
Abstract
Coastal freshwater wetlands are amongst the world’s most modified but poorly researched ecosystems and some of the most vulnerable to climate change. Here, we examine vegetation resilience in coastal wetlands of subtropical Australia to altered salinity and flooding regimes likely to occur with climate change. We conducted field surveys and glasshouse experiments to examine plant diversity and regeneration responses of understorey and canopy species across four habitats. Vegetation composition, but not richness, varied between seaward and inland habitats while soil seed bank diversity was greatest in more inland sites. Experimental salinity and flooding treatments strongly influenced emergence from seed banks with most species germinating under fresh, waterlogged conditions and very few in saline treatments. Composition of emerging seedling assemblages was similar across habitats and treatments but differed considerably from the extant vegetation, indicating a relatively minor role of soil seed banks in sustaining current vegetation structure in this wetland. An exception to this was Sporobolus virginicus (marine couch) which was common in both the vegetation and seed banks suggesting a high capacity for this species to re-establish following disturbances. Seedlings of dominant canopy species also reacted strongly to increased salinity treatments with decreased survivorship recorded. Overall, our findings suggest a high probability of constrained vegetation regeneration in this wetland in response to key projected climate change disturbances with implications for vegetation diversity at a landscape scale including declines in the extent and diversity of more landward vegetation communities and expansion of salt-tolerant marshes dominated by Sporobolus virginicus.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Coastal freshwater wetlands (CFWs) are highly valued for their diverse biota and ecological services (Millennium Ecosystem Assessment 2005). Typically positioned between estuarine and freshwater systems, CFWs are neither fully saline nor exclusively freshwater. Rather, the main water sources are fresh (i.e. rainfall, terrestrial runoff, streamflow, groundwater) with saline intrusion events also occurring during the largest tides and storm surge events (Office of Environment and Heritage (OEH) 2011). The unique hydrology of these systems, highlighted by the variability in moisture and salinity, creates distinct vegetation communities (David 1996; Olmstead and Armentano 1997; Ross et al. 2003; Todd et al. 2010). CFWs provide many ecosystem services that are valuable to the organisms which inhabit these areas as well as the human populations living around them including; but not limited to, habitat provision and diversity, nutrient cycling, flood mitigation, storm buffering and carbon sequestration (Greenway 2001; Millennium Ecosystem Assessment 2005; Saintilan et al. 2018).
CFWs are commonly found in areas subject to significant urban development (i.e. along rivers, coastal regions) and in many places, large areas of these wetlands have been cleared, drained and filled to provide space for cultivation, grazing and urban developments (Tait 1994; Johnson et al. 1999). Despite their ecological importance and vulnerability to human pressures, CFWs are amongst the most poorly researched wetlands in the world (Finlayson et al. 1999; Leigh et al. 2015; Saintilan et al. 2018). Furthermore, CFWs are likely to be highly vulnerable to climate change impacts due to their topographic location and proximity to intense human activity which will constrain their capacity to be resilient and adapt to future changes (Rogers et al. 2014a; Peirson et al. 2015). In this paper, resilience is defined as “the rate at which a system regains structure and function following a stress or perturbation” (Park and Allaby 2013). Most research concerning the ecology and resilience of CFWs has occurred in the USA (i.e. Baldwin et al. 2001; Hopfensperger and Engelhardt 2008; Craft et al. 2009) with a few studies in Australia (Johnson et al. 1999; Zoete 2001; Boon 2012), most notably in Kakadu National Park in northern Australia (Knighton et al. 1991; Finlayson 2005; Bowman et al. 2010; Finlayson et al. 2013). Given this limited knowledge, there is an urgent need to better understand the resilience and vulnerability of these ecosystems to climate change to inform the development of effective management strategies.
Extremes in climate-related pressures, such as fires, cyclones, floods and storm surges, represent major disturbances to CFWs with the potential to alter nutrient flows and remove or change vegetation communities (Cahoon 2006; Hauser et al. 2015). While unmodified and intact wetlands may be able to cope with such disturbances through inherent mechanisms of resilience and adaptive capacity, human alteration of these systems typically reduces these capacities, placing them at risk of extinction (Middleton 1999). Projected climatic changes, including altered rainfall patterns, warmer temperatures, increased occurrence of extreme weather events, rising sea levels and salinisation of wetlands and groundwater, are therefore likely to trigger major shifts in the structure and function of CFWs (OEH 2011; IPCC 2014; Craft et al. 2009). Anticipated impacts, however, are difficult to manage as the effects of many climate threats and their interactions are poorly understood for these systems, although modelling generally suggests potential for substantial changes (Shoo et al. 2014; Rogers et al. 2014b; Leigh et al. 2015). Modelling of the future distributions of coastal wetlands based on climatic envelopes in south-east Queensland, for instance, suggests that mangrove forests are likely to become the dominant vegetation type in this coastal landscape as they migrate landward in response to rising sea levels (Saintilan and Williams 1999; Cahoon et al. 2006; Nicholls et al. 2007; Saintilan and Rogers 2013; Shoo et al. 2014), with sedgelands and samphire vegetation types potentially being extirpated as a result (Shoo et al. 2014). While CFWs may similarly shift in a landward direction, there tends to be significant spatial constraints on these ecosystems because of urban developments and unsuitable hydrology. Consequently, there is a strong potential for CFWs to become ‘squeezed’ or face local extinction (Boon 2012; Rogers et al. 2014a). Ecological processes which might drive patterns of landward migration and/or extirpation of wetland ecosystems and subsequent landscape transformation, however, are largely unknown.
Regeneration of vegetation communities is critical to the recovery of an ecosystem after disturbance, the lack of which could lead to a transition to an alternative stable state (May 1977; Westman 1978). The regeneration niche concept is therefore important to help describe and predict the response of species post disturbance, where all five stages of regeneration ((1) seed production, (2) seed dispersal, (3) germination, (4) establishment and (5) further development of immature plant) must be realised for regeneration, which contributes to resilience, to be successful (Grubb 1977). Because of the critical importance of vegetation to wetland structure and function, the resilience of wetland ecosystems to climate change depends strongly on vegetation responses to projected threats such as altered hydrology and salinity (Capon et al. 2013). Many freshwater wetland plants are well adapted to variable hydrologic regimes, but tend to be strongly and negatively impacted by salinity (Flynn et al. 1995; Neubauer 2013; Sutter et al. 2014; Herbert et al. 2015). Increased saline groundwater intrusion and increased periods of saltwater inundation have already led to large-scale changes in vegetation community structure, including tree mortality, in many American tidal freshwater marshes (Leck and Simpson 1995; Baldwin et al. 2001; Anderson et al. 2013). Changes in hydrology also drive shifts in coastal wetland vegetation composition, with both floods and droughts reducing species richness and abundance (Baldwin et al. 2001). Soil seed banks are often important for the resilience of wetland understorey vegetation, especially in hydrologically variable wetlands (Hopfensperger and Engelhardt 2008), providing a mechanism of regeneration following a range of disturbances including floods (Casanova and Brock 2000; Capon and Brock 2006), fire (Kimura and Tsuyuzaki 2011), cyclone (Middleton 2009) and drought (Capon 2003, 2007; Brock 2011). The regenerative capacity of soil seed banks, however, may also be affected by projected climatic changes, both with respect to the composition and structure of the soil seed banks, as well as the establishment responses of the soil seed bank species. In most freshwater wetlands, for instance, increasing salinity typically reduces and homogenises the diversity of plant assemblages emerging from the soil seed banks, resulting in declines in wetland vegetation diversity (Brock et al. 2005). Additionally, the structure and composition of wetland soil seed banks tend to reflect hydrological regimes and disturbances (Reid and Capon 2011; Capon and Reid 2016).
