
Abstract
Invasions of exotic grasses are increasing the spatial extent and temporal frequency of wildfire. Mitigation efforts to prevent wildfire depend on understanding the biophysical agents that drive spatial and temporal distributions of exotic grasses. We examined the growth and reproduction of the exotic grass red brome (Bromus rubens L.) in response to precipitation timing in the fall period, fertile-island topography, and fire history in the Mojave Desert. Tiller density, shoot height, shoot biomass, and seed production were measured. Early precipitation timing (mid-October versus late November) triggered early germination and led to greater red brome density (1.5-fold), shoot height (1.9-fold), shoot biomass (6.8-fold) and seed production (3.9-fold). Soil cores collected from beneath shrubs (fertile-islands) versus shrub inter-spaces also produced red brome with enhanced shoot height (1.5-fold), and seed production (1.9-fold). The legacy effect of wildfires on soils (10 years after the fires) had no significant effect on the growth or reproduction of red brome. Soils beneath shrubs had significantly less gravel and higher mineral nitrogen (N) concentrations than soils from shrub inter-spaces. Soil gravel % was negatively correlated with red brome density, height, biomass and seed production while mineral N tended to have weak positively correlations with red brome density and seed production. These results suggest that the growth and reproduction of red brome is influenced by fall precipitation timing and soil characteristics. Management planning for fire intervention should consider fall precipitation patterns, soil characteristics and shrub topography as important determinants of red brome growth and fire potential.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The introduction of non-native species into new ecosystems has been one of the most significant anthropogenic alterations on planet earth (Vitousek et al. 1997). Introduced species alter biogeochemical cycles, reduce biological diversity and negatively impact economics and human health (Vitousek et al. 1997). Among the most widespread and ecologically problematic invasions is the introduction and expansion of exotic grasses in arid ecosystems (D’Antonio and Vitousek 1992; Brooks et al. 2004a). The major deserts of North America are invaded by the exotic grasses red brome (Bromus rubens L.) and cheatgrass (Bromus tectorum L.) (Salo 2005; Rice et al. 2008; Balch et al. 2013). These Eurasian originating species are associated with increased fire extent and frequency in the Great Basin and Mojave Deserts (Brooks et al. 2004a; Bukowski and Baker 2012; Balch et al. 2013). Exotic grasses often fill spaces between shrubs creating continuous fuels that can cause large-scale fire. By altering the disturbance regime, these exotic grasses remove long-lived native species and increase competition for soil resources, which has cascading effects on the biological community and its function (Huenneke and Noble 1996; Horn et al. 2015b; St. Clair et al. 2016).
Wildfire has the capacity to drastically alter the resilience of desert plant communities. Many desert shrubs are sensitive to fire due to prolonged and historically episodic recruitment of native perennials (Callison et al. 1985; Lei 1999; Haubensak et al. 2009; Germino et al. 2016). The loss of native perennial shrubs following fire is further exacerbated by the positive response that exotic grasses have to burned landscapes that can lead to invasive grass fire cycles (Brooks and Berry 2006; Brooks 2012; Abella et al. 2012; Horn and St. Clair 2017). Emergence of invasive grass-fire regimes, with more frequent fire, has caused and will likely continue to promote state changes in vegetation that can fundamentally alter functional interactions within the biological community (Bestelmeyer et al. 2003; Brooks and Matchett 2006; Brooks 2012; Provencher et al. 2016; Horn and St. Clair 2017). The ability of invasive grass-fire regimes to transform entire landscapes makes understanding the mechanisms driving the growth and expansion of exotic grasses a high priority for management intervention (Huenneke and Noble 1996; Vitousek et al. 1997; Rice et al. 2008).
