
Abstract
A temperature increase higher than the global mean is likely for Central Europe until the end of the century. Historic climate records reveal that the temperature in eastern Austria has already increased by about 2 °C over the last 50 years. We investigated the responses of ecophysiological and productivity indicators to climate change of the Vienna Woods, the neighbouring forests of the Austrian capital, to assess potential impacts on forest functioning. In this impact analysis, we used the biogeochemical mechanistic model Biome-BGC and ran it with 50 years of historic climate data and regional climate change projections based on IPCC emission scenarios A1B and B1 until 2100. We projected sustained productivity until the end of the twenty-first century. Lowered soil water potentials, however, seem to limit a productivity increase, especially in the low elevation areas, while the canopy leaf area, annual soil water outflow and annual mean water use efficiency are projected to remain constant or increase. We conclude that the forests in the greenbelt of Vienna are not severely prone to negative functional climate change effects, and therefore, key forest functions concerning welfare and recreation can be maintained in the Vienna Woods.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The role of forests in and around densely populated areas is of increasing interest since they cover a broad range of environmental, social and economic issues (Konijnendijk et al. 2005; Carreiro et al. 2008; Konijnendijk 2008; Young 2010). Functioning forests secure the traditional use of forests for timber production as well as the provision of a variety of important non-timber forest products and services, such as welfare and recreation (Oleyar et al. 2008; Dobbs et al. 2011). Climate change impact studies deal with potential latitudinal (Woodall et al. 2010) and altitudinal shifts in tree species (Peñuelas et al. 2007), increasing biotic and abiotic disturbances (Dukes et al. 2009; Seidl et al. 2014), socioeconomic impacts (e.g. creation of new, ‘green’ jobs, Renner et al. 2008), climate change adaptation (Lindner et al. 2010) and mitigation options (Millar et al. 2007; Zheng et al. 2013) and climate change impact on ecophysiology and productivity (Hyvönen et al. 2006; Luo et al. 2008). Latter will impact forest ecosystem functioning with possible consequences on welfare and recreation, for example, due to the stabilising effects of forests for the urban microclimate (Peters and McFadden 2010) or role of forests in the water supply (Furniss et al. 2010).
In the present study, we are interested in the climate change impact on ecophysiology- and productivity-based key forest functions of the famous ‘Wienerwald’ or the ‘Vienna Woods’ that borders the Austrian capital. This European beech (Fagus sylvatica L.) dominated forest is located in the transition zone between the eastern edge of the Alps and the Pannonic lowlands and includes a UNESCO biosphere reserve. The forest provides important ecosystem services to Vienna’s urban population, such as recreation, welfare, fresh air, climate, fresh water, protection against soil erosion and timber. The forest is explicitly managed to attain these goals (Foet 2010; Albrecht 2011).
European beech is one of the most dominant tree species in Europe with a wide distribution in Central and Western Europe. In the last decades, beech forests have experienced increased growth rates in Central Europe, mostly attributed to increased nitrogen deposition, higher temperatures and CO2 levels (Pretzsch et al. 2014). European beech avoids regions with long, very cold winters or long dry periods in summer and is often said to require a minimum annual precipitation of 600 mm (Forstreuter 2002), although the actual minimum will depend on the distribution of precipitation among the seasons and on the soil conditions (Ellenberg and Leuschner 2010). Due to the species’ sensitivity to water supply (Granier et al. 2007), the future of beech at a southern or lower elevation distribution limit is uncertain under expected climate change (Mátyás et al. 2010; Hlásny et al. 2011; Tegel et al. 2013).
For this study, we used biogeochemical mechanistic modelling to assess potential climate change-driven impacts on forest functioning (not including storm or biotic disturbances) using the process-based ecosystem model Biome-BGC as a diagnostic tool. We employed 50 years of local historic climate data, four regional climate change scenarios for the horizon 2100 and corresponding atmospheric CO2 concentrations to drive the model. The scenarios are based on the IPCC emission scenarios A1B and B1, and both are used with and without an additional precipitation scenario. We were interested in (1) investigating changes of ecophysiology and productivity measures and (2) deriving indications of changes in forest functions relevant for welfare and recreation within the Vienna Woods.
Materials and methods
Study area
The study region covers the Vienna Woods, the largest broadleaf forest in Austria which lies south-west of the capital city of Vienna. The area is located at the eastern edge of the Limestone Alps and comprises non-forested areas such as agricultural land and some smaller settlements. The total study region area is about 140,000 ha within the provinces of Lower Austria and Vienna. A major part of the area (105,000 ha) is a UNESCO biosphere reserve. Elevation ranges from 140 to 1,000 m. Annual mean temperature and annual mean precipitation (mean 1960–2009) range from 5.9 to 10.6 °C and from 550 to 1,290 mm, respectively. Current nitrogen deposition rates are between 7 and 26 kg N ha−1 year−1 (Eastaugh et al. 2011).
The Vienna Woods cover highly productive broadleaved European beech (F. sylvatica L.) dominated forests that face strong recreational demands by the urban population. For many centuries, the Vienna Woods were royal hunting grounds. Wood production has become increasingly important over the last 400 years. Plans of the later nineteenth century to clear the forest and build housing were dismissed due to broad public resistance. Today, the forests are managed by the Vienna Forest Administration and the Austrian Federal Forests. Although timber production is still important, the forests are explicitly managed to address societal demands. For instance, approximately 95 % of the forest area is used for recreational purposes with around 500,000 visitors annually.
According to the European forest types (Larsson 2001), the predominant beech forests can be categorised as 5a ‘lowland and submontane beech forest’, where F. sylvatica is the dominating tree species due to its high competitiveness and often forms monospecific and monolayered forests. The lower elevations also feature oak–hornbeam (Quercus robur L., Carpinus betulus L.) forests, and a less dense downy oak (Quercus pubescens Willd.) forest can be found in the drier parts of the area on calcareous bedrock (Landsteiner 1990).
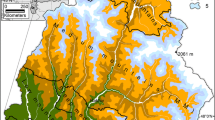
Figure 1 gives an overview of the forest cover in the study region with its dominating forest types (broadleaved, mixed, etc.). It also shows the 782 grid points (1 km × 1 km) within the forest area, where the analyses were performed. Due to the predominance of beech, we treated the whole forest area as a beech forest. Climate change assessments, ecophysiology considerations and analysis of potential productivity responses are done for the forested part only. For more detailed regional analysis, we selected grid points from three sub-regions in the north-east (NE), the south-east (SE) and west (W, Fig. 1). Each sub-region is characterised by a strong elevation gradient. The NE plots comprise all simulation plots within the city of Vienna boundaries. Information on the forest cover was obtained from a GIS raster layer provided by the Federal Research and Training Centre for Forests, Natural Hazards and Landscape (BFW). The GIS layer is based on data of the Austrian National Forest Inventory of the period 2000–2002 and LANDSAT data from 2000 to 2003 (Koukal 2004).

Forest area with the main forest categories in the study area; the 782 forest simulation points on a 1 km × 1 km grid, and three sub-regions with simulation points in the north-east (NE), south-east (SE) and west (W)
Climate data
Daily weather data are key ecosystem model drivers. The climate data used for this study consisted of daily weather data using (1) historic records from the year 1960 to 2009 and (2) climate change scenarios for the years 2010–2100.
