
Abstract
To adapt forest ecosystems and forest management to climate change, it is essential to know which forest regions and which tree species are resilient to climate variability and which ones are possibly affected most by past and anticipated future changes. In this contribution, for the main forest regions of Türkiye and six tree species, recent climate variability and trends were quantified and statistically correlated to record tree defoliation and vitality. Climate variables considered are maximum temperature (Tmax), minimum temperature (Tmin), mean temperature (Tmean), and total precipitation (Prcp), which are compared to forest health responses recorded as part of the International Cooperative Programme on Assessment and Monitoring of Air Pollution Effects on Forests (ICP Forests) on 277 plots across forests along the Black Sea and Mediterranean regions. In addition, long-term data on satellite measurements of the normalized difference vegetation index (NDVI) were extracted for the same 277 plots for the period 2008–2020. Firstly, 30 years (1991–2020) of reanalysis of climate variables from ECMWF were extracted for all plots; secondly, individual correlations and cross-correlations of climate variables and tree health and vitality were computed for the period 2008–2020 (significance level of 95%) for the four most dominant species from the Black Sea forests (F. orientalis, Q. cerris, P. sylvestris, P. orientalis) and two species from Mediterranean forests (P. brutia and C. libani). Temperature showed a stronger effect on most species than precipitation. Finally, time-lagged correlations were analyzed for seven-time lags (significance level of 95%) to evaluate legacy effect. The analysis revealed that different tree species from the two regions show different responses to climate variables. Species in the Mediterranean region are more resistant to droughts and climatic variations. Legacy effects of defoliation and NDVI have lasted for at least 2 years.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
Climate variability and extremes have been shown to increasingly pose a risk to forest ecosystems (Haynes et al. 2014; Sanchez-Salguero et al. 2017; Seidl et al. 2017; Limaki et al. 2021; Song et al. 2018). There are numerous studies that link climatic factors to forest health and vitality through abiotic (climate and weather extremes, air pollutants, forest fires) and biotic (pests, disease) factors (Marco et al. 2014; Gea-Izquierdo et al. 2019; Gottardini et al. 2020). Prediction of these factors is important for adapting future forest plans and sustainable forest ecosystem management (Deng et al. 2021). Beside investigating climatic conditions, use of remote sensing and defoliation data is crucial for assessing forest health. Many studies for many regions on different tree species have been conducted to improve the assessment of defoliation and tree responses to climate variability using remote sensing (Fernandez-Manso et al. 2011; Townsend et al. 2012; Zhu et al. 2018; Pasquarella et al. 2021) A study by Kovalev et al. 2021 shows that use of seasonal normalized difference vegetation index (NDVI) dynamics makes it possible to classify plots according to the tree conditions. Results of assessing impact of extreme climatic events on aspen defoliation using NDVI note that there are predictable patterns of aspen defoliation that identify aspen stands vulnerable to climate extremes (Currit and St Clair 2010).
Türkiye also faces a number of forest stressors including fires, climate change, droughts, insects, and fungi, the part where the fires are most intense is the coastline from Antalya to Istanbul (Ozturk et al. 2010). Since the 1970s droughts due to the decreasing trend in total amount of precipitation in the Mediterranean region has been one of the confronted problems (Turkes 2012). There were 2.793 forest fires in 2021, resulting in the loss of 139.503 hectares of forest in Türkiye, which is approximately eleven times greater than the average loss for the period 2017–2020 (GDF 2022). Since the early 1980s, daily maximum and minimum temperatures in Türkiye have increased (Toros 2012b). Changes in the frequency, intensity, and duration of weather extremes with droughts and flash floods caused by extreme temperatures and precipitation events were experienced (Tayanç and Toros 1997; Kömüşçü et.al. 1998; Toros 2012a; Baltacı 2017). The number of hot days, the number of heatwaves, and the duration of heatwaves were found to be significantly correlated (95% significance level) with fire occurrences in the period 1997–2006 in the western part of Türkiye (Unal et al. 2013). According to a recent trend analysis of heatwave characteristics, the frequency and length of heat waves in Türkiye have increased since 1950, and the Black Sea and the Mediterranean coasts have experienced the most significant changes in heat wave characteristics since the 1980s (Erlat et al. 2021). As given above, despite the abundance of literature showing climate variability and extreme behaviors in Türkiye, the studies that explain how these results effect on forest dominant tree species are insufficient. Some of the available studies related to this subject; a study by Ayan et al. (2022) based on the prediction of warmer conditions in the future with different scenarios, Fagus orientalis Lipsky. populations of the southern slopes of the North Anatolian Mountains were predicted to shrink and the populations in main distribution areas of Istranca Mountains and Northern Anatolia would be reduced, particularly populations in transition zone Central Anatolia. Also, it would lose its isolated-marginal populations in the southeast of Türkiye. The global habitat suitability area of Cedrus libani would enlarge in the forecasted scenarios with respect to the present, although it would be more restricted in 2070 due to the altitudinal shift (López-Tirado et al. 2021). This paper aims to explore which climate variables impact forest health in Türkiye for the period 2008–2020, so unlike previous studies, NDVI, defoliation and climate variables were conducted together for dominant tree species in the Black Sea and Mediterranean forests.
The International Co-operative Programme on Assessment and Monitoring of Air Pollution Effects on Forest (ICP Forests), ICP monitored plots in Türkiye provide a unique dataset to analyze the details of forest health responses to climate and their changes over time. In this study, collected data at the ICP sites combined with climate and remote-sensed data could contribute to understand the drivers of forest health conditions in Türkiye; hereby, provide crucial knowledge towards sustainable and climate-adapted management of Turkish forests. No such studies have been conducted in the Mediterranean and Black Sea regions. These forest regions are the backbones of commercial forestry in Türkiye and their health conditions are of concern in Türkiye (GDF 2020).
In this study, we investigate how observed climate variability and change impacted tree crown defoliation in different geographic regions of Türkiye through the three main points as follows:
-
Identify recent climate trends in the main forest regions and for the distribution areas of different tree species in Türkiye
-
If the climate changed recently identify which forest regions and which tree species are sensitive to climate variability, expressed by defoliation and vitality (using NDVI)
-
If a sensitivity is identified and determine which climate variables (precipitation temperature) explains over which time span most of the observed defoliation or change in NDVI.
To address those questions we aim to explain surveyed tree crown conditions assessed at ICP plots and also changes in satellite-derived NDVI for 277 plots across Türkiye for the period 2008–2020. We focus on six dominant tree species including Oriental beech (Fagus orientalis), Türkiye oak (Quercus cerris), Scots pine (Pinus sylvestris), Oriental spruce (Picea orientalis), Turkish pine (Pinus brutia), and Lebanese cedar (Cedrus libani). We focus on the two main forest regions, the Black Sea coast and the Mediterranean coast. Within the scope of this work, we will use individual correlations between short term climate variability and tree responses in terms of defoliation and NDVI, cross-correlations between climate variables and defoliation and NDVI, respectively, and finally time-lagged correlations to assess legacy effects and the time-scale of climate variables on forest health. Results will be analyzed both in a spatial (geographical) and temporal (decadal) context.
2 Data and methodology
For Europe and beyond, the International Co-operative Programme on Assessment and Monitoring of Air Pollution Effects on Forest (ICP Forests) has provided an international comprehensive framework and monitoring system since 1985 to assess the spatio-temporal variation of forest conditions at selected plots (Lorenz 1995). The goal of the ICP forest network is to provide a comprehensive compilation of information on the condition of forests (health, productivity) in Europe, including Türkiye. ICP assesses the impacts of natural and human-induced disturbances in order to understand cause-effect relationships between these factors and the condition of forest ecosystems (Lorenz 2010). The ICP Forest assessments exist at two monitoring intensity levels: In 2020, there were 5852 Level I plots across Europe on a grid of 16 × 16 km and 623 Level II plots that are a selected forest ecosystem. Forty-two countries including Türkiye have participated in the program. Initially, ICP assessments in Türkiye were launched in the 1990s, but they were discontinued due to lack of qualified staff and funding (Tolunay et al. 2010; Karakas et al. 2007; Koray et al. 2007). Since 2007, continuous observation has been implemented in Türkiye with 597 Level I plots and 52 Level II plots as of 2019 (Michel et al. 2020).
Defoliation is measured at Level I sites. Defoliation is regarded as a measure of tree health because it represents the percentage of leaves missing in the current crown in comparison to a fully foliated reference tree (Potočić et al. 2021). In the ICP forest plot surveys, there are two types of reference trees: local reference trees and absolute reference trees. Most countries including Türkiye have adopted local reference trees as standards (Eichhorn et al. 2016). Local reference trees or conceptual trees are the most suitable trees with full foliage for a particular site, considering factors such as altitude, latitude, tree age, site conditions, and social status. Their defoliation is set to 0%. Defoliation based on local reference trees is the most assessed variable in forest monitoring and international field exercises dating back to the 1980s (Michel et al. 2020). Even though accepting crown defoliation as a core parameter of forest health, it has also caused some critics due to the bias that can be caused by the observer and different measuring methods (Innes 1993; Eichhorn et al. 2010). Nevertheless, it is commonly considered an approved indicator for forest health (De Marco et al. 2014; Zierl 2004).
2.1 Study area
2.1.1 Ecology of forests
Fundamental studies were conducted to categorize phyto-geographical regions in Türkiye. Three types of phyto-geographical regions are defined for Türkiye which are the Euro-Siberian, Mediterranean, and Irano-Turanian (Davis 1971; Zohary 1971, Avcı 1996). These regions have a particular vegetation and climatic characteristic. The Euro-Siberian region covers the entire Black Sea coast and the Black Sea coast of Thrace, climate throughout the year with high precipitation ratio and low seasonal temperature differences and foggy weather are climatic conditions (Çolak and Rotherham 2006, Atalay 2002) The Mediterranean region includes western and southern part of the Anatolia and the Amanos mountains. Maquis and dry forests at high elevations and dry, hot summers with warm and rainy winters are the characteristics of the region (Çolak and Rotherham 2006, Atalay 2002). These two regions are mostly forested, and also the most densely populated areas (Atalay et al. 2014). There are different types of forests with more than 450 tree species and shrubs (Çolak and Rotherham 2006). The detailed investigations mentioned above into forest structure groups according to ecological regions in Türkiye, which helped to determine our study areas. Hence priority was given to the tree species of the Black Sea/northern Marmara ecological regions (hereafter “Black Sea Forests”) and Mediterranean/western Aegean ecological regions (hereafter “Mediterranean Forests”). Based on the most common trees found (Bahadır and Emet 2010), four species from the Black Sea forests (F. orientalis, Q. cerris, P. sylvestris, P. orientalis) and two species from Mediterranean forests (P. brutia and C. libani) were selected. A study by are cited in text but not provided in the reference list.
