
Abstract
The objective of this study is to assess toll-like receptor-9 (TLR9) expression in CD3-positive T lymphocytes and CD19-positive B lymphocytes in systemic sclerosis (SSc) patients and to study their relation to the extent of skin fibrosis, disease characteristics, and severity as well as the functional status. Fifty-five female SSc patients and 30 matched controls were included. Skin thickness was scored according to the modified Rodnan skin score (mRss). The severity of major organ involvement was assessed using the Medsger severity score (MSS). Scleroderma health assessment questionnaire (SHAQ) was measured to evaluate patients’ functional status. Expression of TLR9 in CD3-positive T lymphocytes and CD19-positive B lymphocytes was studied using flow cytometry. The mean age of the patients was 40.5 ± 9.1 years, and their disease duration was 6.7 ± 3.3 years. There were 21 (38.2%) with diffuse (dcSSc) and 34 (61.8%) with limited cutaneous (lcSSc) subtypes. There was a significant increase in the expression of TLR9/CD3 and TLR9/CD19 in the SSc patients (44.9 ± 18.1 and 24.1 ± 9.6) compared to that in the control (1.4 ± 0.97 and 1.3 ± 0.94; p < 0.0001 for both, respectively) being higher in those with dcSSc. TLR9/CD3 expression was significantly increased in SSc patients with arthralgia/arthritis and digital resorption compared to those without. The TLR9/CD3 significantly correlated with the mRss and MSS (r = 0.37, p = 0.006 and r = 0.31, p = 0.02; respectively). Both the TLR9/CD3 and TLR9/CD19 expressions were significantly correlating (r = 0.53, p < 0.0001). On regression analysis, only TLR9/CD3 was a significant risk factor of the mRss and MSS (β = 0.43, p = 0.009 and β = 0.33, p = 0.015, respectively). TLR9, especially TLR9/CD3, is highly expressed in SSc patients particularly those with dcSSc subtype and could form a potential marker for skin fibrosis and disease severity.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Scleroderma (SSc) is a complex and heterogeneous connective tissue disease mainly characterized by autoimmunity, vascular damage, and fibrosis that mostly involve the skin and lungs [1].
Toll-like receptors (TLRs) represent a family of pattern recognition receptors involved in the recognition and in the defense of the host from invading microorganisms. They sense a wide range of pathogen-associated molecular patterns. TLR activation plays a critical role in the activation of the downstream signaling pathway by interacting and recruiting several adaptor molecules. Studies suggested that TLRs cascade system plays an important role in the pathogenesis of many autoimmune rheumatic diseases such as SSc, systemic lupus, and Bechet disease [2]. In SSc, TLR signaling is likely to be involved in tolerance breakthrough and chronic inflammation via combined Fc gamma receptors and TLR recognition of immune complexes [3]. TLRs on both immune and non-immune cells represent not only the first line of defense against microbial pathogens but are also increasingly being implicated in the progression of chronic autoimmune and fibrotic diseases [4]. Elevated expression of TLR9 has been demonstrated previously in the lungs of patients with idiopathic pulmonary fibrosis (IPF), where its levels were shown to correlate with rapid disease progression [5]. The question of whether increased TLR9 expression in SSc skin correlates with disease severity or activity, or can predict disease progression, awaits further study.
