in whistles.
Discussion
Adult/Subadult Vocal Repertoire
We identified five calls for adult/subadult Pithecia chrysocephala, which differs from the predicted 7–18 calls based on the work of Buchanan (1978) and Henline (2007) on P. pithecia, and Keiran (2012) on P. aequatorialis. This smaller repertoire is potentially due to the amount of gradation seen in whistle, chuck, and trill calls, which we could not objectively categorise into separate calls. In Henline (2007), chucks are split into chucks and churks. It is possible that Henline (2007) overlooked gradation between calls because of their smaller sample sizes, meaning that calls appearing as several groups rather than variations in one group. We also noted soft growls and throat rattles, two other calls recorded by Henline (2007) in the current study. Only one call from Henline (2007) was not present; the Z-trill, a low-frequency call used when an individual became separated from the group. This may not have been present during this study owing to masking from background noise or simply because individuals did not stray far from the group.
Comparing spectrograms, only chuck calls were visually similar in Pithecia pithecia and P. chrysocephala. Throat rattle and scream are similar in P. chrysocephala and P. aequatorialis, as are the juvenile cry and yip. The calls of P. aequatorialis are less visually similar to P. chrysocephala than P. pithecia, potentially reflecting the geographical and evolutionary distance between these species, with the range of P. aequatorialis being further from P. chrysocephala than that of P. pithecia (IUCN 2019).
Outside Pithecia species, similar repertoire sizes exist in Aotus species (4–6 calls) and species from the Callicebinae, (6–13), two groups with group sizes and structure similar to those of Pithecia (Andrew 1963; Cäser et al. 2012; Kantha et al. 2009; Kinzey et al. 1977; Moynihan 1964, 1966; Robinson 1979; ). This provides some support for the social complexity hypothesis. However, other groups with more complex social structures and larger groups also show similar repertoire sizes, such as uacaris, Cacajao spp. (9–12 calls) and brown woolly monkeys, Lagothrix lagothricha (6), suggesting that other factors may have influenced the evolution of call repertoire sizes in New World monkeys (Bezerra et al. 2010; Casamitjana 2002; Fontaine 1981). Another possibility is that these repertoires are similarly complex, but in different ways. For example, P. chrysocephala have smaller but more stable groups in terms of size and composition, while the larger Cacajao spp. groups show high fission–fusion dynamics and spend most time with a smaller subunit (7 individuals in Cacajao ouakary: Barnett et al. 2018). Increased vocal complexity may have evolved in P. chrysocephala to deal with the social complexity of interacting with the same individuals consistently over time, which could be comparable to the level of complexity Cacajao spp. face living in subunits (Freeberg et al. 2012).
Of the calls we recorded, throat rattles were the longest, while chucks were the shortest. In terms of frequencies, trills were highest and soft growls lowest. The structure of these calls and the contexts in which Pithecia chrysocephala uses them is consistent with the hypothesis that high-frequency calls are typically friendly, and low-frequency calls are hostile (Morton 1977). The long durations of throat rattle calls also agree with evidence that longer calls are used in high-stress situations (Briefer 2012). Individuals did not use soft growls when in direct visual contact with other groups, and so this call may function to maintain the spacing between the two groups at a distance, similar to the loud calls of Callicebinae spp. (Robinson 1981).
Among the calls, whistles, chucks, and trills were present in a wide number of contexts, while throat rattles and soft growls were used specifically in intragroup conflicts. whistles, chucks, and trills were also the only calls to show gradation, which may be associated with their more variable use, allowing communication of more detail across differing contexts (Green 1975).