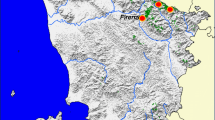
Please provide references in the list or delete these citations. Eşen et al. (2015) notes that F. orientalis is the principal commercial tree in Türkiye and mostly growing in Black Sea Region. Q. cerris is a native and widely distributed species and oak species constitute 76.4% of broadleaf forests in Türkiye (Özer 2014). One main distributional areas of P. sylvestris in Türkiye are the western and eastern part of Black Sea region and mixed P. sylvestris, forests are commonly associated with F. orientalis and P. orientalis in the eastern part of Black Sea region (Atalay et al. 2014). P. orientalis is a native species in the Eastern Black Sea Region (Tüfekçioğlu et al. 2011). P. brutia is one of the important forest trees of the Mediterranean Region, reaches its greatest distribution in southern Türkiye and one of the most valuable commercial trees (Kayacık 1980). C. libani naturally distributed in Türkiye, Syria, Lebanon, and Southern Türkiye has the largest forest area of C. libani in the world with 324,453 hectares (Boydak and Çalıkoğlu 2008). Of the 277 plots, 138 plots were attributed to Black Sea forests and 139 plots to Mediterranean forests (Fig. 1). Two plots from the Mediterranean, and 25 from the Black Sea forests, in total 27 plots out of 277 had two of the species present, F. orientalis (73), Q. cerris (25), P. sylvestris (50), P. orientalis (16), P. brutia (118), C. libani (22) as seen in Fig. 1.

© European Union, Copernicus Land Monitoring Service 2018, EEA)
The locations of forests (green areas), ecological regions (red labels refer to the climate regions as called in the text) and the 304 ICP plots in Türkiye with the 277 plots contain at least one of the six dominant tree species in the study highlighted by different symbols: F. orientalis (73), Q. cerris (25), P. sylvestris (50), P. orientalis (16), P. brutia (118), and C. libani (22). 27 ICP plots contain more than one selected tree species. Land Cover was retrieved from CORINE Land Cover data (
According to GDF (2009), the forested areas in Türkiye cover 21.2 million hectares, constituting 27.2% of the total land area. The forested areas in Türkiye are mostly concentrated in the Black Sea region (BSR), Marmara region (MR), Aegean region (AR), and Mediterranean region (MeR), while Central Anatolia (CAR), East Anatolia (EAR), and Southeastern Anatolia (SEAR) regions are forest-poor regions (Fig. 1). The forests in Türkiye are separated into roughly 50% productive and 50% unproductive. Sixty percent of the forests in Türkiye are coniferous and 40% are broad-leaved dominated trees. The majority of broad-leaved trees are Quercus sp (oak) tree species, which constitute 30.2% of the total forestry regions and the majority of conifers are Pinus nigra (black pine) and P. brutia (red pine), which constitute 19.8% and 25.2% of the total forestry regions. Regarding the geographical areas of Türkiye, the forested area by region is as follows: Black Sea (24.4%), Mediterranean (19.6%), Aegean (15.9%), Marmara (12.8%), Middle Anatolia (13.8%), East Anatolia (8.3%), and Southeast Anatolia (5.8%). In Türkiye, ICP Forest level I plots are found mostly in the Black Sea and Mediterranean regions since most forested areas are located here (GDF 2020).
2.1.2 Climate of the study area
The plots in the Black Sea forests experience dominantly temperate climates that are year-round humid with warm summers (Cfb); the plots in the Marmara ecological regions are also found with climates that experience hot summers (Cfa) while the higher-altitude mountain forests and forests of the Backward Black Sea Plateau experience continental cold climates with warm summers (Dfb) (Kottek et al. 2006). Generally, most of the Black Sea forest plots are wet in every season, the share of summer (sum of months June, July, and August, hereafter “JJA”) precipitation in the annual total is about 19.4%. In contrast, the Mediterranean forests are in temperate climates with dry and hot summers (Csa), partially also continental cold climates with dry and warm summers (Dsa) in the interior. For the plots in the Mediterranean forests, summer precipitation (JJA) in the annual accounts for only about 5.7% (Atalay 1997).
2.2 Data sources
2.2.1 Defoliation data
In 2020, 607 plots are available over Türkiye. In this study 277 plots out of 607 were selected considering only the six most dominant tree species (Table 1) from main forestry regions of Türkiye. For defoliation data in the 277 plots were provided by the Ministry of Agriculture and Forestry for the period 2008–2020 which is the longest available period for defoliation. Defoliation is categorized into five classes, from 0% (no defoliation) to 100% (dead tree). It has five different classes based on reference trees on the plots such as Class 0 up to 10% (none), Class 1 > 10–25% (slight), Class 2 > 25–60% (moderate), Class 3 > 60–100% (severe), and Class 4 = 100% (standing dead trees only) (Michel et al. 2020). Class 4, namely dead trees, is not included in this study. Measurement of defoliation is conducted only once each year during the months of June, July, August, and September. In this study, the plots whose measurements were completed in the summer season (June, July, August; hereafter JJA), were used. September is an autumn month that has different characteristics than the summer months and was not considered in this study. In addition, the percentage of missing data in the defoliation data as follows by tree species; F. orientalis is 12.8%, Q. cerris 25%, P. sylvestris 16%, P. orientalis 6%, P. brutia 14%, and C. libani 16%.
2.2.2 Climate data
Climate variables were obtained from the fifth generation ECMWF reanalysis data ERA5-land (C3S 2017) obtained for the period 2008–2020 at a spatial resolution of 0.25° × 0.25° and an hourly temporal resolution. For each plot, data from the grid cell containing the coordinates of the corresponding ICP plot were extracted. For each ICP plot, daily maximum air temperature at 2 m (Tmax), daily minimum air temperature at 2 m (Tmin), and daily mean air temperature at 2 m (Tmean) were calculated from hourly ERA5 data over the entire period 2008–2020. Similarly, daily precipitation (Prcp) was calculated as the sum of hourly ERA5 precipitation data for each ICP plot.
The standardized precipitation index (SPI) and standardized precipitation evapotranspiration index (SPEI) were used similar to other studies investigating drought effects on vegetation (Sirdaş and Sen 2003; Polong et al. 2019; Danandeh Mehr and Vaheddost 2020). Previous studies showed that temperature has a critical role in drought severity (Stagge et al. 2015; Abbasi et al 2019; Eris et al. 2020). While SPI (McKee et al. 1993) works solely based on precipitation, SPEI (Vicente-Serrano et al. 2010) additionally includes maximum and minimum temperature to estimate evapotranspiration (Frank et al. 2017). Hargreaves (Hargreaves 1994) was used to estimate reference evapotranspiration since besides maximum and minimum temperature data, latitude data is also available for ICP plots. SPI and SPEI calculations were conducted using R packages SPI and SPEI (Begueria and Vicente-Serrano 2023) for each ICP plot separately over different periods, including 1, 2, 3, 6, 9, 12, and 24-month backwards based on ERA5 Tmax, Tmin, and Prcp data for the period 2008–2020.
2.2.3 Remote sensing data
In addition to ground-based defoliation assessments as surveyed at ICP plots, satellite-based metrics such as normalized difference vegetation index (NDVI) can also show responses of vegetation vitality at the stand-level to varying climate variables. NDVI has been used to assess how climate variability affects vegetation growth especially in climate-sensitive areas (John et al. 2013; Zhang et al. 2016). Studies on climate change and its responses of NDVI of vegetated areas have been conducted globally over the past few decades (Pravalie et al. 2014; Schmidt et al. 2014; Shawul and Chakma 2020; Salvador and de Brito 2018), and the number of the studies for Türkiye due to its location, intricate climate, climate change sensitivity, and unique topography has increased (Arekhi et al. 2018; Akkoyunlu et al. 2019; Koç et al. 2020). For satellite-based metric, we used MOD13A3 version 6 at 1 × 1 km resolution to retrieve monthly NDVI for the period 2008 to 2020 from application for extracting and exploring analysis ready samples (AρρEEARS) (https://lpdaac.usgs.gov). AppEEARS provides an efficient way to use spatial temporal and layer parameters.
2.3 Statistical analysis methodologies
2.3.1 Trend analysis
Long-term in Tmean, Tmax, Tmin, and Prcp were calculated for the 30-year climate period 1991–2020 using linear regressions on the ERA5 reanalysis data separately for each grid cell in which an ICP plot was located and separately for the full-year and the summer (JJA) period. Long-term trends in NDVI and mean plot defoliation (defoliation) per species were calculated over the 13-year period 2008–2020 (summer only), due to limited data availability.
2.3.2 Correlation analysis
Pearson correlation coefficients (Rodgers and Nicewander 1988) were computed to assess linear relationships between summer (JJA) average Tmean, Tmax, Tmin, Prcp, NDVI and summer defoliation to check for correlations between interannual climate variability and tree responses for the period 2008–2020. Statistical significance was evaluated according to linear regression results at a 95% confidence interval (p-values ≤ 0.05 (*)), and were evaluated at three different levels with p-values ≤ 0.05 (*), p-values ≤ 0.01 (**), and p-values ≤ 0.001 (***).
2.3.3 Principal component analysis
For analyzing cross correlations between the above climate variables, NDVI and defoliation for the period 2008–2020 (summer only), additionally principal component analysis (PCA) was used (Anderson 1958; Kendall 1968). The method has aimed at the selection of significant components and the dimensionality reduction. Specifically, PCA identifies the directions of maximum variance in high-dimensional data and projects it onto a smaller dimensional subspace while retaining most of the original information (Hartmann et al. 2018). PCA has been widely adopted in various fields such as meteorology, climatology, and forestry to detect elementary features of physical phenomena by simplifying data sets (Camperlin and Diop 2003; Sneyers et al. 1989; Pandžić et al. 2009; Tadić et al. 2019). This study has applied PCA to identify the main components of climate affecting forest health conditions of different tree species.
2.3.4 Time-lagged correlations
To test if there are delayed effects or legacy effects of climate extremes on tree health, time-lagged correlations were performed. Correlations with a moving window were calculated using the Pearson correlation coefficients between Tmax, Tmean, Tmin, Prcp, and Tmean anomalies over periods of increasing length back from a measurement when NDVI/defoliation was measured. Time-lagged correlations were performed for the period 2008–2020 using seven different time windows backwards from a date when defoliation and/or NDVI were measured. These time windows are considering the time from the defoliation/NDVI measurement date back to the previous 1, 2, 3, 6, 9, 12, and 24 months (here after T1, T2, T3, T6, T9, T12, T24) with respect to the defoliation date of the year based on plots from each species. For each time window Tmax, Tmean, Tmin, Prcp, and Tmean anomalies were calculated relative to the same period of the year in the entire 2008–2020 period. The significant tests of the partial correlation coefficients were based on the Student t-tests at a significance level of 95%.
