
Abstract
Probiotics can modulate gut-liver axis. Effects of administration of multistrain probiotics (MP) on liver damage and overall health of trypanosomic rats were investigated. Thirty male rats randomly assigned to five groups A–E (n = 6), and MP containing Bifidobacterium BB-12, Lactobacillus delbrueckii LBY-27, Lactobacillus acidophilus LA-Z, Streptococcus thermophilus STY-31 and Lactobacillus paracasei LC-01 were used for supplementation. From day 0 post-supplementation (PS), rats in groups A–C received 1 × 109, 5 × 109 and 10 × 109 CFUs of MP daily and respectively. Groups D and E received no MP supplementation being the infected and uninfected controls respectively. On day 7PS, groups A–D were challenged with approximately 1 × 106 Trypanosoma brucei intraperitoneally. On days 0, 7 and 16, PS, serum alanine and aspartate aminotransferases (ALT and AST) were determined spectrophotometrically, while parasitaemia, bodyweight and mortality were monitored. By day 7, PS groups B-D had similar ALT activities and AST-ALT ratios, while all groups had similar AST activities. On day 16, PS all infected groups (A–D) had similar and significantly (p < 0.05) lower AST activities and AST-ALT ratio compared to the uninfected control, while ALT activities were similar and significantly elevated in groups A-D. Logarithmic parasitaemia levels and AST-ALT ratio showed random moderate negative correlation (r = − 0.390; p = 0.059). No significant variations in bodyweight were seen, and all infected groups had 100% mortality by day 27 post-supplementation. Supplementation with MP was innocuous to liver function in naïve rats, but failed to improve liver-function parameters, bodyweight or survivability of trypanosomic rats.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Probiotics are live micro-organisms or the components thereof, which can offer one or more health benefits when administered or consumed by humans or animals. Promising outcomes on their usefulness in management of several health conditions have been reported, including applications in certain cancers (Ali et al. 2019), gastrointestinal disorders (Gogineni et al. 2013), plasmodium infection (Villarino et al. 2016), trypanosomosis (Okolo et al. 2020), etc. Besides their use as growth promoters, probiotics alongside prebiotics are now being explored as possible complement or replacement for antibiotics in poultry, livestock and cultured fishes which are known to be easily susceptible to infectious disease (Gogineni et al. 2013; Aronu et al. 2019; Omeje et al. 2019).
The gut-liver axis is a well-established concept in gastrointestinal biopathology. Dysbiosis of gut microflora has been linked to several hepatopathies including alcoholic liver disease, liver fibrosis and fatty liver disease (Sharma et al. 2013; Engen et al. 2015). In a review, Khalesi et al. (2018) concluded that certain probiotic strains could significantly improve the activities of major liver enzymes including alanine aminotransferase and aspartate aminotransferase in experimental models of hepatopathies or in patients suffering liver diseases. Furthermore, safety of probiotic strains in human or animal consumers is an important developing issue. There have been confirmed reports of bacterial sepsis and aggravation of illnesses following treatment with some probiotic strains (Land et al. 2005).
Severe hepatopathy characterized by marked elevation of activities of liver function enzymes is one of the major features of African animal trypanosomosis caused by Trypanosoma brucei infections (Anosa 1988, Federico et al. 2016; Okolo et al. 2019a). The disease takes a heavy toll on animal health in Africa, where also the annual loss in monetary terms has been put at over $4.5 billion dollars (Yaro et al. 2016; Aneke et al. 2016; Abenga and Idowu, 2018). Experimental trials using strains of Lactobacillus casei (Bautista-Garfias et al. 2008) and Saccharomyces cerevisiae (Eze et al. 2012) against Trypanosoma spp. infections yielded positive results. However, studies evaluating the safety of probiotics strains or their ability to improve the liver damage which characterizes trypanosomosis are lacking. The rationale for the current study is to assess whether some proven probiotic strains can ameliorate or avert liver damage commonly seen in trypanosomosis.