The magnitude and distribution of precipitation are critical factors in the germination and growth potential of invasive brome grasses (Beatley 1974; Salo 2004; Meyer and Allen 2009; Horn et al. 2015a). Bromus spp. can germinate in spring, fall, and winter when temperatures are optimal for a sufficient time-span and soil moisture is favorable (Allen et al. 1995; Roundy et al. 2007; Horn et al. 2015a; Chambers et al. 2016). Fall and winter precipitation enhance the production of exotic annual grasses in deserts of North America (Westerling et al. 2003).Unlike cheatgrass, red brome has not been reported to undergo secondary dormancy, and precipitation and temperature conditions are most favorable for red brome germination in fall and winter in the Mojave Desert (Salo 2004; Bykova and Sage 2012; Jurand et al. 2013; Horn et al. 2015a). High productivity of Bromus spp. also appears to be correlated with episodic wet periods associated with ENSO (El Niño–Southern Oscillation) and PDO (Pacific Decadal Oscillation) cycles in which optimal temperatures overlap with high moisture resulting in a longer growing season (Mack and Pyke 1983; Salo 2004; Bykova and Sage 2012; Abella et al. 2012; Horn et al. 2015a). What is lacking is an understanding of how fall precipitation timing influences the growth and reproductive potential of exotic grasses and its potential for altering fire behavior. With the continued expansion of invasive plants across desert systems and impending shifts in precipitation and temperature with climate change, it is critical that we understand the influence of precipitation timing with greater precision.
The spatial distribution of exotic grasses is influenced by soil properties that can be highly heterogeneous across desert landscapes (Schlesinger and Pilmanis 1998; Brooks 2009; Miller et al. 2009; Reisner et al. 2013). Through its influence on water and nutrient availability, soil characteristics strongly influences plant function and plant community development in deserts (Noy-Meir 1973; Schlesinger and Pilmanis 1998; Austin et al. 2004). Exotic grasses tend to respond positively to increases in soil moisture and nutrients, particularly nitrogen (N) (Brooks and Berry 2006; Hunter and Omi 2006; Abella et al. 2011a). However, the specific effects of soil texture, and its interactions with precipitation timing, on the development and reproduction of red brome are not well documented.
Soil resource availability in desert shrublands is spatially variable based on “fertile islands” that develop beneath shrubs (Havstad et al. 2006). Long-lived shrubs enrich underlying soils with plant-accumulated organic matter, finer-textured soil, and improved moisture conditions through shading (Walker et al. 2001; Ewing et al. 2007). Many plants preferentially establish within fertile islands and exotic grasses are no exception (Brooks 1999a; Walker et al. 2001; Drezner 2006; Abella 2010). Fertile islands have substantially higher concentrations of nutrients which facilitates plant establishment (Brooks 1999a, b). Disturbance such as fire removes shrub canopies but the legacy effects of fertile-islands soils can persist (Ravi and D’Odorico 2009). While it is generally understood that fertile islands have positive effects on plant establishment and growth, previous research has not examined how soil conditions and chemistry beneath desert shrubs affect annual invasive grass growth independent from shade effects.
Fire can have important influences on soil processes that feedback on plant growth. Brome densities can increase following fire (Shinneman and Baker 2009; Brooks 2012; Abella et al. 2012). Fire can increase plant available nutrients by modifying soil structure and texture through secondary effects such as increased erosion (Lei 1999; Esque et al. 2010; Allen et al. 2011). Additionally, shrub mortality decreases competition for soil resources (Phillips and MacMahon 1981). However, the effects of fire on desert soil processes are inconsistent (Allen et al. 2011) and its feedbacks on exotic grass emergence, growth and reproduction are not well characterized.
The objective of this study was to identify biophysical variables that influence the growth and reproduction potential of red brome, which is a dominant plant invader in the Mojave and Sonoran Deserts and has potential for expansion into the Great Basin Desert. We examined the growth and reproductive responses of red brome to precipitation timing (mid-October vs. late November precipitation pulse), soil heterogeneity (soil texture and fertile island topography), and fire history (soils from burned vs. unburned landscapes) to test the legacy effects of fire on invasive annual grass growth. The study was conducted in the northeastern region of Mojave Desert that has recently experienced widespread wildfires driven by the invasion of red brome. We hypothesize that: (1) early fall precipitation will stimulate red brome density, growth and reproduction; and (2) shrub-islands, and fire-disturbed soils would increase red brome growth and reproduction.