In wetlands where the soil seed bank does not contribute significantly to the resilience and regeneration capacity of the vegetation community, resilience could be facilitated by perennial species which can regenerate asexually (Combroux et al. 2002). The presence of a rhizomatous stem/root system and the ability to produce roots from cuttings (clonal fragments) are adaptations which allow dominant marsh species, such as the common reed Phragmites australis, many salt marsh grasses (e.g. Sporobolus virginicus and Spartina alterniflora) and sedge (Scirpus americanus) species, to withstand disturbance and allow them to regenerate and spread rapidly throughout wetland areas (Alvarez et al. 2005; Xiao et al. 2010; Marsh et al. 2016).
The resilience of wetland vegetation in CFWs to the combined effects of altered hydrology and salinity remains poorly understood in most places. Here, we sought to address major knowledge gaps concerning coastal freshwater wetland vegetation in eastern Australia and its potential responses to the changes in hydrology and water quality (i.e. salinity), which are likely to be driven by climate change. We investigated patterns in vegetation diversity and composition along a seaward-landward gradient exhibiting stark zonation of dominant canopy species. We examined the resilience of understorey vegetation along this gradient with respect to inundation of fresh- and saltwater by investigating patterns of seedling emergence from the soil seed bank. We also explored the effects of a similar range of flooding and salinity conditions on the seedling establishment of the dominant canopy species characterising the major habitat types along this gradient. Overall, we sought to understand the resilience of vegetation in this CFW in order to make predictions concerning the capacity of vegetation to regenerate following major climate-driven disturbances in these coastal freshwater wetlands and infer consequences for wetland biodiversity in these landscapes under projected climate change.
Methods
Study area
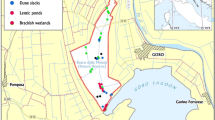
The study was conducted in the Boondall Wetlands Reserve in south-east Queensland (SEQ; 27° 20′ 3″ S:153° 04′ 5″ E). The climate of this region is subtropical with summer mean temperatures reaching 29 °C and winter minimums of 9.8 °C. Rainfall is variable and falls predominantly in summer months with a mean annual rainfall of 1140 mm (Bureau of Meteorology (BOM) 2017). The study area is low lying, rarely exceeding 1 m above sea level, and includes numerous creeks which drain into Moreton Bay (Brisbane City Council 2017). The tidal range of the nearest monitoring point (Brisbane Bar 27° 22′ S:153° 10′ E), based on the last 10 years, is minimum 0.201 m ± 0.079 m to maximum 2.668 m ± 0.1228 m (based on mean lowest astronomical tide; BOM 2017). The Boondall Wetland Reserve is part of the Moreton Bay Ramsar site but is constrained by urban development on all but its ocean side (Fig. 1). CFWs in SEQ are palustrine wetlands, i.e. vegetated, non-river or channel systems where emergent vegetation represents greater than 30% of ground cover (Department of Environment and Heritage Protection (DEHP) 2013). The CFWs within Boondall wetlands fall into the sub-class of coastal and subcoastal floodplain wetlands (tree swamp) and saline swamp (DEHP 2013).

Map of south-east Queensland, inset. Boondall Wetlands study site and plot locations within each habitat type: ▲ = Sporobolus, + = Casuarina, ♦ = Melaleuca, • = Eucalyptus (QLD Department of Natural Resources, Mines and Energy 2018)
Wetland zonation
Clear zonation is evident in the vegetation which comprises several relatively distinct communities (hereafter referred to as habitats), each dominated by a single canopy species, that occur along a gradient of increasing distance from the coast; marine couch (Sporobolus virginicus) marsh, swamp she-oak (Casuarina glauca) woodland, swamp paperbark (Melaleuca quinquenervia) woodland and eucalypt (Eucalyptus tereticornis) woodland (Supp 1). C. glauca and M. quinquenervia trees form a distinct monoculture canopy in their respective habitats; however, Eucalyptus tereticornis is commonly found alongside Corymbia tessellaris, Alphitonia excelsa and Melaleuca spp. Understorey vegetation communities are distinctly different between habitats with increased diversity in Melaleuca and Eucalyptus habitats compared to Sporobolus and Casuarina habitats. Perennial species are common throughout the understorey of all habitats but annual species are only found in the Casuarina, Melaleuca and Eucalypt habitats. The soil seed bank is most abundant in Eucalyptus habitats but was more diverse in Melaleuca habitats. Sporobolus habitats have the most depauperate soil seed bank but contain the largest number of perennial species. Sporobolus virginicus is common in the soil seed bank for all habitats except Eucalyptus, where the annual weed Ageratum houstonianum is most common.
Data collection
Extant vegetation survey
Extant vegetation was surveyed in April 2016, in five randomly positioned sites (5 × 5 m) in each habitat type. April was an optimal time for vegetation survey and soil seed bank collection as majority of common Queensland wetland plants flower during the spring and summer months (September–February; Gunn 2001) and allowed for any summer flow events to subside. Sites within habitats were positioned at least 100 m apart from each other. Within each site, the presence of all understorey (< 1 m) and overstorey (> 1 m) plant species was recorded. Five random subsamples of soil (15 × 15 × 10 cm) were collected from each site and aggregated into a single bag for use in experiments. Experimentation was conducted in the glasshouse facility at Griffith University, Brisbane, south-east Queensland (SEQ).
Soil seed bank experimental design
Aggregated soil samples were divided into five plastic containers (17 × 12 × 3 cm), with drainage holes in the bottom, to a depth of 2.5 cm. Each container was placed inside a larger container (27.5 × 21.5 × 13 cm) to apply the water and salinity treatments; (1) flooding with saltwater, (2) waterlogging with saltwater, (3) flooding with freshwater, (4) waterlogging with freshwater and (5) ambient rainfall. Flooded samples were submerged to a level of 6.5 cm above the soil surface while waterlogging was achieved by maintaining the water level at the soil surface. Salinity treatments were created by dissolving superfine salt into freshwater to create a 35 dS/m mixture (equivalent to seawater; Ocean Nature Synthetic Sea Salt). Ambient rainfall treatments involved waterlogging samples with freshwater once a week but without placing samples in larger containers to allow water to drain away.
Experimental containers were randomly arranged on tables within the glasshouse. Eight samples of potting mix (Brunnings Potting Mix) were used as controls to identify the presence of ‘fly in’ seeds, i.e. seedlings which were not present at the sampling site but which ended up in the glasshouse by any means. The experiment ran for 16 weeks from May to September 2016 during which time water levels were adjusted as necessary every 3–4 days. Glasshouse temperatures ranged between minimums of 8–15 °C and maximums of 27–43 °C during the experimental period. Counts of emerging seedlings were conducted every 3–4 days, with species identifications made where possible larger seedlings were removed and transplanted into deeper seedling tubes (5 × 5 × 12 cm) to promote flowering for identification. This technique is known as the emergence method, which has been used in previous studies as an accurate measure of viable seeds in wetland soils (Leck and Simpson 1995; Baldwin et al. 1996; Baldwin et al. 2001; Peterson and Baldwin 2004).
Seedling experimental design
This study examined responses to flooding and salinity treatments of seedlings of Sporobolus virginicus, Casuarina glauca, Melaleuca quinquenervia and Eucalyptus tereticornis, representing the dominant canopy species of the four major habitat types. Fifty-two seedlings of each species were sourced from Wallum Nurseries (Gumdale, QLD). Individuals (height 100–200 mm) were repotted into 300-mm (90-mm diameter) PVC tubes, covered at one end with fine mesh and a 20-mm base layer of vermiculite (Brunnings Vermiculite) to allow for water uptake. Soil from the corresponding habitat type was used to repot each seedling. Seedling height was recorded for each individual prior to repotting and four seedlings per species were reserved to determine initial biomass (see below).
