
Abstract
Mycorrhizal symbioses (mycorrhizas) of Ericaceae, including ericoid mycorrhiza (ErM), have been mainly studied in the Northern Hemisphere, although the highest diversity of ericaceous plants is located in the Southern Hemisphere, where several regions remain largely unexplored. One of them is South America, which harbors a remarkably high diversity of Ericaceae (691 species and 33 genera) in a wide range of environmental conditions, and a specific mycorrhizal type called cavendishioid. In this review, we compile all available information on mycorrhizas of Ericaceae in South America. We report data on the mycorrhizal type and fungal diversity in 17 and 11 ericaceous genera, respectively. We show that South American Ericaceae exhibit a high diversity of habitats and life forms and that some species from typical ErM subfamilies may also host arbuscular mycorrhiza. Also, a possible geographical pattern in South American ErM fungal communities is suggested, with Sebacinales being the dominant mycorrhizal partners of the Andean clade species from tropical mountains, while archetypal ErM fungi are common partners in southern South America species. The gathered information challenges some common assumptions about ErM and suggests that focusing on understudied regions would improve our understanding of the evolution of mycorrhizal associations in this intriguing family.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Mycorrhizas are symbiotic associations between most land plants and specific fungal taxa (Smith and Read 2008). The mycorrhizal association is estimated to be present in more than 90% of plant species and is distributed in almost all terrestrial ecosystems (Brundrett and Tedersoo 2018; van der Heijden et al. 2015). In this interaction, plants provide carbon to the fungal partners, which in exchange deliver soil nutrients to plants, among other non-nutritional benefits (Delavaux et al. 2017). Depending on their function, structure, and on plant and fungal identity (Genre et al. 2020; Smith and Read 2008), mycorrhizas are classified into four types: arbuscular mycorrhiza (AM), ectomycorrhiza (EM), and ericoid (ErM) and orchid mycorrhizas (OrM). AM is the most ancestral type (Maherali et al. 2016; Strullu-Derrien et al. 2018), in which fungi from the Glomeromycetes colonize the roots of 80% of plant species, forming intracellular structures such as arbuscules or coils (Smith and Read 2008). EM symbiosis, on the other hand, evolved independently several times along plant and fungal evolution (Martin et al. 2016; Tedersoo and Brundrett 2017). This association involves mostly trees that dominate woodland and forest communities in boreal, Mediterranean, and temperate regions and some tropical forests. EM fungi belong to Basidiomycota, Ascomycota, and more rarely Zygomycota (Tedersoo et al. 2010), and form a hyphal sheath around root tips and penetrate the intercellular space forming a network between cortical cells called Hartig net (Smith and Read 2008). Finally, the last, most recent mycorrhizal types in plant evolution are OrM and ErM (Strullu-Derrien et al. 2018). In both cases, fungal partners form hyphal coils within root cells of orchids and Ericaceae species, respectively, but beyond penetration of the cell wall, the cell membrane remains intact and allows exchanges. Whereas orchid mycorrhizas are present in almost all Orchidaceae (Selosse et al. 2022), representing 10% of vascular plant species, ErM are present in some Ericaceae that account for only 1.5% of vascular plants (Brundrett and Tedersoo 2018). Despite this relatively low plant diversity, ErM occupy a broad range of ecosystems, from heathlands of the Northern Hemisphere, open forests, Mediterranean woodlands, and tropical cloud forests, to the dry sand plains of Australia (Cairney and Meharg 2003). Moreover, ErM plants colonize ecosystems estimated to hold more than 20% of the terrestrial carbon stock, playing a pivotal role in the soil carbon cycle (Perotto et al. 2018). Yet, ErM remains the least studied and the least understood type of mycorrhizal association (Vohník 2020).
The Ericaceae family is highly diverse in terms of its mycorrhizal associations (Lallemand et al. 2016): while the earliest-diverging genus Enkianthus retained the ancestral AM (Obase et al. 2013), the subfamilies Monotropoideae and Arbutoideae form ectendomycorrhizas (a type of EM with intracellular colonization), and the subfamilies Ericoideae, Cassiopoideae, Harrimanelloideae, Styphelioideae, and Vaccinioideae, a monophyletic clade known as the “early anther inversion clade” (EAI clade; Kron et al. 2002), form ErM (Selosse et al. 2007).
The ericoid mycorrhizal fungi (ErMF) include fungi from the Ascomycetes—typically species from Hyaloscypha s. str. (formerly known as the Hymenoscyphus ericae aggregate) and Oidiodendron maius—and from the Basidiomycetes—including mostly taxa from the Serendipitaceae (Vohník et al. 2016; Weiss et al. 2016). However, the range of potential ErMF is much broader than previously believed (Vohník 2020), arising questions about how accurately our current knowledge of ErM symbiosis reflects its true diversity (Vohník et al. 2023). ErMF form hyphal coils in the rhizodermal cells of the fine roots (also called hair roots) of Ericaceae, but some of them are also common root endophytes in non-ericaceous plants (Almario et al. 2017; Weiss et al. 2016), i.e., colonize the tissues biotrophically and diffusely without causing any morphological symptom. In addition, as shown by their in vitro cultivability and genomic features (Perotto et al. 2018), many ErMF have strong saprobic capabilities, in contrasts to AM and most EM fungi (Miyauchi et al. 2020), and some also grow as free-living decomposers (Rice and Currah 2006).
ErM plants are frequently characterized as colonizers of infertile and acidic soils, with a high content of recalcitrant polyphenolic compounds and slow decomposition of soil organic matter (Perotto et al. 2018; Van Geel et al. 2020). On the other hand, most of the ErM research has focused on a limited number of species found in cold habitats primarily in the Northern Hemisphere (Albornoz et al. 2021; Vohník et al. 2023), while the highest diversity of Ericaceae plants is actually found in the Southern Hemisphere, in tropical, subtropical, and Mediterranean regions, where ErM interactions are highly understudied (Albornoz et al. 2021). For example, while the vast majority of species of Erica, the largest genus in Ericaceae, are restricted to the Cape Floristic Region of South Africa (Linder 2003), little is known about their mycorrhizal associations in this region (Kohout and Tedersoo 2017).
South America is one of the regions where mycorrhizal associations remain largely unexplored, despite its vast plant and fungal diversity and endemism (Mujica et al. 2019; Bueno et al. 2017). It boasts four notable centers of Ericaceae diversity (Luteyn 2002), yet our understanding of its mycorrhizal associations in these habitats remains limited. This is at least in part because information published in Spanish and Portuguese in local journals and databases is typically not included in global compilations. Indeed, non-English literature may contain unique knowledge and ignoring it introduces biases in global literature reviews (Konno et al. 2020; Nuñez and Amano 2021). Hence, here we present the first review of existing knowledge about mycorrhizal associations of Ericaceae in South America (for previous local efforts, see Pacheco Flores et al. 2022). Our aim was to collect information about the mycorrhizal type and the diversity of fungal species associated with South American ericaceous species. Doing so, we hope to contribute to a broader understanding of the mycorrhizal associations in Ericaceae on a global scale.