3 Results
3.1 Climatic conditions
3.1.1 Annual averages, variability, and change
The average annual total precipitation (Prcp) in the period 1991–2020 in the Black Sea forest plots was 1033 mm (range 559–2453 mm). Generally, highest annual Prcp is found on the forest plots along the Eastern Black Sea coast. Annual average air temperature (Tmean) in the Black Sea forest plots was 9.90 °C (3.42–15.04 °C). The average annual Prcp in the forest plots of the Mediterranean forests for the same period is 810 mm (356–1275 mm). Annual Tmean in the Mediterranean forest plots was on average 14.68 °C (7.81–19.73 °C).
The average interannual variability as assessed by the standard deviation of annual Tmean from 1991 to 2020 is 0.92 K (0.69–1.26 K) for the plots of the Black Sea forests, and 0.74 K (0.56–1.07 K) for the plots of the Mediterranean forests. Average interannual Prcp variability is 122.45 mm or 11.8% of the average annual total (66.67–297.10 mm) for the plots of the Black sea forests, and 144.73 mm or 17.8% (63.80–255.69 mm) for the plots of the Mediterranean forests. The interannual variabilities in Tmean and Prcp by tree species are shown in Table 2.
Over the period from 1991 to 2020, for n = 277 plots, namely all investigated plots a statistically significant positive trend of annually averaged Tmean was found (Fig. 2a). Averaged over all plots of the Black Sea forests, annual Tmean experienced a statistically significant trend of + 0.70 K/decade. For the Mediterranean forests annual Tmean experienced a statistically significant trend of + 0.57 K/decade. Only for n = 7 plots (four from the Eastern Black Sea region, three from the Western Mediterranean region) a statistically significant positive trend of annual total Prcp was found. A total of n = 31 plots, all from the Middle Black Sea region, experienced a statistically significant negative trend of annual total precipitation and n = 239 plots had no statistically significant trend (Fig. 2e). Averaged over all plots of the Black Sea forest plots, annual Prcp had no statistically significant trend, Similarly, averaged over all Mediterranean forest plots, annual Prcp had no statistically significant trend. The trends in Tmean and Prcp separated by tree species are shown in Table 2.

a–e Statistically significant trends (95% confidence interval) are shown for the 1991–2020 period in a annual mean temperature, b summertime mean temperature, c annual max temperature, d summertime max temperature, e annual total precipitation, f summertime total precipitation. Gray points refer to the plots having no significant trend for the period
3.1.2 Summer season averages, variability, and change
The average summertime (JJA) total precipitation (Prcp) in the period 1991–2020 in the Black Sea forest plots was 239.62 mm (range 71.32–744.38 mm). Again, highest summertime Prcp is found on forest plots along the Eastern Black Sea coast. The average summertime Prcp in the forest plots of the Mediterranean forests for the same period is 53.07 mm (3.21–214.86 mm). The share of JJA Prcp in the annual total Prcp for the Black Sea and Mediterranean are 6.53% and 21.49%, respectively. Summertime average (JJA) average air temperature (Tmean) in the Black Sea forest plots was 18.80 °C (14.07–23.93 °C) and in the Mediterranean plots it was 24.17 °C (18.71–28.13 °C).
Figure 3 contrasts summer (JJA) Tmean and total Prcp individually from all 277 selected plots; the tree species are shown in different colors. Clear ecoclimatic differences for different tree species and regions are revealed: Generally highest JJA Tmean and lowest JJA Prcp are experienced by forest plots containing P. bruita, while C. libani is found in similarly dry (Mediterranean) conditions, but with generally lower Tmean The forest species of F. orientalis, Q. cerris, and P. sylvestris are found over a range of summertime climatic conditions of the western Black Sea region, while forests with P. orientalis experience clearly more humid conditions of the eastern Black sea with highest summertime Prcp.

Total summer (JJA) precipitation vs average summer (JJA) Tmean (°C) for all 277 plots for the 2008–2020 period (includes 304 dots)
The average interannual variability of summer air temperatures as assessed by the standard deviation of JJA Tmean from 1991 to 2020 is 0.86 K (0.68–1.30 K) for the plots of the Black Sea forests, and 0.73 K (0.54–0.97 K) for the plots of the Mediterranean forests. Average interannual summer precipitation variability based on JJA Prcp is 65.08 mm or 27.16% (22.05–163.61 mm) for the plots of the Black sea forests, and 25.83 mm or 48.67% (3.66–77.63 mm) for the plots of the Mediterranean forests. The interannual variability in Tmean and Prcp by tree species is shown in Table 3.
Over the period from 1991 to 2020, for n = 209 plots a statistically significant positive trend of summer (JJA) Tmean was found and n = 68 plots had no statistically significant trend (Fig. 2b). Averaged over all plots of the Black Sea forests, JJA Tmean experienced a statistically significant trend of + 0.61 K/decade. For the Mediterranean forests, JJA Tmean experienced a statistically significant trend of + 0.30 K/decade. Similarly, for n = 53 plots a statistically significant positive trend in summertime (JJA) total Prcp was found. n = 6 plots had a statistically significant negative trend of JJA total Prcp and n = 218 plots had no statistically significant trend (Fig. 2f). Averaged over all plots of the Black Sea forests, summer (JJA) Prcp has no statistically significant trend. For the Mediterranean forests JJA Prcp has no statistically significant trend. The trends in JJA Tmean and Prcp by tree species are shown in Table 3.
3.2 Forest health indicators
3.2.1 Defoliation
Defoliation was assessed on all 277 plots from 2008 to 2020 on more than 4300 individual trees (Table 4). Overall, Q. cerris had the highest fraction of trees with strong defoliation. 20% of all Q. cerris trees assessed were in defoliation Class 2 or 3. This is followed by P. Brutia (12% of all P. Brutia trees assessed are in defoliation Class 2 or 3) and F. orientalis (12%) and P. sylvestris (10%), P. orientalis and C. libani showed the lowest fraction of trees in defoliation Classes 2 and 3 among the six species studied with 7 and 3%, respectively. P. orientalis had 67% of all trees in Class 0 (no defoliation). F. orientalis (44%), C. libani (48%), P. sylvestris (47%) experienced about half of the trees in Class 0 (no defoliation), Q. cerris (29%) and P. brutia (21%) had fewest trees with no defoliation. The number of trees into each defoliation class by year are given in Table 4.
Defoliation of all selected tree species during the 2008–2020 period was 17.14% in the Black Sea forests and 18.36% in the Mediterranean forests. Separated by species, the defoliation during the 2008–2020 period was 16.68% for F. orientalis, 22.67% for Q. cerris, 17.47% for P. sylvestris, 13.51% for P. orientalis: 19.03% for P. brutia, and 15.40% for C. libani. Interannual variation in summertime defoliation was recorded as follows: F. orientalis, 6.48%; Q. cerris, 7.73%; P. sylvestris, 5.31%; P. orientalis, 4.93%; P. brutia, 4.60%; C. libani, 4.04%.
Over the 13 years of the dataset, four tree species experienced a statistically significant trend in summertime defoliation, namely F. orientalis (− 9.8%/decade***), Q. cerris (− 8.7%/decade**), P. sylvestris (− 5.8%/decade***), and P. orientalis (− 3.9%/decade**), P. brutia (0.4% /decade) and C. libani (1.6%/decade) experienced no statistically significant trends.
3.2.2 NDVI
The 13-year summertime (JJA) NDVI averages for the 2008–2020 period for plots containing the species are as follows: Plots containing F. orientalis 0.84 (0.70–0.91), Q. cerris 0.72 (0.49–0.87), P. sylvestris: 0.70 (0.39–0.90), P. orientalis: 0.81 (0.63–0.89), P. brutia: 0.52 (0.26–0.74), C. libani: 0.47 (0.29–0.63). The NDVI interannual variability for the summertime (JJA) 2008–2020 period by species are as follows: Plots containing F. orientalis, 0.02 (0.01–0.07); Q. cerris, 0.03 (0.02–0.07); P. sylvestris, 0.02 (0.01–0.05); P. orientalis, 0.02 (0.01–0.04); P. brutia, 0.03 (0.01–0.06); and C. libani, 0.03 (0.02–0.04).
The following trends in summertime (JJA) NDVI were identified for the 2008–2020 period for plots containing the corresponding species: F. orientalis, + 0.03**; Q. cerris, + 0.04*; P. sylvestris, + 0.04**; P. orientalis, + 0.03***; P. brutia, + 0.04**; C. libani, + 0.05 ** per decade.
3.3 Individual correlations between climate variables and tree health
Correlations between summertime (JJA) averaged climate variables (Tmax, Tmean, Tmin, Prcp) and tree health indicators (defoliation, NDVI) were calculated for each tree species separately; all plots containing a species were combined to calculate correlations irrespective of geographic location in Table 5. As shown in Fig. 4, it is evident that average Tmax, Tmean, Tmin, total Prcp of JJA, and defoliation values for each species have also varied considerably between plots.

Box plot graphs of average Tmean, daily Tmin, and daily Tmax for JJA (°C), total precipitation of JJA (mm), and defoliation of JJA (%) for 2008–2020 period based on tree species from 277 plots over Black Sea and Mediterranean regions
In addition, correlations between climatic variables and forest health conditions (defoliation, NDVI) were calculated for each individual plot; and the Pearson correlation coefficient (r) values between the average JJA Tmax and total JJA Prcp plot averaged defoliation and NDVI are shown in Fig. 5. All correlations are calculated for the 2008–2020 period. In addition to Tmax, r values have also been calculated for Tmean and Tmin, however, Tmax results have been highlighted because results were more pronounced. Although it has been found that summertime (JJA) Tmax and total Prcp vary with geographic location, a clear regional distinction in the climate variability is found between Mediderranean forests and Black Sea forests: P. brutia and C. libani have mainly varied in total precipitation of JJA between 25 and 250 mm, in average maximum temperature of JJA between 23 and 34° C. F. orientalis, Q. cerris, P. sylvestris, and P. orientalis have changed between 100 and 800 mm in total precipitation of JJA and changed between 19 and 29 °C for the average maximum temperature of JJA.

Results of the Pearson correlation coefficient (r) between total Prcp of JJA-average Tmax of JJA defoliation and NDVI based on tree species for the 2008–2020 period
Figure 5 also gives the opportunity to interpret the ranges of climate variability in which tree responses are higher. It is seen that some plots with low precipitation values are highly correlated with NDVI. Part of the P. orientalis plots, which experience higher total Prcp, show low/negative correlation with values > 500 mm between NDVI and Prcp.
3.4 Cross-correlations between climate variables and tree response
For the PCA, the initial variables are Tmean, Tmax, Tmin, Prcp, defoliation, and NDVI (Fig. 6). The target variable is defoliation class. Each dot in Fig. 6 refers to a plot and single year (2008–2020), the number of dots is different for the species since the number of plots varies for each tree species and for each year. Selected plots containing Q. cerris and P. sylvestris had defoliation values reported up to Class 3, while the other four species showed maximum defoliation only up to Class 2. The most common defoliation values reported are in classes 0 and 1, which means that none or slight defoliation are more frequent at a plot level.