Repotted seedlings were given a 5-day establishment period of regular watering to allow for initial readjustments. Twelve seedlings of each species were randomly allocated to each treatment and divided between 16 large containers (740 × 535 × 440 cm: four for each treatment) to conduct these treatments: (1) flooding with saltwater, (2) flooding with freshwater, (3) ambient rainfall and (4) flooding with saltwater for 7 days after which water was replaced with freshwater, to simulate a storm surge followed by large freshwater flows. Flooding was achieved by submerging seedlings 10 cm above the soil surface with salt added as per the soil seed bank experiment. Water and salinity treatments were applied within the outer containers and levels were adjusted every 3–4 days. The rain treatment was included as a control with seedlings receiving neither flooding nor salinity stress. Seedling height (i.e. length from soil surface to tip of stem including the longest leaf) was recorded weekly, as were observations of adventitious root growth and salt secretion on stems and leaves were recorded. The experimental period was 31st August to 14th October 2016, during which the glasshouse temperature ranged between minimums of 8–15 °C and maximums of 27–43 °C. At the end of the experimental period, a random sample of four seedlings per species, per treatment, was harvested to determine biomass. Plants were removed from the PVC tube and the roots washed to remove excess soil, then placed in a 60 °C oven for 48 h. Roots were separated from stems and leaves and weighed to determine above- and below-ground biomass (Sharpe and Baldwin 2012; Isla et al. 2014). Four seedlings of each species, which were set aside at the beginning of the experiment, were used as an initial biomass measure to compare against.
Data analysis
Extant vegetation composition
Differences of species richness between habitat types extant understorey vegetation assemblages were explored by analysis of variance (ANOVA) and Tukey’s post hoc tests. Changes in vegetation composition across habitats was investigated through non-metric multidimensional scaling (nMDS) and analysis of similarity (ANOSIM; Euclidean distance) using a species presence/absence data transformation. Pairwise beta-diversity (β-diversity) was used to further investigate compositional differences. Differences in the abundance, species richness and composition of seedlings emerging from the soil seed banks of each habitat type were similarly examined for each site using an ANOVA model and Tukey’s post hoc tests.
Soil seed bank and seedling treatment effects
Effects of experimental treatments on the richness and abundance of emergent seedlings were investigated with ANOVA and Tukey’s post hoc test. Differences in the composition of emergent species in relation to experimental treatments were investigated using nMDS and an ANOSIM (Euclidean distance) based on a species presence/absence data transformation. Responses of canopy seedlings to experimental treatments were investigated, first by calculating changes in height over time, then by one-way ANOVA and Tukey’s post hoc tests to explore effects of experimental treatments within each species. Biomass results were standardised across species by subtracting the average initial biomass to determine the change in biomass due to treatments. Factorial ANOVA and Tukey’s post hoc test were used to identify treatment effects on total biomass, above- and below-ground biomass for each species. Prior to data analyses, data was checked against assumptions of normality and presence/absence data transformations were performed where required.
All univariate analyses were conducted in MS Excel and RStudio (R core team 2013). Multivariate statistics (nMDS and ANOSIM) were conducted in the statistical program R 3.3.3 using the package vegan for nMDS and ANOSIM functions and, ggplot2 for univariate graphics (R core team 2013).
Results
Extant vegetation composition
Overstorey species typically followed the given zonation description. A total of 25 species, representing 16 families, were identified in the understorey vegetation, mainly comprising forbs (12 spp.), grasses and sedges (4 spp.; Table 1). Poaceae (3 spp.) and Chenopodiaceae (3 spp.) were the most frequently represented families. Vines and grasses were common in the Melaleuca and Eucalypt habitats with Parsonsia straminea, Dianella revoluta and Imperata cylindrica present in both. Sporobolus virginicus was the most frequently recorded and widespread species, present in all four habitats. Passiflora suberosa, Parsonsia straminea and Commelina diffusa were also common, occurring in all habitats except the Sporobolus marsh.
Species richness of understorey vegetation did not significantly differ between habitat types (F = 2.133; p = 0.136) although fewer species typically occurred in the Sporobolus marsh habitat (Supp. Fig. 1).
Understorey vegetation composition varied significantly between habitat types (ANOSIM R = 0.633; p < 0.0001; Supp. Fig. 2). The species driving this differentiation included Sporobolus virginicus, which was unsurprisingly dominant in the Sporobolus marsh habitat; the forbs Commelina diffusa, Passiflora suberosa and Einadia hastata were associated with the Casuarina woodland habitat; and the perennial herb Dianella revoluta and grass Imperata cylindrica were associated with the Melaleuca woodland and Eucalyptus woodland habitat. Distinct differences in species composition were also detected between habitats, with the Sporobolus marsh being significantly different to the Melaleuca and Eucalyptus habitats (Table 1).
Soil seed bank composition
Over 1111 individual seedlings germinated during the soil seed bank experiment, representing at least 20 species, 18 of which were identified to at least genus. Across all habitat types, the most abundant species were Ageratum houstonianum (232), Sporobolus virginicus (620) and Eleocharis spp. (125). Six of the 18 identified species were annuals, all of which only emerged from Casuarina, Melaleuca and Eucalyptus samples. The Cyperaceae family was the most dominant of the 11 families with four species recorded. Seedlings belonging to the Poaceae, Asteraceae and Apiaceae families were also common, each being represented by two species. At a site level, both the abundance (F = 2.716, p = 0.049, df = 3) and species richness (F = 5.54, p = 0.0015, df = 3) of emerging seedlings were significantly higher from the soil seed banks of the Eucalyptus habitat than those of all other habitat types (Supp. Fig. 3a & b). The composition of emerging seedling assemblages, however, did not vary significantly in relation to habitat (ANOSIM R = 0.064; p < 0.0001).
Comparison of extant and soil seed bank assemblages
Only seven species were common to both the extant understorey vegetation and the soil seed bank (Ageratum houstonianum, Alternanthera denticulata, Commelina diffusa, Cyperus polystachyos, Einadia hastate, Sesuvium portulacastrum, Sporobolus virginicus), all of which are perennial except for Ageratum houstonianum and Alternanthera denticulata. With the exception of S. virginicus a grass and C. polystachyos a sedge, all of these common species were forbs. Ten species, mostly forbs and sedges, were recorded exclusively in the soil seed bank while 16 species were only recorded from the extant understorey vegetation including several vines and woody shrubs (e.g. Parsonsia straminea, Passiflora suberosa, Eustrephus latifolius) and several grass species (i.e. Imperata cylindrical and Chloris spp.). The number of species common between the extant vegetation and soil seed bank did not vary significantly in relation to habitat type (F = 2.809; p = 0.0728), although, β-diversity was the highest when comparing the extant and emergent communities of the Sporobolus marsh (Table 2).
Effects of salinity and flooding on emergence from soil seed banks
The abundance (F = 11.24; p < 0.0001, df = 4) and richness (F = 26.62; p < 0.0001, df = 4) of seedlings emerging from the soil seed bank varied significantly in response to the flooding and salinity treatments. Overall, more seedlings representing more species emerged under fresh-waterlogged treatments (Fig. 2). Seedling abundances and richness did not differ significantly between rain and fresh-flooded treatments but both were significantly higher than those from salt-flooded and salt-waterlogged treatments (Fig. 2). Seedling assemblages emerging from Eucalyptus and Melaleuca habitats under fresh-waterlogged treatment had the highest species richness, while no species emerged from samples from these habitats under salt-waterlogged or salt-flooded treatments (Fig. 2). The composition of emerging seedling assemblages did not vary significantly in relation to experimental treatments or habitat type (ANOSIM R = 0.254; p < 0.0001).