For this study, proprietary multistrain probiotic mix containing Bifidobacterium BB-12, Lactobacillus delbrueckii LBY-27, Lactobacillus acidophilus LA-5, Streptococcus thermophilus STY-31 and Lactobacillus paracasei LC-01 was used. Well-documented health benefits of these probiotic strains in terms of positive immunomodulation and antioxidative capacities have been discussed (Hajifaraji et al. 2018; Okolo et al. 2020). The aim of this study was to evaluate the effect of treatment with the probiotic mix on liver function enzymes in naïve rats, and following infection with Trypanosoma brucei, to determine the effect of such treatment on liver aminotransferases and the overall well-being and survivability of the experimental models.
Materials and methods
Location and experimental animals
The research was carried out in the research laboratory of the Department of Veterinary Medicine, University of Nigeria Nsukka, located within geographical coordinates 6.85667° N 7.39583° E. Thirty (30) male post-pubertal Sprague Dawley rats were used for the study. They were housed in fly-proof cages and provided with water and proprietary rat feed fed ad libitum. Ethical judgements for use of the experimental animals in this study were based on the guidelines of the Animal Use and Care Committee of the Faculty of Veterinary Medicine, University of Nigeria, which agrees with the NIH procedures (NIH 2011).
Trypanosomes and probiotic strains
The Trypanosoma spp. used for the study was isolated from a dog patient diagnosed of canine trypanosomosis at the Veterinary Teaching Hospital, University of Nigeria Nsukka. The isolate was further identified as Trypanosoma brucei at the Department of Veterinary Parasitology and Entomology, University of Nigeria. Live multistrain probiotics blend sourced from CHR® (Netherlands) was used in the study. There were about 25 × 109 CFU of probiotic organisms in 1 g of the freeze dried culture. The blend contained equal proportion of five strains of probiotic organisms namely, Bifidobacterium BB-12, Lactobacillus delbrueckii LBY-27, Lactobacillus acidophilus LA-5, Streptococcus thermophilus STY-31 and Lactobacillus paracasei LC-01.
Study design, supplementation and infection with Trypanosoma brucei
A randomized controlled experimental design was used. Following 2 weeks of acclimatization, rats were randomly assigned to five groups A–E (n = 6). Groups A to C received 1 × 109 CFU, 5 × 109 CFU and 10 × 109 CFU of the multistrain probiotics (MP) respectively on a daily basis. Groups D and E did not receive any supplementation with probiotics being the infected and the uninfected controls respectively, rather each rat in groups D and E received 1 ml of distilled water, the vehicle for probiotics administration in groups A–C. Supplementation with multistrain probiotics (MP) started from the beginning (day 0 post-supplementation, PS) to the end of the study. Probiotics were delivered to rats in indicated groups as a suspension in 1 ml of distilled water delivered through oral gavages. The isolated trypanosomes were multiplied in a donor rat, and on day 7 post-supplementation (that is day 7 of ongoing supplementation with probiotics), rats in groups A to D were challenged intraperitoneally with approximately 1x 106Trypanosoma brucei suspended in phosphate buffered saline. Supplementation with MP continued in groups A to C till the end of the experiment.
Assays, bodyweights and survivability scores
From 48 h post-inoculation, the challenged groups were monitored for establishment of the infection using wet mount technique. Briefly, a drop of peripheral blood, taken from a tail tip snip, was let unto a clean glass slide and covered with a cover slip to make a wet mount preparation for examination under light microscope. Parasitaemia was estimated using rapid matching technique (Herbert and Lumsden 1976). On days 0, 7 and 16 post-supplementation, approximately 1.5 ml of blood was collected from the retrobulbar plexus of the eye of rats into plain Eppendorf tubes and, after clotting, was centrifuged at 3000 rpm for 5 min to separate the serum for biochemical assays. The activities of serum alanine aminotransferase (ALT) and aspartate aminotransferase (AST) were determined spectrophotometrically (Reitman and Frankel 1957) using commercially available kits (Randox®, UK). The AST-ALT ratio was computed on days 0, 7 and 16 post-supplementation for further evaluation of liver function (Gong et al. 2015). The computation was done by dividing the serum AST value by the corresponding ALT value for each subject. The body weights of rats in experimental groups were measured using electronic weighing balance (Mettler Toledo®, USA) on days 0, 14 and 21 post-supplementation. The survivability scores [(number of surviving animals in a group/total number of animals in a group) × 100] of the experimental groups were computed after noting the occurrence of mortalities in the groups.