PCA results for the annual 2008–2020 period based on total precipitation of JJA, average Tmean, Tmin, Tmax of JJA, defoliation, and NDVI. The colored areas show the range of the four defoliation classes (Defo_Class) with respect to climate variables for each species. The arrows in PCA represent the vectors that correspond to the initial variables from which principal components have been computed. An angle between two vectors represents the correlation between the variables: small angles show a strong positive correlation, right angles indicate a lack of correlation, and opposite angles indicate a high negative correlation (Rossiter 2014)
Aside from C. libani and P. sylvestris, other species show a negative correlation between defoliation and NDVI. In cases where defoliation and NDVI are positively correlated, as in C. libani and P. sylvestris, the defoliation class is 2, meaning that defoliation has increased by 25–60%.
Except for P. sylvestris, the correlation between NDVI and Prcp is positive or strongly positive. P. orientalis has the smallest angle between NDVI and Prcp, showing a strong positive correlation.
NDVI correlates positively with Tmin for P. orientalis, P. sylvestris and P. brutia and also correlates positively with Tmean for Q. cerris, P. sylvestris, and P. orientalis. Except for P. Sylvestris and F. orientalis, NDVI has right angles with Tmax so there is no correlation between NDVI and Tmax. Similarly, F. orientalis and C. libani also have right angles between NDVI and Tmean. Where Prcp and NDVI correlate positively, the defoliation degree is either none or slight as in P. orientalis, F. orientalis, and Q. cerris. As a result of the PCA, differences in the variables behaviors based on species become more evident, which demonstrates that the relationships between variables are species-specific.
3.5 Spatio-temporal correlation analysis
3.5.1 Climate variables, drought indices and defoliation
Pearson correlation coefficients (r) between defoliation and drought indices, and climate variables on the 277 plots for the period 2008–2020 are shown in Fig. 7. On the maps it is evident that the correlation between Tmax or Tmean and defoliation is generally positive in the Mediterranean and along the western Black Sea Coast, on the other hand there is generally a negative correlation between defoliation and drought indices in these regions. The eastern Black Sea and the eastern Mediterranean regions show the most positive correlations between Prcp and defoliation, furthermore the same behavior is valid for the correlations between drought indices and defoliation.

Pearson correlation coefficient (r) values between climate variables anomalies (total Prcp of JJA and average Tmean, Tmin, and Tmax of JJA)—drought indices and defoliation for the 2008–2020 period for all 277 plots
3.5.2 Climate variables, drought indices and NDVI
Based on the analysis of the relationship between climate variables and NDVI, the plots and the species in the Eastern Black Sea region have different correlation values than the rest of the region. For the Eastern Black Sea, correlations between average Tmax, Tmean, Tmin and the NDVI are high; however, the correlations between the total Prcp and the NDVI are lower than the rest of the Black Sea region. Also, the correlations between Tmin and NDVI show a different pattern for both of the regions (Fig. 8). The number of the plots with positive correlation have increased greatly for the species in the Black Sea forests and for the Mediterranean forests, the strong negative correlation seen in Tmax and Tmean was replaced by weak negative values in Tmin. Unlike in the Black Sea forests, there is no regionally differentiated behavior in Mediterranean forests. The NDVI has mostly positive correlation with Prcp (91% of the plots), but has negative correlation with Tmean (70% of the plots), Tmax (79% of the plots), and Tmin (52% of the plots). For the relationships between NDVI and monthly, 3-month SPEI, SPI is mostly positive by 90%, 92%, 90%, and 93% of the plots, respectively. Most of the negative correlations belong to the plots found in the Eastern Black Sea Region.

Pearson correlation coefficient (r) values between climate variables anomaly (total precipitation of JJA and average Tmean, Tmin, and Tmax of JJA)—drought indices and NDVI for the 2008–2020 period for all 277 plots
3.6 Evaluation of time lagged correlations
In order to determine on which time-scales the climatic drivers have strongest correlations with measured NDVI, defoliation and drought indices, correlations between climate variables and tree health/vitality indicators were analyzed for seven different time lags. First, monthly mean Tmax, monthly mean Tmean, monthly mean Tmin, total Prcp over the entire period, monthly Tmean anomaly, and monthly Prcp anomaly were determined for the previous 1, 2, 3, 6, 9, 12, and 24 months (here after T1, T2, T3, T6, T9, T12, T24). The time windows were chosen based on the measurement date of defoliation as reported for each plot separately, back 1, 2, 3, 6, 9, 12, and 24 months from the date of the defoliation measurement. For NDVI and drought indices, correlations were calculated for the month of the defoliation measurement and compared to the climate variables in the variable windows (Figs. 9 and 10). Significance tests of the correlation coefficients were separately calculated for each time window based on the Student t-tests at a significance level of 95%.

Pearson correlation coefficients (r) between average Tmean, cumulative Prcp and NDVI for the previous 1, 2, 3, 6, 9, 12, and 24 months (time lags were calculated based on plot location from the day defoliation was measured in summertime) for the entire 2008–2020 period

Pearson correlations coefficient (r) between climate variables and defoliation for windows of 1, 2, 3, 6, 9, 12, and 24 months backwards (the windows are calculated from the day the defoliation is measured) for the 2008–2020 period
F. orientalis and Q. cerris experienced the strongest legacy effect in correlation Prcp anomalies and NDVI. Median values for F. orientalis and Q. cerris in T1, T2, T3, T6, T9, T12, and T24 are − 0.06, 0.04, 0.10, − 0.16, 0.15, 0.16, and 0.20 and 0.02, 0.10, 0.17, 0.20, 0.10, 0.14, and 0.38, respectively. However, for all time-lags there is no significant correlation. P. sylvestris experienced the strongest legacy effect for the correlation between Tmean anomalies and NDVI, median values in T1, T2, T3, T6, T9, T12, and T24 were 0.26, 0.35, 0.38, 0.20, 0.15, 0.16, and 0.33, respectively. In this case, correlations are significant in all time-lags. P. orientalis which has the wettest conditions, experienced the strongest legacy effect in correlation between Tmean anomalies and NDVI; median values in T1, T2, T3, T6, T9, T12, and T24 were 0.17, 0.37, 0.33, 0.17, 0.20, 0.27, and 0.48, respectively. P. brutia and C. libani experienced the strongest legacy effect in correlation Prcp anomalies and NDVI; however, correlations are not significant except for P. brutia in T2, T3, and T6 (SI). Median values for P. brutia and C. libani in T1, T2, T3, T6, T9, T12, and T24, respectively, were 0.12, 0.22, 0.27, 0.11, 0.21, 0.19, and 0.30 and − 0.15, 0.24, 0.29, 0.11, 0.24, 0.17, and 0.21.
Overall P. brutia experienced the strongest legacy effect for the correlations (Figs. 9 and 10). Also Mediterranean species, P. brutia and C.libani showed stronger and more consistent correlations than Black Sea species. Regarding the median line of the time lag, the majority of the tree species showed different behaviors in T6 and beyond compared to T1, T2, and T3.
In addition, there is no significant correlation for Prcp anomaly and NDVI for all species and time lags except for P. brutia in T2, T3, and T6, for Prcp anomaly and defoliation in T2, T3, T6, T9, T12, T24 for P. orientalis, and in T6, T9, T12, T24 for C. libani. For some of the time-lagged correlations, F. orientalis, Q. cerris, and P. sylvestris have no statistically significant correlation between Tmax, Tmean, Tmin, and defoliation.
4 Discussion
All tree species experience higher variability of summertime Prcp than annual Prcp proportionally to the total summer and annual precipitation. The Black Sea forests experience higher summertime Prcp and lower interannual relative summertime Prcp variability. Also the Black Sea region has a slightly higher interannual summertime temperature variability, with values of 0.79 K, 0.80 K, 0.97 K, 0.86 K for plots containing F. orientalis, Q. cerris, P. sylvestris, and P. orientalis, respectively. When considering selected tree species, tree species common for the Black Sea forests (e.g., F. orientalis, Q. cerris, P. sylvestris, P. orientalis) show a higher rate of defoliation than Mediterranean forests tree species (e.g., P. brutia, C. libani) which experiences significantly lower summertime precipitation. Overall defoliation rate, when all species were included in the analysis, the average defoliation of plots in the Mediterranean forest was slightly higher than in the Black Sea forests. Despite the large differences in precipitation and temperature values between the two forests, the fact that the defoliation rate difference is very low, indicates that the species of the Mediterranean region; C. libani and P. brutia are better adapted to climate variability (Sevik and Ertürk 2015; Messinger et al. 2015).
All four tree species dominantly found in the Black Sea forests show a statistically significant decreasing trend in defoliation over the 2008–2020 period. Simultaneously, the plots containing these species show concurrently a statistically increasing trend in NDVI. This means that changes in the tree-level indicators are also reflected in changes to plot-level data over decadal scale. However, on an interannual scale only Q. cerris showed a statistically significant correlation between annual summertime NDVI at plot level and annual summertime defoliation measurements at tree level.
For the Mediterranean climate, the natural vegetation of the coastal zone consists of pine species, which have a high demand for warmth and light and are resistant to drought, and, in places where they are destroyed, maquis cover the place which is consists of small trees or shrubs with an average height of 1–2 m and always green. In high places, coniferous larch, cedar and fir forests dominate, for the Black Sea climate; the natural vegetation consists of broad-leaved moist forests in the coastal areas and coniferous forests that grow in cold and humid conditions in the higher elevations (Atalay 1997, 2010; Günal 2013; Dönmez 1985). A study by Efe and Sönmez 2015 state that most of the broad-leaved forest trees are humid species and form “humid forests.” These are mostly spread in the Black Sea Region. Conifers, especially P. brutia, which is a common species of the Mediterranean region, are considered a “dry forest” element. As a result, Mediterranean forests are generally more drought and less dense than Black Sea forests, which is an explanation why NDVI could statistically significantly increase but defoliation does not show a significant trend.