Diversity of Ericaceae in South America
Ericaceae is one of the most diverse families of the Angiosperms, with ca. 4500 species globally ranging from sea level to high mountains over 5000 m (POWO 2023). The most recent classification system of Ericaceae comprises eight sub-families, and 20 tribes with 121 genera (POWO 2023; Kron et al. 2002). The family is distributed worldwide, especially in tropical and temperate regions, except for Antarctica (Kron and Luteyn 2005). The radiation of Ericaceae is mostly of boreotropical with subsequently more recent diversification in tropical Asia, Southern Africa, and tropical South America (Kron and Luteyn 2005). This radiation is probably associated with the availability of oligotrophic habitats, linked to the evolution of low specific leaf areas and specialized mycorrhizal associations (Schwery et al. 2015). The areas of higher species richness are localized in the montane tropics of South America, Asia, New Guinea, Australia, and Africa (Kron and Luteyn 2005). The members of this family grow in a wide range of habitats and with diverse life history strategies ranging from shrubs and trees to various often ignored kinds of epiphytes: climbing epiphyte, climbing shrub, epiphytic shrub, scrambling epiphyte, scrambling shrub, and true lianas (POWO 2023; Kron et al. 2002). It also includes achlorophyllous, mycoheterotrophic species relying on their mycorrhizal fungi for carbon nutrition (Merckx 2013).
In South America, the Ericaceae are well represented with 691 species (POWO 2023), mostly belonging to the subfamilies Ericoideae, Styphelioideae, and Vaccinioideae in the EAI clade and one species of the subfamily Monotropoideae (Fig. 1). From the 121 genera of Ericaceae, 33 are native (27%), and 15 are found exclusively in South America (12.4%). The endemic genera include four monotypic genera from Chile and Argentina (Lebetanthus), Ecuador (Periclesia), and Bolivia (Polyclita and Rusbya) (Table 1). Colombia (277 spp.), Ecuador (229), and Peru (154) contribute to most of the species’ diversity, with approximately 320 species representing about 45% of the total existing in South America (Table 1). The largest genera, Cavendishia (83), Thibaudia (78), Psammisia (59), Gaultheria (57), and Gaylussacia (48), encompass nearly 50% of species found in South America. Luteyn (2002) identified four biogeographical regions or centers of diversity of Ericaceae in South America: the tropical Andes, the Guyana Highlands, Southeast Brazil, and temperate Andes.

Phylogeny of the Ericaceae subfamilies with the most parsimonious scenario for the evolution of mycorrhizal types (according to Freudenstein et al. 2016 and Lallemand et al. 2016) and their global distribution (following Kron and Luteyn 2005). The tribes Pyroleae and Monotropeae are included in the subfamily Monotropoideae, but according to Freudenstein et al. (2016) and Schwery et al. (2015), Pyroleae is a sister group to Monotropeae + Arbutoideae. AM, the ancestral arbuscular mycorrhiza; CVM, cavendishioid mycorrhizas (a type of EEM); EEM ectendomycorrhizas; ErM ericoid mycorrhizas. Note that the acquisition of ErM corresponds to the rise of the “early anther inversion clade” (EAI clade; Kron et al. 2002). AM reports are not necessarily functional, further research is needed to confirm their functionality in ericaceous roots. The geographical distribution of subfamilies is presented alongside each one, with the same corresponding color. Subfamilies distributed in South America are underlined
The greatest diversity of all described neotropical Ericaceae is found in the tropical Andean region, with more than 500 species (Pedraza-Peñalosa and Luteyn 2015). These plants have adapted to the moist and cool environments at altitudes between 1000 and 3000 m above sea level (Setaro et al. 2006a; Pacheco Flores et al. 2022). 95% of them belong to the subfamily Vaccinioideae, tribe Vaccinieae, and form the monophyletic clade known as the “Andean clade” (Kron et al. 2002). Biogeographical analysis indicates that the ancestor to the Andean + Mesoamerican-Caribbean groups may have been widespread in the Andes and the mountains of Central America and the Antilles (Kron and Luteyn 2005). The diversification within the Andean clade has been indicated as very recent (since the late Miocene) and the taxa currently found in Central America have recently arrived through dispersal from the northern Andes (Kron and Luteyn 2005). However, recent studies have shown that the diversity of Ericaceae in montane habitats is the result of a higher diversification rate rather than dispersal from non-montane habitats (Schwery et al. 2015). Thus, the Andean orogeny may account for the current distribution patterns of many species-rich genera within Ericaceae in South America. This biogeographical event is among the most significant factors for radiation of vascular plants and the biogeographic history of neotropical species in South America (Luebert and Weigend 2014; Antonelli et al. 2009). Although it has often been suggested that mycorrhizal associations have been crucial for the diversification of Ericaceae, this has not been evaluated in South America. This region, where diversification events are associated with the formation of new habitats, allows to test for a key role of mycorrhizal associations.
To summarize, South America provides habitats different from those commonly used to investigate ErM. Furthermore, the wide variety of life forms of Ericaceae in South America contrasts with the 85% of shrubs reported to ErM plants from global compilations (Soudzilovskaia et al. 2020). This diversity of life forms and habitats positions South America as a highly important location for studying the functioning of ErM, to enhance current knowledge extrapolated from shrubs in cold climates of the Northern Hemisphere.
Mycorrhizal types of South American Ericaceae
We performed a systematic review to gather all available scientific information (to our best knowledge) about the mycorrhizal associations of South American ericaceous species. For each of the 33 genera native to South America, we conducted thorough searches in multiple scientific databases (Google Scholar, Web of Science, and SciELO) using the keywords “mycorrhiza,” “ericoid,” “colonization,” and “fungi” along with the specific genus name. Additionally, we expanded our search by including the name of each South American country along with the same keywords to maximize article findings. Most importantly, we performed the same search using the keywords in Spanish and Portuguese. Subsequently, we critically reviewed each publication to keep only those that examined mycorrhizal types of Ericaceae species native to South America or the identity of their fungal partners, excluding non-native species within native genera. Our collection of articles considered works written in English (64%), Spanish (16%), and Portuguese (20%; Table S1). Additionally, we included theses that were available online and provided complete information about methods and observations.
Our search yielded information on the mycorrhizal type for 51% (n = 17) of the genera present in South America, while no information was found for the remaining 16 genera (Table 1). Based on our findings, Ericaceae species in South America exhibit cavendishioid (CVM), ericoid (ErM), monotropoid, and arbuscular mycorrhizal (AM) associations. These results partially agree with the phylogenetic pattern of mycorrhizal types observed in the Ericaceae family, with the subfamily Ericoideae displaying ERM, while the subfamily Vaccinioideae showcases both ERM and CVM associations (Lallemand et al. 2016; Freudenstein et al. 2016). However, we also found several reports of AM associations in Vaccinioideae species, which agrees with reports of AM in this subfamily in other regions (see below).