Historic climate data
The historic daily minimum and maximum temperature (T min, T max) and precipitation (Prcp) data came from climate stations provided by the Austrian Central Institute for Meteorology and Geodynamics (ZAMG). To produce daily weather data on a 1 km × 1 km grid, we applied DAYMET (Thornton et al. 1997). DAYMET is a climate interpolation and simulation tool adapted and validated for Austria (Thornton et al. 2000; Petritsch 2002; Hasenauer et al. 2003). For the period of 1960–2009, daily T min, T max and Prcp were interpolated from 25 and 15 stations, respectively, and solar radiation (Srad) and vapour pressure deficit (VPD) were calculated based on the temperature and precipitation for each forest grid point. A digital elevation map (100 m × 100 m resolution) was required to address elevation and aspect-related variations in the interpolation procedure. For details on the algorithms, we refer to Petritsch and Hasenauer (2009).
A linear regression analysis of the interpolated grid point data revealed significant trends in the historic temperature for the period 1960–2009 (Table 1, results section). The average 50-year increase in temperature derived from the trends was 2.6 °C for T min and 1.4 °C for T max. The Prcp and VPD trends were not significant (Table 1, results section). Figure 2 shows for the period of 1960–2009 the regional distribution of the mean of daily average temperature [(T min + T max) × 0.5], and the annual precipitation as well as the trends in average temperature and precipitation.

Mean average temperature (top left), annual average temperature change (top right), mean annual precipitation (bottom left) and annual precipitation change (bottom right) between 1960 and 2009. The data were calculated using DAYMET 100 m × 100 m grid interpolation. Average temperature calculated as the mean of daily minimum and maximum temperature. Trend calculated as the slope of the linear regression line through annual average values (temperature) or annual sums (precipitation)
Climate change scenarios
The regional climate change scenarios were obtained from the regional climate model (RCM) climate limited-area model (CLM) v2.4.11 (Hollweg et al. 2008). CLM projections for Germany lie within the range of other regional climate change projections (Jacob et al. 2012; Deutscher Wetterdienst 2014). The experiments CLM_A1B_1_D3 and CLM_B1_1_D3 (Lautenschlager et al. 2009a, b) are European climate simulations on a 0.2° horizontal grid resolution (grid cell size ~15 km × 22 km) for the years 2001–2100. They used global greenhouse gas emission scenarios A1B and B1 (IPCC 2000). The atmospheric CO2 concentration in the A1B scenario is projected to reach 718 ppm, and 548 ppm in the B1 scenario by the end of the twenty-first century. The IPCC A1B scenario assumes a rapid economic growth, the global population peaking mid-century and an introduction of new and more efficient technologies affecting all energy sources. The B1 scenario addresses a high level of environmental and social consciousness, a fast change in economic structure towards a service and information economy combined with a coherent approach to sustainable development (low material, clean and resource-efficient technologies), but without additional climate initiatives. The global population also peaks mid-century and declines thereafter.
Daily T min, T max, Prcp and Srad data for the scenario A1B and B1 were obtained. They were not directly suitable as input for our ecosystem modelling because the RCM does not provide temperature and precipitation for single locations (the 782 forest grid points). Moreover, when switching from the observed daily weather data to the RCM scenario data, typically problematic discontinuities in the climate data occur. This phenomenon is especially pronounced in mountainous regions where discrepancies may be several degrees Celsius and more than 100 % in precipitation (Formayer et al. 2010).
For each day of the year, we calculated trend lines for T min, T max and Prcp by scenario to derive absolute changes between the years 2010 and 2100. The difference among the scenario grid cells turned out to be small; thus, we continued to use averaged values. Figure 3 gives the absolute T min and T max change for the scenario A1B and B1 for each day of the year. A polynomial regression was fit across all 365 daily changes. We obtained our 2010–2100 temperature scenarios time series for the 782 grid points by randomly repeating the meteorological years of the observation period 1960–2009 and then linearly increasing these T min and T max data with the smoothed daily temperature trends of the scenarios (Fig. 3). For Prcp and Srad, similar approaches were attempted. However, the changes yielded were negligible (not shown). Instead, we assumed Prcp to increase in winter and decrease in summer, its maximum at 30 % (Fig. 2). For Srad, we did not introduce a scenario because the projected changes were minor (not shown), but used the unchanged Srad values. VPD was calculated from the temperature scenarios using the DAYMET algorithm. All daily meteorological variables were always based on a common day from the observation period to ensure physical congruency. This resulted in four different climate change scenarios for the period 2010–2100:

Projected T min and T max change between 2010 and 2100 for each day of the year (lines polynomial regression) according to the CLM A1B (top) and B1 simulations (middle) and the sinusoidal scenario of relative precipitation change between 2010 and 2100 for each day of the year (bottom). The actual pattern of rainfall and non-rainfall days is retained
-
1.
A1B and no change in precipitation (A1B).
-
2.
A1B and precipitation changes as outlined above (A1B + Prcp).
-
3.
B1 and no change in precipitation (B1).
-
4.
B1 and precipitation changes as outlined above (B1 + Prcp).
The linear trends of annual T min and T max, averaged for the 782 forest grid points for the period 2010–2100, gave a 100-year increase in T min of 4.2 °C and T max of 4.5 °C for A1B, and 2.3 °C (T min) and 2.6 °C (T max) for B1 (Table 1, results section). In Table 2 (results section), we provide the periodical mean values for the 20-year period 1960–1979, 1990–2009 and 2081–2100. Linear trends of Prcp for 2010–2100 were not significant, and VPD increased significantly in both A1B and B1 (Table 1, results section). The residuals of the linear regression model for T min, T max, Prcp and VPD were analysed for temporal autocorrelation (using a Durbin-Watson test). No indication of our data not meeting the assumption of the regression analysis was found (DW > 1.9).
The annual average temperature development from 1960 to 2009 and from 2010 to 2100 for the A1B and B1 scenarios and the annual precipitation from 1960 to 2009 and from 2010 to 2100 with and without our precipitation scenario were averaged for all 782 forest grid points to show annual fluctuations and temporal developments (Fig. 4).

Annual average temperature (top) and annual precipitation (bottom) averaged over the 782 forest grid points between 1960 and 2100, including linear trend lines. Values of the climate observation period 1960–2009 are interpolated with DAYMET. For temperature in the period 2010–2100, the meteorological years of the period 1960–2009 are randomly repeated and temperature is linearly increased with the A1B or B1 scenario temperature trends. For precipitation in the period 2010–2100, the meteorological years of the period 1960–2009 are randomly repeated or in addition rescaled with the sinusoidal precipitation scenario
The ecosystem model
For our study, we used the ecosystem model Biome-BGC (Thornton 1998) version 4.1.1 (Thornton et al. 2002), with improvements related to species representation and the self-initialisation process (Pietsch et al. 2005; Pietsch and Hasenauer 2006). The model simulates energy, water, carbon and nitrogen cycles within a given ecosystem on a daily time step. The model is driven by the daily meteorological data, i.e. daily minimum and maximum temperature, incident solar radiation, vapour pressure deficit and precipitation. Vegetation-specific properties of the modelled ecosystem are listed in an ecophysiological parameter set (White et al. 2000; Pietsch et al. 2005). Atmospheric CO2 content, nitrogen deposition and fixation, aspect, elevation and physical soil properties (depth, texture) are input parameters for the model. The model predicts stomatal conductance to CO2 and water vapour by reducing species-specific maximum stomatal conductance by a series of multiplicative reductions based on solar radiation, VPD, soil water potential (Psi, Ψ) and daily minimum temperature (Thornton et al. 2002). The multiplicators take values from 1 (no limitation) to 0 (total stomatal closure). Threshold values of VPD [600, 3,000 Pa], Ψ [−0.34, −1.70 MPa] and T min [0, −8 °C] are species specific (Pietsch et al. 2005, values for beech in brackets). Photosynthesis is calculated separately for the sunlit and shaded canopy fractions, based on the Farquhar photosynthesis routine (Farquhar et al. 1980). Growth, the allocation of newly assimilated carbon to the different plant compartments and to the storage pools for following years growth, is limited by the carbon requirements for autotrophic respiration (i.e. for maintenance respiration, what needs to be fulfilled first, and for growth respiration) and by the availability of nitrogen to fulfil the species-specific C/N ration requirements of the different plant compartments. Maintenance respiration is a temperature-dependent function of the nitrogen content of the plant compartment (Ryan 1991), and growth respiration is a function of the carbon allocated to the different plant compartments (Larcher 1995). Both leaves flushing in spring and litter-fall in autumn are soil temperature dependent. Decomposition of dead organic material is regulated by temperature and soil water status. Easily decomposable labile and more recalcitrant cellulose and lignin plant proportions go in different decomposition pathways. Leaching and volatile loss of and competition for soil mineral nitrogen between growing plants and decomposing microorganisms are considered in the model. The water cycle considers canopy interception and stomatal conductance/transpiration, evaporation from the canopy and from the soil, snow sublimation, storage of water in the soil and outflow (Of) of water exceeding saturation and field capacity. The soil water-holding capacity at saturation is derived from soil depth and texture, based on empirical pedotransfer functions (Clapp and Hornberger 1978; Cosby et al. 1984; Saxton et al. 1986). Evaporation and transpiration are estimated using the Penman–Monteith equation and depend on air temperature, air pressure, VPD, solar radiation and the transport resistance of water vapour and sensible heat; soil evaporation also depends on the number of days since the last rain event and transpiration on the stomatal, cuticular and boundary layer conductance. For further detail, we refer to Thornton (1998), White et al. (2000), Thornton et al. (2002), Pietsch and Hasenauer (2006) and Pietsch and Hasenauer (2009). For this study, phenology required an adaptation in order for it to account for altered phenology through increasing temperatures. Based on the original algorithm, we calculated the required long-term daily average temperature mean (T avg, mean) from the available climate data (1960–2100) in a moving window that has the same length as the historic climate records (50 years).