Both intracellular TLRs, such as TLR3, TLR7, and TLR9 and also cell surface TLRs, especially TLR2 and TLR4, play an essential role in the development of autoimmune diseases including SSc and afford multiple therapeutic targets [6]. A critical role of TLR4 signaling in the development of tissue fibrosis makes it a potential therapeutic target in SSc [7]. CD19 deficiency suppresses fibrosis and autoantibody production by inhibiting TLR4 signals in scleroderma [8]. TLR9 is not present at the cell surface; rather, it is synthesized in the endoplasmic reticulum in an inactive form, which upon cell activation, traffics through the Golgi complex to endosomal compartments, where it undergoes proteolytic cleavage [9]. There is strong evidence supporting a pathogenic role of TLR9 in fibrotic skin disease of SSc patients [10]. Epstein-Barr virus is able to persistently infect human SSc fibroblasts inducing an aberrant innate immune response and the TLR activation induces the expression of selected interferon-regulatory factors, transforming growth factor-β1 (TGF-β1), and several markers of fibroblast activation that all play a key role in determining the pro-fibrotic phenotype in SSc [1]. Furthermore, fibroblast activation mediated through TLR9 might be an important factor contributing to the progression of skin fibrosis in SSc [10]. There are emerging evidences for the implication of B cells in the pathogenesis of SSc. B cells from patients with SSc overexpress CD19, a stimulatory B cell receptor (BCR). Expression of interleukin-10 (IL-10) by regulatory B (Breg) cells after stimulation with TLR9 was impaired in patients with SSc [11]. The altered TLR-mediated activation of dendritic cells (DCs) may be responsible for T helper type 2 (Th2)-skewed T cell activation in SSc that may be orchestrated by fibrogenic T cell cytokines, such as IL-4 and IL-13. Targeting DCs could thus offer new avenues for therapeutic intervention [12].
The aim of this work was to assess TLR9 expression in CD3-positive T lymphocytes and CD19-positive B lymphocytes in SSc patients and to study their relation to the extent of skin fibrosis, disease characteristics, and severity as well as the functional status.
Methods
Study subjects
Fifty-five adult female systemic sclerosis patients were recruited from the rheumatology outpatient clinic, Cairo University Hospitals. All patients fulfilled the American College of Rheumatology criteria for systemic sclerosis [13] and grouped as limited (lcSSc) or diffuse cutaneous (dcSSc) subtypes according to LeRoys et al. classification system [14]. Patients with any other associated rheumatic diseases were excluded. Thirty age-matched normal females were included as control. The study conforms to the 1995 Helsinki declaration and was approved by Cairo University Hospitals’ ethical committee. Informed consent was obtained from all patients. Thorough history taking and physical examination were performed to all patients. Skin thickness was scored according to the modified Rodnan skin score (mRss) method [15]. The severity of involvement of major organs was assessed using the Medsger severity score (MSS) [16]. Scleroderma health assessment questionnaire (SHAQ) was measured [17]. Relevant laboratory investigations were performed to patients. Furthermore, plain X-rays of the hands and wrists, high-resolution chest computed tomography (HRCT), and forced vital capacity (FVC) were carried out to all patients.
Flow cytometry for TLR expression in peripheral blood
Samples were split into two tubes, each containing 1 × 106 cells and incubated with surface monoclonal antibodies against CD3- and CD19-conjugated phycoerythrin (PE; BDPharmingen, San Diego, CA, USA), at a concentration of 20 L/1 × 106 cells, in darkness at room temperature for 30 min. The cells were then fixed and permeabilized using an intracellular TLR staining kit according to the manufacturer’s protocol (Imgenex, San Diego, CA, USA). The cells were then incubated with monoclonal antibodies against TLR9 conjugated with fluorescein isothiocyanate (FITC) and their corresponding isotype controls (Invivogen, San Diego, CA, USA), at a concentration of 4 L/1 × 106 cells, in darkness at room temperature for 30 min. Cells were then washed in PBS and assessed using flow cytometry.
Statistical analysis
Statistical Package for Social Science (SPSS) program version 15 was used for analysis of data. Data was summarized as mean ± SD or median and range. Data were tested for normality by Shapiro-Wilk test. Mann-Whitney U test was used for comparing and analysis of two quantitative data. Spearman’s correlation was used for detection of the relation between TLR9/CD3 and TLR9/CD19 with the other variables. Multivariable linear regression tests were used with mRss and MSS were the outcome variables and the results expressed as beta coefficient (β) and p value. p value was considered significant if < 0.05.
