
Abstract
Human activities are increasing wildfires and livestock activity in arid ecosystems with potential implications for the spread of invasive grasses. The objective of this study was to test whether fire history and cattle activity alter soil resource gradients, thereby affecting patterns of Bromus rubens L. (red brome) invasion. Six paired burned and unburned transect lines (1-km long) were established in the northeast Mojave Desert along the boundaries of four independent wildfire scars. At 100-m transect increment points, we measured the distance to the two nearest cowpats, and two random points and measured the density, height, biomass, and seed production of red brome, soil moisture and inorganic nitrogen (N). Cattle activity was 29% greater along burned transects compared to unburned transects (P < 0.05). Red brome height, density, and seed production were 11–34% greater along burned transects than unburned transects (P < 0.05). Red brome height, biomass, density, and seed production were twofold to tenfold greater next to cowpats compared to random points (P < 0.05). Soils along burned transects and beneath cowpats had greater soil inorganic N (P < 0.05), which was positively correlated with red brome density, height, biomass, and seed production (R2 = 0.60–0.85, P < 0.0001). Transgenerational effects were evident as seeds from red brome next to cowpats had 27% higher germination than seeds collected from random points. Positive responses of red brome to increased inorganic N related to fire and cattle activity may contribute fine fuel infill that drives invasive grass-fire cycles in deserts.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Humans throughout their history have transported plant and animal species beyond their natural range resulting in non-native species introductions across Earth’s continents (Perrings et al. 2005). The spread of exotic species increased during the agricultural revolution with trade and migration (Hulme 2009). However, it expanded most dramatically following the industrial revolution due to mechanized travel, immigration, and globalization (Hulme 2009). One of the most dramatic and widespread exotic species introductions was cattle (Bos taurus), domesticated by humans more than 8500 years ago in western Asia and later distributed globally (Kumar et al. 2016). Cattle arrived in North America with the Spaniards in the early 1500s (Bowling 1942). Today more than 106 million head of cattle, or about 12% of the global total, are distributed across North America, including desert rangelands (Commission for Environmental Cooperation 2015).
More than three centuries after the arrival of cattle in North America, red brome, an annual winter grass from southern Europe, was introduced to western North America, likely during a period of rapid expansion in immigration and commerce in California (Salo 2005). Red brome quickly spread and invaded the Mojave, Great Basin, and Sonoran deserts over subsequent decades (Salo 2005), particularly during cycles of warmer Pacific decadal oscillation regimes that produced abundant winter precipitation (Mantua 2000). Like Bromus tectorum L. (cheatgrass), its early establishment during the fall period fuels rapid growth the following spring resulting in soil resource depletion that can negatively impact native plants (Bishop et al. 2020). In addition, following seed drop in late spring, red brome’s tissues become dry, producing landscape fuel conditions that increase the frequency and size of wildfires in deserts of western North America (Brooks and Matchett 2006; Horn and St Clair 2017).
Animals play a central role in facilitating the spread and establishment of plant invaders through seed dispersal (Hogan and Phillips 2011), soil and vegetation disturbance (Chambers et al. 2016), and nutrient enrichment (Auman et al. 1998). Livestock is known to disperse seeds both through ingestion (endozoochory) (Mouissie et al. 2005) and attachment (epizoochory) (Chuong et al. 2016). In addition, dung from various large mammals contains a variety of exotic plant seeds (Mouissie et al. 2005; Dovrat et al. 2012). Microsite conditions created by dung piles can also alter plant community composition (Auman et al. 1998; Dai 2000). However, much less is known about the role that endozoochory and dung piles play in the productivity of invasive annual grasses that are driving changing fire regimes in deserts of western North America (Eichberg et al. 2007). This is a critical knowledge gap for two reasons: first, livestock grazing is widespread in deserts of North America; and the stress gradient hypothesis (Callaway 2007) predicts that dung pats may provide microsite conditions that overcome water and nutrient limitations to the establishment success of invasive annual grasses in deserts (López et al. 2016).