Modelling procedure
The general modelling procedure within Biome-BGC uses a self-initialisation procedure including the dynamic mortality routine of Pietsch and Hasenauer (2006) to stabilise the pool sizes at a dynamic equilibrium (negligible changes i.e. in soil carbon stocks between two mortality cycles). Historic land use, i.e. forest management, was considered by a sequence of clear cuts and plantings (3–4) and thinnings for management-related impacts on the state and productivity of the forest system (Pietsch and Hasenauer 2002). In this study, the rotation length was set to 130 years. Viennese forests experienced intensive management which was addressed by employing the thinning routine for beech as described in Petritsch (2008). For the scheduling of the management interventions, we randomly assigned a current age (in 2009) between 1 and 130 years to the simulation plots. The implemented forest management routine allows that during clear-cut and thinning intervention user-specified total amounts or shares of carbon and nitrogen are transferred from the living biomass pools (stem including branches, fine and coarse roots, leaves) to the dead biomass pools (coarse woody debris, litter), but carbon and nitrogen can also be extracted from the system.
The available daily climate data from 1960 to 2009 are repeatedly and randomly used for the simulations before 1960 (self-initialisation, historic land use). From 1960 to 2009, the observed climate data were used in the natural order, which is followed by the different climate change scenarios (A1B, A1B + Prcp, B1, B1 + Prcp). For the early phases of simulations, the CO2 concentration was kept at a preindustrial level of 278 ppm. The historic anthropogenic CO2 increase followed IPCC’s mean global annual atmospheric CO2 concentration data set IS92a starting in 1765 (IPCC 1992; Enting et al. 1994). For the climate change simulations, the CO2 concentrations were used as prescribed by the emission scenarios A1B and B1. Preindustrial nitrogen deposition was estimated by Holland et al. (1999) at 1 kg N ha−1 year−1, whereas current annual nitrogen deposition was taken from a nitrogen deposition map combining nitrogen deposited in wet and dry form, published by Eastaugh et al. (2011). The historic development of the nitrogen deposition was approximated by the trend in the atmospheric CO2 concentration. For future simulations, the nitrogen deposition rates were kept constant at current levels. Soil texture and depth were interpolated from the Austrian National Forest Soil Survey (Petritsch and Hasenauer 2007).
Model validation
We obtained stand (age, volume) and site (elevation, slope, aspect) and management information for 32 experimental beech plots from the region which were established and are maintained by the Institute of Silviculture at the University of Natural Resources and Life Sciences, Vienna. Of these plots, ten are directly located within the study region. The remaining plots are found in the south within 65 km of the study area, and one plot is 120 km away. The experimental plots cover a wide age range (>150 years), have a documented management history and serve as model validation plots.
We applied the same modelling procedure as described above to the 32 experimental beech plots. The scheduling of the management, however, was adapted to the documented management history and the age of the plots.
Ecophysiological indicators
Daily weather, atmospheric CO2 concentrations and nitrogen deposition rates affect the ecophysiological processes of forests either directly or through feedback between pools and fluxes. Certain ecophysiological indicators possess a strong ability to highlight potential reactions to changes in climate because they are decisive in the functioning and the productivity of forests. For our study, such model variables are the transpiration (Tr), the evaporation (Ev), the water use efficiency [WUE, ratio between net primary production (NPP) and transpiration], the soil water potential (Psi,Ψ), outflow of water through deep percolation (Of) and the net soil nitrogen mineralisation rate (Nmin). Psi is used by Biome-BGC as a surrogate for the leaf water potential. It can be taken as a measure of the system dryness. Furthermore, it influences the transpiration, evaporation, CO2 uptake through the stomata and decomposition. Nmin is mediated by soil temperature and Psi and is important for sustained ecosystem productivity.
Ecosystem productivity indicators
Productivity indicators, such as leaf area index (LAI), gross primary production (GPP), NPP, the ratio NPP/GPP and net ecosystem exchange (NEE), are used to assess the potential productivity changes of forests. In our analysis, we covered the full heterogeneity in site productivity in the study region. Thus, we consider any reductions in productivity (carbon cycle) through the close link to other ecosystem processes and cycles (water, nitrogen, energy) as a potential threat to forest functioning and thus the provision of ecosystem services. This includes the provision of fresh air, shading and cooling, intact soil (mediated by the litter input and root growth) for proper filtering of drinking water, protection against soil erosion and flooding and the pleasure wandering about in a lush, healthy forest.
GPP strongly depend on daily weather, the nitrogen availability and feedback loops with LAI. The ratio NPP/GPP is a measure of the effective forest productivity. A decreas of NPP/GPP indicates a higher plant internal respiration and a lower biomass build-up. The total ecosystem carbon balance is given by the NEE (g C m−2 day−1). NEE is NPP reduced by heterotrophic respiration, which is dependent on temperature, soil water content and soil mineral nitrogen availability.
Analyses and results
Model validation
We applied Biome-BGC to the 32 experimental beech plots in the region. We converted the simulation results for stem carbon to timber volume using biomass expansion factors (Pietsch et al. 2005) and compared the predicted volume with the observed stand volume (Fig. 5). The mean predicted and observed volumes are 362 and 364 m3 ha−1, respectively. Standard deviation of the differences between predicted and observed values is 81 m3 ha−1 or 22.2 % of the observed mean volume. The paired Student’s t test revealed no significant difference between predicted and observed volume (Δ = −2 m3 ha−1, t = 0.153 < t α = 0.025, N = 31 = 2.04). The linear regression analysis of observed versus predicted volume (Fig. 5) gives an r 2 of 0.80, an intercept of −34 m3 ha−1 and a slope of 1.10. The volume residual analysis showed no trends versus elevation, slope, aspect and mean stand age (Fig. 5). The confidence (CI) and prediction intervals (PI) according to Reynolds (1984) revealed unbiased results, with a CI of −31 to +27 (−8.6 to +7.4 %) and a PI of −169 to +165 (−46.5 to +45.3 %). The CI examines divergences between the expected differences if the model is repeatedly used; the PI provides the range of the error in future applications. This suggests that Biome-BGC produces consistent and unbiased results and is a suitable diagnostic tool for our analysis.