Results
The present study included 55 SSc female patients with a mean age of 40.5 ± 9.1 years and disease duration of 6.7 ± 3.3 years as well as 30 matched female control of comparable age (38.4 ± 8.6 years; p = 0.28). The SSc patients were 21 (38.2%) with dcSSc and 34 (61.8%) with lcSSc. Table 1 shows the demographic features, clinical characteristics, medications received, and scores in both dcSSc and lcSSc patients. All the patients had Raynaud’s phenomenon and were receiving a median steroid dose of 10 mg/day (5–15 mg/day). ANA positivity and anti-scl70 were significantly different between dcSSc and lcSSc patients (p < 0.01), Table 2.
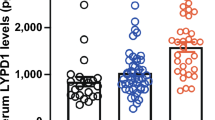
There was a significant increase in the expression of TLR9/CD3 and TLR9/CD19 in the SSc patients (44.9 ± 18.1 and 24.1 ± 9.6) compared to that in the control (1.4 ± 0.97 and 1.3 ± 0.94; p < 0.0001 for both, respectively) (Fig. 1). The TLR9 expression difference in the presence and absence of some clinical manifestations and laboratory manifestations is shown in Table 3.

TLR9/CD3 and TLR9/CD19 expression in diffuse and limited cutaneous systemic sclerosis patients and control. TLR9 toll-like receptor 9, CD cluster of differentiation, dcSSc diffuse cutaneous systemic sclerosis, lcSSc limited cutaneous SSc
Table 4 demonstrates the correlation of TLR9/CD3 and TLR9/CD19 with demographic features, laboratory findings, and the studied scores. There was a significant positive correlation between TLR9/CD3 and TLR9/CD19 expressions (r = 0.53, p < 0.0001). On regression analysis including the age, disease duration, laboratory investigations, and TLR9/CD3 and TLR9/CD19 as independent factors, only TLR9/CD3 was a significant risk factor of the mRss and MSS (β = 0.43, p = 0.009 and β = 0.33, p = 0.015, respectively).
Discussion
Systemic sclerosis is an autoimmune disease in which a breach of tolerance has occurred and leads to fulminant autoimmunity, dysregulated cytokines, pro-fibrotic mediators, and activation of fibroblasts leading to fibrosis via collagen deposition. It has become apparent in recent years that the innate immune system and specifically TLRs are important in disease pathogenesis—activate an innate immune response ultimately leading to release of a variety of factors that initiate and perpetuate fibrosis in SSc [18].
Progressive organ fibrosis due to prolonged fibroblast activation accounts for the intractable nature and high mortality of SSc [19]. Innate immunity is triggered by microbial and endogenous (damage-associated) ligands that are recognized through TLRs expressed on both immune and non-immune cells. The endosomal pathogen sensor TLR9 recognizes nucleic acids and plays vital roles in antiviral defenses [20]. The expression and functional role of TLR9 in the context of fibrosis and SSc were of particular interest [21].
In the present study, there was a significantly increased expression of the TLR9/CD3 and TLR9/CD19 in the SSc patients especially the diffuse cutaneous subtype. In agreement, SSc patients showed upregulation of TLR9 in skin biopsies and myofibroblasts were found to be the major cellular source for it [10]. TLR9, which can be stimulated by nucleosome expression, was upregulated in the affected T cells, B cell IgG production, and proliferation of lymphocytes in SSc [22] and overexpression of CD19 has been reported [11]. An increased TLR response was observed in SSc patients with more inflammatory cytokine secretion [23].
In the current study, the TLR9/CD3 significantly correlated with clinical determinant of skin fibrosis (mRss). In SSc, TLR9 was reported to elicit fibrotic responses mediated via endogenous TGF-β; the damage-associated TLR9 ligands in the skin trigger localized activation of TLR9 signaling, TGF-β production, and consequent fibroblast activation [10]. TLRs in SSc are now considered as emerging therapeutic targets [18]. Disrupting the fibrotic process with inhibitors targeting TLR9 might therefore represent a novel approach to SSc therapy [10]. TLRs have been shown to play a key role in SSc and may represent the link between immune activation, common in SSc, and tissue fibrosis [24].