Arid ecosystems are defined by vegetation that is both water and nitrogen limited (Yahdjian et al. 2011; Gill et al. 2018). The distribution and productivity of invasive annual brome grasses in deserts are responsive to soil resource pulses (Belnap et al. 2016; Horn and St Clair 2017), often resulting in prolific seed production (Horn et al. 2017). Soil moisture and nutrients vary along gradients in desert landscapes, being concentrated beneath native shrubs (islands of fertility) and decreasing in inter-shrub spaces (Schlesinger et al. 1996). This tends to produce high-density grasses beneath shrubs with sparser production between shrubs, limiting the continuity of fine plant fuels that drive desert fires (Lucero et al. 2021). Cattle which are prolific dung producers (Chastain and Camberato 2004), move and forage between shrubs in desert landscapes (France et al. 2008) and, as a result, leave their droppings in the more resource-limited inter-shrub spaces. Dung piles may influence three potential resources that could, in theory, fuel the spread and growth of invasive annual grasses in inter-shrub spaces, thereby increasing fire potential: seeds, nutrients, and moisture (Dai 2000). While greater resource availability around cattle dung may increase invasibility directly by priming growth and seed production, it can also influence invader fecundity by affecting seed quality in subsequent generations (Luzuriaga et al. 2006). There is some evidence for transgenerational effects in annual brome grasses (Steinger et al. 2000), but the role of plant resource availability in cross-generational seed viability of invasive brome grasses is limited.
In the northeast Mojave Desert, widespread cattle grazing, and wildfires overlap with red brome invasion providing an opportunity to test how post-fire conditions and cattle dung deposition affect soil resource availability and its influence on the establishment and fecundity of red brome. We hypothesized that: (1) post-fire environments and cowpats would increase the growth and seed production of red brome via increases in soil moisture and nitrogen, and (2) soil resource enrichment in the vicinity of cowpats and along burn transects would create a transgenerational effect by improving the quality of F1 generation seed.
Materials and methods
This study was conducted in the Beaver Dam Wash in the northeastern Mojave Desert (Latitude 37.0837 N, Longitude 114.0119 W, elevation 1216 m) in May 2020. The 30-year mean annual precipitation from the nearest Climate Station is 26.5 cm with a mean annual temperature of 16 °C (Western Reginal Climate Center 2019). The study area is characterized by native shrubs of which Coleogyne ramosissima (Torr.), Larrea tridentata (DC. Coville), and Ambrosia dumosa (A. Gray Payne) are the most common. Two yucca species are common in the study area: Yucca baccata (Torr.) and Yucca brevifolia (Engelm). Common native herbaceous plants included Sphaeralcea ambigua (A. Gray), Baileya multiradiata (Harv. & A. Gray), Astragalus nuttaliianus (DC.), Plantago spp. (P. patagonica and P. ovata), and Descurainia pinnata (Walter Britton). The soil in the study area is a gravelly sandy loam (Soil Survey Staff 2015). In recent years, plant invasions have altered fire regimes in North American Deserts by increasing fire size and frequency (Brooks et al. 2004; Horn and St Clair 2017). Invasive annual grasses, Schismus arabicus Nees., and Bromus Rubens L., responded positively to early fall precipitation in 2004, resulting in the build-up of fine fuels that triggered multiple, large-scale fires in the study area (Beaver Dam Wash) in the summer of 2005 (Horn et al. 2017). Active cattle allotments exist in our study area in the Beaver Dam Wash, and we germinated viable red brome seeds from cowpat samples from our study area indicating that cattle graze red brome.
Study design
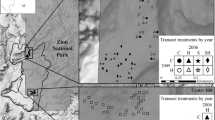
A network of six 1-km-long paired transect lines were positioned along adjacent burned/unburned boundaries within four independent fires that occurred during the summer of 2005: Westside (June, 27,059 ha), Duzak (July, 7065 ha—within Utah), Burgess 1 (July, 60 ha), and Burgess 2 fires (July, 712 ha) (Fig. 1). The fires were ignited by lightning and burn severity was classified as high in each of the four burns (Lybbert et al. 2017). Paired burned/unburned transects were placed 200 m on either side of the burn boundary. Three transect pairs were placed in the Burgess 1, Burgess 2 and Duzak fires. The remaining three transect pairs were spread across the Westside fire, which was fourfold larger than the Duzak fire and several orders of magnitude larger than the Burgess 1 and 2 fires making up most of the area burned during the 2005 fires. Transect pairs were selected on the above criteria using GIS mapping software to prevent on-site selection bias.