AM in ericoid mycorrhizal subfamilies
We found literature suggesting the potential colonization of AM in five genera of Vaccinioideae (Agarista, Gaultheria, Gaylussacia, Psammisia, and Vaccinium) in South America. Although this subfamily is commonly referred to as ErM (e.g., Kohout 2017; Lallemand et al. 2016), several studies have reported the presence of AM in some of its species. For example, Koske et al. (1990) showed the presence of spores, vesicles, and arbuscules in the roots of Vaccinium calycinum, V. dentatum, V. reticulatum, and Styphelia tameiameiae from Hawaii, in addition to the presence of typical ErM structures (see Figs. 1–4 and 5–7, pages 65–66). Similarly, in addition to ErM, the shrub Vaccinium oldhamii exhibited AM fungal mycelium and structures like vesicles and arbuscules in roots, and the presence of the AM fungus Rhizophagus diaphanous was detected (Baba et al. 2016; see Fig. S3). Likewise, Lancheros (2012; see Fig. 28, page 50) found AM and ErM structures in V. meridionale in Colombia, while Santos et al. (1995, no photos) and Baião and Kasuya (2007, no photos) detected AM associations in Brazilian Agarista spp. Similarly, John (1980, no photos) reported AM structures in Satyria sp. in Brazil. In addition to ErM, AM associations were also observed in Gaultheria poeppigi in southern South America (Urcelay 2002, see Fig. 1, page 90; Menoyo et al. 2007, no photos) and in G. myrsinoides from the Colombian Paramo (Guerrero 1996, no photos). The same dual mycorrhizal colonization was observed in Indian G. fragantissima by Das and Kayang (2012, see Fig. 1, page 138). On the contrary, AM structures were not detected in five Gaultheria species from Patagonian and temperate Valdivian Forest in Argentina by Vohník et al. (2023).
The ability to form AM in addition to ErM appears to be extended to other subfamilies of the EAI clade that are thought to be exclusively ErM. For example, in Ericoideae, Empetrum nigrum var. japonicum and Ledum palustre var. diversipilosum possessed hyphae and vesicles of AM fungi (AMF) in addition to ErM in the crater basin of Volcano Esan in Hokkaido, Japan (Fukuchi et al. 2011, see Fig. 4, page 9), and several species of Rhododendron presented AMF hyphal coils, intraradical vesicles, and chlamydospores in central Himalaya (Chaurasia et al. 2005, see Fig. 1, page 318). Similarly, in Styphelioideade Bellgard (1991) observed AM in Epacris microphylla and Leucopogon juniperus in the Hawkesbury Sandstone soils in Australia. Nevertheless, it is necessary to evaluate whether the AM-like structures observed in these ErM subfamilies are indeed functional, i.e., taking part in the mycorrhizal bi-directional nutrient transfer. In fact, non-mycorrhizal plants grown in the presence of the AM mycelium can exhibit AMF structures, frequently hyphae and vesicles, but never arbuscules (DeMars and Boerner 1996; Veiga et al. 2012, 2013; Mnasri et al. 2017). On the other hand, it is important to acknowledge the possibility of misidentifying roots from adjacent non-ericaceous AM vegetation as ericaceous roots. Such misclassification could result in misleading conclusions and deserves careful consideration.
The high number of reports of AM in the EAI clade from different sources suggests that at least some species of these subfamilies may have dual mycorrhizal capacity, like mycorrhizal plants forming EM and AM associations (Teste et al. 2020). Ancestral trait reconstruction has shown that the ancestor of Ericaceae most probably had AM (Selosse et al. 2007; Fig. 1). Enkianthus, the most basal extant ericaceous genus forms AM (Obase et al. 2013), although the possibility of also forming ErM has been proposed (Vohník 2020). Then, there was an evolutionary transition to ectendomycorrhizal associations, which are present in Monotropoideae and Arbutoideae (Richard et al. 2005; Selosse et al. 2007), and a transition to ErM in the five remaining subfamilies of EAI (Freudenstein et al. 2016; Fig. 1). Vaccinioideae, however, show an additional acquisition of CVM (Setaro et al. 2006a, b). This model suggests that AM was lost in the transition to ectendomycorrhiza (Selosse et al. 2007). However, the presence of AM in Vaccinioideae (and probably in other ErM subfamilies) suggests either a partial reversion to AM (Brundrett and Tedersoo 2018) or the persistence of an ancestral (plesiomorphic) trait that might have never been lost in Ericaceae (Freudenstein et al. 2016). Further genomic, phylogenomic, and ecological research is needed to test these hypotheses.
Cavendishioid mycorrhiza in the Andean clade
CVM was first described by Setaro et al. (2006a), who discovered a mycorrhizal association exhibiting a hyphal sheath, Hartig net, and intracellular colonization in roots of Cavendishia nobilis in the tropical mountain rainforest of South Ecuador. CVM can be classified as an independent evolution of ectendomycorrhizas, similar to the fungal associations found in the subfamily Arbutoideae. The investigated C. nobilis was associated with a remarkable diversity of Sebacinales, although to a lesser degree, some Leotiales were also detected (Setaro et al. 2006a). Subsequently, CVM was observed in 15 species belonging to nine different genera from the Andean clade (Setaro et al. 2006b; Table 1). Ericaceous species from the same habitat but not belonging to the Andean clade (i.e., Gaultheria spp., Bejaria, and Vaccinium) did not show CVM suggesting that this type of association is mostly phylogenetically constrained, rather than resulting from certain environmental conditions.
Interestingly, almost all examined species belonging to the Andean clade and presenting CVM have not been reported to form AM (Table 1), except for the genus Psammisia in which Bermudez and Benzing (1989) observed possible CVM and AM structures in one plant. In contrast, the Ericaceae species with AM structures were sampled outside the tropical Andean range, mostly in Brazil and in southern South America (Table 1). These findings show the variability of the mycorrhizas of Ericaceae in South America, as well as some endemic traits. Further research is needed to evaluate how environmental conditions and/or biogeographical and historical drivers determine the type of mycorrhizal association in Ericaceae.
Monotropoid mycorrhizas in South America?
Monotropa uniflora is a member of Monotropoideae, which comprises 15 genera and ca. 50 species. They lack chlorophyll and are mycoheterotrophic, meaning they depend on carbon compounds obtained through fungal mycelial connections with surrounding EM host plants (Merckx 2013). Monotropoideae establish a distinct type of ectendomycorrhiza called monotropoid mycorrhiza, characterized by a hyphal mantle, Hartig net, and intracellular “fungal pegs” (Massicotte et al. 2005). Monotropoid mycorrhizas display a high level of specificity, as each lineage of Monotropoideae associates with specific groups of EM fungi within single genera (Bidartondo and Bruns 2002).
Monotropoideae has a widespread distribution in temperate regions of the Northern Hemisphere, mostly found in dense, humid forests characterized by abundant organic litter and restricted light availability for autotrophic growth (Leake 1994). Monotropa uniflora is the only species of this subfamily that occurs in South America. This species is also predominantly distributed in the Northern Hemisphere, but in America, its presence aligns with the natural distribution of Pinaceae, extending from the highlands of Mexico and Central America into Belize, and further south into northern South America (Luteyn et al., 2002). In this region, it can be found in western Colombia, in native Quercus forests as well as introduced pine plantations (Luteyn 2002). Although M. uniflora is classically considered a single species along its distribution, molecular studies suggest that the populations from Asia, North America, and Central America are molecularly divergent and phylogenetically distinct (Neyland and Hennigan 2004). The South American taxon/taxa may thus belong to one or several distinct species, which remains to be assessed.
Surveys across almost all its distribution have shown that M. uniflora forms specific mycorrhizal associations with members of the genus Russula (Russulaceae; Bidartondo and Bruns 2001). Accordingly, in Abies religiosa and Pseudotsuga menziessi forests in Mexico, M. uniflora associates with several species of Russula (Kong et al. 2015). Currently, there is a lack of information regarding the mycorrhizal associations of M. uniflora in South America, where the study of mycoheterotrophy, in general, remains largely unexplored. Considering the distinct climate and floristic composition compared to the majority of its distribution, it can be expected that more tropical Russula species could be involved. Yet, the taxonomic status of M. uniflora and potential differences in mycorrhizal partners (even among Russulaceae) await further analyses in South America.