Boxplot of differences in species richness of emergent assemblages of major wetland habitat types between water and salinity treatments
Effects of flooding and salinity on growth of seedlings
Seedling survival was variable between species and experimental treatments. Treatment effects were greatest for Eucalyptus tereticornis seedlings of which < 10% survival occurred in both saltwater treatments while 100% survival was recorded in freshwater treatments. In contrast, seedling survival was > 60% for seedlings of Sporobolus virginicus regardless of treatment (Fig. 3). Melaleuca quinquenervia seedlings consistently showed lower rates of survival than seedlings of Casuarina glauca and both had lower survival rates in saltwater treatments compared to freshwater (Fig. 3). The treatment effects shown in Fig. 3 were not statistically significant.

Boxplot of the proportion of mortality of dominant wetland canopy species in water and salinity treatments
Amongst surviving seedlings, growth during the experiment varied in relation to species (F = 10.95; p < 0.0001; Fig. 4) and treatment. Higher average growth was evident in all species under rain and fresh-flooded conditions (F = 19.06; p < 0.0001, df = 9; Fig. 4) with significant interactions between species and treatment also detected (F = 19.06; p < 0.0001; Fig. 4), indicating differences in responses between species. Eucalyptus tereticornis seedlings exhibited significantly more growth under rain and fresh-flooded treatments than all other species or treatments (Fig. 4). Salinity significantly decreased survivorship (measured as a decrease in height) over the course of the experiment in all species except C. glauca (Fig. 4). Height of S. virginicus was significantly greater in the storm-surge treatments compared to salt-flooded treatments and was reduced in the fresh-flooded and rain treatment (Fig. 4). The height of M. quinquenervia and C. glauca seedlings did not significantly differ between treatments although a positive trend was recorded in fresh-flooded individuals of M. quinquenervia and in rain treatments for C. glauca. (Fig. 5). Sporobolus virginicus increased in height over time in response to all treatments except salt-flooded. The storm-surge treatments resulted in an initial decrease in survivorship of S. virginicus followed by an increase in height after saltwater was replaced with freshwater (Fig. 5). Only E. tereticornis exhibited a significant difference in above- and below-ground biomass across treatments where rain and fresh-flooded treatments were significantly greater (Fig. 6).

Boxplot of the average change in height for major wetland canopy species between water and salinity treatment

Change in mean height (± standard deviation) of seedlings over experimental period for aE. tereticornis, bS. virginicus and cC. glauca

Boxplots of a average above-ground biomass and b below-ground biomass of dominant wetland species of Boondall wetlands in response to different water and salinity treatments
Discussion
Climate change is predicted to significantly impact the eastern coast of Australia. Increased intensity of extreme rainfall events, a rise in sea level of 0.3–0.65 m by 2090 and an increase in height of extreme sea level events (i.e. king tides and storm surges) are all predicted with high to very high confidence (Reisinger et al. 2014; Dowdy et al. 2015). These predictions have been simulated in this study with freshwater inundation to represent the increased flooding from extreme rainfall events, and saltwater manipulations to represent saline groundwater inundation, increased height of storm surge and extreme sea level events and long-term increases in sea level. Due to the low-lying nature of the study area (generally < 1 m above sea level), it is likely that large proportions of this wetlands will be influenced by increased extreme sea level events and will become inundated with sea level rise.
Vegetation in many coastal wetlands exhibits a distinctive zonation in relation to increasing distance from the shoreline with variations in salinity and hydrology (He et al. 2011; Swarth et al. 2013; Janousek and Folger 2014). In coastal freshwater wetlands of south-east Queensland, this is reflected by clear changes in the distribution of dominant canopy species. Our results indicate that understorey vegetation also differs significantly across these habitats with a general increase in diversity in a landward direction along this gradient, possibly reflecting a decline in saline influences and a higher availability of freshwater in less tidally influenced, inland positions. Hydrology and salinity are likely to be major drivers of these shifts in vegetation structure and composition (Baldwin et al. 2001; Swarth et al. 2013), with salt tolerance playing a particularly key role in shaping species distributions within coastal wetlands (Odum 1988; Marcum and Murdoch 1992; Janousek and Folger 2014). In the wetlands studied here, low species diversity was observed in the Sporobolus marshes, possibly reflecting the greater saline influences here, compared to other habitats. Greater freshwater availability and/or the absence of saline water may support the higher plant diversity found in the Melaleuca and Eucalypt habitats. The decreased dominance of S. virginicus in the understorey of the Melaleuca and Eucalypt habitats might also permit the establishment of a more diverse community as S. virginicus is known to form thick, mat-like ground covers with a dense layer of roots, possibly inhibiting the germination and growth of other species (Marcum and Murdoch 1992; Boon 2012).
Role of the soil seed bank
Regeneration from the soil seed bank (germination) is an important stage of the regeneration niche (Grubb 1977). The soil seed banks of the CFWs in the Boondall wetlands are dominated by annual herbaceous species and appear to be relatively species depauperate, compared to many inland Australian wetlands (e.g. Capon 2003; James et al. 2007), although were similar in species diversity to tidal freshwater wetlands in the USA, (e.g. Baldwin et al. 2001; Peterson and Baldwin 2004). Soil seed bank composition does not differ substantially between the major habitat types examined here despite significant variation in the extant vegetation suggesting that there is a common soil seed bank species pool throughout all habitats sampled across the wetland (Baldwin et al. 1996; Cronk and Fennessy 2001). Given this similarity throughout the wetland, it is possible that hydrology and salinity do not play a role in structuring the soil seed bank composition for this CFW; however, there may be some role in seed viability and germination success, as has been found in tidal marshes in North America (Baldwin et al. 1996; Peterson and Baldwin 2004). Such homogeneity in wetland soil seed banks is likely due to widespread dispersal of propagules via wind and water (Baldwin et al. 1996; Capon 2003, 2007; Wall and Stevens 2015). The lack of substantial variation in the soil seed bank also suggests that compositional differences in the extant vegetation are due to effects of drivers such as hydrology and salinity on plant establishment and growth, rather than spatial differentiation in the availability of propagules (e.g. Webb et al. 2006).
Soil seed bank assemblages in tidal freshwater marshes frequently have different composition to extant vegetation communities (Wilson et al. 1993; Hopfensperger et al. 2009; Liu et al. 2006) and similar results have been observed in some inland wetlands (Reid and Capon 2011). In some cases, tidal freshwater marsh seed banks strongly resemble the species composition of extant vegetation in adjacent open areas, suggesting that the seed bank provides an indication of what would occur in the area if the canopy was removed (Cronk and Fennessy 2001). Rather than providing sources of regeneration for dominant and productive understorey species, the soil seed banks may instead contribute to the resilience of understorey plant diversity when certain conditions occur and the standing vegetation is removed, e.g. following recession of floodwaters or after fire (e.g. coastal dunes Pierce and Cowling 1991; alpine fields Chambers 1993). The homogeneity and minimal similarity of the soil seed bank in this study could also be attributed to temporal trends in seed availability and viability periods (i.e. the length of time a seed is viable for in the soil seed bank). No seedlings of C. glauca, M. quinquenervia or E. tereticornis were observed in the field survey or emerging from the soil seed bank. These species typically store seeds in an aerial seed bank and flower from September to October, March to July and June to November respectively; however, all species have seed capsules that persist on the tree for at least 1 year (Gunn 2001). Suggesting that conditions for germination of these seeds may not have occurred during experimentation or that these seeds do not persist in the soil seed bank.
Changes in flow patterns and salinity concentrations can have significant effects on the viability and germination of seeds from the wetland soil seed bank (Leck and Simpson 1995; Peterson and Baldwin 2004; Middleton 2016), further highlighting the role of alternative regeneration mechanisms for wetland resilience. Baldwin et al. (1996) found that short-term saline inundation had little effect on germination from soil seed banks suggesting that soil seed banks may be important for wetland vegetation regeneration following transient salinity disturbances such as storm surge events (Baldwin et al. 1996; Casanova and Brock 2000; Capon and Brock 2006). In contrast, longer term saline flooding, such as that likely to occur as a result of rising sea levels, is more likely to have negative influences on seed bank viability and regeneration capacity. This is particularly important as the combined impacts of reduced germination and an increased frequency and intensity of disturbance events with a changing climate (e.g. storm surge) have the potential to trigger rapid shifts in vegetation structure, as observed by Baldwin and Mendelssohn (1998).