Volume observations versus predictions for validation plots including the trend analysis of standardised volume residuals versus site and stand parameters
Ecophysiological indicators
We calculated the ecophysiological indicators for the 782 forest grid points between 1960 and 2100. Linear regression (Table 1) and periodical mean values (Table 2) were calculated for the annual sums (Tr, Ev, Of, Nmin), annual means (WUE) or seasonal means (Ψ, mean for months June–August), averaged over all grid points. Annual evaporation is projected to increase in the A1B scenario, whereas transpiration is projected to decrease under A1B + Prcp and B1 + Prcp (Tables 1, 2) and so is the sum of Ev and Tr, evapotranspiration (ET, not shown). Annual WUE increased in every period and scenario (Tables 1, 2) and shows considerable annual variation (Fig. 6). In contrast, the summer time soil water potential shows persistent decrease, except for the B1 scenario (Tables 1, 2). The changes in mean monthly Psi between the 1960s and the 2090s are shown in Fig. 6, where the decrease in the summer months is apparent. Regional differences in mean summer Psi (Fig. 8c) and elevation trends (Fig. 9d–f) are evident. The annual Of from the bulk soil is projected to increase under A1B + Prcp and B1 + Prcp (Tables 1, 2). Net soil nitrogen mineralisation rates show a positive response to all four scenarios with highest rates in A1B, followed by A1B + Prcp (Tables 1, 2).

Modelled mean annual water use efficiency (WUE, NPP/transpiration, g C mm−1 H2O) (top) and modelled monthly soil water potential (Psi, MPa) averaged for single decades (each tick indicates a month with Psi values averaged over the respective decade, bottom) using the historic climate observations and the two future temperature scenarios (A1B, B1) as well as a combined temperature—sinusoidal precipitation scenarios. Averaged over the 782 forest grid points
Ecosystem productivity indicators
The productivity indicators LAI, GPP, NPP and NEE for the period 1960–2100 were calculated for each of the 782 forest grid points (Fig. 7; Tables 1, 2). All four measures exhibit an increase for the period of 1960–2009. Among the climate change scenarios, A1B exhibits the highest increase for all indicators, whereas B1 + Prcp shows no change. A regional forest map with NPP averaged for 1990–2009 reveals the spatial variation of productivity in the region (Fig. 8a). Elevation gradients and differences between mean values for 1990–2009 and 2081–2100 are apparent (Fig. 9g–i). High productivity is apparent in the vicinity of Vienna, sub-region NE (Figs. 8a, 9g). A second map shows the relative changes of the period 2081–2100 versus 1990–2009 for A1B, the scenario with the highest productivity increase (Fig. 8b). The highest relative increase of up to 25 % is primarily found in high elevations (Figs. 8a, 9g–i). Under A1B + Prcp, NPP increases are less than half compared to the A1B scenario (Table 1; Fig. 9g–i). We calculated the correlation between the relative NPP increase (Fig. 8b) and the summer Psi (Fig. 8c). The correlation coefficient of 0.52 indicates strong correlation.

Modelled average annual GPP, NPP and NEE using the historic climate observations and the two future temperature scenarios (A1B, B1) as well as a combined temperature—sinusoidal precipitation scenarios. Averaged over the 782 forest grid points

(a) Mean annual NPP for the period 1990–2009 for the forested area in the study region and (b) relative annual NPP changes to the period 2081–2100, (c) and mean summer time (June–August) soil water potential Psi for the period 2081–2100 with the scenario A1B; interpolated with ArcGIS Kriging interpolation tools using the standard settings of ordinary Kriging

Elevation trends for annual nitrogen deposition (a–c), summer (June–August) Psi (d–f) and annual NPP (g–i) averaged for the periods 1990–2009 and 2081–2100 for A1B and A1B + Prcp for selected forest grid points from the sub-regions in the north-east (NE, a, d, g), south-east (SE, b, e, h) and west (W, c, f, i); sub-regions are defined in Fig. 1
Discussion
Climatic effects
For the last 50 years (1960–2009), an increase in daily minimum temperature of 2.6 °C and in daily maximum temperature of 1.4 °C (Table 1) with large regional differences in the study area has been evident (Fig. 2). Until the end of the twenty-first century, climate change scenarios project an increase in average temperature of ~4.4 °C (A1B) and ~2.6 °C (B1); however, the projected changes vary for T min and T max, along with the seasons (Figs. 3, 4). Past (1960–2009) precipitation changes were not significant (Table 1), and CLM did not project strong Prcp trends for A1B and B1. Therefore, we conducted our study with unchanged Prcp and with our own Prcp scenario, assuming increased winter and decreased summer precipitation following assessments of the IPCC for Central Europe (Christensen et al. 2007).
Higher temperatures have led to a prolongation of the growing season in Austria (Hasenauer et al. 1999) and can partly explain higher productivity in Austrian forests (Eastaugh et al. 2011). A moderate temperature increase fastens turnover and mineralisation processes (Lloyd and Taylor 1994) and therefore potentially increases ecosystem productivity. Overdieck et al. (2007), for example, show higher growth rates of juvenile beech trees with higher temperatures compared to growth rates at elevated CO2 in a non-water-limited system. Temperature responses are represented by Biome-BGC in many of the modelled steps in the carbon and nitrogen cycles and include known negative effect of temperature on the carbon balance (Thornton 1998). For example, autotrophic respiration increases with temperature. Oxygenation reactions during photosynthesis (photorespiration) increase faster with temperature than carboxylation reactions (CO2 assimilation, Lambers et al. 2008). Higher maintenance respiration changes the carbon balance of forests and plays a significant role during the process of carbon starvation, where prolonged stomatal closure inhibits photosynthesis and carbon stores are depleted (McDowell et al. 2011). As an immediate effect of higher temperature, higher VPD (Tables 1, 2) increases the evaporative demand, the driving force for ET.
Impacts on ecophysiological indicators
Changes in the ET rates and precipitation patterns modify the Of (Tables 1, 2). In combination with a projected decrease in ET, an increase in stand Of is projected for A1B + Prcp and B1 + Prcp. The analysis of the sub-regions indicates that in both high and low elevations, our assumed Prcp decrease in summer and increase in winter causes a decrease in ET and Of in summer but an increase in Of in winter (not shown), causing the increase in annual Of.
The WUE is projected to increase for all the scenarios, especially for A1B + Prcp (Tables 1, 2). This confirms results of Forstreuter (2002), who experimentally found higher WUE for beech under elevated CO2 concentrations and of Peñuelas et al. (2008), who detected increased intrinsic WUE with increasing CO2 and temperature for mature beech trees in Catalonia (Davi et al. 2006) also predict continued increase of WUE for two French beech forest sites from 1960 to 2100.
We project lowered summer Psi for all but one scenario. The exception is the B1 scenario because neither Tr nor Ev are projected to change (Tables 1, 2). Regional differences in mean summer Psi under higher temperatures (Fig. 8c) are accompanied by strong elevation trends (Fig. 9d–f). This means drier soil conditions in low elevations and thus a stronger climate change impact, especially for the SE (SE sub-region). An additional decrease in summer Prcp has a strong negative effect on the soil water status (Fig. 9d–f) and, in the following, stomatal conductance and thus CO2 uptake are limited (compare following chapter). Low Psi also limits mineralisation processes. A correlation between annual Nmin and Psi, however, cannot be directly derived from our results because different trends in primary production and thus litter production disguise this effect.