TLR9/CD3 further significantly correlated with the Medsger severity score. It is also noteworthy that the severity was significantly increased in dcSSc compared to that of lcSSc patients. Among patients with dcSSc, the proportion of TLR9-positive myofibroblasts was significantly greater in those with late-stage disease compared with those with early-stage disease [10]. Damaged cells trigger inflammation through activation of TLRs and TLR9 may play a role in the development of liver fibrosis and hepatocellular carcinoma [25]. Mice lacking TLR9 showed attenuation of fibrosis in the liver [26]. TLR9-dependent activation of myofibroblasts might contribute to the progression of skin fibrosis in patients with SSc and in mice with experimentally induced scleroderma [10].
In the present study, TLR9/CD3 expression was significantly increased in SSc patients with arthralgia/arthritis and digital resorption compared to those without. Serum nucleosome levels, which were able to stimulate TLR9 expression, were higher in SSc patients with joint affections compared with those without; however, it was non-significant [23]. Further data on the activation of B cells in rheumatoid arthritis (RA) being produced by co-activation of the antigen receptor and TLR9 and through TLR9 autoantibody production may sustain the inflammatory response [27]. Many studies showed that TLR9 were overexpressed not only in peripheral blood but also in human synovial tissue of RA patients [28]. TLR9, but not TLR3 or TLR10, gene polymorphism was linked to the susceptibility of RA but not the autoantibodies in Turkish population [29]. Active RA patients show an increased expression of TLR9 on monocyte subsets and display higher production of inflammatory cytokines [30]. TLR stimulation of dendritic cells resulted in higher secretion of IL-6 and tumor necrosis factor-α (TNF-α) in early dcSSc and lcSSc patients [12].
Importantly, patients receiving MTX and CYC tended to have an increased TLR9/CD3 and TLR9/CD19 expression compared to those not receiving. SSc patients with higher disease severity and more organ involvement such as arthritis or IPF are managed with immunosuppressive medications and that may be cause of high TLR9 expression in these patients. TLR9 antagonist reduced dermal and lung fibrosis in SSc model mice [22]. There is an increasing interest in ligands of nucleic acid-sensing TLR, especially TLR9, for pharmacological intervention in various diseases. Antimalarial drugs have been reported to act as TLR7/9 antagonists [31]. TLR-mediated signaling provides numerous potential therapeutic targets for development of therapies for the treatment of multi-systemic autoimmune diseases [32].
The present study has several strengths. To our knowledge, the present study is the first one to reveal the correlation of serum expression of TLR9 with several clinical and laboratory features of fibrosis and severity in SSc patients. This study opens window for prospective research to address the expression of TLR9 on different CD4(+) and CD8(+) T cell sub-populations (naïve, central memory, effector memory, and terminally differentiated effector memory). Nevertheless, this study has some limitations. First, the study population was all female SSc, which may limit the generalizability of the results. Second, most of patients had long disease duration and the expression of TLR9 in the early stage of the disease needs to be addressed. Third, prospective data to evaluate the role of TLR9 as early marker for fibrosis progression were not available.
In summary, TLR9, especially TLR9/CD3, is highly expressed in SSc patients particularly those with dcSSc subtype and could form a potential marker for skin fibrosis and disease severity. Disrupting the fibrotic process, especially in dcSSc patients, with inhibitors targeting TLR9 or its downstream signaling pathways might represent a novel therapeutic approach.