Juvenile Vocal Repertoire
The juvenile repertoire of Pithecia chrysocephala comprised six calls. No juvenile calls are described by Buchanan (1978), Henline (2007), or Keiran (2012), so we did not base juvenile call names and groupings on previous works. Similar to adult P. chrysocephala, juveniles performed graded chucks and whistles across a variety of contexts. Juvenile trills are split into two calls: trills resemble the adult version, and trews are used exclusively during nursing. The juvenile trill did not appear in several contexts, unlike in the adult version, possibly as the establishment of this call in their repertoire had not advanced as much as whistles and chucks by the stage of development the juvenile had reached when we made the recordings (e.g., Snowdon and Elowson 2001). Juvenile equivalents of adult whistles were longer and higher in frequency than adult calls, while chucks were shorter and higher and trills shorter and lower than adult calls. The higher frequencies are likely due to the smaller body size of juveniles (e.g., Hauser 1993). The lower frequency of trills compared to those in adults is not as predicted, but could be due to the trill calls becoming more distinct from structurally similar trews, which are low-frequency calls.
The only juvenile exclusive calls we recorded were the trew, peep, and cry vocalizations. Trew calls were low and repetitive, similar to a purr and used only while nursing. Cacajao calvus, Leontopithecus rosalia, and mantled howler monkeys (Alouatta palliata) display similar calls (Carpenter 1934; Fontaine 1981; Kleiman et al. 1988). In these cases, the purr-like vocalizations are also present in a parental care context, possibly functioning to solicit nursing from the mother or communicate contentment (Bradshaw and Cameron-Beaumont 2000; Leyhausen 1979). Peeps were the highest frequency calls in the juvenile repertoire and are present in resting, social, and request contexts. Similar to adult trills, the highest frequency call is present only in friendly contexts (Morton 1977). The structure of the juvenile cry is in line with the finding that infant distress calls are often a “chevron” shape on spectrograms, which is common across multiple mammal species (Lingle et al. 2012). However, cry calls are not as high frequency as expected for a distress call, potentially because the call must be low enough to travel when from separated infants to the group.
The Relationship Between Context and Call Structure
There was a structural difference in chuck calls between feeding and hostile contexts. However, this occurred only in terms of call duration. This finding did not support our prediction that more hostile calls would be lower in frequency than friendly ones (Morton 1977). The higher stress level of the situation could explain the higher than expected call frequency during hostile contexts (e.g., Linhart et al., 2015). The chuck call in Pithecia chrysocephala is similar in use to the tchó call in golden-backed uacaris, Cacajao ouakary, which also occurred at higher than expected frequencies in hostile contexts (Bezerra et al. 2010). The authors proposed that this finding was due to the call being used to communicate appeasement rather than hostility in conflict situations. If this is the case for chuck calls, then they support the motivational–structural rule.
The Relationship Between Call Structure and Sex
Call structures for whistle and trill calls did not differ across sex, contra our prediction that males would produce longer calls with lower fundamental frequencies than females do. We found partial support for our predictions for chuck calls, which were significantly different in duration between sexes, but not across other parameters. This similarity in calls between sexes may be due to the sex difference in body mass being only mild to moderate (Norconk 2011). However, other primate species with little size dimorphism display differences in call structure, such as Leontopithecus rosalia, and Callithrix jacchus (Benz et al.1990; Norcross et al. 1999). One explanation for this could be that these species show internal differences which alter call structure, as is the case for L. rosalia (Hershkovtiz 1977). Another possibility is that structural differences do exist for Pithecia chrysocephala, but in parameters we did not measure, such as frequency range and highest frequency.
Potential Applications
Our findings have several potential applications. Monitoring the use of chuck calls in captive groups could act as a means of assessing stress levels, as this is primarily an alert/conflict call. Similar to those in captive Sapajus apella, higher frequency chucks and more frequent use could indicate higher stress levels (Jacobsen et al. 2010). Vocalizations are also applicable as enrichment, as carried out with Callithrix jacchus (Watson 2014), to encourage more positive behaviors. In this case, playing a group’s whistle and trill calls back to them could support behaviors such as grooming and play. However, excessive grooming is undesirable, so playback should be brief. Additional research on the use of whistles and trills across contexts is necessary, as using the wrong calls could cause distress. Soft growl calls from another group could also be used as enrichment to encourage natural behaviors such as territorial defense (e.g., Farmer 2011). While these calls are less ambiguous than whistles and trills, their use is for intergroup encounters and could increase stress levels.