Map of the transect network in the study area in the Mojave Desert along the Utah–Nevada border 80-km northeast of Las Vegas Nevada. The six 1-km paired transect were arranged along the burn boundaries of the Duzak, Westside, Burgess 1 and Burgess 2 fires that burned in the summer of 2005
Field measurements
Distance to the nearest cowpat on each side of the transect lines were measured at each 100 m increment point (a total of 20 cowpats per transect) to estimate cowpat densities using point distance methods (Diggle 1975). Red brome establishment was concentrated in distinct rings that extended from the edge of the cowpats (Fig. 2), since cowpats varying dramatically in size and shape using a standard size quadrat would have done a poor job of capturing red brome density concentrated around the edges and interior of cowpats of different dimensions. Estimating cowpat influence on red brome density was, therefore, estimated by counting all individuals established from the outer ring edge to the center of the cowpat. Accordingly, the area of red brome establishment relative to each cowpat used for density calculations was estimated by measuring the longest ring edge extent and multiplying it by the ring edge distance at its perpendicular angle using a measuring tape. Based on dozens of measurements at our study site, we determined that the average area inside of the red brome rings around cowpats was 0.15 m2. As a result, a 0.15 m2 quadrat was constructed for measuring red brome density at random points where cowpats were absent as a control. Random sampling points were located by throwing the quadrat over the shoulder to avoid bias. For each transect, we had a total of 20 sampling points around cowpats and 20 random sampling points. All field measurements, and plant and soil samples were collected on April 20–24, 2020.

Photos of prolific Bromus rubens (red brome) growth and seed production around the a edges of a cowpat and b recently decomposed cowpat along our transect lines. Photo scale is approximately 1 m−2
To estimate red brome height, we randomly measured 10 red brome individuals collected from each sampling location. We harvested the red brome plants established within the area where density measurements were made using clippers at soil level and placed them in a paper bag. The dried cowpats were removed from the soil surface, and several sub-samples from the top of the cowpat were collected and placed in a paper bag. A soil moisture probe (Spectrum Technologies, Aurora, Illinois) with 7.6 cm length rods were inserted into the soil where the cowpat had been removed to measure moisture content. A soil probe was used collect two soil samples to a depth of 7 cm beneath cowpats. Soil moisture measurements and soil samples were also collected at random (control) points for comparison. Soil samples were pooled and placed in a soil sample bag for analysis of inorganic N.
Lab measurements
Red brome plant samples were allowed to air dry at 25 °C for 60 days and were weighed for vegetative and seed biomass using a balance. Allometric equations were used to estimate seed counts from seed head biomass (Bishop et al. 2020).
Soil samples were air-dried on-site and then transported to the Brigham Young University Environmental Analytical Laboratory (Provo, UT, USA), dried at 50 °C for 72 h, and homogenized. Nitrate and ammonium concentrations (mg NO3–N per kg and mg NH4–N per kg) were extracted using two mol/L KCl following the cadmium reduction method (Keeney and Nelson 1982) and the salicylate–sodium nitroprusside method (Rowland 1983). Samples were analyzed using a rapid flow injection analyzer to calculate soil nitrate and ammonium levels (Quick Chem 8500; Lachat Instruments, Loveland, CO, USA). Inorganic N concentrations were calculated as the sum of nitrate and ammonium.
Fecundity experiments: seed germination and seedling growth
Red brome seeds were collected from the plants measured and harvested next to cowpat margins or non-cowpat controls. Seeds were homogenized and pooled by treatment group from each transect line. Three sets of 25 seeds from each pooled seed sample were placed on moist germination paper discs in three separate Petri-dish at 20 °C for 14 days to test for the effect of burn condition and cowpats on germination rates in the F1 generation seed. To test for transgenerational effects on seedling emergence, nine red brome seeds pooled from each transect were placed in containers (10 × 10 × 10 cm) and filled with soil collected from the study site that had been sieved to remove any seedbank. Containers were placed in outdoor study plots in Provo, Utah starting March 15, 2022, and were watered to saturation every 3 days. Seedling emergence from soil was counted 10 days after beginning the experiment.
Statistical analysis
Mixed-effects linear models were used to examine the effects of burn status (burned or unburned transect) and cowpat or control on red brome density, height, biomass, and seed production. In our mixed models, transect type (unburned, burned) and cowpat were fixed effects, and transect was specified as a random effect. Tukey’s HSD was used to test mean differences across burn conditions and cowpat effects on red brome plants from the field and germination and seedling growth studies. Assumptions of normality and homoscedasticity were visually inspected using residual plots (e.g., histogram, scatterplot, and qqnorm of residuals). To meet equal variance assumptions, data were log-transformed when appropriate. Linear regression models characterized red brome productivity responses to gradients of inorganic N and soil moisture along the transect network. All calculations, models, assumption analyses, and statistical significance determinations were performed using JMP Pro statistical software (version 16).
Results
Responses along burned transects and cowpat microsites
Cowpat density was 29% greater along burned transects (138 per ha ± 13) compared to unburned transects (98 per ha ± 13) (P = 0.03). Red brome height, density, and seed production were 11%, 28%, and 34% greater along burned transects than unburned transects, respectively (P < 0.05) (Fig. 3). Red brome biomass was not statistically different between burned and unburned transects (P = 0.71).