References
Farina A, Cirone M, York M, Lenna S, Padilla C, Mclaughlin S et al (2014) Epstein-Barr virus infection induces aberrant TLR activation pathway and fibroblast-myofibroblast conversion in scleroderma. J Investig Dermatol 134(4):954–964
Gianchecchi E, Fierabracci A (2015) Gene/environment interactions in the pathogenesis of autoimmunity: new insights on the role of toll-like receptors. Autoimmun Rev 14(11):971–983
Santegoets KCM, van Bon L, van den Berg WB, Wenink MH, Radstake TRDJ (2011) Toll-like receptors in rheumatic diseases: are we paying a high price for our defense against bugs? FEBS Lett 585(23):3660–3666
Bryant CE, Gay NJ, Heymans S, Sacre S, Schaefer L, Midwood KS (2015) Advances in toll-like receptor biology: modes of activation by diverse stimuli. Crit Rev Biochem Mol Biol 50(5):359–379
Trujillo G, Meneghin A, Flaherty KR, Sholl LM, Myers JL, Kazerooni EA et al (2010) TLR9 differentiates rapidly from slowly progressing forms of idiopathic pulmonary fibrosis. Sci Transl Med 2(57):57ra82
Liu Y, Yin H, Zhao M, Lu Q (2014) TLR2 and TLR4 in autoimmune diseases: a comprehensive review. Clin Rev Allergy Immunol 47(2):136–147
Takahashi T, Asano Y, Ichimura Y, Toyama T, Taniguchi T, Noda S et al (2015) Amelioration of tissue fibrosis by toll-like receptor 4 knockout in murine models of systemic sclerosis. Arthritis Rheumatol 67(1):254–265
Yoshizaki A, Iwata Y, Komura K, Ogawa F, Hara T, Muroi E et al (2008) CD19 regulates skin and lung fibrosis via toll-like receptor signaling in a model of bleomycin-induced scleroderma. Am J Pathol 172(6):1650–1663
Chockalingam A, Brooks JC, Cameron JL, Blum LK, Leifer CA (2009) TLR9 traffics through the Golgi complex to localize to endolysosomes and respond to CpG DNA. Immunol Cell Biol 87(3):209–217
Fang F, Marangoni RG, Zhou X, Yang Y, Ye B, Shangguang A et al (2016) Toll-like receptor 9 signaling is augmented in systemic sclerosis and elicits transforming growth factor beta-dependent fibroblast activation. Arthritis Rheumatol 68(8):1989–2002
Mavropoulos A, Simopoulou T, Varna A, Liaskos C, Katsiari CG, Bogdanos DP et al (2016) Breg cells are numerically decreased and functionally impaired in patients with systemic sclerosis. Arthritis Rheumatol 68(2):494–504
van Bon L, Popa C, Huijbens R, Vonk M, York M, Simms R et al (2010) Distinct evolution of TLR-mediated dendritic cell cytokine secretion in patients with limited and diffuse cutaneous systemic sclerosis. Ann Rheum Dis 69(8):1539–1547
Hoogen F, Khanna D, Fransen J, Johnson SR, Baron M, Tyndall A et al (2013) Classification criteria for systemic sclerosis: an American College of Rheumatology/European League against Rheumatism collaborative initiative. Arthritis Rheumatol 65(11):2737–2747
LeRoy EC, Black C, Fleischmajer R, Jablonska S, Krieg T, Medsger TAJ et al (1988) Scleroderma (systemic sclerosis): classification, subsets and pathogenesis. J Rheumatol 15(2):202–205
Clements P, Lachenbruch P, Siebold J, White B, Weiner S, Martin R et al (1995) Inter and intraobserver variability of total skin thickness score (modified Rodnan TSS) in systemic sclerosis. J Rheumatol 22(7):1281–1285
Medsger TAJ, Bombardieri S, Czirjak L, Scorza R, Della Rossa A, Bencivelli W (2003) Assessment of disease severity and prognosis. Clin Exp Rheumatol 21(3 Suppl 29):S42–S46
Steen VD, Medsger TAJ (1997) The value of the health assessment questionnaire and special patient-generated scales to demonstrate change in systemic sclerosis patients over time. Arthritis Rheum 40(11):1984–1991