The IUCN recommend a new red list assessment for Pithecia chrysocephala since its classification as a full species (Mittermeier and Roosmalen 2015). The population trends of this species are currently unknown, and while currently considered Least Concern they occupy an area experiencing rapid deforestation (INPE 2016). Knowledge of vocalizations could be used to conduct population censuses of this species without relying on visual contact. This could be highly beneficial, as sakis can be difficult to follow visually in the wild (Pinto et al. 2013). Censuses could also use playbacks of calls, but, as this species is territorial, hearing another group’s calls could cause distress.
Future Studies and Conclusion
Our study provides information previously unknown on the vocalization of Pithecia chrysocephala. Our findings offer a foundation for future studies into the vocalizations and behavior of P. chrysocephala, such as investigations into vocal development and further examination of the association between context and call structure, as well as more data for comparisons of vocal behavior across the Pitheciidae. There is also potential for using our results in captive welfare and field research on P. chrysocephala, potentially to monitor stress levels, enrich the captive environment, or make censuses of these cryptic animals easier.
References
Altmann, J. (1974). Observational study of behaviour: Sampling methods. Behaviour, 49, 227–266.
Alvares, C. A., Stape, J. L., Sentelhas, P. C., de Moraes, G., Leonardo, J., & Sparovek, G. (2013). Köppen’s climate classification map for Brazil. Meteorologische Zeitschrift, 22, 711–728.
Andrew, R. J. (1963). The origin and evolution of the calls and facial expressions of the primates. Behaviour, 20, 1–109.
Ayres, J. M. (1986). Uakaris and Amazonian flooded forest. PhD thesis, University of Cambridge.
Barnett, A. A., Silla, J. M., de Oliveira, T., Boyle, S. A., Bezerra, B. M., et al (2017). Run, hide, or fight: Anti-predation strategies in endangered red-nosed cuxiú (Chiropotes albinasus, Pitheciidae) in southeastern Amazonia. Primates, 58, 353–360.
Barnett, A. A., Todd, L. M., & Tománek, P. A. (2018). Ecologia do uacari-de-costas-douradas (Cacajao ouakary) (Pitheciidae) na bacia Amazônica. In B. Urbani, M. Kowalewski, R. Grasseto Teixeira da Cunha, S. de la Torre, & L. Cortés-Ortiz (Eds.), La primatología en Latinoamérica 2: A primatologia na America Latina 2 (pp. 219–228). Caracas: Instituto Venezolano de Investigacoes Cientificas.
Benz, J. J., French, J. A., & Leger, D. W. (1990). Sex differences in vocal structure in a callitrichid primate, Leontopithecus rosalia. American Journal of Primatology, 21, 257–264.
Bezerra, B., Casar, C., Jerusalinsky, L., Barnett, A., Bastos, M., et al (2017). Pitheciid vocal communication: What can we say about what they are saying? Ethnobiology and Conservation, 6, 1–23.
Bezerra, B. M., Souto, A. S., & Jones, G. (2010). Vocal repertoire of golden-backed uakaris (Cacajao melanocephalus): Call structure and context. International Journal of Primatology, 31, 759–778.
Boersma, P. (2001). Praat, a system for doing phonetics by computer. Glot International, 5(9/10), 341–345.
Bouchet, H., Blois-Heulin, C., & Lemasson, A. (2013). Social complexity parallels vocal complexity: A comparison of three non-human primate species. Frontiers in Psychology, 4, 390.
Boyle, S. A. (2016). Pitheciids in fragmented habitats: Land cover change and its implications for conservation. American Journal of Primatology, 78, 534–549.
Bradbury, J. W., & Vehrencamp, S. L. (1998). Principles of animal communication (2nd ed.). Sunderland, MA: Sinauer Associates.
Bradshaw, J. W. S., & Cameron-Beaumont, C. L. (2000). The signaling repertoire of the domestic cat and its undomesticated relatives. In D. C. Turner & P. Bateson (Eds.), The domestic cat: The biology of its behaviour (pp. 67–94). Cambridge: Cambridge University Press.