Data analysis
Data from biochemical assays and body weight monitoring were analysed using Fisher’s analysis of variance (ANOVA) or Welch’s ANOVA where heteroscedasticity was to be accommodated. Variations in means were separated using the least significant difference (LSD) at post hoc analysis. The survivability scores of rats in experimental groups were analysed using descriptive statistics and presented with a chart. Test for correlation between logarithmic parasitaemia levels and the AST-ALT ratio was determined using Pearson’s product moment correlation. The analyses were done using computer package SPSS version 16.0. Significance was accepted at p < 0.05. The results were presented as means ± standard deviations using tables.
Results
The baseline values (day 0 post-supplementation) of the serum alanine and aspartate aminotransferases and the AST-ALT ratios were similar and unremarkable across all the groups (Tables 1 and 2, Fig. 1). On day 7 post-supplementation (PS), group A had significantly (p < 0.05) higher mean serum alanine aminotransferase (ALT) than all the other experimental groups (Table 1). Infection with Trypanosoma brucei established successfully in groups A–D within 4 days post-inoculation. By day 16 PS, group E (uninfected untreated) had significantly lower [Levene’s statistics (df 4, 25) = 3.945; Sig 0.013; Welch’s F(df 4, 10.179) = 747.377; p < 0.01] ALT activity when compared to those of other experimental groups, and there were no significant (p < 0.05) variations in mean ALT activity between the infected control (group D) and groups treated with probiotics (groups A, B and C) (Table 1).

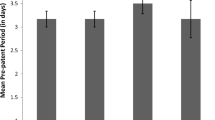
The AST-ALT ratio on days 0, 7 and 16 post-supplementation of rats treated with multistrain probiotics and infected with Trypanosoma brucei. A, infected +1 billion CFU multistrain probiotics; B, infected +5 billion CFU multistrain probiotics; C, infected +10 billion CFU multistrain probiotics; D, infected + untreated; E, uninfected + untreated. For day 0, F(df 4,25) = 0.4, p = 0.807; for day 7, F(df 4,25) = 3.259, p = 0.028; for day 16, F(df 4, 25) = 234.241, p < 0.01
There was no significant variation [F(df 4, 25) = 1.598; Sig. = 0.206] in mean serum AST values across all experimental groups on day 7 PS (Table 2). This was followed by a sharp fall in AST values on day 16 PS. Group E (uninfected untreated) had significantly higher [Levene’s statistic (df 4, 25) = 8.978, Sig. < 0.01; Welch’s F(df 4, 11.145) = 72.403, p < 0.001] mean AST values compared to all other experimental groups on day 16 PS (Table 2).
On day 7 post-supplementation, the uninfected control (group E) had significantly [F(df 4, 25) = 3.259; p = 0.028] higher mean AST-ALT ratio when compared to probiotic-treated groups A and C; however, the ratio was similar to those of groups B and D (Fig. 1). By day 16 PS, all infected groups (groups A–D) had similar mean AST-ALT ratios which were significantly [F(df 4, 25) = 234.24; p < 0.01] lower than that of the infected control (Fig. 1). A moderately strong negative correlation was seen between the equivalent logarithm number of trypanosomes per ml of blood and the AST-ALT ratio on day 16 post-supplementation. The correlation was, however, not statistically significant (Pearson’s correlation coefficient = − 3.90; p = 0.059) (Fig. 2).

Scatter plot showing correlation between Log10 number of trypanosomes/mL of blood and the AST-ALT ratio on day 16 post-supplementation in rats treated with a multistrain probiotics and infected with Trypanosoma brucei. Pearson product moment correlation coefficient = − 0.39; p = 0.059
There was no statistically significant variation (p < 0.05) in mean body weight of the experimental groups on days 0, 14 and 21 post-supplementation (Table 3). However, the uninfected control (group E) gained more weight when compared with the infected groups. On day 17 PS, groups A (infected +1Billion CFU of Probiotics), B (infected +5Billion CFU of Probiotics) and C (infected +10Billion CFU of Probiotics) had a percentage survivability score of 66.67%, while groups D and E had 100% survivability score (zero mortality) (Fig. 3). By day 24 PS, while group B had 0% survivability score (100% mortality), groups A, C and D had a survivability score of 16.67%. However, by day 27 post-supplementation, all the infected groups (groups A–D) recorded zero survivability score (100% mortality) (Fig. 3).