Ericoid mycorrhizal fungal diversity in South America
Globally, most of the knowledge on the ErMF diversity comes from the temperate and boreal ecosystems of the Northern Hemisphere (Vohník 2020). As stated above, ericoid mycorrhizal fungi are mainly categorized within Ascomycota and Basidiomycota (Perotto et al. 2018; Fehrer et al. 2019; Vohník 2020). Among Ascomycota, the most significant ErMF include the Hyaloscypha hepaticicola aggregate—formerly referred to as the Hymenoscyphus ericae aggregate and the Rhizoscyphus ericae aggregate (Fehrer et al. 2019), comprising H. gryndleri (Vohník et al. 2022), H. hepaticicola, and H. variabilis (Vohník et al. 2013)—and Oidiodendon maius and various Leohumicola species (Vohník 2020). Among Basidiomycota, the family Serendipitaceae in the Sebacinales (formerly Sebacinales Clade B, encompassing the single genus Serendipita; Weiss et al. 2016; Vohník et al. 2016; Selosse et al. 2007) and several non-sebacinoid fungi (Vohník et al. 2012; Kolařík and Vohník 2018) have also been defined as ErMF. However, despite the increasing identification of potential ErMF worldwide, the dominant perspective on the identity of ErMF has been focused on the ascomycetous H. hepaticicola and Oidiodendon maius (Vohník et al. 2023).
In South America, the information about the identity of mycorrhizal fungi associated with Ericaceae is very scarce, with only 11 out of 33 genera being assessed. Most of the information comes from species in the Andean clade (Setaro et al. 2006a) and from Gaultheria spp. in southern South America (Selosse et al. 2007; Bruzone et al. 2015, 2017; Vohník et al. 2023). Most identified fungi associating with Ericaceae belong to Sebacinales. Specifically, studies conducted by Setaro et al. (2006a, b, 2013), Setaro and Kron (2011), and Kottke et al. (2008) have identified several Serendipitaceae as the principal mycorrhizal partners in Ericaceae species from tropical mountain forests in Ecuador. Most of these plant species form CVM, but plants with ErM in the same habitat also showed associations with Serendipitaceae (Setaro et al. 2006b). Serendipitaceae encompasses species that form mycorrhizas with orchids (Selosse et al. 2022), grow as nonspecific root endophytic taxa (Selosse et al. 2009; Varma et al. 2012; Riess et al. 2014), and form mycorrhizal structures in a wide range of Ericaceae hosts (Selosse et al. 2007; Vohník et al. 2016; Weiss et al. 2016).
On the other hand, the diversity of ErMF found in southern South America associated with the genus Gaultheria includes both basidiomycetous and ascomyceouts ErMF. Using specific primers for Sebacinales, Selosse et al. (2007) identified Serendipita spp. as mycorrhizal partner in G. poeppiggi in the Cordoba Mountain Ranges, Argentina. Later, Bruzone et al. (2015) isolated Sebacinales from Gaultheria spp. roots, but they were not able to isolate H. hepaticicola, and the authors suggested this prominent ErMF could be absent in the Southern Hemisphere (Bruzone et al. 2015). However, in a subsequent study Bruzone et al. (2017) isolated the commonly reported ErMF H. hepaticicola and H. bicolor. In addition, Geml et al. (2014) detected the presence of potential ErMF (Helotiales and Sebacinales) in soils of the subtropical Andean forests (1500–3000 m a. s. l.). Recently, species of Gaultheria from different ecosystems in Patagonia revealed a diverse fungal community, including various species with uncertain symbiotic status (Vohník et al. 2023) strongly different from ErMF from Northern Bohemia (Central Europe), and strong differentiation among the investigated Patagonian sites. Particularly, the archetypal ericaceous mycobionts H. hepaticicola, H. variabilis, and O. maius were scarce or even absent in most samples from Patagonia.
In conclusion, whereas Sebacinales are the dominant mycorrhizal partners of the Andean clade species from tropical mountains, some archetypal ErMF are common partners of some Gaultheria spp. in southern South America. Regional differences could be related to different habitats and/or, more likely, to plant preferences of mycorrhizal fungi. However, further research is needed to draw any conclusion. First, more studies are needed to understand how the structure of mycorrhizal fungal communities varies across different environmental conditions within the same plant species or genera in South America. For example, Gorzelak et al. (2012) compared the community structure of ErMF associated with Vaccinium membranaceum along an elevation gradient in Canada, showing that vegetation types influence the distribution of mycorrhizal fungi. Similarly, Bougoure et al. (2007) observed differences in fungal communities associated with Calluna vulgaris among different habitat types, including forest, heathland, and their transition zone.
The case of the genus Gaultheria
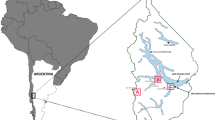
Gaultheria, a genus comprising ca. 130 species, can be found across various regions including Australia, Asia, the Indomalaya region, New Zealand, as well as tropical and temperate regions of the Americas (Middleton 1991). In South America, Gaultheria has the broadest distribution within Ericaceae, comprising 57 species ranging from northern Venezuela to southern Chile (Fig. 2). This genus is widely distributed in temperate regions, while in tropical regions it is more confined to high altitudes in the Andes and in coastal mountain ranges of Brazil.

Diversity of habitats of the genus Gaultheria (Ericaceae) in South America. a Gaultheria poeppiggi in recent volcanic soils in Conguillío National Park, within the temperate Andes of south-central Chile (photo by Nicolás Arcos). b G. poeppiggi in shrublands within the Cordoba Mountain Range, located in central Argentina (photo by Carlos Urcelay). c G. pumila in the high Andean steppe of Catedral Mountain, near Bariloche, in Argentine Patagonia (photo by Natalia Fernández). d G. pumila growing in Sphagnum sp. bogs on Navarino Island, within the extreme southern Magellan and Chilean Antarctica Region (photo by Felipe Osorio). e G. reticulata growing in tropical mountain forests in Loja, Ecuador (photo by María Cristina Córdova). f G. anastomosans in the Páramos of Chingaza, in the Cundinamarca Department, Colombia (photo by Universidad de Los Andes, Colombia). g G. eriophylla in the Atlantic Forest of Petropolis, Río de Janeiro, Brazil (photo by Joey Santoro)
Probably due to its broad distribution, Gaultheria is the ericaceous genus with the highest number of publications regarding mycorrhizal type and fungal identity in South America (Table 2). These surveys cover a large portion of the distribution of the genus, including tropical mountain forests in Ecuador (Setaro et al. 2006a, b), the Paramo in Colombia (Guerrero 1996; Sarmiento 2020), temperate forests in Southern Chile and Argentina (Godoy and Marin 2019; Bruzone et al. 2015, 2017; Vohník et al. 2023), and the Córdoba Mountain Range in Argentina (Urcelay 2002). In these habitats, the genus Gaultheria exhibits predominantly ErM associations and associates with Sebacinales and ascomycetous ErMF (Table 2). While the species sampled in the tropical mountains of Ecuador associate mostly with Sebacinales (Setaro et al. 2006b, 2013; Setaro and Kron 2011), the species studied in Argentina showed a wider diversity of archetypal ErMF, including H. hepaticicola, H. bicolor, and Oidiodendron aff. maius (Bruzone et al. 2015, 2017; Vohník et al. 2023). Yet, in some Gaultheria species, these archetypal ErMF were low in abundance or absent (Bruzone et al. 2015, 2017), and some common ErMF from the Northern Hemisphere, H. variabilis and O. maius, were lacking (Vohník et al. 2023). Also, some Gaultheria species from Argentine Patagonia presented highly diverse fungal communities and a strong spatial differentiation across relatively short distances (Vohník et al. 2023). Remarkably, under certain conditions, Gaultheria establishes additional AM associations (Table 2), e.g., G. poeppigi in Córdoba Mountain range (Urcelay 2002) or G. myrsinoides in the Páramo (Guerrero 1996). Moreover, CVM has also been observed in the roots of Gaultheria in Costa Rica (Rains et al. 2003), and probably this type of association was what Sarmiento (2020) reported as ECM in G. anastomosans.