Regarding defoliation and NDVI trends for the 2008–2020 period, there is a decrease in NDVI and an increase in defoliation across most plots in 2008–2009 and 2013–2014 (SI) which is explained by extreme drought conditions at the beginning of the study period in 2008 and also during the year 2013. 2008 experienced the lowest total annual precipitation of the last 40 years across Türkiye. Mediterranean forests experienced the lowest and second lowest total annual precipitation of the last 20 years in 2008 and 2013. For the Black Sea forests, conversely 2013 was the year with the lowest and 2008 was the year with the second lowest total annual precipitation over the last 20 years (TSMS 2022). In line with this, the temperature and precipitation values at the forest plots showed that 2008 is one of the driest years for the summertime for the forests as well. In summer 2008, the Mediterranean forest plots experienced a compound effect namely very low precipitation and unusually high temperatures at the same time. For the Black Sea forest plots, 2008 and 2013 were on average the third and fifth driest years for the summertime over the 1991–2020 period. For the Mediterranean forest plots, the JJA period in 2008 and 2013 were the second and fourth driest summer periods over the 1991–2020 reference period. In addition, summer 2008 was the third hottest summer for the Mediterranean forest plots over the reference period. The years following the droughts, 2009 and 2014, had a higher amount of precipitation than the long term average but still a lower NDVI and a higher defoliation compared to the 13-year average. Tree damages in 2008 and 2013 were severe and 2009 and 2014 showed legacy effects from the previous year’s drought as expected, even though they did not experience severe conditions. From 2010 to 2020, 91.7% of the plots (254) show an upward trend in NDVI and regarding each plot; 55% for F.orientalis, 50% for Q.cerris, 46% for P. sylvestris, 69% for P. orientalis, 34% for P. brutia, and 36% for C.libani show either decreasing or no trend in defoliation (SI, Fig. S11).
F. orientalis showed statistically significant trends between mean plot NDVI and Tmax and Prcp. With increasing precipitation and decreasing maximum and mean temperature on plots (SI, Fig. S2-S5), NDVI of F. orientalis plots significantly increased during the 2008–2020 period. For Q. cerris, P. sylvestris, P. brutia significant correlations between NDVI and Tmin and Tmean were found, but no influence on precipitation variability could be detected. P. orientalis and C. libani tree species did not show any direct correlations between climate variables and NDVI.
Regarding Tmax, Tmean, Tmin, and Prcp variables, P. brutia and C. libani have a different range from the four tree species from the Black Sea region. At this point, the ecological characteristics of the species reveal themselves. F. orientalis and Q. cerris do not need very special conditions to grow, F.orientalis has a wide distribution along with the Black Sea Region (Kandemir and Kaya 2009). Q. cerris exhibits strong morphological variability and ecological adaptability (Simeone et al. 2019). P. orientalis exists in more specific conditions. It actually reflects the characteristics of extreme wet conditions in the Eastern Black Sea (Atalay et al. 2014). P. brutia is one of the light-tree species and has a very high-temperature demand. It is the most resistant pine species to temperature and drought (Fady et al. 2003). A study by Messinger et al. (2015) concluded that the exceptional drought tolerance of C. libani was confirmed by comparing tree ring growth in natural stands at Elmali, southwest Türkiye. Although the rainfall total at Elmali was very low (68 mm) and mean air temperatures were high (18 °C) during summertime. C.libani tolerates extreme cold in winter and prolonged droughts during summer. Semerci et al. (2017) studied drought stress of P. sylvestris in different regions from Europe and Türkiye, there are significant differences among populations of P. sylvestris, while populations in northern Europe have significantly weaker tolerances to drought compared to southern and high-elevation populations, which exhibit generally lower growth rates and more resistance to drought. In addition, P. brutia has been found to be highly drought-resistant and can effectively be used for reforestation in disturbed areas (Sevik and Ertürk 2015). Recently, there has already been an increase in the mean annual temperature and severity of summer droughts in the Mediterranean region (Abbasnia and Toros 2018). Having drought-resistant tree species in the Mediterranean region is really promising since future climate scenarios indicate that the Mediterranean region will be among the most heavily affected parts of the world in terms of increased frequency and intensity of drought events (Ozturk et al. 2015; Türkeş et al. 2018).
The correlations between climate variables and defoliation and drought indices have revealed that the temperature effect in the regions is stronger than the precipitation. However not only climate variables are affecting defoliation in the forests but also ecological factors such as insect calamities (Ataman 1967; Yüksel and Akbulut 2005; Ülgentürk et al. 2012; GDF 2022), non-sustainable forest management (Baskent et al. 2008), illegal logging (Gunes and Elvan 2005) may cause defoliation. The results of our study are subject to these uncertainties.
The spatial variability in JJA average Tmean and Prcp is larger for the tree species in the Black Sea region, which is due to the fact that they can be found over larger geographic areas and are adaptive to different climates (Fig. 3). While the Black Sea forests are more suitable for the species with lower summertime temperature and higher summer precipitation demand, the Mediterranean forests are mainly suitable for species with high summertime temperature and low summer precipitation demand. A detailed study by Günal (2013) has reported similar climate evaluation for the regions.
In 2009, defoliation was assessed on 563 plots including 12,290 trees over Türkiye. The mean defoliation was 18.9% for conifers, and 21.5% for broadleaves, respectively. In 2009, on 18.7% of the monitored trees, defoliation was more than 25%. P. brutia had the highest defoliation among conifers (Fischer et al. 2010). Similarly, in this study P. brutia had the highest defoliation (19.03%) for the 2008–2020 period among the assessed conifers including P. brutia, C. libani, P. sylvestris, and P. orientalis. The altitude of the distribution area of P. brutia is lower than other coniferous species (Table 1), The Mediterranean region has high average and maximum temperatures (Atalay 1997). The shortlength of the growing period (Boydak 2004) and the high average and maximum temperatures may encourage this species to reduce water loss by transpiration as a precaution against drought. On the other hand, it is the most important conifer species from the commercial perspective; its wood is used for many purposes: construction, industry, carpentry, firewood, and pulp in Türkiye (Fady et al. 2003; Petrakis et al. 2007; Tolunay et al. 2008).
In particular, forest fires in the western Mediterranean are considered a great risk (Özcan et al. 2018); however, this may not be the case in the Black Sea region. Tüfekçioğlu et al. (2011) based on RegCM3, reported that temperatures will increase 2–4 °C in the Black Sea region, and precipitation will increase 200–300 mm in the eastern part of the region in the next century. They also concluded that while fire could pose a threat to the western part of the Black Sea region, it does not threaten the eastern part—P. orientalis ecosystems, since their ecosystems have a humid climate through the summer. According to Bilgili (1997), fires in Türkiye are distributed as follows: 41% in the Aegean, 25% in the Mediterranean, 22% in the Marmara, and 13% in the rest. Summer is the season of most forest fires in Türkiye where extreme temperature and drought are observed, followed by spring with 48%, (Bahadır 2010). In Fig. 4, the findings support previous studies on forest fire tendency in the Mediterranean region mentioned earlier due to the low precipitation and high temperatures of the plots.
In this study, PCA confirmed the climatological results of previous studies (Türkeş 2012; Kadıoglu 2000; Toros 2012a) that explained the general characteristics of precipitation and temperature in Türkiye. Relationships between the parameters are species-specific due to spatial factors like altitude, latitude, distance from sea (Usta et al. 2018) along with ecosystem differences (Atalay 2002). PCA results have revealed the correlation differences between climate variables and forest responses for the six tree species.
In this paper, the geographic distribution of the species (Günal 2013) and the exposure to different climatic features (Atalay et al. 2014) in local environments may have influenced the individual correlation results to have low correlation values. The occurrence of defoliation can be the result of weather extremes, air pollution, or biological factors (Popa et al. 2017), so tree responses to climate variability can be different even in the same subregion due to compounding factors or factors not even linked to weather extremes. Defoliation is also one of the most difficult and expensive data to collect, the ICP network (Level I) provides an opportunity to investigate the relationships between tree defoliation and climate variables, considering a large number of species and large spatial variations of species (Fischer et al. 2012).
The Student t-test (95% significance level) was performed for the seven time lags (T1, T2, T3, T6, T9, T12, T24) with climate variables including Tmean, Prcp, Tmax, Tmin, Prcp anomalies, Tmean anomalies, drought indices including the SPI, the SPEI and defoliation and NDVI, respectively. A study by de la Cruz et al. (2014) for the period similarly also reported that there is a relationship between climate variables and defoliation was proved by statistical methodologies and previous years’ thermal conditions determine current year defoliation conditions for the 1987–2012 period. Similarly, to other studies (Wu et al. 2018; Li et al. 2020) in the past, the results revealed that the legacy effect of parameters continued even for at least the previous 2 years, except for Prcp anomaly and NDVI. The studies conducted on relatively similar regions also showed there was no significant correlation between Prcp anomaly and NDVI. For instance, no significant correlations were found in the forest of the northeast, in some parts of southern China as well as in some desert areas of the northwest (Chen et al. 2020). No significant correlations between Prcp anomaly and NDVI were found in major parts of the mountain ranges, in some semi-desert and desert areas of southern Kazakhstan and Turkmenistan as well as in the forest steppe in northern and northeastern Kazakhstan (Gessner et al. 2013). Detailed information about the topography and climate properties is provided by Atalay et al. (2014) and Günal (2013).
In this study, defoliation analyses have been done with 13 years of data (2008–2020). It is the longest available period for the ICP plots across Türkiye however historical knowledge and climate responses of tree species could be presented better with longer data periods of data. So the survey should be continued to get precise results. Also timely coordinated defoliation observation for the same tree species can be one of contributions to better understanding correlation with climate variables and satellite metrics.
5 Summary and conclusion
The most important findings of this study are the following:
-
Tree level defoliation and plot-level NDVI show concurrent trends in Black Sea forests species F. orientalis, Q. cerris, P. sylvestris, and P. orientalis.
-
Forest health as assessed by NDVI and defoliation in the Black Sea region is statistically linked to climate variability, strongest effects were detected for F. orientalis.
-
Forest health assessed by NDVI and defoliation could not be linked to climate variability in the Mediterranean forests.
-
Legacy effect of climate variabilities on NDVI and defoliation was observed for up to 2 years. This is a fundamental result to better understanding of the tree species responses.
-
Regarding defoliation rate and interannual variabilities, Mediterranean species, P. brutia and C. libani, are more resistant to drought and climate variabilities.
Annual climate variables trends showed that Tmean and Tmax have statistically significant and remarkable trends for each species for the 1991–2020 period. Summer climate variables trends showed that F. orientalis, Q. cerris, P. sylvestris, and P. orientalis in the Black Sea region have statistically significant trend in Tmean and Tmax for the 1991–2020 period however the tree species in the Mediterranean region have no statistically significant trend. On the other hand, Prcp has a significant trend for none of the tree species in annual and summertime trends for the 1991–2020 period.
According to the individual correlations and PCA result of this study, it was concluded that correlation assessment between ground-based (defoliation)-satellite-based (NDVI) data and climate variables made on the basis of the tree species regarding defoliation classes is more reliable than the individual correlation assessment based on the tree species.
Although the highest interannual Prcp variability was seen in the Mediterranean region there was no increasing trend for defoliation. It means P. brutia and C. libani have higher adaptive capacity to drought. Similarly, statistically significant positive NDVI trends were seen in the Mediterranean region and the trend value is slightly higher than the Black Sea region on average per decade. Additionally, the tree species in the Black Sea region experienced a decreasing defoliation trend in a range − 3.9 to − 9.8%/decade.