Asexual reproduction and vegetative regeneration in perennial species is important for wetland vegetation resilience when seed germination is inhibited by effects of salinity and flooding (Baldwin et al. 1996; Combroux et al. 2002). In this study, germination abundance, but not diversity, was largely inhibited by saline waterlogging and flooding. Such constraints on this regenerative pathway could result in a reduction of understorey vegetation diversity, favouring species like S. virginicus, which are able to regenerate through vegetative pathways (Leck 1989; Cronk and Fennessy 2001). Species such as the common reed, Phragmites australis, salt marsh grasses (e.g. Sporobolus virginicus and Spartina alterniflora) and sedge (Scirpus americanus) species possess rhizomatous root systems and clonal fragment propagation mechanisms, and have high seed production rates, allowing them to regenerate and spread rapidly throughout wetland areas (Alvarez et al. 2005; Xiao et al. 2010; Marsh et al. 2016). These species are minimally affected by periods of saline and freshwater inundation (Baldwin et al. 2001; Baldwin et al. 2010; Sutter et al. 2014), being able to recover from periods of saline inundation when followed by freshwater flows (Mauchamp and Mésleard 2001). Spartina alterniflora is known to recolonise areas of marsh where dieback has occurred due to disturbance (Alber et al. 2008; Marsh et al. 2016). A similar method of recolonisation after disturbance could be seen with Sporobolus virginicus; however, this has not been reported on. Sporobolus virginicus is commonly the dominant species in salt marshes of eastern Australia, often forming dense grass mats with a thick root zone extending up to 50 cm under the marsh surface (R. Geiger, pers. obs.). This dominance could further inhibit germination from the soil seed bank and the establishment of seeds transported via other mechanisms (e.g. wind and water).
Seedling establishment responses to flooding and salinity
The resilience of wetland systems is dependent on the rate of regeneration post disturbance. The development of seedlings and the further development of the immature plant are important stages in the regeneration niche (Grubb 1977). Although the soil seed bank does not seem to have a significant role in sustaining the vegetation of the study wetland, the success of seedlings which have germinated is vital. This study found, however, that increased flooding and salinity, likely with rising sea levels, can have a significant effect on seedling establishment and development. A general negative trend was found between saltwater treatments and all recorded variables in most species, demonstrating the widespread negative effect of salinity even amongst plants that are known to have some degree of salt tolerance (i.e. S. virginicus and C. glauca). The most pronounced impacts of salinity were observed with respect to height, with growth in most species being reduced by salt by more than half of that observed under freshwater treatments. Of the seedlings investigated in this study, M. quinquenervia and E. tereticornis were observed to be less tolerant of saline flooding, with seedling mortality recorded in almost all saltwater-flooded seedlings and detrimental effects on growth and biomass recorded, even under short pulses of saline flooding. The ongoing effect of saline water intrusion and rising sea levels has been widely recorded for tidal freshwater forests of southern USA, with many authors noting reductions in species diversity, productivity and regeneration (Krauss et al. 2009; Anderson et al. 2013; Xijun et al. 2017). Widespread tree mortality events and reduced seedling regeneration have also been recorded in US tidal freshwater forests experiencing saltwater intrusion and rising sea levels (Doyle et al. 2007; Anderson et al. 2013; Xijun et al. 2017), which have resulted in a transition to tidal marsh or saltmarsh habitat (Krauss et al. 2009; Anderson et al. 2013). A similar fate could be predicted for coastal freshwater wetlands in Southeast Queensland, with a reduction in regeneration capacity from both the soil seed bank and reduced seedling survival. This results in a vegetation community that is susceptible to local extinction if canopy species are removed through disturbance (e.g. cyclones, fire, long-term saline inundation).
Coastal wetlands in eastern Australia are highly valuable for habitat provision and diversity, nutrient cycling, flood mitigation, storm buffering and carbon sequestration (Greenway 2001; Millennium Ecosystem Assessment 2005; Saintilan et al. 2018). Subtropical and temperate coastal saltmarsh are listed as vulnerable under the Environment Protection and Biodiversity Conservation Act (EPBC 1999; Department of Sustainability, Environment, Water, Population and Communities 2013). The increase of saltmarsh area, predicted here through the reduced regenerative capacity and dieback of Casuarina, Melaleuca and Eucalypt habitats on the landward side of the saltmarsh, is beneficial for the species which rely on saltmarsh habitat (i.e. bats, colonial waterbirds and the native water mouse, Xeromys myoides; Department of Sustainability, Environment, Water, Pollution and Communities 2013). The landward Coastal Swamp Oak (Casuarina glauca) forest of New South Wales and SEQ are also protected in the EPBC Act (1999), however as an endangered ecological community due to the very minimal distribution both locally and nationally. The transgression of saltmarsh into these swamp oak communities is concerning considering the extent of urban and agricultural development in SEQ, largely reducing the area that is suitable for swamp oak communities, and the further landward Melaleuca and Eucalypt wetlands, to migrate into.
Coastal squeeze is a key threat of coastal wetland communities, particularly in SEQ where urban and agricultural development is significant. Planners of coastal environments should consider the expansion of existing coastal protected areas to enable the migration of wetlands and integrate modelling of vegetation transitions into decisions-making tools (Rogers et al. 2014a; Shoo et al. 2014). Vegetation communities of Melaleuca and Eucalypt forested swamp should be considered for protection under the national EPBC act 1999 and under state ecosystem protection policies. The conversion of current coastal agricultural/horticultural land back into vegetated wetland could further reduce the risk of CFW loss, providing area for vegetation migration. The zonation and emergence responses found in this study are likely to apply to many CFWs along the coast of Australia; however, given the range of climatic zones within the country, and the variation in climate change predictions, it is important to further investigate these ecosystems. The deficiency of information on small-scale elevation change within coastal wetlands is a key knowledge gap realised in this study. Further investigations should endeavour to include a measure of elevation; however, this could be difficult given the small changes observed in CFWs. Information on the vulnerability of CFWs to both climate change and human development would benefit planners and decision makers, helping to prioritise the conservation of wetlands. Further investigation into all aspects of CFWs within Australia is required to facilitate this; however, priority could be given to the investigation of processes that are known to contribute to the resilience of a wetland system (i.e. sediment accretion; Cahoon 2006; Fagherazzi et al. 2012; Stagg et al. 2016, and vegetative regeneration) and to the identification of areas which may become important areas for CFW migration.