Impacts on ecosystem productivity indicators
Depending on the scenario, the ecosystem productivity indicators LAI, GPP, NPP and NEE reveal a continued increase or preservation of the current productivity level (Fig. 7; Table 1). For both A1B and B1, the additional scenario of reduced summer and increased winter precipitation reduces LAI, GPP, NPP and NEE compared to the no-Prcp change scenarios. This indicates a response to limited water supply during the growing season corresponding to the lower summer Psi (compare previous chapter). Spatial patterns in the productivity indicators are the result of the combined effect of spatially varying temperature, precipitation, soil properties and nutrient supply. The highest NPP values for the period 1990–2009 are evident for areas with high nitrogen deposition rates, such as the forests in the immediate vicinity of the city of Vienna (Figs. 8a, 9a, g). The analysis of the three sub-regions (Fig. 1) reveals that responding to decreasing summer Psi towards lower elevations (Fig. 9d–f), annual NPP decreases with elevation (Fig. 9g–i). Therefore, the relative NPP increase (mean 2081–2100 compared to mean 1990–2009, shown for A1B or A1B + Prcp) is smaller in low elevations (Figs. 8b, c, 9g–i). The positive effect of increasing temperatures on productivity thus is more pronounced in higher elevations, whereas in lower elevations the productivity increase is limited by the water supply. The results are consistent with findings from the climate change impact study by Hlásny et al. (2011). They predict, using a multi-model approach, that beech forest productivity will increase in higher elevations but show less increase or even drought-induced decline in growth in the lower part of the species distribution range. The decreasing ratio NPP/GPP (Tables 1, 2) demonstrates a reduction in the efficiency of plant primary production because more carbon is required for maintenance respiration with increasing temperature. NEE (NPP minus heterotrophic respiration) is positive and increased during the past 50 years (Table 1). Although NEE is projected to further increase only together with increasing NPP (A1B), results suggest that the forest will remain a carbon sinks in all scenarios (Fig. 7). The underlying modelling assumptions are that timber is extracted during thinnings and final harvest, whereas leaves and roots are left in the forest to decompose. The detected patterns in GPP and ecosystem respiration increase are consistent with reports from water and carbon fluxes of two French EUROFLUX beech sites (Davi et al. 2006) where in a climate sensitivity study up to 60 % GPP and 100 % NEE increase for the period 1960–2100 are projected.
A debated phenomenon in climate change impact studies is the downregulation of productivity due to nitrogen or generally nutrient limitation under CO2 enrichment (Oren et al. 2001; Luo et al. 2004; Iversen and Norby 2008). Nutrient cycling, as implemented in Biome-BGC as a detailed nitrogen cycle, is important to detect possible downregulation of productivity (Medlyn et al. 2011). Our results show no clear sign of downregulation due to nitrogen limitation as the relative increase in NPP (Fig. 8b) is similar in regions with high N deposition (sub-region NE, inside city boundaries of Vienna, Fig. 9a) and areas with lower N deposition (e.g. lower elevations of sub-region SE; Fig. 9b). Also, increasing N mineralisation (Table 1) indicates no worsening nutrient limitation (Luo et al. 2004). Our results of no N limitation agree well with the estimated exceeding of the critical loads of eutrophication-causing nitrogen deposition by up to 15 kg N ha−1 year−1 in the study region (Obersteiner and Offenthaler 2008).
An uncertainty in the forest response to climate change is the potential limiting effect of elevated CO2 on the stomatal conductance. Such a response is not included in Biome-BGC due to inconsistent results from field studies on woody species (Saxe et al. 1998; Norby et al. 1999; Thornton et al. 2002). Effects vary among plant functional groups and are generally lower for woody plant species than for grasses and herbaceous plants (Ainsworth and Rogers 2007). Also for F. sylvatica, the reports range from no response to conduction reductions of more than 30 % (Dufrêne et al. 1993; Liozon et al. 2000; Forstreuter 2002; Keel et al. 2007). Reduced stomatal conductance with elevated CO2 could improve the WUE and increase productivity under water-limiting conditions (Saxe et al. 1998; Medlyn et al. 2011), but opposite results have also been reported (Warren et al. 2011).
In this climate change impact study, we did not model the risks of an increase in storm damage and the spread of forest-damaging agents (pathogens, insects), on the one hand, because storms and biotic disturbances are not part of the Biome-BGC model. On the other hand, susceptibility to storm damage is low for beech, compared to other dominant tree species, for example, Norway spruce (Schütz et al. 2006; Thom et al. 2013). Beech is also considered less susceptible to biotic damaging agents, although several different beech pests and pathogens are known (Steyrer 2009; Tomiczek et al. 2011). They include a number of secondary damaging agents (that follow drought periods and wind damages), which research pays special attention to at the xeric edge of the natural distribution range of beech (Lakatos and Molnár 2009, Rasztovits et al. 2013). A recent study in the biosphere reserve Vienna Woods, however, found no bark beetle damage even on wind-throw sites (Steyrer and Wieshaider 2010).
Forest management planning may be influenced by drought effects and require heightened attention to stand structure, harvesting regimes and (ground-) water management in the future (Lindner et al. 2010; Seidl et al. 2011; Hlásny et al. 2014). For this study, we did not consider alternatives to our implemented forest management routine. A careful adaptation of the shelterwood system may be necessary under climate change for Chakraborty et al. (2013) report a higher risk of crown dieback and even total mortality in dry years for beech regeneration in the understory of adult trees due to competition for water. Since Biome-BGC does not simulate different tree layers, this effect could not be tested. Alternative future management scenarios could also include increased biomass extraction for energy production (e.g. fine branches or leaves), consequently changing NEE and the nutrient cycle, or shortened rotation periods to make use of the increasing productivity. Shortened rotation periods could reduce the risk of red heartwood in the beech stems that develops with age (Knoke 2003) and also reduce the risk of disturbances (i.e. storms).
Impacts on welfare and recreation
The projected impact of climate change on ecophysiology and productivity and thus on forest functions of the Vienna Woods may have implications on welfare and recreation.
The projected reduction in transpiration (Tables 1, 2) will reduce evaporative cooling in the canopy. The extent to which this cooling in the canopy will effect potential cooling effects within a forest stand is difficult to assess (Oke et al. 1989). Changes in the ET rates, precipitation patterns and WUE are projected to influence Of and thus potentially drinking water sources (Tables 1, 2). Our projections of stable productivity (see Fig. 7; Table 1) fulfil the very general requirement of stable forests for sustainable drinking water supply (Richards et al. 2012). The preservation of beech forests is beneficial because of its high effectiveness in filtering out nitrates that typically harm drinking water quality (von Wilpert et al. 2000). Following Bartsch (2000), problems with drinking water quality cannot be excluded, where forests on shallow soils are disturbed and low pH in the surface soil increases nitrate leaching and hinders fast vegetation growth after gap creation.
The projected increase in LAI (Table 2) may have several positive implications: (1) a high LAI is important for flood protection because water is intercepted, evaporated and transpired, and thus, run-off rates are reduced (Badoux et al. 2006; Pötzelsberger and Hasenauer 2015); (2) a well-developed canopy provides shade for recreation seeking visitors; (3) a high LAI guarantees continued litter production, and therefore, an organic-rich well-structured soil that filters, adsorbs and transforms pollutants and therefore is important for clean drinking water supply (Dudley and Stolton 2003; Blume et al. 2010). Such a well-developed soil, in combination with a dense canopy and litter layer, is also more resistant to erosion (Brang et al. 2001; Salles et al. 2002).
NEE is projected to stay positive under all climate change scenarios and of unaltered forest management, implying that these forests act as carbon sinks and thus help mitigating climate change. In a strict sense, this climate change mitigation effect applies to a situation, where the extracted wood is stored permanently and is not burned or left to decompose somewhere outside the forest.