Percentage survivability scores of rats treated with a multistrain probiotics and infected with Trypanosoma brucei
Discussion
The aim of supplementation with the multistrain probiotics prior to infectious challenge with T. brucei was to single out the effect of the treatment on the functional integrity of the liver. It was observed that supplementation with the multistrain probiotic mix in naïve rats resulted in significant elevation of activities of serum alanine aminotransferase (ALT), although not in a dose response fashion. There was about 20% increase in serum ALT activity in a probiotic-treated group when compared to the control. Although elevated activities of ALT and aspartate aminotransferase (AST) are thought to result mainly from leakage of the enzymes from damaged hepatocytes into the extracellular fluid (Zangeneh et al. 2018), further evidences from the current study do not corroborate the existence of supplementation-induced liver damage in the rats considering that both the treatment groups and the control group had similar AST and AST-ALT ratios. The AST-ALT ratio can be used as a more robust indicator of severity of liver damages, and for differentiating aetiologies of hepatopathies. For instance, in human patients, the ratio is usually < 1 in acute and chronic hepatitis but usually increases to > 1 when fibrosis ensues, and > 2 in alcoholic hepatopathy (William and Hoofnagle 1988; Ohgo et al. 2009). In rats, elevated AST-ALT ratio in comparison to a control group indicates severe liver damage (Gong et al. 2015). It is conceivable therefore that the reported elevation in serum ALT activities in the probiotic-treated groups may not be due to liver damage. Another possible cause of elevated serum ALT activity is from ALT produced in vivo by exogenous bacteria (such as probiotics) during metabolism (Jing and Zhang, 2011); however, such causal relationship was not tested in the current study.
Following inoculation of test rats with Trypanosoma brucei, the infection established successfully within few days. Trypanosoma brucei is a remarkably tissue invasive trypanozoon—a feature that enables it to cause severe liver pathologies (Uilenberg, 1998; Federico et al. 2016). By day 16 of supplementation, ALT activities had peaked in all infected groups, suggesting the presence of liver injury. This is the typical clinicopathologic picture seen during progression of trypanosomosis in domestic and laboratory animal species (Akpa et al. 2008; Okolo et al. 2019a). The data obtained did not indicate that treatment with the probiotics elicited any form of hepatoprotective effect in the trypanosomic rats. In fact, when the probiotic-treated groups were compared to the infected control, their serum ALT and AST activities were similar irrespective of the dosage administered and did not compare favourably to those of the uninfected control.
Remarkably, the mean serum AST activities fell sharply following the establishment of the infection in all infected groups. It is very unlikely that the low AST activities observed in this study was induced by treatment with the multistrain probiotics bearing in mind that the infected control group, similarly, had very low serum AST activity. It is probable, therefore, that the decline was a trypanosomosis-induced clinical pathology which the probiotics treatment failed to reverse to normal. More often than not, trypanosomosis is characterized by elevated AST activity, and in fact, only few reports (Sow et al. 2014; Nongo et al. 2015) exist where trypanosomosis was characterized by decline in serum aspartate transaminase activity in animals as was seen from this study. The basis for low activities (below reference ranges) of ALT or AST in vertebrates remains unclear, and its association with any defined disease state is still inconclusive. However, in haemodialysis patients, some workers have attributed this condition to vitamin B6 deficiency precipitated by the existing disease condition (Ono et al. 1995).
After establishment of the infection, the AST-ALT ratio followed similar pattern as the serum AST activity, where all the infected groups, with or without probiotics treatment, had low and similar AST-ALT ratios. This shows that depressed AST levels had an overwhelming effect on the ratio, thereby minimizing its conventional diagnostic usefulness. When the logarithmic parasitaemia levels were plotted against the AST-ALT ratio, a random moderately strong, negative correlation was observed. The two possible explanations to this phenomenon is either that rising parasitaemia levels do not directly correlate with increasing liver pathology in trypanosomosis, or that simply the remarkably depressed AST component of the ratio successfully distorted the picture. Nonetheless, the AST-ALT ratio versus parasitaemia correlation was not statistically significant.