References
Alber M, Swenson EM, Adamowicz SC, Mendelssohn IA (2008) Salt marsh dieback: an overview of recent events in the US. Estuar Coast Shelf Sci 80(1):1–11. https://doi.org/10.1016/j.ecss.2008.08.009
Alvarez MG, Tron F, Mauchamp A (2005) Sexual versus asexual colonization by Phragmites australis: 25-year reed dynamics in a Mediterranean marsh, southern France. Wetlands 25:639–647. https://doi.org/10.1672/0277-5212(2005)025[0639:SVACBP]2.0.CO;2
Anderson CJ, Lockaby BG, Click N (2013) Changes in wetland forest structure, basal growth, and composition across a tidal gradient. Am Midl Nat 170:1–13. https://doi.org/10.1674/0003-0031-170.1.1
Baldwin AH, Kettenring KM, Whigham DF (2010) Seed banks of Phragmites australis-dominated brackish wetlands: relationships to seed viability, inundation, and land cover. Aquat Bot 93(3):163–169. https://doi.org/10.1016/j.aquabot.2010.06.001
Baldwin AH, Egnotovich MS, Clarke E (2001) Hydrologic change and vegetation of tidal freshwater marshes: field, greenhouse, and seed-bank experiments. Wetlands 21(4):519–531. https://doi.org/10.1672/0277-5212(2001)021[0519:hcavot]2.0.co;2
Baldwin AH, McKee KL, Mendelssohn IA (1996) The influence of vegetation, salinity, and inundation on seed banks of oligohaline coastal marshes. Am J Bot 83:470–479. https://doi.org/10.1002/j.1537-2197.1996.tb12728.x
Baldwin AH, Mendelssohn IA (1998) Response of two oligohaline marsh communities to lethal and nonlethal disturbance. Oecologia 116:543–555. https://doi.org/10.1007/s004420050620
Boon P (2012) Coastal wetlands of temperate eastern Australia: will Cinderella ever go to the ball? Mar Freshw Res 63:845–855. https://doi.org/10.1071/MF12205
Bowman D, Prior LD, De Little SC (2010) Retreating Melaleuca swamp forests in Kakadu National Park: evidence of synergistic effects of climate change and past feral buffalo impacts. Austral Ecology 35(8):898–905. https://doi.org/10.1111/j.1442-9993.2009.02096.x
Brisbane City Council, (2017) ‘Boondall Wetlands’, <https://www.brisbane.qld.gov.au/environment-waste/natural-environment/bushland-reserves/boondall-wetlands>. Accessed on 20/05/2017
Brock MA (2011) Persistence of seed banks in Australian temporary wetlands. Freshw Biol 56:1312–1327. https://doi.org/10.1111/j.1365-2427.2010.02570.x
Brock MA, Nielsen DL, Crossle K (2005) Changes in biotic communities developing from freshwater wetland sediments under experimental salinity and water regimes. Freshw Biol 50(8):1376–1390. https://doi.org/10.1111/j.1365-2427.2005.01408.x
Capon SJ (2003) Plant community responses to wetting and drying in a large arid floodplain. River Res Appl 19(5–6):509–520. https://doi.org/10.1002/rra.730
Cahoon DR (2006) A review of major storm impacts on coastal wetland elevations. Estuar Coasts 29(6A):889–898. https://doi.org/10.1007/BF02798648
Capon SJ (2007) Effects of flooding on seedling emergence from the soil seedbank of a large desert floodplain. Wetlands 27:904–914. https://doi.org/10.1672/0277-5212(2007)27[904:EOFOSE]2.0.CO;2
Capon SJ, Brock MA (2006) Flooding, soil seed bank dynamics and vegetation resilience of a hydrologically variable desert floodplain. Freshw Biol 51(2):206–223. https://doi.org/10.1111/j.1365-2427.2005.01484.x
Capon SJ, Chambers LE, MacNally R, Naiman RJ, Davies P, Marshall N, Pittock J, Reid M, Capon T, Douglas M, Catford J, Baldwin DS, Stewardson M, Roberts J, Parsons M, Williams SE (2013) Riparian ecosystems in the 21st centuary: hotspots for climate change adaptation? Ecosystems 16(3):359–381. https://doi.org/10.1007/s10021-013-9656-1
Capon S, Reid M (2016) Vegetation resilience to mega-drought along a typical floodplain gradient of the southern Murray-Darling Basin, Australia. J Veg Sci 27(5):1–12. https://doi.org/10.1111/jvs.12426
Casanova M, Brock MA (2000) How do depth, duration and frequency of flooding influence the establishment of wetland plant communities? Plant Ecol 147:237–250. https://doi.org/10.1023/A:1009875226637
Chambers JC (1993) Seed and vegetation dynamics in an alpine herb field - effects of disturbance type. Can J Botany - Revue Canadienne de Botanique 71(3):471–485. https://doi.org/10.1139/b93-052
Combroux ICS, Bornette G, Amoros C (2002) Plant regenerative strategies after a major disturbance: the case of a riverine wetland restoration. Wetlands 22(2):234–246. https://doi.org/10.1672/0277-5212(2002)022[0234:PRSAAM]2.0.CO;2
Craft C, Clough J, Ehman J, Joye S, Park R, Pennings S, Guo H, Machmuller M (2009) Forecasting the effects of accelerated sea-level rise on tidal marsh ecosystem services. Front Ecol Environ 7(73–78). https://doi.org/10.1890/070219
Cronk JK, Fennessy SM (2001) Wetland plants: biology and ecology: Chapter 7 Community Dynamics in Wetlands. CRC Press. Boca Raton, Fla: Lewis Publishers
David PG (1996) Changes in plant communities relative to hydrologic condition in the Florida Everglades. Wetlands 16:15–23. https://doi.org/10.1007/BF03160642
Department of Sustainability, Environment, Water, Population and Communities (2013) Conservation advice for subtropical and temperate coastal saltmarsh. Canberra: Department of Sustainability, Environment, Water, Population and Communities. Available from: http://www.environment.gov.au/biodiversity/threatened/communities/pubs/118-conservation-advice.pdf
Department of Environment and Heritage Protection, Palustrine ecology, WetlandInfo, Queensland viewed 17 October 2017, <https://wetlandinfo.ehp.qld.gov.au/wetlands/ecology/aquatic-ecosystems-natural/palustrine/>. Accessed 17 Oct 2017
Dowdy A, Abbs D, Bhend J, Chiew F, Church J, Ekstrom M, Kirono D, Lenton A, Lucas C, McInnes K, Moise A, Monselesan D, Mpelasoka F, Webb L, Whetton P (2015) Climate change in Australia projections for Australia’s natural resource management regions: cluster reports. In: Ekstrom M, Whetton P, Gerbing C, Grose M, Webb L, Risbey J (eds) East Coast Cluster Report. CSIRO and Bureau of Meterology, Australia
Doyle TW, Conner WH, Ratard M, Inabinette LW (2007) Assessing the impact of tidal flooding and salinity on long-term growth of bald cypress under changing climate and riverflow. Conner, W. H., Doyle, T. W., Krauss, K. W. (Eds.) Ecology of Tidal Freshwater Forested Wetlands of the Southeastern United States. Springer, Netherlands, pp. 411–445
Environmental Protection and Biodiversity Conservation Act 1999 (Aust) No. 91, 1999, ch5.13, Australian Government
Fagherazzi S, Kirwan ML, Mudd SM, Guntenspergen GR, Temmerman S, D’Alpaos A, van de Koppel J, Rybczyk JM, Reyes R, Craft C, Clough J (2012) Numerical models of salt marsh evolution: ecological, geomorphic, and climatic factors. Rev Geophys. https://doi.org/10.1029/2011RG000359
Finlayson CM, Davidson NC, Spiers AG, Stevenson NJ (1999) Global wetland inventory – current status and future priorities. Mar Freshw Res 50(8):717–727. https://doi.org/10.1071/MF99098
Finlayson CM (2005) Plant ecology of Australia’s tropical floodplain wetlands: a review. Ann Bot 96(4):541–555. https://doi.org/10.1093/aob/mci209
Finlayson CM, Davis JA, Gell P, Kingsford RT, Parton KA (2013) The status of wetlands and the predicted effects of global climate change: the situation in Australia. Aquat Sci 75:73–93. https://doi.org/10.1007/s00027-011-0232-5
Flynn KM, McKee KL, Mendelssohn IA (1995) Recovery of freshwater marsh vegetation after a saltwater intrusion event. Oecologia 103:63–72. https://doi.org/10.1007/BF00328426
Greenway M (2001) Wetlands - wonderlands or wastelands. Keynote paper Regional Wetlands Forum, Wetland Care Australia, Lismore
Grubb PJ (1977) The maintenance of species-richness in plant communities: The importance of the regeneration niche. Biol Rev Camb Philos Soc 52:107–145. https://doi.org/10.1111/j.1469-185X.1977.tb01347.x
Gunn BV (2001) Australian Tree Seed Centre Operations Manual. Canberra: CSIRO Forestry and Forest Products; 2001. legacy:482. doi:https://doi.org/10.4225/08/585821a89fece
Hauser S, Meixler MS, Laba M (2015) Quantification of impacts and ecosystem services loss in New Jersery coastal wetlands due to Hurricane Sandy storm surge. Wetlands 35:1137–1148. https://doi.org/10.1007/s13157-015-0701-z
He YL, Li XZ, Craft C, Ma ZG, Sun YG (2011) Relationships between vegetation zonation and environmental factors in newly formed tidal marshes of the Yangtze River estuary. Wetl Ecol Manag 19(4):341–349. https://doi.org/10.1007/s11273-011-9220-8
Herbert ER, Boon P, Burgin AJ, Neubauer SC, Franklin RB, Ardon M, Hopfensperger KN, Lamers LPM, Gell P (2015) A global perspective on wetland salinization: ecological consequences of a growing threat to freshwater wetlands. Ecosphere 6(10):43. https://doi.org/10.1890/es14-00534.1
Hopfensperger KN, Engelhardt KAM (2008) Annual species abundance in a tidal freshwater marsh: germination and survival across an elevational gradient. Wetlands 28(2):521–526. https://doi.org/10.1672/07-117.1
Hopfensperger KN, Engelhardt KAM, Lookingbill TR (2009) Vegetation and seed bank dynamics in a tidal freshwater marsh. J Veg Sci 20(4):767–778. https://doi.org/10.1111/j.1654-1103.2009.01083.x
IPCC (2014) Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change [Core Writing Team, R.K. Pachauri and L.A. Meyer (eds.)]. IPCC, Geneva, Switzerland, 151 pp.