Conclusion
With biogeochemical mechanistic ecosystem modelling, we were able to project changes in ecophysiological and productivity indicators for the Vienna Woods under two different temperature scenarios combined with two variants for precipitation (increased winter and decreased summer precipitation, or no change). Changes in the carbon, water and nutrient pools and fluxes are diverse due to multiple feedback loops and different temperature dependencies of biological and geochemical processes. Changes in precipitation pattern plus temperature increase are projected to decrease annual ET, increase the WUE and increase water Of in winter. Annual NPP and NEE are projected to increase for one of the four scenarios. Lower summer soil water potentials especially in lower elevations limit the productivity increase. Overall, we project that the Vienna Woods will continue to fulfil their key forest functions.
References
Ainsworth EA, Rogers A (2007) The response of photosynthesis and stomatal conductance to rising [CO2]: mechanisms and environmental interactions. Plant Cell Environ Environ 30:258–270. doi:10.1111/j.1365-3040.2007.01641.x
Albrecht FK (2011) GIS-gestützte Analyse der gesellschaftlichen Leistungen des Wiener Wald- und Wiesengürtels (GIS aided analysis of the social services of the Greenbelt Vienna). Master thesis, University of Natural Resources and Life Sciences, Vienna
Badoux A, Witzig J, Germann PF et al (2006) Investigations on the runoff generation at the profile and plot scales, Swiss Emmental. Hydrol Process 20:377–394. doi:10.1002/hyp.6056
Bartsch N (2000) Element release in beech forest gaps. Water Air Soil Pollut 122:3–16
Blume H-P, Brümmer GW, Horn R et al (2010) Scheffer/Schachtschabel Lehrbuch der Bodenkunde (Textbook of soil science), 16th edn. Spektrum Akademischer Verlag, Heidelberg
Brang P, Schönenberger W, Ott E, Gardner B (2001) Forests as protection from natural hazards. In: Evans J (ed) Forest handbook. Blackwell Scinece, Oxford, pp 53–81
Carreiro MM, Song Y-C, Wu J (2008) Ecology, planning, and management of urban forests: an international perspective. Springer, New York
Chakraborty T, Saha S, Reif A (2013) Decrease in available soil water storage capacity reduces vitality of young understorey European Beeches (Fagus sylvatica L.)—a case study from the Black Forest, Germany. Plants 2:676–698. doi:10.3390/plants2040676
Christensen JH, Hewitson B, Busuioc A et al (2007) Regional climate projections. In: Solomon S, Qin D, Manning M et al (eds) Climate change 2007: physical science basis. Contribution of working group I to the fourth assessment report of the intergovermental panel on climate change. Cambridge University Press, Cambridge, pp 847–940
Clapp RB, Hornberger GM (1978) Empirical equations for some soil hydraulic properties. Water Resour Res 14:601–604. doi:10.1029/WR014i004p00601
Cosby BJ, Hornberger GM, Clapp RB, Ginn TR (1984) A statistical exploration of the relationships of soil moisture characteristics to the physical properties of soils. Water Resour Res 20:682–690. doi:10.1029/WR020i006p00682
Davi H, Dufrêne E, Francois C et al (2006) Sensitivity of water and carbon fluxes to climate changes from 1960 to 2100 in European forest ecosystems. Agric For Meteorol 141:35–56. doi:10.1016/j.agrformet.2006.09.003
Deutscher Wetterdienst (2014) Regionaler Klimawandel—Klimamodelle im Vergleich (Regional climate change—comparison of climate models). http://www.dwd.de/bvbw/appmanager/bvbw/dwdwwwDesktop?_nfpb=true&_pageLabel=dwdwww_start&T99803827171196328354269gsbDocumentPath=Navigation/Oeffentlichkeit/Homepage/Klimawandel/ZWEK__T__node.html?__nnn=true. (Accessed 16 Dec 2014)
Dobbs C, Escobedo FJ, Zipperer WC (2011) A framework for developing urban forest ecosystem services and goods indicators. Landsc Urban Plan 99:196–206. doi:10.1016/j.landurbplan.2010.11.004
Dudley N, Stolton S (2003) Running pure: the importance of forest protected areas to drinking water. The World Bank/WWF International, Washington D.C., USA
Dufrêne E, Pontallier J-Y, Saugier B (1993) A branch bag technique for simultaneous CO2 enrichment and assimilation measurements on beech (Fagus sylvatica L.). Plant Cell Environ 16:1131–1138. doi:10.1111/j.1365-3040.1996.tb02071.x
Dukes JS, Pontius J, Orwig D et al (2009) Responses of insect pests, pathogens, and invasive plant species to climate change in the forests of northeastern North America: what can we predict? Can J For Res 39:231–248. doi:10.1139/X08-171
Eastaugh CS, Pötzelsberger E, Hasenauer H (2011) Assessing the impacts of climate change and nitrogen deposition on Norway spruce (Picea abies L. Karst) growth in Austria with BIOME-BGC. Tree Physiol 31:262–274. doi:10.1093/treephys/tpr033
Ellenberg H, Leuschner C (2010) Vegetation Mitteleuropas mit den Alpen (Vegetation ecology of central Europe), 6th edn. Ulmer, Stuttgart
Enting IG, Wigley TML, Heimann M, Scientific C (1994) Future emissions and concentrations of carbon dioxide: key ocean/atmosphere/land analyses. Division of Atmospheric Research technical paper no. 31. CSIRO, Australia
Farquhar GD, von Caemmerer S, Berry JA (1980) A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 149:78–90
Foet M-C (2010) Der Wiener Grüngürtel: Leistungen und Nutzen für die Gesellschaft (The Vienna green belt). Master thesis, University of Natural Resources and Life Sciences, Vienna
Formayer H, Haas P, Nadeem I (2010) Regional climate model scenarios of CC-WaterS. Explanatory report. University of Natural Resources and Life Sciences, Vienna
Forstreuter M (2002) Auswirkungen globaler Klimaänderungen auf das Wachstum und den Gaswechsel (CO2/H2O) von Rotbuchenbeständen (Fagus sylvatica L.) (Impacts of global climate change on the growth and gas exchange of European beech stands). Landschaftsentwicklung und Umweltforschung - Schriftenreihe der Fakultät VII Architektur Umwelt Gesellschaft, 119, Technische Universität Berlin
Furniss MJ, Staab BP, Hazelhurst S et al (2010) Water, climate change, and forests. US Department of Agriculture, Forest Service, Pacific Northwest Research Station
Granier A, Reichstein M, Bréda N et al (2007) Evidence for soil water control on carbon and water dynamics in European forests during the extremely dry year: 2003. Agric For Meteorol 143:123–145. doi:10.1016/j.agrformet.2006.12.004
Hasenauer H, Nemani RR, Schadauer K, Running SW (1999) Forest growth response to changing climate between 1961 and 1990 in Austria. For Ecol Manag 122:209–219. doi:10.1016/S0378-1127(99)00010-9
Hasenauer H, Merganicova K, Petritsch R et al (2003) Validating daily climate interpolations over complex terrain in Austria. Agric For Meteorol 119:87–107. doi:10.1016/S0168-1923(03)00114-X
Hlásny T, Barcza Z, Fabrika M et al (2011) Climate change impacts on growth and carbon balance of forests in Central Europe. Clim Res 47:219–236. doi:10.3354/cr01024