Regarding tree species, while the Black Sea region is more sensitive to precipitation than temperature, the Mediterranean region is more sensitive to temperature than precipitation. However, regarding both our statistically significant correlation results for different time lags and the extreme events and climate projections based on the previous studies, for the long run, the temperature has a greater influence on tree species in the regions due to expected temperature rises. In terms of the time lag, the results show that the legacy effects of climate variables, satellite-based data (NDVI), and ground-based data (defoliation) last for 2 years. Nevertheless, no significant correlations between Prcp anomaly and NDVI were found for the species except for the P. brutia in T2, T3, and T6. Similarly, the earlier studies conducted similar results for the densely populated and mountainous regions. While there are clear correlations found between NDVI and climatic anomalies, there was no clear relationship found on the plot-level between NDVI and defoliation data. This paper is one of the first to apply the subject of climate variability and forest health to the ICP Forest plots in Türkiye, highlighting the importance of integrating these fields together. Similar to the studies in the past, it has been shown once again that due to the special topographic structure of Türkiye and climate variabilities, further studies on tree species and vegetation should be conducted by taking into consideration sub-climate zones.
Data and materials availability
Available after request.
Code availability
Available after request.
References
Abbasi A, Khalili K, Behmanesh J, Shirzad A (2019) Drought monitoring and prediction using SPEI index and gene expression programming model in the west of Urmia Lake. Theor Appl Climatol 138:553–567. https://doi.org/10.1007/s00704-019-02825-9
Abbasnia M, Toros H (2018) Analysis of long-term changes in extreme climatic indices: a case study of the Mediterranean climate, Marmara Region. Türkiye Pure Appl Geophys 175:3861–3873. https://doi.org/10.1007/s00024-018-1888-8
Akkoyunlu BO, Baltaci H, Tayanc M (2019) Atmospheric conditions of extreme precipitation events in western Türkiye for the period 2006–2015. Nat Hazards Earth Syst Sci 19:107–119.https://doi.org/10.5194/nhess-19-107-2019
Anderson TW (1958) An introduction to multivariate statistical analysis. John Wiley & Sons, New York
Arekhi M, Yesi A, Ozkan UY, Sanli FB (2018) Detecting treeline dynamics in response to climate warming using forest stand maps and Landsat data in a temperate forest. For Ecosyst 5:23. https://doi.org/10.1186/s40663-018-0141-3
Atalay İ (2010) Applied Climatology. Meta Press, Izmir
Atalay İ, Efe R, Öztürk M (2014) Ecology and classification of forests in Türkiye. Procedia Soc Behav Sci 120:788–805. https://doi.org/10.1016/j.sbspro.2014.02.163
Atalay İ (1997) Türkiye Coğrafyası (Türkiye Geography in Turkish). Ege University, Bornova Izmir
Atalay İ (2002) Ecoregions of Türkiye. Ministry of Forestry Publication No: 163 (266), Ankara, Türkiye
Ataman O (1967) Dogu Karadeniz ladin ormanlarında Ips sexdentatus kabuk bocegi tahribatı ve mücadele esasları. Tek Bult Derg 6(22):79–89
Avcı M (1996) The floristic regions of Turkey and geographical approach for anatolian diagonal. Review of the Department of Geography, University of Istanbul, International Edition, vol 3. pp 59–91
Ayan S, Bugday E, Varol T, Özel HB, Thurm EA (2022) Effect of climate change on potential distribution of oriental beech (Fagus orientalis Lipsky.) in the twenty-first century in Türkiye. Theor Appl Climatol 148:165–177. https://doi.org/10.1007/s00704-022-03940-w
Bahadır M (2010) Türkiye’de (1998–2007) görülen orman yanginlarinin yüzey ve rakamsal sorgulama analizi. Nat Sci 5:146–162
Bahadır M, Emet K (2010) Türkiye’de Ana İklim Tiplerini Karakterize Eden Belli Başlı Ağaç Türlerinin Cbs İle Analizi. TÜBAV Bilim Dergisi 3:94–105
Baltacı H (2017) Meteorological analysis of flash floods in Artvin (NE Türkiye) on 24 August 2015. Nat Hazards Earth Syst Sci 17:1221–1230. https://doi.org/10.5194/nhess-17-1221-2017
Baskent EZ, Başkaya Ş, Terzioğlu S (2008) Developing and implementing participatory and ecosystem based multiple use forest management planning approach (ETÇAP): Yalnızçam case study. For Ecol Manage 256:798–807
Beguería S, Vicente-Serrano S M (2023) SPEI: calculation of the standardized precipitation-evapotranspiration index.
Bilgili (1997) Forests and forest fires in Türkiye. Accessed 21.04.2021 https://gfmc.online/iffn/country/tr/tr_2.html
Boydak M (2004) Silvicultural characteristics and natural regeneration of Pinus brutia Ten.—a review. Plant Ecol 171:153–163
Boydak M, Çalıkoğlu M (2008) Toros Sediri’nin (Cedrus libani A. Rich.) Biyolojisi ve Silvikültürü. Ormancılığı Geliştirme ve Orman Yangınları ile Mücadele Hizmetlerini Destekleme Vakfı Yayını, Lazer Ofset Matbaası. p 284
Camperlin P, Diop M (2003) Application of daily rainfall principal component analysis to the assessment of the rainy season characteristics in Senegal. Clim Res 23:159–169. https://doi.org/10.3354/CR023159
Chen Z, Wang W, Fu J (2020) Vegetation response to precipitation anomalies under different climatic and biogeographical conditions in China. Sci Rep 10:830. https://doi.org/10.1038/s41598-020-57910-1
Çolak AH, Rotherham ID (2006) A review of the forest vegetation of Turkey: its status past and present and its future conservation. Biology and Environment: Proceedings of the Royal Irish Academy 106B(3):343–354
Copernicus Climate Change Service (C3S) (2017): ERA5: Fifth generation of ECMWF atmospheric reanalyses of the global climate. Copernicus Climate Change Service Climate Data Store (CDS). Accessed 21.04.2021. https://cds.climate.copernicus.eu/cdsapp#!/home
Currit N, St Clair SB (2010) Assessing the impact of extreme climatic events on aspen defoliation using MODIS imagery. Geocarto International 25:2:133-147.https://doi.org/10.1080/10106040903051975
Danandeh Mehr A, Vaheddoost B (2020) Identification of the trends associated with the SPI and SPEI indices across Ankara. Türkiye Theor Appl Climatol 139:1531–1542. https://doi.org/10.1007/s00704-019-03071-9
Davis PH (1971) Distribution patterns in anatolia with particular reference to endemism plant life of South-West Asia. The Botanical Society of Edinburg, Edinburg, pp 15–28
de la Cruz AC, Gil PM, Fernández-Cancio Á, Minaya M, Navarro-Cerrillo RM, Sánchez-Salguero R, Grau JM (2014) Defoliation triggered by climate induced effects in Spanish ICP Forests monitoring plots. For Ecol Manag 331:245–255. https://doi.org/10.1016/j.foreco.2014.08.010
De Marco A, Proietti C, Cionni I, Fischer R, Screpanti A, Vitale M (2014) Future impacts of nitrogen deposition and climate change scenarios on forest crown defoliation. Environ Pollut 194:171–180. https://doi.org/10.1016/j.envpol.2014.07.02
Deng Y, Wang M, Yousefpour R, Hanewinkel M (2021) Abiotic disturbances affect forest short-term vegetation cover and phenology in Southwest China. Ecol Indic 124. https://doi.org/10.1016/j.ecolind.2021.107393
Development Plan (DP) (2018) 11th Development plan: forest and forestry products. Ministry of Development-Türkiye. https://www.sbb.gov.tr/wp-content/uploads/2020/04/Ormancilik_ve_Orman_UrunleriCalismaGrubuRaporu.pdf. Accessed 12.05.2022
Dönmez Y (1985) Bitki Coğrafyası. İstanbul Üniv. Coğ. Enst. Yay. No.3213, İstanbul
Efe R, Sönmez S (2006) The regional distribution of the forest vegetation according to the ecological and floristic properties in Turkey. IV. National Geography Symposium (Regional Differences in Turkey in the EU Process), 25-26 May 2006, Ankara (in Turkish)
Eichhorn J, Roskams P, Ferretti M, Mues V, Szepesi A, Durrant D (2010) Visual assessment of crown condition and damaging agents 49 p. manual part IV. In: Manual on methods and criteria for harmonized sampling, assessment, monitoring and analyses of the effects of air pollution on forests. UNECE ICP Forests Programme Coordinating Centre, Hamburg. ISBN: 978–3–926301–03–1. http://www.icp-forests.org/Manual.htm
Eichhorn J, Roskams P, Potočić N, Timmermann V, Ferretti M, Mues V, Szepesi A, Durrant D, Seletković I, Schröck H‐W, Nevalainen S, Bussotti F, Garcia P, Wulff S (2016) Part IV: visual assessment of crown condition and damaging agents. In: UNECE ICP Forests Programme Coordinating Centre (ed.): manual on methods and criteria for harmonized sampling, assessment, monitoring and analysis of the effects of air pollution on forests. Thünen Institute of Forest Ecosystems, Eberswalde, Germany, 49 p
Eris E, Cavus Y, Aksoy H, Burgan HI, Aksu H, Boyacioglu H (2020) Spatiotemporal analysis of meteorological drought over Kucuk Menderes River Basin in the Aegean Region of Türkiye. Theor Appl Climatol 142:1515–1530. https://doi.org/10.1007/s00704-020-03384-0
Erlat E, Türkeş M, Aydin-Kandemir F (2021) Observed changes and trends in heatwave characteristics in Türkiye since 1950. Theor Appl Climatol 145:137–157. https://doi.org/10.1007/s00704-021-03620-1
Ertugrul M, Varol T, Kaygin AT, Ozel HB (2017) The relationship between climate change and forest disturbance in Türkiye. Fres Environ Bull 26(6):4064–4074
Eşen D, Soysal GE, Yıldız O (2015) Regenerating Eastern Beech (Fagus orientalis Lipsky) With gaps of various sizes in the Western Black Sea Region of Turkey. Düzce Üniversitesi Orman Fakültesi Ormancılık Dergisi 11:71–82
Fady B, Semerci H, Vendramin GG (2003) EUFORGEN Technical Guidelines for genetic conservation and use forPinus halepensis and Pinus brutia. International Plant Genetic Resources Institute, Rome, 6 pages
Fernández-Manso A, Quintano C, Fernández-Manso O (2011) Forecast of NDVI in coniferous areas using temporal ARIMA analysis and climatic data at a regional scale. Int J Remote Sens 32(6):1595–617