Isla R, Guillén M, Aragüés R (2014) Response of five tree species to salinity and waterlogging: shoot and root biomass and relationships with leaf and root ion concentrations. Agrofor Syst 88(3):461–477. https://doi.org/10.1007/s10457-014-9705-6
James CS, Capon SJ, White MG, Rayburg SC, Thoms MC, (2007) Spatial variability of the soil seed bank in a heterogeneous ephemeral wetland system in semi-arid Australia. Plant Ecol 190(2):205–217. https://doi.org/10.1007/s11258-006-9201-9
Janousek CN, Folger CL (2014) Variation in tidal wetland plant diversity and composition within and among coastal estuaries: assessing the relative importance of environmental gradients. J Veg Sci 25(2):534–545. https://doi.org/10.1111/jvs.12107
Johnson AKL, Ebert SP, Murray AE (1999) Distribution of coastal freshwater wetlands and riparian forests in the Herbert River catchment and implications for management of catchments adjacent the Great Barrier Reef Marine Park. Environ Conserv 26(3):229–235. https://doi.org/10.1017/s0376892999000314
Kimura H, Tsuyuzaki S (2011) Fire severity affects vegetation and seed bank in a wetland. Appl Veg Sci 14(3):350–357. https://doi.org/10.1111/j.1654-109X.2011.01126.x
Knighton AD, Mills K, Woodroffe CD (1991) Tidal-creek extension and salt water intrusion in northern Australia. Geology 19(8):831–834. https://doi.org/10.1130/0091-7613(1991)019<0831:tceasi>2.3.co;2
Krauss KW, Duberstein JA, Doyle TW, Conner WH, Day RH, Inabinette LW, Whitbeck JL, (2009) Site condition, structure, and growth of baldcypress along tidal/non-tidal salinity gradients. Wetlands 29(2):505–519. https://doi.org/10.1672/08-77.1
Leck MA (1989) Wetland Seed Banks. San Diego, CA: Academic Press
Leck MA, Simpson RL (1995) Ten year seed bank and vegetation dynamics of a tidal freshwater marsh. Am J Bot 82:1547–1557. https://doi.org/10.1002/j.1537-2197.1995.tb13857.x
Leigh C, Bush A, Harrison ET, Ho SS, Luke L, Rolls RJ, Ledger ME (2015) Ecological effects of extreme climatic events on riverine ecosystems: insights from Australia. Freshw Biol 60:2620–2638. https://doi.org/10.1111/fwb.12515
Liu GH, Li W, Li EH, Yuan LY, Davy AJ (2006) Landscape-scale variation in the seed banks of floodplain wetlands with contrasting hydrology in china. Freshw Biol 51(10):1862–1878. https://doi.org/10.1111/j.1365-2427.2006.01621.x
Marcum KB, Murdoch CL (1992) Salt tolerance of the coastal salt marsh grass, Sporobolus virginicus (L.) Kunth. The New Phytologist 120:281–288. https://doi.org/10.1111/j.1469-8137.1992.tb05665.x
Marsh A, Blum LK, Christian RR, Ramsey E III, Rangoonwala A (2016) Response and resilience of Spartina alterniflora to sudden dieback. J Coast Conserv 20(4):335–350. https://doi.org/10.1007/s11852-016-0445-9
Mauchamp A, Mésleard F (2001) Salt tolerance in Phragmites australis populations from coastal Mediterranean marshes. Aquat Bot 70(1):39–52. https://doi.org/10.1016/S0304-3770(00)00140-6
May RM (1977) Thresholds and breakpoints in ecosystems with a multiplicity of stable states. Nature 269(5628):471–477. https://doi.org/10.1038/269471a0
Middleton BA (1999) Wetland restoration, flood pulsing and disturbance dynamics. New York, USA: John Wiley and Sons
Middleton BA (2009) Regeneration of coastal marsh vegetation impacted by hurricanes Katrina and Rita. Wetlands 29(1):54–65. https://doi.org/10.1672/08-18.1
Middleton BA (2016) Effects of salinity and flooding on post-hurricane regeneration potential in coastal wetland vegetation. Am J Bot 103(8):1420–1435. https://doi.org/10.3732/ajb.1600062
Millennium Ecosystem Assessment (2005) Ecosystems and human well-being: wetlands and water synthesis. Washington DC, USA: World Resources Institute
Neubauer SC (2013) Ecosystem Responses of a Tidal Freshwater Marsh Experiencing Saltwater Intrusion and Altered Hydrology. Estuar Coasts 36(3):491–507. https://doi.org/10.1007/s12237-011-9455-x
Nicholls RJ, Wong WP, Burkett VR., Codignotto JO, Hay JE, McLean RF, Ragoonaden S, Woodroffe CD (2007) Coastal systems and low-lying areas. In M. L. Parry, O. F. Canziani, J. P. Palutikof, P. J. van der Linden, & C. E. Hanson (Eds.), Climate change 2007: impacts, adaptation and vulnerability. Contribution of working group II to the fourth assessment report of the Intergovernmental Panel on Climate Change (pp. 315–356). Cambridge: Cambridge University Press
Odum W (1988) Comparative ecology of tidal freshwater and salt marshes. Annu Rev Ecol Syst 19:147–176. https://doi.org/10.1146/annurev.ecolsys.19.1.147
Office of Environment and Heritage. (2011) Freshwater wetlands on coastal floodplains of the NSW north coast, Sydney basin and, South east corner bioregions - endangered ecological community listing: NSW Scientific Committee. Viewed 10 October 2016, <http://www.environment.nsw.gov.au/determinations/FreshwaterWetlandsEndSpListing.htm.>
Olmstead I, Armentano TV (1997) Vegetation of shark slough, everglades national park. SFNRC Technical Report 97-001, South Florida Natural Resources Center, Holmstead, pp 41
Park C, & Allaby M (2013) Resilience. In A Dictionary of Environment and Conservation: Oxford University Press. Retrieved 17 Oct. 2017, from <https://doi.org/10.1093/acref/9780199641666.001.0001/acref-9780199641666-e-6865>
Peirson, W., Davey, E., Jones, A., Hadwen, W., Bishop, K., Beger, M., Capon, S., Fairweather, P., Creese, B., Smith, T. F., Gray, L., Tomlinson, R. (2015). Opportunistic management of estuaries under climate change: a new adaptive decision-making framework and its practical application. J Environ Manag, 163(1), 214–223. doi:https://doi.org/10.1016/j.jenvman.2015.08.021
Peterson JE, Baldwin AH (2004) Seedling emergence from seed banks of tidal freshwater wetlands: response to inundation and sedimentation. Aquat Bot 78(3):243–254. https://doi.org/10.1016/j.aquabot.2003.10.005
Pierce SM, Cowling RM (1991) Disturbance regimes as determinants of seed banks in coastal dune vegetation of the southeastern cape. J Veg Sci 2(3):402–412. https://doi.org/10.2307/3235933
R Core Team (2013) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL http://www.R-project.org/