In general, extreme temperatures occur in these areas, with very low temperatures during the winter and warm conditions during the summer (Luebert and Pliscoff 2006). Most of these habitats are characterized by acidic soils, low nutrient availability, and high organic matter content, which is a common environment for most ErM plants (Smith and Read 2008; Kohout 2017). Accordingly, ErMF has an enzymatic pool similar to saprophytic soil fungi, which is crucial for nutrient (N and P) mobilization from recalcitrant organic sources (Martino et al. 2018). This may explain the presence of Gaultheria in habitats with soil rich in organic matter in South America. Nevertheless, Gaultheria also occurs in soils derived from recent volcanic depositions in temperate Andes (Bruzone et al. 2017; Muñoz et al. 2021) that contain extremely low organic matter, mainly associated with pioneer species such as Berberis sp. (Berberidaceae), Orites myrtoidea (Proteaceae), and Acaena antarctica (Rosaceae). Further research is needed to understand the role of ErMF in these soils; however, one possible benefit obtained from these fungi could be related to resistance to high levels of available metals, especially aluminum, in the soil solution (Oliva et al. 2012; Daghino et al. 2016). Because of its broad distribution in South America and globally, and due to its probable capacity to establish different types of mycorrhizal associations, Gaultheria offers an excellent model to study which environmental factors promote the establishment of one or more mycorrhizal types.
Discussion
ErM is the least studied type of mycorrhizal association (Vohník 2020). Furthermore, the understanding of the evolution and ecology of mycorrhizal associations within the Ericaceae family, which encompasses AM, ectendomycorrhizas (including arbutoid, monotropoid, and cavendishioid mycorrhizas), and ErM, still lacks a detailed understanding, even though broad models exist at higher phylogenetic levels (Lallemand et al. 2016; Freudenstein et al. 2016). This family is very intriguing in terms of the fast evolution and diversification of its mycorrhizal associations, about which many dogmas need to be revisited (Albornoz et al. 2021), particularly since current knowledge is predominantly derived from temperate habitats in the Northern Hemisphere. Here we compiled all available scientific information about South American Ericaceae mycorrhizal types and mycorrhizal fungal diversity, revealing a significant lack of studies concerning ErM in this region, particularly within 15 out of the 33 local genera. The fact that ErM mycorrhizas are understudied in South America has been acknowledged in previous publications (Kohout 2017; Albornoz et al. 2021; Vohník 2020). However, our study represents the first empirical attempt to quantify this information gap. Our findings align with a similar lack of information for ErM reported for other non-European regions, such as Africa, Central Asia, and Oceania (Soudzilovskaia et al. 2020).
Nevertheless, the existing data of mycorrhizal associations in South American Ericaceae about ErM helps to contrast some affirmations that can be found in current mycorrhizal literature, which can be summarized in the following: (1) “ErM plants mostly occur in cold tundra and boreal habitats” (Tedersoo et al. 2020). Because ErM plants are important components of vegetation in these biomes (Kohout 2017), it has been assumed that they are the typical habitats for ErM plants at the global scale. However, here we showed that ErM plant species exhibit remarkable diversity in tropical habitats in South America. Globally, the highest richness of Ericaceae occurs in tropical and subtropical regions of the Southern Hemisphere, where several species-rich genera are associated with mountain ranges, such as Rhododendron in New Guinea Highlands, Vaccinium in the Andes, and Erica in the Cape Floristic Region in South Africa (Kohout 2017; Schwery et al. 2015). Further studies are needed to expand our knowledge on the functioning of ErM in these habitats. (2) “ErM plants are predominantly shrubs” (Soudzilovskaia et al. 2020). Given that ErM research has focused mainly on shrubs from the Northern Hemisphere, ErM literature supports a prevailing notion that ErM hosts are mostly shrubs. Contrary, ErM plant species in South America display various life forms, from small shrubs to trees, including different types of epiphytes. However, the functioning of ErM symbiosis across plants with different life forms has not been assessed yet. (3) “Subfamilies from the EAI clade form exclusively ErM” (Lallemand et al. 2016). While these subfamilies of Ericaceae have been regarded as exclusively ErM, evidence in South America suggests a high degree of flexibility in terms of their mycorrhizal types, including species that establish both AM and ErM associations. However, further research is needed to assess the functionality of these AM associations.
In conclusion, we showed that including regions that have been historically less studied contributes to broadening of the generalized understanding of ericoid mycorrhiza. We hope that this approach will inspire researchers from South America and other regions where ErM symbiosis remains largely unexplored, i.e., the Amazon basin and Guiana Highlands in South America (Fig. S1) and the Cape Floristic Region in South Africa. Additionally, we have highlighted the value of studying globally distributed genera, such as Gaultheria, to expand our understanding of biogeographic patterns of ErM. All these efforts will improve our understanding of fungal symbioses within what is arguably the most complex plant family in terms of mycorrhizal associations.
References
Albornoz FE, Dixon KW, Lambers H (2021) Revisiting mycorrhizal dogmas: are mycorrhizas really functioning as they are widely believed to do? Soil Ecology Letters 3:73–82
Almario J, Jeena G, Wunder J, Langen G, Zuccaro A, Coupland G, Bucher M (2017) Root-associated fungal microbiota of nonmycorrhizal Arabis alpina and its contribution to plant phosphorus nutrition. Proceedings of the National Academy of Sciences, USA 114:E9403–E9412
Antonelli A, Nylander JAA, Persson C, Sanmartín I (2009) Tracing the impact of the Andean uplift on neotropical plant evolution. Proceedings of the National Academy of Sciences, USA 106:9749–9754
Baba T, Hirose D, Sasaki N, Watanabe N, Kobayashi N, Kurashige Y, Karimi F, Ban T (2016) Mycorrhizal formation and diversity of endophytic fungi in hair roots of Vaccinium oldhamii Miq. in Japan. Microbes and environments, ME16011
Baião EB, Kasuya CM (2007) Associações micorrízicas em Agarista Coriifolia (Ericaceae). 58º Congresso Nacional de Botanica-São Paulo.