Materials and methods
Site description
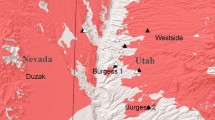
The soil cores used in this study were collected in the summer of 2014 in the Beaver Dam Wash (37.15 °N, 114.01 °W) located in the northeastern region of the Mojave Desert in the southwest corner of Utah, USA (Fig. 1). The experiment was conducted at Lytle Research Preserve which is centrally located in the Beaver Dam Wash. Soils are described as a very gravelly sandy loam but textures within that description vary depending on location (Soil Survey Staff 2011). Vegetation was typical of mid-elevation (~1000 m) Mojave shrubland dominated by creosote bush (Larrea tridentata (DC) Cov.), blackbrush (Coleogyne ramosissima Torr.), and white bursage (Ambrosia dumosa (A. Gray) Payne). Joshua trees (Yucca brevifolia Engelm.) were also prevalent. The most common native forbs and grasses across the landscape were desert marigold (Baileya multiradiata), desert globemallow (Sphaeralcea ambigua), and low woollygrass (Dasyochloa pulchella). Red brome was nearly ubiquitous across the study landscape from which the cores were collected. The prevalence of red brome likely contributed to the expansion of recent wildfires in the northeastern Mojave Desert (Brooks and Matchett 2006; Horn et al. 2015a). Since 1984 over 550,000 hectares have burned in the Mojave Desert (Brooks and Matchett 2006). The majority of this occurred in 2005 (385,357 ha). These fires were highly concentrated in the northeastern region of the Mojave. In 2005 the Beaver Dam Wash experienced 4 independent wildfires: Westside (23,782 ha), Duzak (132,395 ha), Burgess 1 (60 ha) and Burgess 2 (543 ha). The soil cores were collected along the boundaries of these fires.

Map of the study landscape showing the 4 wildfires (Duzak, Burgess1, Burgess2 and Westside with darker shaded polygon) that occurred in 2005. Soil cores were collected from burned landscapes (triangles) and in adjacent unburned islands or landscapes (circles)
Experimental design
We tested the growth responses of red brome to precipitation timing, shrub-island topography, and fire history in a full-factorial design, replicated 8 times on soil cores collected along the boundaries of the four wildfires that occurred in the summer of 2005 (Fig. 1) for a total of 64 samples. We collected soil cores from two burned and two unburned locations for each of the 4 wildfires. Explanatory variables included shrub-island topography, fire history, and the timing of water addition. Soil cores were collected from beneath current or former (burned areas) shrub canopies of creosote bush and adjacent shrub interspaces, nested within adjacent burned or unburned areas across the burn boundaries of the four fires that occurred in 2005 (Fig. 1). Precipitation timing treatments occurred 6 weeks apart in either mid-October or late November.
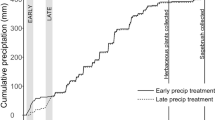
Soil cores were extracted using PVC tubing (10 cm diameter, 20 cm length) driven into the soil with a wooden block and a rubber mallet in early September 2014. Soil surrounding the cylinders was excavated and cores were retrieved with the soil column intact. Mesh screen was secured around the bottom of each core to prevent the soil from shifting and to allow water to flow naturally through the bottom of the core. In mid-September the soil cores were watered for 10 days to saturation to promote germination of the seed bank. Seedlings that established from seed bank were removed carefully by hand to eliminate any background seed bank that was present in the collected soil cores. The soil cores were allowed to dry completely for 3 weeks in preparation for the experiment. The experiment was organized by randomizing the soil cores with all treatment combinations in each of the 8 experimental blocks. A 1-m tall mesh screen fence was placed around the perimeter of the experiment to prevent rodent activity in the experimental plots. Forty red brome seeds were placed on the top of each soil core immediately before initial watering. The seeds were collected from several locations around the Lytle Preserve and they were then mixed thoroughly. The early precipitation cores starting receiving watering treatments October 13, 2014 and the late precipitation cores received their 10-day watering period 6 weeks later starting on November 24, 2014 (Fig. 2). Very little ambient precipitation fell in the six-week period between October 13 and November 24, so initial water timing was the only significant difference between watering treatments (Fig. 2). During the 10-day watering periods 1 cm of water (medium size storm amount) was added each day in the morning. To avoid seed loss due to wind or seed predation by insect, or germination in response to natural precipitation we did not add red brome seed to the late watering soil cores until November 24, the first day of the late watering treatment. The watering totals were based on the precipitation amounts received in October 2004 in the Beaver Dam Wash as recorded at Badger Springs meteorological station (mesowest.utah.edu) and at Brigham Young University’s Lytle Ranch Preserve (https://climate.usurf.usu.edu).