It has been opined that consumption of probiotics may improve the nutritional value of diets through enhanced digestion of feed (Gogineni et al. 2013), which would translate to improved energy balance, body condition and weight gain. Nonetheless, no significant variation in body weight was observed among the probiotic-treated groups and the controls. Positive effect on weight gain or loss was simply not there or may have required a longer period of administration of the probiotics in order to be seen. Such long period of supplementation was practically impossible in the present study considering that full blown trypanosomosis in rat models can invariably lead to a hundred percent mortality within 1 month. Treatment with the probiotic strains did not improve the survivability or overall health of the infected groups; in fact, by day 27 post-supplementation, all the infected groups recorded a hundred percent mortality.
The study shows that trypanosomic rats did not benefit meaningfully from probiotic supplementation and treatment with respect to serum ALT and AST activities, weight gain and survivability. This finding is similar to the report of Firouzi et al. (2015) who demonstrated that following treatment with multistrain probiotic mix containing Lactobacillus acidophilus, Lactobacillus casei, Lactobacillus lactis, Bifidobacterium bifidum, Bifidobacterium longum and Bifidobacterium infantis no significant improvement were observed in ALT and AST activities in diabetic patients. A similar study by Asemi et al. (2015) also reported non-improvement in liver function parameters following probiotic treatment in diabetic patients. However, several other reports (Kirpich et al. 2008; Khalesi et al. 2018) of improvement in liver function indicators following treatment with probiotics contrast our findings. The major reason for this contrast may be difference in strains of probiotic organisms administered in the various studies. It is known that the bioactivities of probiotics are strain specific (Gogineni et al. 2013; Okolo et al. 2020), which implies that the same biological effect may not be elicited by probiotic organisms belonging to different strains although they share the same genus and species. The strains employed for this study may simply not boast any hepatoprotective activity, whereas they may be useful in other conditions. In addition, probiotic products are usually dispersed in inactive excipients (Good and Wu 2017; Okolo et al. 2019b), and Khalesi et al. (2018) showed that inclusion of prebiotics (non-digestible dietary supplements) in the probiotic-excipient mix may enhance the bioactivity of the probiotics by providing fermentable nutrients for multiplication of probiotic strains in vivo. The probiotic preparation used in the current study was void of prebiotics, and it therefore remains to be verified whether the presence of prebiotics could make a desirable difference in a study such as this. Further studies towards developing specific probiotic strains with activity on the gut-liver axis, and towards exploring probiotic and prebiotic combinations will be worthwhile.
Conclusion
In summary, while supplementation with the multistrain probiotic mix was innocuous to liver function in naïve rat models, in rats suffering trypanosomosis, its usefulness for prevention or management of liver damage with respect to serum AST and ALT activities, AST-ALT ratio or the improvement of body weight and survivability was clearly absent.
References
Abenga J, Idowu T (2018) Packed cell volume and bone marrow cytological responses in Trypanosoma vivax- induced acute trypanosomosis in Nigerian Sokoto Red goats. Comp Clin Pathol 27:455–460
Akpa PO, Ezeokonkwo RC, Eze CA, Anene BM (2008) Comparative efficacy assessment of pentamidine isethionate and diminazene aceturate in the chemotherapy of Trypanosoma brucei brucei infection in dogs. Vet Parasitol 151:139–149
Ali S, Batool S, Butt FJ, Nasreen S, Tahir HM (2019) Hepatocellular carcinomas risk factors, diagnosis, prevention and frontier to probiotic treatment. Punjab Univ J Zool 34:149–158