Hlásny T, Barcza Z, Barka I et al (2014) Future carbon cycle in mountain spruce forests of Central Europe: modelling framework and ecological inferences. For Ecol Manag 328:55–68. doi:10.1016/j.foreco.2014.04.038
Holland EA, Dentener FJ, Braswell BH, Sulzman JM (1999) Contemporary and pre-industrial global reactive nitrogen budgets. Biogeochemistry 46:7–43. doi:10.1007/BF01007572
Hollweg H-D, Böhm U, Fast I et al (2008) Ensemble simulations over Europe with the regional climate model CLM forced with IPCC AR4 global scenarios. Technical report 3. Max-Planck-Institut für Meteorologie, Hamburg
Hyvönen R, Ågren GI, Linder S et al (2006) The likely impact of elevated [CO2], nitrogen deposition, increased temperature and management on carbon sequestration in temperate and boreal forest ecosystems: a literature review. New Phytol 173:463–480
IPCC (1992) Climate change 1992: the supplementary report to the IPCC scientific assessment. Cambridge University Press, Cambridge
IPCC (2000) Emission scenarios: a special report of IPCC working group III. Cambridge University Press, Cambridge
Iversen CM, Norby RJ (2008) Nitrogen limitation in a sweetgum plantation: implications for carbon allocation and storage. Can J For Res 38:1021–1032. doi:10.1139/X07-213
Jacob D, Bülow K, Kotova L et al (2012) Climate Service Center report 6, Regionale Klimaprojektionen für Europa und Deutschland: Ensemble-Simulationen für die Klimafolgenforschung. Climate Service Center, Hamburg
Keel SG, Pepin S, Leuzinger S, Körner C (2007) Stomatal conductance in mature deciduous forest trees exposed to elevated CO2. Trees 21:151–159. doi:10.1007/s00468-006-0106-y
Knoke T (2003) Predicting red heartwood formation in beech trees (Fagus sylvatica L.). Ecol Model 169:295–312. doi:10.1016/S0304-3800(03)00276-X
Konijnendijk CC (2008) The forest and the city: the cultural landscape of urban woodland. Springer, New York
Konijnendijk C, Nilsson K, Randrup T, Schipperijn J (2005) Urban forests and trees: a reference book. Springer, Berlin
Koukal T (2004) Nonparametric assessment of forest attributes by combination of field data of the Austrian forest inventory and remote sensing data. PhD thesis, University of Natural Resources and Life Sciences, Vienna
Lakatos F, Molnár M (2009) Mass Mortality of Beech (Fagus sylvatica L.) in South-West Hungary. Acta Silv Lign Hung 5:75–82
Lambers H, Chapin FS, Pons TL (2008) Plant physiological ecology, 2nd edn. Springer, New York
Landsteiner V (1990) Wienerwald (Vienna Woods). Österreichische Bundesforste (Austrian Federal Forests), Vienna
Larcher W (1995) Physiological plant ecology. Springer, Berlin
Larsson T-B (2001) Ecological bulletins 50: biodiversity evaluation tools for European forests. Wiley, Hoboken
Lautenschlager M, Keuler K, Wunram C et al (2009a) Climate simulation with CLM, scenario A1B run no. 1, data stream 3: European region MPI-M/MaD. World Data Center for Climate, Hamburg. doi:10.1594/WDCC/CLM_A1B_1_D3
Lautenschlager M, Keuler K, Wunram C et al (2009b) Climate simulation with CLM, scenario B1 run no. 1, data stream 3: European region MPI-M/MaD. World Data Center for Climate, Hamburg. doi:10.1594/WDCC/CLM_B1_1_D3
Lindner M, Maroschek M, Netherer S et al (2010) Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For Ecol Manag 259:698–709. doi:10.1016/j.foreco.2009.09.023
Liozon R, Badeck F, Genty B et al (2000) Leaf photosynthetic characteristics of beech (Fagus sylvatica) saplings during three years of exposure to elevated CO2 concentration. Tree Physiol 20:239–247
Lloyd J, Taylor JA (1994) On the temperature dependence of soil respiration. Funct Ecol 8:315–323
Luo Y, Su B, Currie WS et al (2004) Progressive nitrogen limitation of ecosystem responses to rising atmospheric carbon dioxide. Bioscience 54:731–739. doi:10.1641/0006-3568(2004)054[0731:PNLOER]2.0.CO;2
Luo Y, Gerten D, Le Maire G et al (2008) Modeled interactive effects of precipitation, temperature, and [CO2] on ecosystem carbon and water dynamics in different climatic zones. Glob Chang Biol 14:1986–1999. doi:10.1111/j.1365-2486.2008.01629.x
Mátyás C, Berki I, Czúcz B et al (2010) Future of beech in southeast Europe from the perspective of evolutionary ecology. Acta Silv Lign Hung 6:91–110
McDowell NG, Beerling DJ, Breshears DD et al (2011) The interdependence of mechanisms underlying climate-driven vegetation mortality. Trends Ecol Evol 26:523–532. doi:10.1016/j.tree.2011.06.003
Medlyn BE, Duursma RA, Zeppel MJB (2011) Forest productivity under climate change: a checklist for evaluating model studies. Wiley Interdiscip Rev Clim Chang 2:332–355. doi:10.1002/wcc.108
Millar CI, Stephenson NL, Stephens SL (2007) Climate change and forests of the future: managing in the face of uncertainty. Ecol Appl 17:2145–2151
Norby RJ, Wullschleger SD, Gunderson CA et al (1999) Tree responses to rising CO2 in field experiments: implications for the future forest. Plant Cell Environ 22:683–714
Obersteiner E, Offenthaler I (2008) Critical Loads für Schwefel- und Stickstoffeinträge in Ökosysteme (Critical loads for sulfur and nitrogen deposition in ecosystems). Environment Agency Austria (Umweltbundesamt), Vienna
Oke TR, Crowther JM, McNaughton KG et al (1989) The micrometeorology of the urban forest [and discussion]. Philos Trans R Soc B Biol Sci 324:335–349. doi:10.1098/rstb.1989.0051
Oleyar MD, Greve AI, Withey JC, Bjorn AM (2008) An integrated approach to evaluating urban forest functionality. Urban Ecosyst 11:289–308. doi:10.1007/s11252-008-0068-5
Oren R, Ellsworth DS, Johnsen KH et al (2001) Soil fertility limits carbon sequestration by forest ecosystems in a CO2-enriched atmosphere. Nature 411:469–472. doi:10.1038/35078064
Overdieck D, Ziche D, Böttcher-Jungclaus K (2007) Temperature responses of growth and wood anatomy in European beech saplings grown in different carbon dioxide concentrations. Tree Physiol 27:261–268
Peñuelas J, Ogaya R, Boada M, Jump AS (2007) Migration, invasion and decline: changes in recruitment and forest structure in a warming-linked shift of European beech forest in Catalonia (NE Spain). Ecography 30:829–837. doi:10.1111/j.2007.0906-7590.05247.x
Peñuelas J, Hunt JM, Ogaya R, Jump AS (2008) Twentieth century changes of tree-ring δ13C at the southern range-edge of Fagus sylvatica: increasing water-use efficiency does not avoid the growth decline induced by warming at low altitudes. Glob Chang Biol 14:1076–1088. doi:10.1111/j.1365-2486.2008.01563.x
Peters EB, McFadden JP (2010) Influence of seasonality and vegetation type on suburban microclimates. Urban Ecosyst 13:443–460. doi:10.1007/s11252-010-0128-5
Petritsch R (2002) Anwendung und Validierung des Klimainterpolationsmodells DAYMET in Österreich (Application and validation of the climate interpolation model DAYMET in Austria). Master thesis, University of Natural Resources and Life Sciences, Vienna
Petritsch R (2008) Large scale mechanistic ecosystem modeling in Austria. PhD thesis, University of Natural Resources and Life Sciences, Vienna
Petritsch R, Hasenauer H (2007) Interpolating input parameters for large scale ecosystem models. Austrian J For Sci 124:135–151