Briefer, E. F. (2012). Vocal expression of emotions in mammals: Mechanisms of production and evidence. Journal of Zoology, 288, 1–20.
Buchanan, D. B. (1978). Communication and ecology of pithecine monkeys with special reference to Pithecia pithecia. PhD thesis, Wayne State University.
Buchanan, K., Burt de Perera, T., Carere, C., Carter, T., Hailey, A., et al (2012). Guidelines for the treatment of animals in behavioural research and teaching. Animal Behaviour, 83, 301–309.
Burke, C. J., Kisko, T. M., Euston, D. R., & Pellis, S. M. (2018). Do juvenile rats use specific ultrasonic calls to coordinate their social play? Animal Behaviour, 140, 81–92.
Campbell, C. J., Fuentes, A., MacKinnon, K. C., Bearder, S. K., & Stumpf, R. M. (2011). Primates in perspective. New York: Oxford University Press.
Carpenter, C. R. (1934). A field study of the behaviour and social relations of howling monkeys. Comparative Psychology Monographs, 10, 168.
Casamitjana, J. (2002). The vocal repertoire of the woolly monkey Lagothrix lagothricha. Bioacoustics, 13, 1–19.
Cäsar, C., Byrne, R. W., Young, R. J., & Zuberbühler, K. (2012). The alarm call system of wild black-fronted titi monkeys, Callicebus nigrifrons. Behavioral Ecology and Sociobiology, 66, 653–667.
Di Fiore, A., Fernandez-Duque, E., & Hurst, D. (2007). Adult male replacement in socially monogamous equatorial saki monkeys (Pithecia aequatorialis). Folia Primatologica, 78, 88–98.
Ey, E., Pfefferle, D., & Fischer, J. (2007). Do age-and sex-related variations reliably reflect body size in non-human primate vocalizations? A review. Primates, 48, 253–267.
Farmer, H. L. A. (2011). Howl vocalisations of captive black and gold howler monkeys (Alouatta caraya): Acoustics, function and applications for welfare. PhD thesis, University of Exeter.
Ferrari, S. F., Iwanaga, S., Ravetta, A. L., Freitas, F. C., Sousa, B. A., et al (2003). Dynamics of primate communities along the Santarem-Cuiba Highway in south-central Brazilian Amazonia. In L. K. Marsh & C. A. Chapman (Eds.), Primates in fragments: Ecology and conservation (pp. 123–144). New York: Kluwer Academic/Plenum.
Fischer, J., Hammerschmidt, K., Cheney, D. L., & Seyfarth, R. M. (2002). Acoustic features of male baboon loud calls: Influences of context, age, and individuality. The Journal of the Acoustical Society of America, 111, 1465–1474.
Fontaine, R. (1981). The uakaris, genus Cacajao. In A. Coimbra-Filho & R. A. Mittermeier (Eds.), Ecology and behavior of Neotropical primates (pp. 443–493). Rio de Janeiro: Academia Brasileira de Ciências.
Freeberg, T. M., Dunbar, R. I., & Ord, T. J. (2012). Social complexity as a proximate and ultimate factor in communicative complexity. Philosophical Transactions of the Royal Society of London B: Biological Sciences, 367, 1785–1801.
Gestich, C. C., Arroyo-Rodríguez, V., Ribeiro, M. C., da Cunha, R. G., & Setz, E. Z. (2019). Unravelling the scales of effect of landscape structure on primate species richness and density of titi monkeys (Callicebus nigrifrons). Ecological Research, 34, 150–159.
Green, S. (1975). Variation of vocal pattern with social situation in the Japanese monkey (Macaca fuscata): A field study. In L. A. Rosenblum (Ed.), Primate behaviour: Development and laboratory research, Vol. 4 (pp. 1–102). New York: Academic Press.
Gros-Louis, J. J., Perry, S. E., Fichtel, C., Wikberg, E., Gilkenson, H., et al (2008). Vocal repertoire of Cebus capucinus: Acoustic structure, context, and usage. International Journal of Primatology, 29, 641–670.