Aneke CI, Ugochukwu CI, Kalu I, Ugochukwu IE (2016) Comparative efficacy of graded doses of diminazene aceturate and fixed doses of iron dextran and vitamin B complex in mice infected with Trypanosoma brucei. Comp Clin Pathol 25:933–939
Anosa VO (1988) Haematological and biochemical changes in human and animal trypanosomiasis II. Rev Elev Med Vet Pays Trop 41:151–164
Aronu CJ, Okolo CC, Akubueze H (2019) Dietary supplementation with aqueous extract of talinum triangulare at different phases of broiler production and effect on growth, serum biochemistry and carcass quality. Indian J Anim Res. https://doi.org/10.18805/ijar.B-1156
Asemi Z, Bahmani S, Shakeri H, Jamal A, Faraji A (2015) Effect of multispecies probiotics supplements on serum minerals, liver enzymes and blood pressure in patients with type 2 diabetes. Int J Diabetes Dev Ctries 35:90–95
Bautista-Garfias-Garfias CR, Alvarez M, Martínez-Gómez F (2008) The inoculation of lactobacillus casei in nih mice induces a protective response against Trypanosoma cruzi (Ninoa strain) infection. Vet Mex 39:19–144
Engen PA, Green SJ, Voigt RM, Forsyth CB, Keshavarzian A (2015) The gastrointestinal microbiome: alcohol effects on the composition of intestinal microbiota. Alcohol Res Curr Rev 37:223–236
Eze JI, Orajaka LJE, Okonkwo NC, Ezeh IO, Ezema C, Anosa GN (2012) Effect of probiotic (Saccharomyces cerevisiae) supplementation on immune response in Trypanosoma brucei brucei infected rats. Exp Parasitol 132:434–439
Federico A, Tuccillo C, Grossi E, Abbiati R, Garbagna N, Romano N, Tiso A, Del Vecchio BC, Loguercio C (2016) The effect of a new symbiotic formulation on plasma levels and peripheral blood mononuclear cell expression of some pro-inflammatory cytokines in patients with ulcerative colitis: a pilot study. Eur Rev Med Pharmacol Sci 13:285–293
Firouzi S, Mohd-Yusof BN, Majid HA, Ismail A, Kamaruddin NA (2015) Effect of microbial cell preparation on renal profile and liver function among type 2 diabetics: a randomized controlled trial. BMC Complement Altern Med 15:1–10
Gogineni VK, Morrow LE, Gregory PJ, Malesker MA (2013) Probiotics: history and evolution. J Anc Dis Prev Rem 1:107–114
Gong XW, Xu YJ, Yang QH, Liang YJ, Zhang YP, Wang GL, Li YY (2015, 2015) Effects of soothing liver and invigorating spleen recipes on the IKKB –NF-kB signaling pathway in Kupffer cells of non-alcoholic steatohepatitis rats. Evid Based Complement Alternat Med:687690. https://doi.org/10.1155/2015/687690
Good D, Wu Y (2017) Excipient characterization. In: Koo OMY (ed) Pharmaceutical excipients. John Wiley and Sons Inc., New Jersey, pp 20–51
Hajifaraji M, Jahanjou F, Abbasalizadeh F, Aghamohammadzadeh N, Abbasi MM, Dolatkhah N (2018) Effect of probiotic supplements in women with gestational diabetes mellitus on inflammation and oxidative stress biomarkers: a randomized clinical trial. Asia Pac J Clin Nutr 7:581–591
Herbert WJ, Lumsden WHR (1976) Trypanosoma brucei: a rapid “matching” method for estimating the host’s parasitemia. Exp Parasitol 40:427–431
Jing X, Zhang S (2011) An ancient molecule with novel function: alanine aminotransferase as a lipopolysaccharide binding protein with bacteriocidal activity. Dev Comp Immunol 35:94–104
Khalesi S, Johnson DW, Campbell K, Williams S, Fenning A, Saluja1 S, Irwin C, (2018) Effect of probiotics and synbiotics consumption on serum concentrations of liver function test enzymes: a systematic review and meta-analysis. Eur J Nutr 57: 2037–2053
Kirpich IA, Solovieva NV, Leikhter SN, Shidakova NA, Lebedeva OV, Sidorov PI, Bazhukova TA, Soloviev AG, Barve SS, McClain CJ, Cave M (2008) Probiotics restore bowel flora and improve liver enzymes in human alcohol-induced liver injury: a pilot study. Alcohol (Fayetteville, NY) 42:675–682
Land MH, Rouster-Stevens K, Woods CR, Cannon ML, Cnota J, Shetty AK (2005) Lactobacillus sepsis associated with probiotic therapy. Pediatrics 115:178–181
NIH (2011) Guide for the care and use of laboratory animals. National Academy Press, Washington DC, pp 11–30
Nongo NN, Tion MT, Apaa TT, Ogunro BN (2015) A case of canine trypanosomosis with epistaxis in a two year old alsatian dog. IOSR J Agric Vet Sci 8:68–72