Bellgard SE (1991) Mycorrhizal associations of plant species in Hawkesbury sandstone vegetation. Austral Journal of Botany 39:357–364
Bermudez D, Benzing DH (1989) Fungi in neotropical epiphyte roots. BioSystems 23:65–73
Besen K (2017) Autoecologia de Gaylussacia brasiliensis (Ericaceae), em restinga da Ilha de Santa Catarina, sul do Brasil. PhD Thesis. Universidade Federal de Santa Catarina. Florianópolis, Brazil
Bidartondo MI, Bruns TD (2001) Extreme specificity in epiparasitic Monotropoideae (Ericaceae): widespread phylogenetic and geographic structure. Mol Ecol 10:2285–2295
Bidartondo MI, Bruns TD (2002) Fine-level mycorrhizal specificity in the Monotropoideae (Ericaceae): specificity for fungal species groups. Mol Ecol 11:557–569
Bougoure DS, Parkin PI, Cairney JWG, Alexander IJ, Anderson IC (2007) Diversity of fungi in hair roots of Ericaceae varies along a vegetation gradient. Mol Ecol 16:4624–4636
Brundrett MC, Tedersoo L (2018) Evolutionary history of mycorrhizal symbioses and global host plant diversity. New Phytol 220:1108–1115
Bruzone MC, Fontenla SB, Vohník M (2015) Is the prominent ericoid mycorrhizal fungus Rhizoscyphus ericae absent in the Southern Hemisphere’s Ericaceae? A case study on the diversity of root mycobionts in Gaultheria spp. from northwest Patagonia. Argentina Mycorrhiza 25:25–40
Bruzone MC, Fehrer J, Fontenla SB, Vohník M (2017) First record of Rhizoscyphus ericae in Southern Hemisphere’s Ericaceae. Mycorrhiza 27:147–163
Bueno CG, Marin C, Silva-Flores P, Aguilera P, Godoy R (2017) Think globally, research locally: emerging opportunities for mycorrhizal research in South America. New Phytol 215:1306–1309
Cairney JWG, Meharg AA (2003) Ericoid mycorrhiza: a partnership that exploits harsh edaphic conditions. Eur J Soil Sci 54:735–740
Carrillo R, Guerrero J, Rodríguez M, Meriño-Gergichevich C (2015) Colonization of blueberry (Vaccinium corymbosum) plantlets by ericoid mycorrhizae under nursery conditions. Ciencia e Investigación Agraria 42:365–374
Castillo CG, Borie F, Godoy R, Rubio R, Sieverding E (2006) Diversity of mycorrhizal plant species and arbuscular mycorrhizal fungi in evergreen forest, deciduous forest and grassland ecosystems of Southern Chile. Journal of Applied Botany and Food Quality 80:40–47
Chaurasia B, Pandey A, Palni L (2005) Distribution, colonization and diversity of arbuscular mycorrhizal fungi associated with central Himalayan rhododendrons. For Ecol Manage 207:315–324
Daghino S, Martino E, Perotto S (2016) Model systems to unravel the molecular mechanisms of heavy metal tolerance in the ericoid mycorrhizal symbiosis. Mycorrhiza 26:263–274
Das P, Kayang H (2012) Root fungal associations in Gaultheria fragrantissima. Journal of Agricultural Technology 8:133–141
Delavaux CS, Smith-Ramesh LM, Kuebbing SE (2017) Beyond nutrients: a meta-analysis of the diverse effects of arbuscular mycorrhizal fungi on plants and soils. Ecology 98:2111–2119
DeMars BG, Boerner REJ (1996) Vesicular arbuscular mycorrhizal development in the Brassicaceae in relation to plant life span. Flora 191:179–189
Fehrer J, Réblová M, Bambasová V, Vohník M (2019) The root-symbiotic Rhizoscyphus ericae aggregate and Hyaloscypha (Leotiomycetes) are congeneric: phylogenetic and experimental evidence. Stud Mycol 92:195–225
Fontenla S, Godoy R, Rosso P, Havrylenko (1998) Root associations in Austrocedrus forests and seasonal dynamics of arbuscular mycorrhizas. Mycorrhiza 8:29–33
Freudenstein JV, Broe BB, Feldenkris ER (2016) Phylogenetic relationships at the base of Ericaceae: implications for vegetative and mycorrhizal evolution. Taxon 65:794–804
Fukuchi S, Obase K, Tamai Y, Yajima T, Miyamoto T (2011) Vegetation and colonization status of mycorrhizal and endophytic fungi in plant species on acidic barren at crater basin of volcano Esan in Hokkaido, Japan. Eurasian Journal of Forest Research 14:1–11
Geml J, Pastor N, Fernandez L, Pacheco S, Semenova TA, Becerra AG, Wicaksono CY, Nouhra ER (2014) Large-scale fungal diversity assessment in the Andean Yungas forests reveals strong community turnover among forest types along an altitudinal gradient. Mol Ecol 23:2452–2472
Genre A, Lanfranco L, Perotto S, Bonfante P (2020) Unique and common traits in mycorrhizal symbioses. Nat Rev Microbiol 18:649–660
Godoy R, Marín C (2019) Chapter 16: Mycorrhizal studies in temperate rainforests of Southern Chile. In: Pagano MC, Lugo MA (eds) Mycorrhizal fungi in South America. Springer, Switzerland, pp 315–341
Godoy R, Romero R, Carrillo R (1994) Estatus micotrófico de la flora vascular en bosques de coníferas nativas del sur de Chile. Rev Chil Hist Nat 67:209–220
Gorzelak MA, Hambleton S, Massicotte HB (2012) Community structure of ericoid mycorrhizas and root-associated fungi of Vaccinium membranaceum across an elevation gradient in the Canadian Rocky Mountains. Fungal Ecol 5:36–45
Guerrero E (1996) Survey of mycorrhiza in a high Andean Paramo. Proceedings of the fourth European Symposium on Mycorrizas. 121–124
John TS (1980) Uma lista de espécies de plantas tropicais brasileiras naturalmente infectadas com micorriza vesicular-arbuscular. Acta Amazon 10:229–234
Kohout P, Tedersoo L (2017) Effect of soil moisture on root-associated fungal communities of Erica dominans in Drakensberg mountains in South Africa. Mycorrhiza 27:397–406
Kohout P (2017) Biogeography of ericoid mycorrhiza. in ed. Tedersoo L. Biogeography of mycorrhizal symbiosis. Switzerland: Springer. 179–193
Kolařík M, Vohník M (2018) When the ribosomal DNA does not tell the truth: the case of the taxonomic position of Kurtia argillacea, an ericoid mycorrhizal fungus residing among Hymenochaetales. Fungal Biol 122:1–18. https://doi.org/10.1016/j.funbio.2017.09.006
Kong A, Cifuentes J, Estrada-Torres A, Guzmán-Dávalos L, Garibay-Orijel R (2015) Russulaceae associated with mycoheterotroph Monotropa uniflora (Ericaceae) in Tlaxcala, Mexico: a phylogenetic approach. Cryptogam, Mycol 36:479–512
Konno K, Akasaka M, Koshido C, Katayama ON, Spake R, Amano T (2020) Ignoring non-English-language studies may bias ecological meta-analyses. Ecol Evol 10:6373–6384
Koske RE, Gemma JN, Englander L (1990) Vesicular-arbuscular mycorrhizae in Hawaiian Ericales. Am J Bot 77:64–68
Kottke I, Haug I, Setaro S, Suárez JP, Weiß M, Preußing M, Nebel M, Oberwinkler F (2008) Guilds of mycorrhizal fungi and their relation to trees, ericads, orchids and liverworts in a neotropical mountain rain forest. Basic Appl Ecol 9:13–23
Kron KA, Luteyn J (2005) Origins and biogeographic patterns in Ericaceae: new insights from recent phylogenetic analyses. Biologiske Skrifter 55:479–500
Kron KA, Judd WS, Stevens PF, Crayn DM, Anderberg AA, Gadek PA, Quinn CJ, Luteyn JL (2002) Phylogenetic classification of Ericaceae: molecular and morphological evidence. Botanical Reviews 68:335–423
Lallemand F, Gaudeul M, Lambourdiere J, Matsuda Y, Hashimoto Y, Selosse MA (2016) The elusive predisposition to mycoheterotrophy in Ericaceae. New Phytol 212:314–319