Fischer R, Lorenz M, Mues V, Iost S, Granke O, Becher G, van Dobben H, Reinds GJ, de Vries W (2010) Forest Condition in Europe, Technical Report of ICP Forests 2010. ICP Forests, Hamburg, p 178
Fischer R, Waldner P, Carnicer J, Coll M, Dobbertin M, Ferretti M, Hansen K, Kindermann G, Lasch-Born P, Lorenz M, Marchetto A, Meining S, Nieminen T, Peñuelas J, Rautio P, Reyer C, Roskams P, Sánchez G (2012) The condition of forests in Europe: 2012 executive report. ICP Forests, Hamburg, Germany, pp. 24. URL: http://www.icp-forests.org/pdf/ER2012.pd
Frank A, Armenski T, Gocic M, Popov S, Popovic L, Trajkovic S (2017) Influence of mathematical and physical background of drought indices on their complementarity and drought recognition ability. Atmos Res 194:268–280. https://doi.org/10.1016/j.atmosres.2017.05.006
Gea-Izquierdo G, Ferriz M, Garcia-Garrido S, Aguin O, Elvira-Recuenco M, Hernandez-Escribano L, Martin-Benito D, Raposo R (2019) Synergistic abiotic and biotic stressors explain widespread decline of Pinus pinaster in a mixed forest. Sci Total Environ 685:963–975. https://doi.org/10.1016/j.scitotenv.2019.05.378
General Directorate of Forestry (GDF) (2009) Ormanlarımızda yayılış gösteren asli ağaç türleri (Primary tree species in Turkish forests). Republic of Türkiye General Directorate of Forestry.https://www.ogm.gov.tr/tr/e-kutuphane-sitesi/Yayinlar/Asli%20A%C4%9Fa%C3%A7%20T%C3%BCrleri.pdf Accessed 21 April 2022
General Directorate of Forestry (GDF) (2020) Forest assets in Türkiye. Republic of Türkiye General Directorate of Forestry. https://www.ogm.gov.tr/tr/ormanlarimiz/Turkiye-Orman-Varligi Accessed 21 April 2022
General Directorate of Forestry (GDF) (2022) Activity report. Republic of Türkiye General Directorate of Forestry. https://www.ogm.gov.tr/tr/faaliyet-raporu Accessed 21 April 2022
Gessner U, Naeimi V, Klein I, Kuenzer C, Klein D, Dech S (2013) The relationship between precipitation anomalies and satellite-derived vegetation activity in Central Asia. Glob Planet Chang 110:74–87. https://doi.org/10.1016/j.gloplacha.2012.09.007
Gottardini E, Cristofolini F, Cristofori A, Pollastrini M, Camin F, Ferretti M (2020) A multi-proxy approach reveals common and species-specific features associated with tree defoliation in broadleaved species For Eco. Manage 467(3):118–151. https://doi.org/10.1016/j.foreco.2020.118151
Günal N (2013) Türkiye’de iklimin doğal bitki örtüsü üzerindeki etkileri. Online Thematic J Turkic Studies 1:1–22
Gunes Y, Elvan OD (2005) Illegal logging activities in Türkiye. Environ Manage 36:220–229. https://doi.org/10.1007/s00267-003-0107-1
Hargreaves GH (1994) Defining and using reference evapotranspiration. J Irrig Drain Eng 120:1132–1139
Hartmann K, Krois J, Waske B (2018) E-Learning project SOGA: statistics and geospatial data analysis. Department of Earth Sciences, Freie Universitaet Berlin
Haynes KJ, Allstadt AJ, Klimetzek D (2014) Forest defoliator outbreaks under climate change: effects on the frequency and severity of outbreaks of five pine insect pests. Global Change Biol 20:2004–2018. https://doi.org/10.1111/gcb.12506
Lorenz M (2010) Objectives, Strategy and Implementation of ICP Forests. Manual Part I, 21 pp. Manual on methods and criteria for harmonized sampling, assessment, monitoring and analysis of the effects of air pollution on forests. UNECE, ICP Forests, Hamburg
Innes JL (1993) Forest health: its assessment and status. Commonwealth Agricultural Bureau, Wallingford
IPCC (2021) Summary for Policymakers. In: Masson-Delmotte, Zhai VP, Pirani A, Connors SL, Péan C, Berger S, Caud N, Chen Y, Goldfarb L, Gomis MI, Huang M, Leitzell K, Lonnoy E, Matthews JBR, Maycock TK, Waterfield T, Yelekçi O, Yu R, Zhou B (eds) Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. October 2021 by the IPCC, Switzerland
John R, Chen JQ, Ou-Yang ZT, Xiao JF, Becker R, Samanta A, Ganguly S, Yuan WP, Batkhishig O (2013) Vegetation response to extreme climate events on the Mongolian Plateau from 2000 to 2010. Environ Res Lett 8:0350333. https://doi.org/10.1088/1748-9326/8/3/035033
Kadioglu M (2000) Regional variability of seasonal precipitation over Türkiye. Int J Climatol 20(14):1743–1760. https://doi.org/10.1002/1097-0088(20001130)20:14%3c1743::AID-JOC584%3e3.0.CO;2-G
Kandemir G, Kaya Z (2009) EUFORGEN technical guidelines for genetic conservation and use of oriental beech (Fagus orientalis). Bioversity International, Rome, Italy. 6 pages
Karakas A, Zengin M, Sarigül M, Özay FS, Uluer K (2007) Bati karadeniz ve marmara bölgesinde hava kirliliginden (SO2) kaynaklanan orman zararlarinin belirlenmesi. kavak ve hizli gelisen orman agaclari arastirma enstitüsü, (In the Western Black Sea and Marmara region forest damage caused by air pollution (SO2) determination. Poplar and fast growing forest trees research institute), Technical Bulletin, No. 206, Izmit, Türkiye
Kayacık H (1980) Orman ve Park Ajraçlarinin Özel Sisternatijri 1. Gynospermae. I.Ü. Or. Fak. yayinlari No. 26421281. Bozak Matbaasi, Istanbul, p 388
Kendall MG (1968) A course of multivariate analysis. Hafner, New York
Koc G, Petrow T, Theiken AH (2020) Analysis of the most severe flood events in Türkiye (1960–2014): which triggering mechanisms and aggravating pathways can be identified?. Water 12:1562. https://doi.org/10.25932/publishup-47733
Kömüsçü AM, Erkan A, Çelik S (1998) Analysis of meteorological and terrain features leading to the Izmir flash flood, 3–4 November 1995. Nat Hazards 18(1):1–25
Koray ES, Güner ST, Cömez A, Çelik N, KaratasR, Karakas A (2007) Eskisehir, sakarya, bilecik,bolu yörelerinde hava kirliliğinin ormanlar üzerine etkisinin belirlenmesi (Determination of the effect of air pollution on forests in Eskisehir, Sakarya, Bilecik and Bolu regions). Çevre ve Orman Bakanlığı (Ministry of Environment and Forest). ESK-02.8302/2002–2007
Kottek M, Grieser J, Beck C, Rudolf B, Rubel F (2006) World map of Köppen-Geiger climate classification. Meteorologishe Zeitschrift 15(3):259–263
Kovalev AV, Voronin VI, Oskolkov VA, Sukhovolskiy VG (2021) Analysis of forest condition based on MODIS remote-sensing data. Contemp. Probl Ecol https://doi.org/10.1134/S199542552107009X
Li P, Zhu D, Wang Y, Liu D (2020) Elevation dependence of drought legacy effects on vegetation greenness over the Tibetan Plateau. Agric for Meteorol 15:295
Limaki MK, Nimvari ME, Alavi SJ, Mataji A, Kazemnezhad F (2021) Potential elevation shift of oriental beech (Fagus orientalis L.) in Hyrcanian mixed forest ecoregion under future global warming. Ecol Modell 455:1–13. https://doi.org/10.1016/j.ecolmodel.2021.109637
López-Tirado J, Vessella F, Stephan J, Ayan S, Schirone B, Hidalgo P (2021) Effect of climate change on potential distribution of Cedrus libani A Rich in the Twenty-First centuury: an Ecological Niche Modeling assessment. New Forests. https://doi.org/10.1007/s11056-020-09798-y
Lorenz M (1995) International co-operative programme on assessment and monitoring of air pollution effects on forests-ICP forests. Water Air Soil Pollut 85:1221–1226. https://doi.org/10.1007/BF00477148
Mátyás C, Ackzell L, Samuel CJA (2004) EUFORGEN technical guidelines for genetic conservation and use for Scots pine (Pinus sylvestris). International Plant Genetic Resources Institute, Rome, Italy. 6 pages
McKee TB, Doesken NJ, Kleist J (1993) The relationship of drought frequency and duration to time scales. In Proceedings of the 8th Conference on Applied Climatology. 17:22 pp. 179–183
Messinger J, Güney A, Zimmermann R, Ganser B, Bachmann M, Remmele S, Aas G (2015) Cedrus libani: a promising tree species for Central European forestry facing climate change? Eur J for Res. https://doi.org/10.1007/s10342-015-0905-z
Michel A, Prescher A-K, Schwärzel K, editors (2020) Forest condition in Europe: the 2020 assessment. ICP forests technical report under the UNECE convention on long-range transboundary air pollution (air convention) Eberswalde: Thünen Institute. https://doi.org/10.3220/ICPTR1606916913000
Müller-Edzards C, Vries WD, Erisman JW (1997) Ten years of monitoring forest conditions in Europe. Studies on temporal development, spatial distribution and impacts of natural and anthropogenic stress factors. UN/ECE
Özcan O, Musaoğlu N, Türkeş M (2018) Assessing vulnerability of a forest ecosystem to climate change and variability in the western Mediterranean sub-region of Türkiye. J for Res 29:709–725. https://doi.org/10.1007/s11676-017-0505-5
Özçelik R, Brooks JR (2012) Compatible volume and taper models for economically important tree species of Türkiye. Ann for Sci 69:105–118. https://doi.org/10.1007/s13595-011-0137-4
Özer YT (2014) Pattern of genetic diversity in Turkey oak (quercus cerris l.) populations, Ph.D. - Doctoral Program. Middle East Technical University
Ozturk M, Gucel S, Kucuk M, Sakcali S (2010) Forest diversity, climate change and forest fires in the Mediterranean region of Türkiye. J Environ Biol 31(1–2):1–9
Ozturk T, Ceber ZP, Türkeş M, Kurnaz ML (2015) Projections of climate change in the Mediterranean Basin by using downscaled global climate model outputs. Int J Climatol 35:4276–4292
Pandžić K, Trninić D, Likso T, Bošnjak T (2009) Long-term variations in water balance components for Croatia. Theor Appl Climatol 95:39–51. https://doi.org/10.1007/s00704-007-0366-5
Pasquarella VJ, Mickley JG, Barker Plotkin A, MacLean RG, Anderson RM, Brown LM, Wagner DL, Singer MS, Bagchi R (2021) Predicting defoliator abundance and defoliation measurements using Landsat‐based condition scores. Remote Sensing in Ecology and Conservation
Petrakis PV, Ioannidis C, Zygomala AM (2007) Biotechnology of Pinus brutia and Pinus halepensis as important landscape plants of the East Mediterranean. Tree For Sci Biotech 1(1):26–38