Reid M, Capon S (2011) Role of the soil seed bank in vegetation responses to environmental flows on a drought-affected floodplain. River Systems 19(3):249–259. https://doi.org/10.1127/1868-5749/2011/019-0022
Reisinger A, Kitching RL, Chiew F, Hughes L, Newton PCD, Schuster SS, Tait A, and Whetton P, (2014) Australasia. In: Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part B: Regional Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change [Barros, V.R., C.B. Field, D.J. Dokken, M.D. Mastrandrea, K.J. Mach, T.E. Bilir, M. Chatterjee, K.L. Ebi, Y.O. Estrada, R.C. Genova, B. Girma, E.S. Kissel, A.N. Levy, S. MacCracken, P.R. Mastrandrea, and L.L. White (eds.)]. Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA, pp. 1371–1438
Rogers, K., Saintilan, N., Copeland, C. (2014aa). Managed retreat of saline coastal wetlands: challenges and opportunities identified from the Hunter River estuary, Australia. Estuar Coasts, 37(1), 67–78. doi:https://doi.org/10.1007/s12237-013-9664-6
Rogers K, Sintilan N, Woodroffe CD (2014b) Surface elevation change and vegetation distribution dynamics in a subtropical coastal wetland: implications for coastal wetland response to climate change. Estuar Coast Shelf Sci 149:46–56. https://doi.org/10.1016/j.ecss.2014.07.009
Ross MS, Reed DL, Sah JP, Ruiz PL, Lewin MT (2003) Vegetation: environmental relationships and water management in Shark Slough, Everglades National Park. Wetlands 11:291–303. https://doi.org/10.1023/B:WETL.0000005541.30283.11
Saintilan N, Rogers K, Kelleway JJ, Ens E, Sloane DR (2018) Climate change impacts on the coastal wetlands of Australia. Wetlands. https://doi.org/10.1007/s13157-018-1016-7
Saintilan N, Rogers K (2013) The significance and vulnerability of Australian saltmarshes: implications for management in a changing climate. Mar Freshw Res 64(1):66–79. https://doi.org/10.1071/mf12212
Saintilan N, Williams RJ (1999) Mangrove transgression into saltmarsh environments in south-east Australia. Glob Ecol Biogeogr 8:117–124. https://doi.org/10.1046/j.1365-2699.1999.00133.x
Sharpe PJ, Baldwin AH (2012) Tidal marsh plant community response to sea level rise: a mesocosm study. Aquat Bot 101:34–40. https://doi.org/10.1016/j.aquabot.2012.03.015
Shoo LP, O’Mara J, Perhans K, Rhodes JR, Runting RK, Schmidt S, Traill LW, Weber LC, Wilson KA, Lovelock CE (2014) Moving beyond the conceptual: specificity in regional climate change adaptation actions for biodiversity in South East Queensland, Australia. Reg Environ Chang 14(2):435–447. https://doi.org/10.1007/s10113-012-0385-3
Stagg CL, Krauss KW, Cahoon DR, Cormier N, Conner WH, Swarzenski CM (2016) Processes contributing to resilience of coastal wetlands to sea-level rise. Ecosystems 19(8):1445–1459. https://doi.org/10.1007/s10021-016-0015-x
Sutter LA, Perry JE, Chambers RM (2014) Tidal freshwater marsh plant responses to low level salinity increases. Wetlands 34(1):167–175. https://doi.org/10.1007/s13157-013-0494-x
Swarth, C. W., Delgado, P., & Whigham, D. F. (2013). Vegetation dynamics in a tidal freshwater wetland: a long-term study at differing scales. Estuaries and coasts, 36(3), 559–574. doi:https://doi.org/10.1007/s12237-012-9568-x
Tait A (1994) Lowland habitat mapping and management recommendations: Tully-Murray catchments: Final Report. Retrieved from Cardwell Shire Catchment Coordinating Committee: Brisbane, Australia
Todd MJ, Muneepeerakul R, Pumo D, Azaele S, Miralles-Wilhelm F, Rinaldo A, Rodriguez-Iturbe I (2010) Hydrological drivers of wetlands vegetation community distribution within Everglades National Park, Florida. Adv Water Resour 33:1279–1289. https://doi.org/10.1016/j.advwatres.2010.04.003
Wall CB, Stevens KJ (2015) Assessing wetland mitigation efforts using standing vegetation and seed bank community structure in neighbouring natural and compensatory wetlands in north-central Texas. Wetl Ecol Manag 23(2):149–166. https://doi.org/10.1007/s11273-014-9366-2
Webb M, Reid M, Capon S, Thoms MC, Rayburg SC, James C (2006) Are flood plain-wetland plant communities determined by seed bank composition or inundation periods. Paper presented at the Sediment Dynamics and the Hydromorphology of Fluvial Systems. Dundee, UK
Westman WE (1978) Measuring the inertia and resilience of ecosystems. BioScience 28(11):705–710. https://doi.org/10.2307/1307321
Wilson SD, Moore DRJ, Keddy PA (1993) Relationships of marsh seed banks to vegetation patterns along environmental gradients. Freshw Biol 29:361–370. https://doi.org/10.1111/j.1365-2427.1993.tb00770.x
Xiao D, Zhang L, Zhu Z (2010) The range expansion patterns of Spartina alterniflora on salt marshes in the Yangtze estuary, China. Estuar Coast Shelf Sci 88(1):99–104. https://doi.org/10.1016/j.ecss.2010.03.015
Xijun L, Conner WH, Song B, Jayakaran AD (2017) Forest composition and growth in a freshwater forested wetland community across a salinity gradient in South Carolina. USA Forest Ecol Manag 389:211–219. https://doi.org/10.1016/j.foreco.2016.12.022
Zoete T (2001) Variation in the vegetation of Melaleuca quinquenervia dominated forested wetlands of the Moreton region. Plant Ecol 152:29–57. https://doi.org/10.1023/A:1011431911988
Acknowledgements
We greatly appreciate the support of B. Brockett and S. Price who assisted with field surveys and experimental setup, B. Coutts who assisted with species identifications and the reviewers who made useful suggestions on improving the manuscript. We thank the staff of Boondall Wetland Education centre for their support and local knowledge.
Funding
Funding was provided by Griffith University School of Environment through the honours program.
Author information
Authors and Affiliations
Corresponding author
Additional information
Editor: Wolfgang Cramer.
Electronic supplementary material
ESM 1
(DOCX 4437 kb)
Rights and permissions
About this article

Cite this article
Grieger, R., Capon, S. & Hadwen, W. Resilience of coastal freshwater wetland vegetation of subtropical Australia to rising sea levels and altered hydrology. Reg Environ Change 19, 279–292 (2019). https://doi.org/10.1007/s10113-018-1399-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10113-018-1399-2