The influence of burned and unburned transects and cowpats on the a seed production, b stem density, c height, and d biomass of red brome. F-values presented with asterisks indicating level of significance for P-values: *P ≤ 0.05, **P ≤ 0.001, ***P ≤ 0.0001. Mean values presented with ± 1 SE
Red brome height (2-fold), biomass (3.2-fold), density (3-fold), and seed production 10-fold) were significantly greater when growing around the margins of cowpats compared to random points lacking cowpats (P < 0.05) (Fig. 3). The burn by cowpat interaction term was not statistically significant for red brome height, biomass, or density (Fig. 3). However, red brome seed production showed a significant burn by cowpat interaction term in which the positive effects of cowpat were amplified along burned transects compared to unburned transects (P = 0.004) (Fig. 3).
Soil resource gradients
Seed germination trials of dried cowpat samples indicated that, on average, only three red brome seedlings germinated per dried cowpat, suggesting that microsite conditions drove dramatic increases in red brome density around cowpats not endozoochory (Fig. 4). Soil moisture, nitrate, ammonium, and inorganic N concentrations were 1.4-fold, 10.3-fold, 3.5-fold, and 6-fold greater in the soil beneath cowpats compared to random soil points (P < 0.05) (Fig. 4). Nitrate, ammonium, and inorganic N concentrations were 2-fold, 1.3-fold, and 1.7-fold greater along burned transects than along unburned transects (P < 0.05) with no significant difference for soil moisture (Fig. 4). Inorganic N showed a strong burn by cowpat interaction term in which the positive effects of cowpats were amplified along burned transects compared to unburned transects (P = 0.01) (Fig. 4). The burn by cowpat interaction terms was not significant for soil moisture.

The influence of burned and unburned transects and cowpats on a inorganic N, b NH4, c NH3, and d soil moisture. F-values presented with asterisks indicating level of significance for P-values: *P ≤ 0.05, **P ≤ 0.001, ***P ≤ 0.0001. Mean values presented with ± 1 SE
Soil inorganic N showed strong positive linear correlations with red brome density (R2 = 0.60, P < 0.001), height (R2 = 0.72, P < 0.001), biomass (R2 = 0.83, P < 0.001) and seed production (R2 = 0.85, P < 0.001) along the transect network (Fig. 5). Soil moisture showed weaker but significant positive correlations with density (R2 = 0.31, P = 0.02) and seed production (R2 = 0.28, P = 0.03) (Fig. 6). Red brome height and density were not significantly correlated with soil moisture.

Regression plots quantifying the relationships of inorganic N with a seed production, b stem density, c height, and d biomass of red brome along the study transect network

Regression plots quantifying the relationships of soil moisture with a seed production and b stem density of red brome along the study transect network
Transgenerational effects on seed germination
There were no statistically significant differences in the germination, emergence, or seed production of red brome seeds collected from burned or unburned transects. However, seeds collected from red brome growing on the margins of cowpats had 27% greater germination rates than seeds collected from red brome growing at random points (P = 0.02) (Fig. 7). A similar trend was seen for seedling emergence, but statistical support was weaker (P = 0.14) (Fig. 7). The fire by cowpat interaction terms in the mixed effects linear model was not statistically significant for seed germination or seedling emergence.

The influence of burned and unburned transects and cowpats on the a germination and b seedling emergence from red brome seed collected along the study transect network. F-values presented with asterisks indicating level of significance for P-values: *P ≤ 0.05, **P ≤ 0.001, ***P ≤ 0.0001. Mean values presented with ± 1 SE
Discussion
Our results showed that microsites around cowpats and burned transects had increased soil resources that influenced red brome’s growth and reproduction patterns (Fig. 3). Livestock dung deposition can disperse seeds (Poschlod and Bonn 1998; Kowarik and Lippe 2008) and create soil resource microsites that can affect plant community composition (Dai 2000). However, more research is needed to understand better how dung deposition may affect plant invasions. Red brome’s growth and seed production are responsive to soil moisture and inorganic N (He et al. 2011; Horn and St Clair 2017), that were significantly elevated under cowpats (Fig. 4). The data supported our first hypothesis that post-fire conditions and cowpats contributed to gradients of inorganic N that were positively correlated with red brome growth and fecundity (Fig. 5). There was also evidence that cowpats (but not post-fire conditions) increased soil moisture (Fig. 4), which also showed positive but weaker correlations with patterns of red brome establishment and growth (Fig. 6). Consistent with our second hypothesis, we observed transgenerational effects in which increased germination occurred in seed collected from red brome growing next to cowpats (Fig. 7).