Petritsch R, Hasenauer H (2009) Tägliche Wetterdaten im 1 km Raster von 1960 bis 2008 über Österreich (Grided Daily weather data between 1960 and 2008 across Austria). Austrian J For Sci 126:215–225
Pietsch SA, Hasenauer H (2002) Using mechanistic modeling within forest ecosystem restoration. For Ecol Manag 159:111–131. doi:10.1016/S0378-1127(01)00714-9
Pietsch SA, Hasenauer H (2006) Evaluating the self-initialization procedure for large-scale ecosystem models. Glob Chang Biol 12:1658–1669. doi:10.1111/j.1365-2486.2006.01211.x
Pietsch SA, Hasenauer H (2009) Photosynthesis within large-scale ecosystem models. In: Laisk A, Nedbal L, Govindjee (eds) Advances in photosynthesis and respiration. Photosynthesis in silico, vol 29. Springer, Dordrecht, pp 441–464
Pietsch SA, Hasenauer H, Thornton PE (2005) BGC-model parameters for tree species growing in central European forests. For Ecol Manag 211:264–295. doi:10.1016/j.foreco.2005.02.046
Pötzelsberger E, Hasenauer H (2015) Forest—water dynamics within a mountainous catchment in Austria. Nat Hazards. doi:10.1007/s11069-015-1609-x
Pretzsch H, Biber P, Schütze G et al (2014) Forest stand growth dynamics in Central Europe have accelerated since 1870. Nat Commun 5:4967. doi:10.1038/ncomms5967
Rasztovits E, Berki I, Mátyás C et al (2013) The incorporation of extreme drought events improves models for beech persistence at its distribution limit. Ann For Sci 71:201–210. doi:10.1007/s13595-013-0346-0
Renner M, Sweeney S, Kubit J (2008) Green jobs: towards decent work in a sustainable, low-carbon world. UNEP, Nairobi
Reynolds MR (1984) Estimating the error in model predictions. For Sci 30:454–469
Richards WH, Koeck R, Gersonde R et al (2012) Landscape-scale forest management in the municipal watersheds of Vienna, Austria, and Seattle, USA: commonalities despite disparate ecology and history. Nat Areas J 32:199–207
Ryan MG (1991) Effects of climate change on plant respiration. Ecol Appl 1:157–167
Salles C, Poesen J, Sempere-Torres D (2002) Kinetic energy of rain and its functional relationship with intensity. J Hydrol 257:256–270. doi:10.1016/S0022-1694(01)00555-8
Saxe H, Ellsworth DS, Heath J (1998) Tree and forest functioning in an enriched CO2 atmosphere. New Phytol 139:395–436
Saxton KE, Rawls WJ, Romberger JS, Papendick RI (1986) Estimating generalized soil-water characteristics from texture. Soil Sci Soc Am J 50:1031. doi:10.2136/sssaj1986.03615995005000040039x
Schütz J-P, Götz M, Schmid W, Mandallaz D (2006) Vulnerability of spruce (Picea abies) and beech (Fagus sylvatica) forest stands to storms and consequences for silviculture. Eur J For Res 125:291–302. doi:10.1007/s10342-006-0111-0
Seidl R, Rammer W, Lexer MJ (2011) Climate change vulnerability of sustainable forest management in the Eastern Alps. Clim Chang 106:225–254. doi:10.1007/s10584-010-9899-1
Seidl R, Schelhaas M-J, Rammer W, Verkerk PJ (2014) Increasing forest disturbances in Europe and their impact on carbon storage. Nat Clim Chang 4:806–810. doi:10.1038/nclimate2318
Steyrer G (2009) Buchenborkenkäfer: Projekt im Biosphärenpark Wienerwald (Beech bark beetle: project in the biosphere reserve Vienna Woods). For Aktuell 45:9–11
Steyrer G, Wieshaider A (2010) Entwarnung im Biosphärenpark Wienerwald: derzeit keine Massenvermehrung des Buchenborkenkäfers (All-clear in the biosphere reserve Vienna Woods: currently no mass outbreak of the beech bark beetle). BFW-Austrian Research Centre For, Vienna. http://bfw.ac.at/rz/bfwcms2.web?dok=8538. (Accessed 16 Oct 2014)
Tegel W, Seim A, Hakelberg D et al (2013) A recent growth increase of European beech (Fagus sylvatica L.) at its Mediterranean distribution limit contradicts drought stress. Eur J For Res 133:61–71. doi:10.1007/s10342-013-0737-7
Thom D, Seidl R, Steyrer G et al (2013) Slow and fast drivers of the natural disturbance regime in Central European forest ecosystems. For Ecol Manag 307:293–302. doi:10.1016/j.foreco.2013.07.017
Thornton PE (1998) Regional ecosystem simulation: combining surface- and satellite-based observations to study linkages between terrestrial energy and mass budgets. PhD thesis, University of Montana, Missoula
Thornton PE, Running SW, White M (1997) Generating surfaces of daily meteorological variables over large regions of complex terrain. J Hydrol 190:214–251. doi:10.1016/S0022-1694(96)03128-9
Thornton PE, Hasenauer H, White M (2000) Simultaneous estimation of daily solar radiation and humidity from observed temperature and precipitation: an application over complex terrain in Austria. Agric For Meteorol 104:255–271. doi:10.1016/S0168-1923(00)00170-2
Thornton PE, Law BE, Gholz HL et al (2002) Modeling and measuring the effects of disturbance history and climate on carbon and water budgets in evergreen needleleaf forests. Agric For Meteorol 113:185–222. doi:10.1016/S0168-1923(02)00108-9
Tomiczek C, Perny B, Cech TL (2011) Zur Waldschutzsituation der Buche (Forest pests and diseases of beech). BFW-Praxisinformation 12:19–21
Von Wilpert K, Zirlewagen D, Kohler M (2000) To what extent can silviculture enhance sustainability of forest sites under the immission regime in Central Europe. Water Air Soil Pollut 122:105–120
Warren JM, Norby RJ, Wullschleger SD (2011) Elevated CO2 enhances leaf senescence during extreme drought in a temperate forest. Tree Physiol 31:117–130. doi:10.1093/treephys/tpr002
White M, Thornton PE, Running SW, Nemani RR (2000) Parameterization and sensitivity analysis of the BIOME-BGC terrestrial ecosystem model: net primary production controls. Earth Interact 4:1–85. doi:10.1175/1087-3562(2000)004<0003:PASAOT>2.0.CO;2
Woodall CW, Nowak DJ, Liknes GC, Westfall JA (2010) Assessing the potential for urban trees to facilitate forest tree migration in the eastern United States. For Ecol Manag 259:1447–1454. doi:10.1016/j.foreco.2010.01.018
Young RF (2010) Managing municipal green space for ecosystem services. Urban For Urban Green 9:313–321. doi:10.1016/j.ufug.2010.06.007
Zheng D, Ducey MJ, Heath LS (2013) Assessing net carbon sequestration on urban and community forests of northern New England, USA. Urban For Urban Green 12:61–68. doi:10.1016/j.ufug.2012.10.003
Acknowledgments
This work was financed by the City of Vienna in support of the EFI Central-East European Regional Office. Our thanks goes to Ervin Rasztovits and Norbert Móricz for their support in retrieving the CLM climate change scenario data. We thank Roland Köck and two anonymous reviewers for their valuable comments and Adam Moreno for English editing.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Martin Moog.
Rights and permissions
About this article

Cite this article
Pötzelsberger, E., Wolfslehner, B. & Hasenauer, H. Climate change impacts on key forest functions of the Vienna Woods. Eur J Forest Res 134, 481–496 (2015). https://doi.org/10.1007/s10342-015-0866-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-015-0866-2