Lancheros HO (2012) Caracterización de las micorrizas nativas en agraz Vaccinium meridionale Swartz y evaluación de su efecto sobre el crecimiento plantular. Master Thesis. Universidad Nacional de Colombia, Bogotá, Colombia
Leake JR (1994) The biology of myco-heterotrophic (‘saprophytic’) plants. New Phytol 127:171–216
Linder HP (2003) The radiation of the Cape flora, southern Africa. Biol Rev 78:597–638
Luebert F, Weigend (2014) Phylogenetic insights into Andean plant diversification. Frontiers in Ecology and Evolution 2.https://doi.org/10.3389/fevo.2014.00027
Luebert F, Pliscoff P (2006) Sinopsis bioclimática y vegetacional de Chile. Editorial Universitaria, Santiago de Chile
Luteyn JL (2002) Diversity, adaptation, and endemism in neotropical Ericaceae: biogeographical patterns in the Vaccinieae. Botanical Reviews 68:55–87
Maherali H, Oberle B, Stevens PF, Cornwell WK, McGlinn DJ (2016) Mutualism persistence and abandonment during the evolution of the mycorrhizal symbiosis. Am Nat 188:E113–E125
Martin F, Kohler A, Murat C, Veneault-Fourrey C, Hibbett DS (2016) Unearthing the roots of ectomycorrhizal symbioses. Nat Rev Microbiol 14:760–773
Martino E, Morin E, Grelet GA, Kuo A, Kohler A, Daghino S, Barry KW, Cichocki N, Clum A, Dockter RB (2018) Comparative genomics and transcriptomics depict ericoid mycorrhizal fungi as versatile saprotrophs and plant mutualists. New Phytol 217:1213–1229
Massicotte HB, Melville LH, Peterson RL (2005) Structural characteristics of root-fungal interactions for five ericaceous species in eastern Canada. Can J Bot 83:1057–1064
Menoyo E, Becerra AG, Renison D (2007) Mycorrhizal associations in Polylepis woodlands of Central Argentina. Can J Bot 85:526–531
Merckx V (2013) Mycoheterotrophy, the biology of plants living on fungi. Springer, Berlin, Germany
Middleton DJ (1991) Infrageneric classification of the genus Gaultheria L. Ericaceae. Bot J Linn Soc 106:229–258
Miyauchi S, Kiss E, Kuo A, Drula E, Kohler A, Sánchez-García M, Morin E, Andreopoulos B, Barry KW, Bonito G et al (2020) Large-scale genome sequencing of mycorrhizal fungi provides insights into the early evolution of symbiotic traits. Nat Commun 11:5125
Mnasri M, Janoušková M, Rydlová J, Abdelly C, Ghnaya T (2017) Comparison of arbuscular mycorrhizal fungal effects on the heavy metal uptake of a host and a non-host plant species in contact with extraradical mycelial network. Chemosphere 171:476–484
Mujica MI, Bueno GC, Duchicela J, Marin C (2019) Strengthening mycorrhizal research in South America. New Phytol 224:563–567
Muñoz G, Orlando J, Zúñiga-Feest A (2021) Plants colonizing volcanic deposits: root adaptations and effects on rhizosphere microorganisms. Plant Soil 461:265–279
Neri de Almeida AF (2019) Fungos micorrízicos arbusculares em Gaylussacia brasiliensis (Spreng.) Meissner em Área de Restinga. Master thesis. Universidade Federal de Santa Catarina, Florianópolis, Brazil
Neyland R, Hennigan MK (2004) A cladistic analysis of Monotropa uniflora (Ericaceae) inferred from large ribosomal subunit (26S) rRNA gene sequences. Castanea 69:265–271
Nuñez MA, Amano T (2021) Monolingual searches can limit and bias results in global literature reviews. Nature Ecology and Evolution 5:264
Obase K, Matsuda Y, Ito S (2013) Enkianthus campanulatus (Ericaceae) is commonly associated with arbuscular mycorrhizal fungi. Mycorrhiza 23:199–208
Oliva SR, Mingorance MD, Leidi EO (2012) Tolerance to high Zn in the metallophyte Erica andevalensis Cabezudo and Rivera. Ecotoxicology 21
Pacheco Flores A, Barcos-Arias M, Naranjo-Morán J, Peña-Tapia D, Moreira-Gómez R (2022) Ericaceous plants: a review for the bioprospecting of ericoid mycorrhizae from Ecuador. Diversity 14:648
Pedraza-Peñalosa P, Luteyn JL (2015) Andean Vaccinium (Ericaceae: Vaccinieae): seven new species from South America. Brittonia 63:257–275
Perotto S, Daghino S, Martino E (2018) Ericoid mycorrhizal fungi and their genomes: another side to the mycorrhizal symbiosis? New Phytol 220:1141–1147
POWO (2023) Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Published on the Internet; http://www.plantsoftheworldonline.org/ Retrieved 8 May 2023
Rains KC, Nadkarni NM, Bledsoe CS (2003) Epiphytic and terrestrial mycorrhizas in a lower mountain Costa Rican cloud forest. Mycorrhiza 13:257–264
Rice AV, Currah RS (2006) Oidiodendron maius: Saprobe in Sphagnum peat, mutualist in ericaceous roots? In: Schulz et al (eds) Microbial root endophytes. Switzerland: Springer. 227–246
Richard S, Millot M, Gardes SMA (2005) Diversity and structuration by hosts of the below-ground mycorrhizal community in an old-growth Mediterranean forest dominated by Quercus ilex L. New Phytol 166:1011–1023
Riess K, Oberwinkler F, Bauer R, Garnica S (2014) Communities of endophytic Sebacinales associated with roots of herbaceous plants in agricultural and grassland ecosystems are dominated by Serendipita herbamans sp. nov. PLoS ONE 9: e94676
Santos OM, De Oliveira NC, de Nováis RF (1995) Observações preliminares sobre fungos micorrízicos vesículo-arbusculares em plantas crescendo em dunas na Bahia. Revista Ceres 42:191–202
Sarmiento D (2020) Caracterización de rasgos florísticos y micorrizales de plantas dominantes de pajonales mixtos en el Páramo de Guerrero, Cundinamarca, Colombia. Undergraduate thesis. Universidad de Los Andes, Colombia
Schwery O, Onstein RE, Bouchenak-Khelladi Y, Xing Y, Carter RJ, Linder HP (2015) As old as the mountains: the radiations of the Ericaceae. New Phytol 207:355–367
Selosse MA, Setaro S, Glatard F, Richard F, Urcelay C, Weiß M (2007) Sebacinales are common mycorrhizal associates of Ericaceae. New Phytol 174:864–878
Selosse MA, Dubois MP, Alvarez N (2009) Do Sebacinales commonly associate with plant roots as endophytes? Mycol Res 113:1062–1069
Selosse MA, Petrolli R, Mujica MI, Laurent L, Perez-Lamarque B, Figura T, Bourceret A, Jacquemyn H, Li T, Gao J et al (2022) The Waiting Room Hypothesis revisited by orchids: were orchid mycorrhizal fungi recruited among root endophytes? Ann Bot 129:259–270
Setaro S, Weiß M, Oberwinkler F, Kottke I (2006a) Sebacinales form ectendomycorrhizas with Cavendishia nobilis, a member of the Andean clade of Ericaceae, in the mountain rain forest of southern Ecuador. New Phytol 169:355–365
Setaro S, Kottke I, Oberwinkler F (2006b) Anatomy and ultrastructure of mycorrhizal associations of neotropical Ericaceae. Mycol Prog 5:243–254
Setaro S, Garnica S, Herrera PI, Suárez JP, Göker MA (2012) A clustering optimization strategy to estimate species richness of Sebacinales in the tropical Andes based on molecular sequences from distinct DNA regions. Biodivers Conserv 21:2269–2285
Setaro S, Kron K (2011) Neotropical and North American Vaccinioideae (Ericaceae) share their mycorrhizal Sebacinales-an indication for concerted migration? PLoS Curr. 3, RRN1227
Setaro SD, Suárez JP, Herrera P, Cruz D, Kottke I (2013) Distinct but closely related Sebacinales form mycorrhizae with coexisting Ericaceae and Orchidaceae in a Neotropical Mountain Area. In Piriformospora indica; Springer: Berlin/Heidelberg, Germany, 81–105.