Fire effects on soils resource gradients and red brome productivity
This study documents two significant fire-related effects. First, the higher density of cowpats along burned transects suggests greater cattle use in burned areas, consistent with another study documenting the increased movement and foraging activity of cattle in post-fire desert environments (Clark et al. 2017). We hypothesize this may be related to greater ease of movement due to reduced shrub cover or shifts toward more palatable forage species (Lybbert et al. 2017). Second, increased red brome growth and seed production along burned transects (Fig. 3), which was correlated with greater soil N (Fig. 5). Red brome establishment and growth have been shown to increase in post-fire environments (Horn et al. 2017; Bishop et al. 2020), which could be related to various factors. For example, a reduction in native shrub cover following a fire can result in declines in rodent populations (Horn et al. 2012) that suppress invasive bromegrass establishment through granivory and folivory (St. Clair et al. 2016; Bishop et al. 2020). A surprising finding in this study was the long-term persistence of elevated inorganic N along burned transects (Fig. 4). Esque et al. (2010) documented short-term inorganic N pulses immediately following desert wildfires, but our data show 1.7-fold greater inorganic N concentrations along burned transects 15 years after the fire (Fig. 4). This may be partially due to N inputs from the fire itself or competition release of soil resources related to fire-related losses of native shrubs or a combination of the two (Horn et al. 2015; Molinari et al. 2019). These results suggest that red brome is perpetuated through post-fire enrichment of inorganic N, which may have important implications for increasing fine fuel continuity that drives invasive grass-fire cycles in desert ecosystems.
Cowpat effects on soil resource gradients and red brome productivity
Previous research has shown the dispersal of exotic species through cattle dung (Mouissie et al. 2005) and the response of a few plant species to cowpats (Auman et al. 1998). This is the first study we are aware of documenting cowpat impacts on a dominant invasive bromegrass that is changing the ecology and fire regimes in deserts of North America where cattle grazing is common. The prolific growth and seed production of red brome growing around cowpats in this study (Figs. 2, 3) appear to be driven by increases in soil moisture and particularly inorganic N enrichment (Figs. 5, 6). Pulses of soil moisture stimulate red brome production in the Mojave Desert sometimes dramatically (Gill et al. 2018; Horn et al. 2017). However, red brome productivity was most strongly correlated with inorganic N gradients (Fig. 5). Elevated soil moisture and inorganic N (as found under the cowpats) can produce synergistic increases in red brome growth (Rao and Allen 2010) consistent with our results (Fig. 4).
High labile C availability in cattle dung promotes microbial decomposition that primes high N mineralization rates (Wang et al. 2018). Evidence for N limitation of red brome growth in desert soil is mechanistically supported by N fertilization studies (Brooks 2003; Rao and Allen 2010). However, this is the first study we are aware of showing strong correlations of annual bromegrass distribution and productivity in response to landscape-scale gradients in inorganic N. Furthermore, the data suggest that fire legacy and cattle via dung deposition significantly contributed to spatial variability in N gradients across the study landscape that appears to influence patterns of red brome growth and reproduction. A critical question left unanswered by this study is: (1) how long does the influence of cowpats on red brome persist across time? We observed partially and fully decomposed cowpats that still appeared to be stimulating red brome growth (Fig. 2), likely through inorganic N enrichment but more research is needed to address this question.
Transgenerational effects
Two factors that influence invasion success are the quantity and quality of seeds produced (Chambers et al. 2016). Due to early germination and rapid growth rates, annual brome grasses vigorously compete for soil resources (Salo et al. 2005), which are utilized for prolific seed production (Rao and Allen 2010). However, a potential tradeoff between the quantity and quality of seeds produced likely varies depending on the resource environment of the parent plant. Our results show that the micro-environment created by cowpats simultaneously increased seed production and seed viability (Figs. 3, 7) likely due to increased soil moisture and N availability (Larigauderie et al. 1988). This suggests that soil resource hotspots may amplify Bromus fecundity by stimulating the production of high numbers of high-quality seeds. This is the first study we are aware of documenting transgenerational increases in seed viability of invasive brome grasses in response to gradients of soil resource availability.
Conclusions
Our results suggest that cattle and fire steepen soil resource gradients that contribute to invasive grass-fire cycles in the Mojave Desert. In the case of cattle, their dung creates soil resource hotspots in resource-limited shrub interspaces that fuels prolific seed production and enhanced seed viability (Figs. 3, 7). The building of red brome seed bank in the inter-shrub space due to prolific seed production next to cowpats (Fig. 3) likely contributes to fine fuel continuity in inter-shrub spaces, particularly in wetter years that spread large-scale wildfires in the Mojave Desert (Brooks et al. 2004; Horn and St Clair 2017). These wildfires facilitated by red brome (Horn and St Clair 2017) appear to create a legacy effect of elevated inorganic N pools in post-fire environments (Esque et al. 2010) that correlate with red brome’s increased growth and reproduction success (Figs. 4, 5). These results suggest that red brome may have the potential to create a positive fire feedback loop mediated by increases in inorganic N that reinforces its growth (Rao and Allen 2010).