O’Reilly S (2017) Toll like receptors in systemic sclerosis: an emerging target. Immunol Lett. https://doi.org/10.1016/j.imlet.2017.09.001
Allanore Y, Simms R, Distler O, Trojanowska M, Pope J, Denton CP et al (2015) Systemic sclerosis. Nat Rev 1:15002
Mogensen TH (2009) Pathogen recognition and inflammatory signaling in innate immune defenses. Clin Microbiol Rev 22(2):240–273 Table of Contents
Bhattacharyya S, Varga J (2015) Emerging roles of innate immune signaling and toll-like receptors in fibrosis and systemic sclerosis. Curr Rheumatol Rep 17(1):474
Yoshizaki A, Taniguchi T, Saigusa R, Fukasawa T, Ebata S, Numajiri H et al (2016) Nucleosome in patients with systemic sclerosis: possible association with immunological abnormalities via abnormal activation of T and B cells. Ann Rheum Dis 75(10):1858–1865
van Bon L, Cossu M, Scharstuhl A, Pennings BWC, Vonk MC, Vreman HJ et al (2016) Low heme oxygenase-1 levels in patients with systemic sclerosis are associated with an altered toll-like receptor response: another role for CXCL4? Rheumatology (Oxford) 55(11):2066–2073
Ciechomska M, Cant R, Finnigan J, van Laar JM, O’Reilly S (2013) Role of toll-like receptors in systemic sclerosis. Expert Rev Mol Med 15:e9
Song IJ, Yang YM, Inokuchi-Shimizu S, Roh YS, Yang L, Seki E (2017) The contribution of toll-like receptor signaling to the development of liver fibrosis and cancer in hepatocyte-specific TAK1-deleted mice. Int J Cancer. https://doi.org/10.1002/ijc.31029
Watanabe A, Hashmi A, Gomes DA, Town T, Badou A, Flavell RA et al (2007) Apoptotic hepatocyte DNA inhibits hepatic stellate cell chemotaxis via toll-like receptor 9. Hepatology 46(5):1509–1518
Sacre SM, Drexler SK, Foxwell BM (2005) Toll-like receptors and rheumatoid arthritis: is there a connection?. In: O’Neill LAJ, Brint E (eds) Toll-like Receptors in Inflammation. Birkhauser, Basel, p 19–40
Tamaki Y, Takakubo Y, Hirayama T, Konttinen YT, Goodman SB, Yamakawa M et al (2011) Expression of toll-like receptors and their signaling pathways in rheumatoid synovitis. J Rheumatol 38(5):810–820
Etem EO, Elyas H, Ozgocmen S, Yildirim A, Godekmerdan A (2011) The investigation of toll-like receptor 3, 9 and 10 gene polymorphisms in Turkish rheumatoid arthritis patients. Rheumatol Int 31(10):1369–1374
Lacerte P, Brunet A, Egarnes B, Duchêne B, Brown JP, Gosselin J (2016) Overexpression of TLR2 and TLR9 on monocyte subsets of active rheumatoid arthritis patients contributes to enhance responsiveness to TLR agonists. Arthritis Res Ther 18(1):10
Sun S, Rao NL, Venable J, Thurmond R, Karlsson L (2007) TLR7/9 antagonists as therapeutics for immune-mediated inflammatory disorders. Inflamm Allergy Drug Targets 6(4):223–235
Fullard N, O’Reilly S (2015) Role of innate immune system in systemic sclerosis. Semin Immunopathol 37(5):511–517
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The study conforms to the 1995 Helsinki declaration and was approved by Cairo University Hospitals’ ethical committee. Informed consent was obtained from all patients.
Disclosures
None.
Rights and permissions
About this article

Cite this article
Gheita, T.A., Sayed, S., Azkalany, G.S. et al. Toll-like receptor 9 in systemic sclerosis patients: relation to modified Rodnan skin score, disease severity, and functional status. Clin Rheumatol 37, 757–763 (2018). https://doi.org/10.1007/s10067-017-3880-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10067-017-3880-6