Comparisons of accumulated precipitation between years (2004–2005 vs. 2014–2015) and early watering versus late watering in 2014–2015 with long dashes representing the early precipitation regimes and short dashes indicating the late precipitation timing. Minimal precipitation in the Beaver Dam Wash between October 13 and November 24, 2014 resulted in minimal differences between total accumulated precipitation in the timing treatments. The early precipitation treatment of 2014–2015 very closely mirrored the timing and initial rate of October precipitation back in 2004 prior to the 2005 fires
Plant measurements
To quantify the effects of treatment conditions on red brome growth and reproduction we measured total number of tillers, shoot biomass, shoot height and seed production. The experiment was terminated at the end of April 2015 when the plants reached their reproductive peak. Average plant height was measured from the soil surface on 10 randomly selected tillers in each core. The relationship between seed head mass and seed number was determined using a regression equation (R2 = 0.96) allowing us to determine total seed production from counts of seed heads. The above ground plant biomass was then harvested, stored in paper sacks, dried for 72 h in a drying oven at 60 °C and weighed. All count data and mass measurements were converted to a per-area basis (divided by surface area of the soil core).
Soil texture and nutrient analysis
Soil cores were homogenized, gravel and larger aggregates were separated and weighed, and the remaining soil fraction was measured for texture and mineral N. Soil texture was determined using a hydrometer and analyzed using percent content of sand, silt, and clay by mass. Ammonium and nitrate were extracted for each soil core using potassium chloride and analyzed undiluted by Flow Injection Analysis (Quick Chem 8500, LACHAT Instruments, Loveland, Colorado, USA) as outlined in protocols developed by Keeney and Nelson (1982).
Statistical analysis
To characterize the effects of precipitation timing, shrub-island topography (beneath shrub vs. shrub interspace) and fire history (burned vs. unburned areas) on red brome growth and seed production, we used a 3-way ANOVA mixed models analysis with precipitation timing, shrub-island topography, and fire history as fixed effects and site as a random effect. Similarly, we analyzed soil texture and mineral N content against shrub-island topography and fire history in a 2-way ANOVA with site as a random effect. Data exploration was conducted according to the methods of Zuur et al. (2010) to test that model assumptions were met. Pearson correlation coefficient was used to examine the relationship between gravel, sand, silt, clay, and mineral N with the growth and reproduction of red brome. Statistics were performed in Program R version 2.14.1 (R Foundation for Statistical Computing, Vienna, Austria).
Results
Variation in soil texture and mineral N
Soil texture and soil mineral N differed with shrub-island topography but no differences in soil characteristics were observed between burned and unburned areas. Soils from shrub interspaces contained 65% more gravel than soils from beneath shrub canopies (Fig. 3a); however, after the gravel fraction was removed, no other soil texture differences were apparent (Fig. 3b–d). Mineral N was 5 times greater beneath shrub canopies than in shrub interspaces however. Differences in mineral N in soil beneath fertile-island versus shrub interspaces was greater in unburned landscapes compared to burned areas (Fig. 3e).

Soil characteristics associated with shrub-island topography and fire history. a Percent gravel was removed before calculating, b sand, c silt, and d clay fractions of the soil, and before determining, e soil mineral nitrogen (N) content. F-values presented with asterisks indicating level of significance for the main effects and interaction term of fire history and shrub topography. Asterisks indicate the following P value levels: *P ≤ 0.05, **P ≤ 0.001, ***P ≤ 0.0001. Mean values presented with ± 1 SE
Effects of precipitation timing, shrub topography and fire history
Early precipitation timing and soils collected beneath shrubs generally had positive effects on red brome productivity while fire history did not. Early precipitation timing significantly increased red brome tiller density (1.5-fold), shoot height (1.9-fold), shoot biomass (6.8-fold) and seed production (3.9-fold) relative to cores that received precipitation starting 6 weeks later in late November (Figs. 4, 5). Soil cores collected beneath shrubs significantly increased shoot height (1.5-fold), and seed production (1.9-fold) relative to cores collected from shrub interspaces (Fig. 5). The positive effect of early precipitation and fertile islands were additive as indicated by the lack of significance of their interaction term (Fig. 5).