Polong F, Chen H, Sun S, Ongoma V (2019) Temporal and spatial evolution of the standard precipitation evapotranspiration index (SPEI) in the Tana River basin, Kenya. Theor Appl Climatol 138:777–792. https://doi.org/10.1007/s00704-019-02858-0
Popa I, Badea O, Silaghi D (2017) Influence of climate on tree health evaluated by defoliation in the ICP level I network (Romania) iForest 10:554–560. https://doi.org/10.3832/ifor2202-009
Potočić N, Timmermann V, Ognjenović M, Kirchner T, Prescher A-K, Ferretti M (2021) Tree health is deteriorating in the European forests (ICP Forests Brief No. 5) Programme Co-ordinating Centre of ICP Forests, Thünen Institute of Forest Ecosystems. https://doi.org/10.3220/ICP1638780772000
Pravalie R, Sîrodoev I, Peptenatu D (2014) Detecting climate change effects on forest ecosystems in Southwestern Romania using Landsat TM NDVI data. J Geogr Sci 24:815–832. https://doi.org/10.1007/s11442-014-1122-2
Quiñonez-Piñón R, Mendoza-Durán A, Valeo C (2007) Design of an environmental monitoring program using NDVI and cumulative effects assessment. Int J Remote Sens 28(7):1643–1664. https://doi.org/10.1080/01431160600887730
Rodgers JL, Nicewander WA (1988) Thirteen ways to look at the correlation coefficient. Am Stat 42(1):59–66. https://doi.org/10.1080/00031305.1988.10475524
Salvador M, de Brito JIB (2018) Trend of annual temperature and frequency of extreme events in the MATOPIBA region of Brazil. Theor Appl Climatol 133(1–2):253–261. https://doi.org/10.1007/s00704-017-2179-5
Sanchez-Salguero R, Camarero JJ, Grau JM, Cruz AC, Gil PM, Minaya M, Fernandez-Cancio A (2017) Analysing atmospheric process and climatic derivers of defoliation to determine forest vulnerability to climate warming. Forest 8:1–13. https://doi.org/10.3390/f8010013
Schmidt M, Klein D, Conrad C, Dech S, Paeth H (2014) On the relationship between vegetation and climate in tropical and northern Africa. Theor Appl Climatol 115:341–353. https://doi.org/10.1007/s00704-013-0900-6
Seidl R, Thom D, Kautz M et al (2017) Forest disturbances under climate change. Nat Clim Change 7:395–402. https://doi.org/10.1038/nclimate3303
Semerci A, Semerci H, Çalışkan B, Çiçek N, Ekmekçi Y, Mencuccini M (2017) Morphological and physiological responses to drought stress of European provenances of Scots pine. Eur J Forest Res 136:91–104. https://doi.org/10.1007/s10342-016-1011-6
Seneviratne S, Nicholls N, Easterling D, Goodess C, Kanae S, Kossin J, Luo Y, Marengo J, McInnes K, Rahimi M, Reichstein M, Sorteberg A, Vera C, Zhang X (2012) Changes in climate extremes and their impacts on the natural physical environment, chapter 3 in IPCC Special Report on Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation. Cambridge University, Cambridge
Şevik H, Ertürk E (2015) Effects of drought stress on germination in fourteen provenances of Pinus brutia Ten. seeds in Türkiye. Turk J Agric Food Sci Tech 3:294–299
Shawul AA, Chakma S (2020) Trend of extreme precipitation indices and analysis of long-term climate in the Upper Awash basin, Ethiopia. Theor Appl Climatol. https://doi.org/10.1007/s00704-020-03112-8
Simeone MC, Zhelev P, Kandemir G (2019) EUFORGEN technical guidelines for genetic conservation and use of Türkiye oak (Quercus cerris), European Forest Genetic Resources Programme (EUFORGEN), European Forest Institute. 6 pages
Sirdaş S, Sen Z (2003) Spatio-temporal drought analysis in the Trakya region. Türkiye Hydrol Sci J 48(5):809–820. https://doi.org/10.1623/hysj.48.5.809.51458
Sneyers R, Vandiepenbeek M, Vanlierde R (1989) Principal component analysis of belgian rainfall. Theor Appl Climatol 39:199–204
Song XP, Hansen MC, Stehman SV, Potapov PV, Tyukavin A, Fermote EF, Townshend JR (2018) Global land change from 1982 to 2016. Nature 560:639–643. https://doi.org/10.1038/s41586-018-0411-9
Stagge JH, Tallaksen LM, Gudmundsson L, Van Loon AF, Stahl K (2015) Candidate distributions for climatological drought indices (SPI and SPEI) abstract. Int J Climatol 35(13):4027–4040. https://doi.org/10.1002/joc.4267
Tadić L, Bonacci O, Brleković T (2019) An example of principal component analysis application on climate change assessment. Theor Appl Climatol 138:1049–1062. https://doi.org/10.1007/s00704-019-02887-9
Tayanç M, Toros H (1997) Urbanization effects on regional climate change in the case of four large cities of Türkiye. Clim Change 35:501–524. https://doi.org/10.1023/A:1005357915441
Tolunay A, Akyol A, Özcan M (2008) Usage of trees and forest resources at household level: a case study of Açagl Yumrutaç Village from the West Mediterranean Region of Türkiye. Res J Forest 2(1):1–14
Tolunay D, Karabıyık B, Temerit A (2010) The result of a nation-wide systematic forest condition survey in Türkiye. IFOREST 4:145–149
Toros H (2012a) Spatio-temporal variation of daily extreme temperatures over Türkiye. Int J Climatol 32(7):1047–1055. https://doi.org/10.1002/joc.2325
Toros H (2012b) Spatio-temporal precipitation change assessments over Türkiye. Int J Climatol 32(9):1310–1325. https://doi.org/10.1002/joc.2353
Townsend PA, Singh A, Foster JR, Rehberg NJ, Kingdon CC, Eshleman KN, Seagle SW (2012) A general Landsat model to predict canopy defoliation in broadleaf deciduous forests. Remote Sens Environ 16(119):255–265
TSMS (2022) Precipitation assessment for 2021. Turkish State Meteorology Service, Ministry of Environment, Urbanization and Climate Change.
Tüfekçioğlu A, Güner S, Tilki F (2011) Climate change and oriental spruce (Picea orientalis) ecosystems in Eastern Black Sea region of Türkiye. Artvin Çoruh Üniversitesi- Orman Fakültesi Dergisi 9(1):101–106
Türkeş M (2012) Türkiye’de gözlenen ve öngörülen iklim değişikliği, kuraklık ve çölleşme. Ankara Üniversitesi Çevrebilimleri Dergisi 4(2):1–32
Türkeş M, Sümer UM, Kılıç G (1995) Variations and trends in annual mean air temperatures in Türkiye with respect to climatic variability. Int J Climatol 15:557–569. https://doi.org/10.1002/joc.3370150507
Türkeş M, Koç T, Sariş F (2009) Spatiotemporal variability of precipitation total series over Türkiye. Int J Climatol 29(8):1056–1074. https://doi.org/10.1002/joc.1768
Türkeş M, Musaoğlu N, Özcan O (2018) Assessing the vulnerability of a forest ecosystem to climate change and variability in the western Mediterranean sub-region of Türkiye: future evaluation. J for Res 29:1177–1186. https://doi.org/10.1007/s11676-017-0500-x
Ülgentürk S, Şahin Ö, Ayhan B, Sarıbaşak H, Kaydan MB (2012) Coccoidea (Hemiptera) species of Taurus cedar (Cedrus libani) in Türkiye. Turkish J Entomol 36(1):113–122
Unal YS, Tan E, Mentes SS (2013) Summer heat waves over western Türkiye between 1965 and 2006. Theor Appl Climatol 112:339–350. https://doi.org/10.1007/s00704-012-0704-0
Usta A, Yılmaz M, Kocamanoğlu YO, Esengül G (2018) Impact of spatial factors on climate variables and species distribution in forest ecosystems under sea influence of Eastern Blacksea Region. Eurasian J for Sci 6(3):83–97
Vicente-Serrano SM, Beguería S, López-Moreno JI (2010) A multiscalar drought index sensitive to global warming: the standardized precipitation evapotranspiration index. J Clim 23(7):1696–1718. https://doi.org/10.1175/2009JCLI2909.1
Wu X, Liu H, Li X, Ciais P, Babst F, Guo W, Zhang C, Magliulo V, Pavelka M, Liu S, Huang Y (2018) Differentiating drought legacy effects on vegetation growth over the temperate Northern Hemisphere. Glob Change Biol 24(1):504–516
Yüksel B, Akbulut S (2005) Doğu ladini ormanlarında Ips sexdeııtatus (Boern.)'un doğal düşmanlarının belirlenmesi. J Faculty Forestry Istanbul Univ 55(2):59–70
Zhang Y, Peng CH, Li WZ, Tian LX, Zhu QA, Chen H, Fang XQ, Zhang GL, Liu GB, Mu XM, Li ZB, Li SQ, Yang YZ, Wang J, Xiao XM (2016) Multiple afforestation programs accelerate the greenness in the‘three north’ region of China from 1982 to 2013. Ecol Indic 61:404–412. https://doi.org/10.1016/j.ecolind.2015.09.041
Zhu C, Zhang X, Zhang N, Hassan MA, Zhao L (2018) Assessing the defoliation of pine forests in a long time-series and spatiotemporal prediction of the defoliation using Landsat data. Remote Sens. https://doi.org/10.3390/rs1003036
Zierl B (2004) A simulation study to analyse the relations between crown condition and drought in Switzerland. For Ecol Manag 188:25–38. https://doi.org/10.1016/j.foreco.2003.07.019
Zohary M (1971) The phytogeographical foundations of the Middle East plant life of South-West Asia. The Botanicalm Society of Edinburg, Edinburg, pp 43–53
Acknowledgements
The authors would like to thank the Ministry of Agriculture and Forestry for permission to use the defoliation data for the plots. Special thanks to Sıtkı Öztürk for his expert assistance.
Funding
Open access funding provided by the Scientific and Technological Research Council of Türkiye (TÜBİTAK). This work is funded by The Scientific and Technological Research Council Of Türkiye (TUBITAK) with the Grant Number 1059B142000733. University of Freiburg provided additional resources during a research exchange of NDC.
Author information
Authors and Affiliations
Contributions
Author contribution All four authors were involved in the conception of the idea. Data collection was performed by NDC and ADŞ. The first analysis was performed by NDC and reviewed by AC. The first draft of the manuscript was written by NDC and AC. The results were reviewed and analyzed by NDC under the supervision of AC and RY. All authors were involved in the development of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Dogan Ciftci, N., Şahin, A.D., Yousefpour, R. et al. Effects of climate trends and variability on tree health responses in the Black Sea and Mediterranean forests of Türkiye. Theor Appl Climatol 155, 3969–3991 (2024). https://doi.org/10.1007/s00704-024-04853-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00704-024-04853-6