Smith SE, Read DJ (2008) Mycorrhizal symbiosis. Academic Press and Elsevier, London
Soudzilovskaia NA, Vaessen S, Barcelo M, He J, Rahimlou S, Abarenkov K, Brundrett MC, Gomes SIF, Merckx V, Tedersoo L (2020) FungalRoot: global online database of plant mycorrhizal associations. New Phytol 227:955–966
Strullu-Derrien C, Selosse MA, Kenrick P, Martin FM (2018) The origin and evolution of mycorrhizal symbioses: from palaeomycology to phylogenomics. New Phytol 220:1012–1030
Tedersoo L, Brundrett M (2017) Chapter 19: Evolution of ectomycorrhizal symbiosis in plants. In: Tedersoo L (ed) Biogeography of mycorrhizal symbiosis, Ecological Studies 230. Springer, Switzerland, pp 407–467
Tedersoo L, May TW, Smith ME (2010) Ectomycorrhizal lifestyle in fungi: global diversity, distribution, and evolution of phylogenetic lineages. Mycorrhiza 20:217–263
Tedersoo L, Bahram M, Zobel M (2020) How mycorrhizal associations drive plant population and community biology. Science 367, eaba1223
Teste FP, Jones MD, Dickie IA (2020) Dual-mycorrhizal plants: their ecology and relevance. New Phytol 225:1835–1851
Urcelay C (2002) Co-occurrence of three fungal root symbionts in Gaultheria poeppiggi DC in Central Argentina. Mycorrhiza 12:89–92
Urgiles N, Haug I, Setaro S, Aguirre N (2016) Introduction to Mycorrhizas in the tropics with emphasis on the Montane Forest in Southern Ecuador. Universidad Nacional de Loja, Ecuador
van der Heijden MGA, Martin FM, Selosse MA, Sanders IR (2015) Mycorrhizal ecology and evolution: the past, the present, and the future. New Phytol 205:1406–1423
Van Geel M, Jacquemyn H, Peeters G, van Acker K, Honnay O, Ceulemans T (2020) Diversity and community structure of ericoid mycorrhizal fungi in European bogs and heathlands across a gradient of nitrogen deposition. New Phytol 228:1640–1651
Varma A, Sherameti I, Tripathi S, Prasad R, Das A, Sharma M, Bakshi M, Johnson JM, Bhardwaj S, Arora M et al (2012) The symbiotic fungus Piriformospora indica: review. In: Hock B (ed) The Mycota, vol IXB – fungal associations, 2nd edn. Springer, Berlin, Germany, pp 231–254
Veiga RSL, Howard K, van der Heijden MGA (2012) No evidence for allelopathic effects of arbuscular mycorrhizal fungi on the non-host plant Stellaria media. Plant Soil 360:319–331
Veiga RSL, Faccio A, Genre A, Pieterse CM, Bonfante P, van der Heijden MGA (2013) Arbuscular mycorrhizal fungi reduce growth and infect roots of the non-host plant Arabidopsis thaliana. Plant, Cell Environ 36:1926–2193
Vohník M (2020) Ericoid mycorrhizal symbiosis: theoretical background and methods for its comprehensive investigation. Mycorrhiza 30:671–695
Vohník M, Sadowsky JJ, Kohout P et al (2012) Novel root-fungus symbiosis in Ericaceae: sheathed ericoid mycorrhiza formed by a hitherto undescribed basidiomycete with affinities to Trechisporales. PLoS ONE 7:e39524. https://doi.org/10.1371/journal.pone.0039524
Vohník M, Mrnka L, Lukešová T, Bruzone MC, Kohout P, Fehrer J (2013) The cultivable endophytic community of Norway spruce ectomycorrhizae from microhabitats lacking ericaceous hosts is dominated by ericoid mycorrhizal Meliniomyces variabilis. Fungal Ecol 6:281–292
Vohník M, Pánek M, Fehrer J, Selosse M-A (2016) Experimental evidence of ericoid mycorrhizal potential within Serendipitaceae (Sebacinales). Mycorrhiza 26:831–846
Vohník M, Figura T, Réblová M (2022) Hyaloscypha gabretae and Hyaloscypha gryndleri spp. nov. (Hyaloscyphaceae, Helotiales), two new mycobionts colonizing conifer, ericaceous and orchid roots. Mycorrhiza 32:105–122
Vohník M, Bruzone MC, Knoblochová T, Fernández NV, Kolaříková Z, Větrovský T, Fontenla SB (2023) Exploring structural and molecular diversity of Ericaceae hair root mycobionts: a comparison between Northern Bohemia and Argentine Patagonia. Mycorrhiza 33:425–447
Weiss M, Waller F, Zuccaro A, Selosse M-A (2016) Sebacinales—one thousand and one interactions with land plants. New Phytol 211:20–40
Acknowledgements
MIM thanks ANID – Fondecyt postdoc for funding support (grant # 3200774). MAS thanks Institut Universitaire de France for the support. We are deeply thankful for the comments of anonymous reviewers on earlier versions of this paper. HH acknowledges the support of the Fondo de Fomento al Desarrollo Científico y Tecnológico, grant number [ID23I10303].
Funding
Agencia Nacional de Investigación y Desarrollo,3200774
Author information
Authors and Affiliations
Contributions
MIM conceived and designed the study. Material preparation and data collection were performed by MIM, CSS, and MC. The first draft of the manuscript was written by all authors. MIM and MAS discussed and edited the first version of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Mujica, M.I., Herrera, H., Cisternas, M. et al. Mycorrhizas in South American Ericaceae. Mycorrhiza 34, 1–18 (2024). https://doi.org/10.1007/s00572-024-01141-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00572-024-01141-z