While this study was conducted in the Mojave Desert, similar issues exist in the other deserts of North America, where invasive annual brome grasses also overlap with cattle grazing (Esque and Schwalbe 2002; Chambers et al. 2016). Addition studies are needed to explicitly determine the spatio-temporal dynamics of post-fire environments and cowpats on annual brome invasions across deserts of North America.
Data availability
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Code availability
Not applicable.
References
Auman BS, Call CA, Wiedmeier RD (1998) Crested wheatgrass establishment in livestock dung deposited on degraded rangeland vegetation types in the Intermountain West, USA. Arid Land Res Manage 12(4):317–333. https://doi.org/10.1080/15324989809381521
Belnap J, Stark JM, Rau BM, Allen EB, Phillips S (2016) Soil moisture and biogeochemical factors influence the distribution of annual Bromus species. Exotic brome-grasses in arid and semiarid ecosystems of the western US. Springer, Cham, pp 227–256
Bishop TB, Gill RA, McMillan BR, St Clair SB (2020) Fire, rodent herbivory, and plant competition: implications for invasion and altered fire regimes in the Mojave desert. Oecologia 192(1):155–167. https://doi.org/10.1007/s00442-019-04562-2
Bowling GA (1942) The introduction of cattle into colonial North America. J Diary Sci 25:129–154
Brooks ML (2003) Effects of increased soil nitrogen on the dominance of alien annual plants in the Mojave desert. J Appl Ecol 40(2):344–353. https://doi.org/10.1046/j.1365-2664.2003.00789.x
Brooks ML, Matchett JR (2006) Spatial and temporal patterns of wildfires in the Mojave desert, 1980–2004. J Arid Environ 67:148–164. https://doi.org/10.1016/j.jaridenv.2006.09.027
Brooks ML, D’antonio CM, Richardson DM, Grace JB, Keeley JE, DiTomaso J et al (2004) Effects of invasive alien plants on fire regimes. Bioscience 54(7):677–688. https://doi.org/10.1641/0006-3568(2004)054[0677:EOIAPO]2.0.CO;2
Callaway RM (2007) Positive interactions and interdependence in plant communities. Springer Science & Business Media, New York
Chambers J, Germino M, Belnap J, Brown C, Schup E, St. Clair SB (2016) Plant community resistance to invasion by Bromus species: the roles of community attributes, Bromus interactions with plant communities, and Bromus traits. Exotic brome-grasses in arid and semiarid ecosystems of the western US. Springer, Cham, pp 275–304
Chastain JP, Camberato JJ (2004) Dairy manure production and nutrient content. Chapter 3a in Confined Animal Manure Managers Certification Program Manual B Dairy Version 1, pp 1–16
Chuong J, Huxley J, Spotswood E, Nichols L, Mariotte P, Suding K (2016) Cattle as dispersal vectors of invasive and introduced plants in a California annual grassland. Rangel Ecol Manage 69:52–58
Clark PE, Nielson RM, Lee J, Ko K, Johnson DE, Ganskopp DC et al (2017) Prescribed fire effects on activity and movement of cattle in mesic sagebrush steppe. Rangel Ecol Manage 70(4):437–447. https://doi.org/10.1016/j.rama.2015.10.009
Commission for Environmental Cooperation (2015) North American ranching industries, beef cattle trade, and grasslands: status and trends. Commission for Environmental Cooperation, Montreal, Canada, p 38
Dai X (2000) Impact of cattle dung deposition on the distribution pattern of plant species in an alvar limestone grassland. J Veg Sci 11(5):715–724. https://doi.org/10.2307/3236578
Diggle P (1975) Robust density estimation using distance methods. Biometrika 62:39–48. https://doi.org/10.1093/biomet/62.1.39
Dovrat G, Perevolotsky A, Ne’Eman G (2012) Wild boars as seed dispersal agents of exotic plants from agricultural lands to conservation areas. J Arid Environ 78:49–54. https://doi.org/10.1016/j.jaridenv.2011.11.011
Eichberg C, Storm C, Schwabe A (2007) Endozoochorous dispersal, seedling emergence and fruiting success in disturbed and undisturbed successional stages of sheep-grazed inland sand ecosystems. Flora-Morphol Distrib Funct Ecol Plants 202(1):3–26. https://doi.org/10.1016/j.flora.2006.01.004