Hauser, M. D., Evans, C. S., & Marler, P. (1993). The role of articulation in the production of rhesus monkey, Macaca mulatta, vocalizations. Animal Behaviour, 45, 423–433.
Henline, W. T. (2007). Vocal repertoire of white-faced sakis (Pithecia pithecia). Master’s thesis, Eastern Kentucky University.
Hershkovitz, P. (1977). Living New World monkeys (Platyrrhini). Chicago: University of Chicago Press.
Hradec, M., Linhart, P., Bartoš, L., & Bolechová, P. (2017). The traits of the great calls in the juvenile and adolescent gibbon males Nomascus gabriellae. PLoS ONE, 12, e0173959.
Inoue, M. (1988). Age gradations in vocalization and body weight in Japanese monkeys (Macaca fuscata). Folia Primatologica, 51, 76–86.
INPE. (2016). Prodes: Assessment of deforestation in Brazilian Amazonia. São Paulo, Brazil: Instituto Nacional de Pesquisas Espaciais (INPE)
IUCN (2019). The IUCN Red List of Threatened Species. Version 2019-2. http://www.iucnredlist.org (accessed August 15, 2019).
Izawa, K., & Yoneda, M. (1981). Habitat utilization of nonhuman primates in a forest of Western Pando. Kyoto University, Vol. IV: Report of New World monkeys, 13–22.
Jacobsen, K. R., Mikkelsen, L. F., & Hau, J. (2010). The effect of environmental enrichment on the behaviour of captive tufted capuchin monkeys (Cebus apella). Laboratory Animal, 39, 269.
Kantha, S. S., Suzuki, J., Hirai, Y., & Hirai, H. (2009). Behavioral sleep in captive owl monkey (Aotus azarae) and squirrel monkey (Saimiri boliviensis). Acta Neurobiologiae Experimentalis, 69, 537–544.
Kieran, T. J. (2012). Identity and vocal repertoire of a Peruvian saki monkey (Pithecia aequatorialis). Master’s thesis, Winthrop University.
Kinzey, W. G., Rosenberger, A. L., Heisler, P. S., Prowse, D. L., & Trilling, J. S. (1977). A preliminary field investigation of the yellow-handed titi monkeys, Callicebus torquatus torquatus, in northern Peru. Primates, 18, 159–181.
Kleiman, D. G., Hoage, R. J., & Green, K. M. (1988). The lion tamarins, genus Leontopithecus. In R. A. Mittermeier, A. F. Coimbra-Filho, & G. A. B. Fonseca (Eds.), Ecology and behaviour of Neotropical primates (pp. 299–347). New York: World Wildlife Fund.
Lehman, S. M., Prince, W., & Mayor, M. (2001). Variations in group size in white-faced sakis (Pithecia pithecia): Evidence for monogamy or seasonal cogregations? Neotropical Primates, 9, 96–101.
Leyhausen, P. (1979). Cat behaviour: The predatory and social behaviour of domestic and wild cats. New York: Garland STPM Press.
Lingle, S., Wyman, M. T., Kotrba, R., Teichroeb, L. J., & Romanow, C. A. (2012). What makes a cry a cry? A review of infant distress vocalizations. Current Zoology, 58, 698–726.
Linhart, P., Ratcliffe, V. F., Reby, D., & Špinka, M. (2015). Expression of emotional arousal in two different piglet call types. PLoS ONE, 10, e0135414.
Marsh, L. K. (2014). A taxonomic revision of the saki monkeys, Pithecia (Desmarest, 1804). Neotropical Primates, 21, 1–165.
Martin, P., Bateson, P. P. G., & Bateson, P. (2007). Measuring behaviour: An introductory guide (3rd ed.). Cambridge: Cambridge University Press.
McComb, K., & Semple, S. (2005). Coevolution of vocal communication and sociality in primates. Biology Letters, 1, 381–385.