Ohgo H, Yokoyama H, Hirose H, Kawabe H, Saito I, Tomita K, Hibi T (2009) Significance of ALT/AST ratio for specifying subjects with metabolic syndrome in its silent stage. Diabe Metab Syndrom Clin Res Rev 3:3–6
Okolo CC, Eze JI, Nweze NE (2020) Hematobiochemical and immunological responses of rats treated with multi-strain probiotics and infected with Trypanosoma brucei. Probiot Antimicro Prot 12:952–960
Okolo CC, Ezeh IO, Uju CN, Nweze NE (2019a) Combination of a probiotic mix and diminazene aceturate in treatment of Trypanosoma brucei infection in Sprague Dawley rats. Vet Sci: Res Rev 5:43–52
Okolo CC, Nweze NE, Okonkwo IA, Omeje OV, Eke IG (2019b) The pharmaceutical excipient, magnesium stearate, depresses lymphocyte counts in vivo but does not lower humoral immune response in Sprague-Dawley rats. EAS J Agric Life Sci 2:317–322
Omeje VO, Okolo CC, Eze DC (2019) Pathogenicity (LD50) and antibiotics sensitivity tests of Aeromonas hydrophila isolated from fishes of the Kainji Lake area. Int J Fish Aquat Stud 7:190–196
Ono K, Ono T, Matsumata T (1995) The pathogenesis of decreased aspartate aminotransferase and alanine aminotransferase activity in the plasma of hemodialysis patients: the role of vitamine B6 deficiency. Clin Nephrol 43:405
Reitman S, Frankel S (1957) Serum transaminase levels. Am J Clin Pathol 28:56–63
Sharma V, Garg S, Aggarwal S (2013) Probiotics and liver disease. Perm J 17:62–67
Sow A, Sidibe I, Kalandi M, Bathily A, Ndiaye NP, Ouedraogo M, Mouiche MM, Sawadogo GJ (2014) Biochemical changes induced by natural infection of trypanosomosis in Burkinabese local donkey breeds. Comp Clin Pathol 23:103–104
Uilenberg G (1998) A field guide for the diagnosis, treatment and prevention of African animal trypanosomosis. Food Agriculture Organization (FAO), Rome, pp 12–60
Villarino NF, LeCleir GR, Denny JE, Dearth SP, Harding CL, Sloan SS, Gribble JL, Campagna SR, Wilhelm SW, Schmidt NW (2016) Composition of the gut microbiota modulates the severity of malaria. PNAS 113:2235–2240
Williams ALB, Hoofnagle JH (1988) Ratio of serum aspartate to alanine aminotransferase in chronic hepatitis. Gastroenterology 95:734–739
Yaro M, Munyard KA, Stear MJ, Groth DM (2016) Combatting african animal trypanosomiasis (AAT) in livestock: the potential role of trypanotolerance. Vet Parasitol 225:45–52
Zangeneh MM, Zangeneh M, Tahvilian R, Moradi R, Zhaleh H, Amiri-paryan A, Bahrami E (2018) Hepatoprotective and hematoprotective effects of Falcaria vulgaris aqueous extract against CCl4-induced hepatic injury in mice. Comp Clin. Path 27:1359–1365
Acknowledgements
We are grateful to the staff of Veterinary Medicine Laboratory, Department of Veterinary Medicine, University of Nigeria, for technical support in the course of this work.
Availability of data and material
Primary data supporting the findings of this research are available from the corresponding author upon request.
Funding
The research was self-funded.
Author information
Authors and Affiliations
Contributions
Besides initial drafting of the manuscript which was done by Chukwuemeka Calistus Okolo, all the authors contributed equally to the design and execution of the research, data analysis and review of final manuscript.
Corresponding author
Ethics declarations
Conflict of interests
The authors declare that they have no conflict of interests.
Ethical approval
All institutional and national guidelines for the care and use of laboratory animals were followed. Ethical considerations for use of the experimental animals in this study were based on the guidelines of the Animal Use and Care Committee of the Faculty of Veterinary Medicine, University of Nigeria, which agrees with the NIH procedures (NIH, 2011).
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Okolo, C.C., Uju, C.N., Chibuzor, S.D. et al. The effects of treatments with multistrain probiotics on serum aminotransferases, AST-ALT ratio, body weight and survivability scores of rats experimentally infected with Trypanosoma brucei. Comp Clin Pathol 29, 1229–1236 (2020). https://doi.org/10.1007/s00580-020-03175-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00580-020-03175-z