Esque TC, Schwalbe CR (2002) Alien annual grasses and their relationships to fire and biotic change in Sonoran desert scrub. Invasive exotic species in the Sonoran region. University of Arizona Press and the Arizona-Sonora Desert Museum, Tucson, AZ, pp 165–194
Esque TC, Kaye JP, Eckert SE, DeFalco LA, Tracy CR (2010) Short-term soil inorganic N pulse after experimental fire alters invasive and native annual plant production in a Mojave desert shrubland. Oecologia 164(1):253–263. https://doi.org/10.1007/s00442-010-1617-1
France KA, Ganskopp DC, Boyd CS (2008) Interspace/undercanopy foraging patterns of beef cattle in sagebrush habitats. Rangel Ecol Manage 61(4):389–393. https://doi.org/10.2111/06-072.1
Gill RA, O’Connor RC, Rhodes A, Bishop TB, Laughlin DC, St Clair SB (2018) Niche opportunities for invasive annual plants in dryland ecosystems are controlled by disturbance, trophic interactions, and rainfall. Oecologia 187(3):755–765. https://doi.org/10.1007/s00442-018-4137-z
He WM, Yu GL, Sun ZK (2011) Nitrogen deposition enhances Bromus tectorum invasion: biogeographic differences in growth and competitive ability between China and North America. Ecography 34(6):1059–1066. https://doi.org/10.1111/j.1600-0587.2011.06835.x
Hogan J, Phillips C (2011) Transmission of weed seed by livestock: a review. Anim Prod Sci 51:391–398. https://doi.org/10.1071/AN10141
Horn KJ, St Clair SB (2017) Wildfire and exotic grass invasion alter plant productivity in response to climate variability in the Mojave desert. Landscape Ecol 32(3):635–646. https://doi.org/10.1007/s10980-016-0466-7
Horn KJ, McMillan BR, St Clair SB (2012) Expansive fire in Mojave desert shrubland reduces abundance and species diversity of small mammals. J Arid Environ 77:54–58. https://doi.org/10.1016/j.jaridenv.2011.10.003
Horn KJ, Wilkinson J, White S, St Clair SB (2015) Desert wildfire impacts on plant community function. Plant Ecol 216(12):1623–1634. https://doi.org/10.1007/s11258-015-0546-9
Horn KJ, Bishop TB, St Clair SB (2017) Precipitation timing and soil heterogeneity regulate the growth and seed production of the invasive grass red brome. Biol Invasions 19(4):1339–1350. https://doi.org/10.1007/s10530-016-1348-2
Hulme PE (2009) Trade, transport, and trouble: managing invasive species pathways in an era of globalization. J Appl Ecol 46(1):10–18. https://doi.org/10.1111/j.1365-2664.2008.01600.x
Keeney DR, Nelson DW (1982) Nitrogen—inorganic forms. In: Page AL (ed) Methods of soil analysis: part 2 chemical and microbiological properties, vol 9. Am Soc Agron, Madison, pp 643–698
Kowarik I, Lippe MVD (2008) Pathways in plant invasions. In: Nentwig W (ed) Biological invasions. Springer, Berlin, Heidelberg, pp 29–47
Kumar S, Kumari R, Sharma V, Yadav MP (2016) Genetics of domestication and world-wide introduction of Bos indicus (Zebu) and Bos taurus (Taurine) cattle. Indian J Anim Sci. https://doi.org/10.56093/ijans.v86i12.68482
Larigauderie A, Hilbert DW, Oechel WC (1988) Effect of CO2 enrichment and nitrogen availability on resource acquisition and resource allocation in a grass Bromus mollis. Oecologia 77(4):544–549. https://doi.org/10.1007/BF00377272
López RP, Squeo FA, Armas C, Kelt DA, Gutiérrez JR (2016) Enhanced facilitation at the extreme end of the aridity gradient in the Atacama desert: a community-level approach. Ecology 97(6):1593–1604. https://doi.org/10.1890/15-1152.1
Lucero JE, Callaway RM, Faist AM, Lortie CJ (2021) An unfortunate alliance: Native shrubs increase the abundance, performance, and apparent impacts of Bromus tectorum across a regional aridity gradient. Basic Appl Ecol 57:41–53. https://doi.org/10.1016/j.baae.2021.09.001
Luzuriaga AL, Escudero A, Pérez-García F (2006) Environmental maternal effects on seed morphology and germination in Sinapis arvensis (Cruciferae). Weed Res 46(2):163–174. https://doi.org/10.1111/j.1365-3180.2006.00496.x
Lybbert A, Taylor J, Defranco A, St. Clair SB (2017) Reproductive success of wind, generalist, and specialist pollinated plant species following wildfire in desert landscapes. Int J Wildland Fire 26:1030–1039