Meyer, D., Hodges, J. K., Rinaldi, D., Wijaya, A., Roos, C., & Hammerschmidt, K. (2012). Acoustic structure of male loud-calls support molecular phylogeny of Sumatran and Javanese leaf monkeys (genus Presbytis). BMC Evolutionary Biology, 12, 16.
Mittermeier, R. A., & van Roosmalen, M. G. (1981). Preliminary observations on habitat utilization and diet in eight Surinam monkeys. Folia Primatologica, 36, 1–39.
Mittermeier, R. A., Veiga, L. M., & Marsh, L. K. (2015). Pithecia chrysocephala. The IUCN Red List of Threatened Species 2015. https://doi.org/10.2305/IUCN.UK.2015-1.RLTS.T43943A70606989.en (accessed August 24, 2018).
Morton, E. S. (1977). On the occurrence and significance of motivation-structural rules in some bird and mammal sounds. American Naturalist, 111, 855–869.
Moss, C. F., Redish, D., Gounden, C., & Kunz, T. H. (1997). Ontogeny of vocal signals in the little brown bat, Myotis lucifugus. Animal Behaviour, 54, 131–141.
Moynihan, M. (1964). Some behaviour patterns of playtyrrhine monkeys I. The night monkey (Aotus trivirgatus). Smithsonian Miscellaneous Collections, 146, 1–84.
Moynihan, M. (1966). Communication in the titi monkey, Callicebus. Journal of Zoology, 150, 77–127.
Norconk, M. A. (2006). Long-term study of group dynamics and female reproduction in Venezuelan Pithecia pithecia. International Journal of Primatology, 27, 653–674.
Norconk, M. A. (2011). Sakis, uakaris, and titi monkeys: Behavioural diversity in a radiation of primate seed predators. In C. J. Campbell, A. Fuentes, K. C. MacKinnon, S. K. Bearder, & R. M. Stumpf (Eds.), Primates in perspective (pp. 122–139). New York: Oxford University Press.
Norconk, M. A., & Setz, E. Z. (2013). Ecology and behavior of saki monkeys (genus Pithecia). In L. M. Veiga, A. A. Barnett, S. F. Ferrari, & M. A. Norconk (Eds.), Evolutionary biology and conservation of titis, sakis and uacaris (pp. 262–271). Cambridge: Cambridge University Press.
Norcross, J. L., Newman, J. D., & Cofrancesco, L. M. (1999). Context and sex differences exist in the acoustic structure of phee calls by newly–paired common marmosets (Callithrix jacchus). American Journal of Primatology, 49, 165–181.
Pinto, L. P., Barnett, A. A., Bezerra, B. M., Boubli, J. P., Bowler, M., et al (2013). Why we know so little: The challenges of fieldwork on the Pitheciids. In L. M. Veiga, A. A. Barnett, S. F. Ferrari, & M. A. Norconk (Eds.), Evolutionary biology and conservation of titis, sakis and uacaris (pp. 145–150). Cambridge: Cambridge University Press.
Pollermann, B. Z., & Archinard, M. (2002). Acoustic patterns of emotions. In E. Keller, G. Bailly, A. Monaghan, J. Terken, & M. Huckvale (Eds.), Improvements in speech synthesis (pp. 237–245). Hoboken, NJ: Wiley.
Range, F., & Fischer, J. (2004). Vocal repertoire of sooty mangabeys (Cercocebus torquatus atys) in the Taï National Park. Ethology, 110, 301–321.
Redican, W. K. (1975). Facial expressions in nonhuman primates. In L. A. Rosenblum (Ed.), Primate behaviour: Developments in field and laboratory research, Vol. 4 (pp. 103–194). New York: Academic Press.
Robinson, J. G. (1979). Vocal regulation of use of space by groups of titi monkeys Callicebus moloch. Behavioral Ecology and Sociobiology, 5, 1–15.
Robinson, J. G. (1981). Vocal regulation of inter-and intragroup spacing during boundary encounters in the titi monkey, Callicebus moloch. Primates, 22(2), 161–172.