Mantua N (2000) The Pacific Decadal Oscillation (PDO). Joint Institute for the Study of the Atmosphere and Ocean (website). http://www.jisao.washington.edu/pdo/. Retrieved from 21 January 2002
Molinari R, Bishop TB, Bekker MF, Kitchen SG, Allphin L, St. Clair SB (2019) Creosote growth rate and reproduction increase in postfire environments. Ecol Evol 9(22):12897–12905. https://doi.org/10.1002/ece3.5771
Mouissie AM, Vos P, Verhagen HMC, Bakker JP (2005) Endozoochory by free-ranging, large herbivores: ecological correlates and perspectives for restoration. Basic Appl Ecol 6(6):547–558. https://doi.org/10.1016/j.baae.2005.03.004
Perrings C, Dehnen-Schmutz K, Touza J, Williamson M (2005) How to manage biological invasions under globalization. Trends Ecol Evol 20:212–215. https://doi.org/10.1016/j.tree.2005.02.011
Poschlod P, Bonn S (1998) Changing dispersal processes in the central European landscape since the last Ice Age: an explanation for the actual decrease of plant species richness in different habitats? Acta Bot Neerl 47:27–44
Rao LE, Allen EB (2010) Combined effects of precipitation and nitrogen deposition on native and invasive winter annual production in California deserts. Oecologia 162(4):1035–1046. https://doi.org/10.1007/s00442-009-1516-5
Rowland AP (1983) An automated method for the determination of ammonium-N in ecological materials. Commun Soil Sci Plant Anal 14(1):49–63. https://doi.org/10.1080/00103628309367341
Salo L (2005) Red brome (Bromus rubens subsp. madritensis) in North America: possible modes for early introductions, subsequent spread. Biol Invasions 7:165–180. https://doi.org/10.1007/s10530-004-8979-4
Salo LF, McPherson GR, Williams DG (2005) Sonoran desert winter annuals affected by density of red brome and soil nitrogen. Am Midl Nat 153(1):95–109. https://doi.org/10.1674/0003-0031(2005)153[0095:SDWAAB]2.0.CO;2
Schlesinger WH, Raikes JA, Hartley AE, Cross AF (1996) On the spatial pattern of soil nutrients in desert ecosystems: ecological archives E077–002. Ecology 77(2):364–374. https://doi.org/10.2307/2265615
Soil Survey Staff (2015) Web soil survey. Natural Resources Conservation Service, United States Department of Agriculture, USA
St. Clair SB, O’Connor R, Gill R, McMillan B (2016) Biotic resistance and disturbance: rodent consumers regulate post-fire plant invasions and increase plant community diversity. Ecology 97(7):1700–1711. https://doi.org/10.1002/ecy.1391
Steinger T, Gall R, Schmid B (2000) Maternal and direct effects of elevated CO2 on seed provisioning, germination, and seedling growth in Bromus erectus. Oecologia 123(4):475–480. https://doi.org/10.1007/s004420000342
Wang J, Wang D, Li C, Seastedt TR, Liang C, Wang L et al (2018) Feces nitrogen release induced by different large herbivores in a dry grassland. Ecol Appl 28(1):201–211. https://doi.org/10.1002/eap.1640
Western Regional Climate Center (2019) NCDC 1981–2010 monthly normals [online], Lytle Ranch, Utah. https://wrcc.dri.edu/cgi-bin/cliMAIN.pl?ut5252. Accessed 3 September 2019
Yahdjian L, Gherardi L, Sala OE (2011) Nitrogen limitation in arid-subhumid ecosystems: a meta-analysis of fertilization studies. J Arid Environ 75(8):675–680. https://doi.org/10.1016/j.jaridenv.2011.03.003
Acknowledgements
We would like to thank the Bureau of Land Management St. George Utah and Lytle Research Preserve for facilitating our work in the Beaver Dam Wash.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
EAS and SIS performed the research, analyzed the data, and co-wrote the article. SBS, BES and SJL helped perform the research and edit the paper. GES and DAS performed the research and compiled the data. SBS designed the study, analyzed the data, and co-wrote the article.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by Yu-Long Feng .
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
St. Clair, E.A., St. Clair, S.I., St. Clair, B.E. et al. Wildfire and cattle legacies on gradients of soil nitrogen underlie patterns of annual brome invasion. Oecologia 203, 361–370 (2023). https://doi.org/10.1007/s00442-023-05470-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-023-05470-2