Salmi, R., Hammerschmidt, K., & Doran-Sheehy, D. M. (2013). Western gorilla vocal repertoire and contextual use of vocalizations. Ethology, 119, 831–847.
Schott, D. (1975). Quantitative analysis of the vocal repertoire of squirrel monkeys (Saimiri sciureus). Zeitschrift für Tierpsychologie
Schwitzer, C., Glatt, L., Nekaris, K. A. I., & Ganzhorn, J. U. (2011). Responses of animals to habitat alteration: An overview focussing on primates. Endangered Species Research, 14, 31–38.
Setz, E. Z., & Gaspar, D. D. A. (1997). Scent-marking behaviour in free-ranging golden-faced saki monkeys, Pithecia pithecia chrysocephala: Sex differences and context. Journal of Zoology, 241, 603–611.
Smith, A. S., Birnie, A. K., Lane, K. R., & French, J. A. (2009). Production and perception of sex differences in vocalizations of Wied's black‐tufted‐ear marmosets (Callithrix kuhlii). American Journal of Primatology: Official Journal of the American Society of Primatologists, 71(4), 324-332.
Snowdon, C. T., & Elowson, A. M. (2001). ‘Babbling’ in pygmy marmosets: Development after infancy. Behaviour, 138, 1235–1248.
Sokoloff, G., & Blumberg, M. S. (2004). Vocalization. In B. Kolb & I. Q. Whishaw (Eds.), The behaviour of the laboratory rat: A handbook with tests (pp. 371–380). Oxford: Oxford University Press.
Strier, K. B. (2007). Communication and cognition. In K. B. Strier (Ed.), Primate behavioural ecology (pp. 311–346). Boston: Pearson Education.
Take, M. (2017) Comparison of the diets among three sympatric primate species in an urban fragmented forest in Manaus, Brazillian Amazon. Master’s thesis, Kyoto University.
Titze, I. R. (1994). Principles of voice production. Englewood Cliffs, NJ: Prentice-Hall.
Tomaszycki, M. L., Davis, J. E., Gouzoules, H., & Wallen, K. (2001). Sex differences in infant rhesus macaque separation–rejection vocalizations and effects of prenatal androgens. Hormones and Behaviour, 39, 267–276.
Vie, J. C., Richard-Hansen, C., & Fournier-Chambrillon, C. (2001). Abundance, use of space, and activity patterns of whitefaced sakis (Pithecia pithecia) in French Guiana. American Journal of Primatology, 55, 203–221.
Waser, P. M. (1982). The evolution of male loud calls among mangabeys and baboons. In T. Snowdon, C. H. Brown, & M. R. Peterson (Eds.), Primate communication (pp. 117–144). Cambridge: Cambridge University Press.
Watson, C. F., Buchanan-Smith, H. M., & Caldwell, C. A. (2014). Call playback artificially generates a temporary cultural style of high affiliation in marmosets. Animal Behaviour, 93, 163–171.
Zimmermann, E. (1995). Acoustic communication in nocturnal prosimians. In L. Alterman, G. A. Doyle, & M. K. Izard (Eds.), Creatures of the dark (pp. 311–330). Boston: Springer.
Acknowledgments
We thank Tianara Sobroza and Makiko Take for their assistance in Brazil. We also thank Instituto National de Pesquisas da Amazonia for providing the opportunity to carry out this project, and Santander for providing a Student Project Grant. We are grateful to three anonymous reviewers for their comments on the original manuscript.
Author information
Authors and Affiliations
Contributions
JM and MSS conceived and designed the study; JM collected and analysed the data; JM, MSS, and AB wrote the article.
Corresponding author
Additional information
Handling Editor: Joanna M. Setchell
Rights and permissions
About this article

Cite this article
Muir, J., Barnett, A. & Svensson, M.S. The Vocal Repertoire of Golden-Faced Sakis, Pithecia chrysocephala, and the Relationship Between Context and Call Structure. Int J Primatol 40, 721–743 (2019). https://doi.org/10.1007/s10764-019-00125-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10764-019-00125-7