Repeat photos of a single block in our experiment in January (top photo) and again in April (bottom photo) 2015 showing that when red brome received rain in mid-October (4 corner cores) versus late November (4 center cores) that it was much more aggressive as evidenced by greater height, tiller density and seed production. Note soil core positions within blocks were randomized; we used this picture because by chance the positions of the precipitation treatment were separated by center cores and corner cores so it is easy to see the precipitation timing effect

Effects of precipitation timing (October vs. November), shrub-island topography (beneath shrub vs. shrub interspace), and fire history on tiller density, shoot height, shoot biomass and seed production of red brome. F-values presented with asterisks indicating level of significance for the main effects and interaction term of precipitation timing and shrub topography. Asterisks indicate the following P value levels: *P ≤ 0.05, **P ≤ 0.001, ***P ≤ 0.0001. Mean values presented with ± 1 SE
Growth and seed production in red brome were negatively correlated with percent gravel content (Table 1). Tiller density and plant height (P = 0.07) showed weak positive correlations with mineral N content (Table 1). Fractions of sand, silt and clay showed no significant correlations with red brome productivity.
Discussion
The spread of wildfire is largely dictated by the distribution and density of vegetation (Bond and Keeley 2005). In native arid shrublands of North America, density and connectivity of vegetation are typically insufficient to create large scale, and frequent wildfires (Humphrey 1974). Invasion of exotic annual grasses has increased the density and connectivity of fuels in arid shrublands facilitating expansive fires and altering the ecological state of arid shrublands to arid annual grasslands (D’Antonio and Vitousek 1992; Brooks 1999b; Brooks and Matchett 2006; Bukowski and Baker 2012). However, the distribution of these exotic grasses (and subsequently the extent and frequency of fire) is not uniform across landscapes, and is influenced by weather patterns, soil properties, and fire history (Brooks 2012; Abella et al. 2012). The objective of this study was to experimentally determine how fall precipitation timing, fertile island topography and fire history influence the growth and reproduction of the exotic invasive grass, red brome. Our data indicate that the timing of fall precipitation and fertile island topography strongly affect the growth and reproductive potential of red brome, while the legacy effects of wildfire mediated through soil properties are less important.
Timing of fall precipitation
Timing of fall precipitation greatly affected plant density, growth and seed production of red brome. High abundances of red brome and large fires have been preceded by intense and prolonged precipitation events (Brooks and Matchett 2006; Horn et al. 2015a; Horn and St. Clair 2017). However, experimental evidence is lacking on the effects of precipitation timing on invasive annual grass growth. Our results indicate that precipitation timing caused significant increases in grass densities, height, and biomass (Figs. 4, 5). Not only is the growing season extended by earlier precipitation but germination success is also increased (Fig. 5a; Horn et al. 2015a). The positive effects of earlier fall precipitation are likely driven by precipitation arriving when temperatures are optimal for germination, which for red brome is approximately 19 °C with maximums not to exceed 30 °C (Horn et al. 2015a). Our data suggests that early fall precipitation events are likely to promote vegetative growth and increased seed production of red brome, which is likely to increase the probability of desert fire.
Fertile-island soils increase red brome densities and localized fuel
Fertile islands associated with desert shrubs contribute to the distribution and maintenance of red brome populations. Succession in the Mojave Desert and other arid systems is strongly dependent on services provided by nurse shrubs (Abella 2010), and it seems no different for red brome (Brooks 1999a). Enhanced plant growth in fertile islands is due in part to soil properties, as we observed increased growth and reproduction on fertile-island soils in the absence of shrub canopies (Fig. 5). In particular, soil mineral nitrogen was noticeably higher in fertile islands (Fig. 3), and nitrogen strongly stimulated red brome growth in several other studies (Noy-Meir 1973; Hooper and Johnson 1999; Brooks 2003; Ewing et al. 2007). Finer textured surface soils of fertile islands (less gravel) can also increase moisture availability to plants relative to coarser-texture inter-shrub soils (Walker et al. 2001; Austin et al. 2004). Greater soil resources and shading by the shrub canopies can increase plant establishment beneath shrub canopies (Schlesinger and Pilmanis 1998; Brittingham and Walker 2000; Walker et al. 2001; Drezner 2006). Once established beneath shrub canopies, seed sources from fertile islands allow expansion of red brome into inter-shrub spaces when moisture conditions become favorable enough for growth on coarser textured soils.
Wildfire effects on red brome growth
Increases in red brome densities have been documented in the Mojave Desert following wildfire (Brooks 2012; Horn and St. Clair 2017). Results from time-series analyses of satellite data have also indicated that red brome increases on burned landscapes and can dominate even several years after fire (Horn and St. Clair 2017). These increases could be induced by nutrient pulses from combusted biomass (Esque et al. 2010). However, the long term persistence of fire-induced nutrient pulses is uncertain (Allen et al. 2011). We were unable to detect any difference in red brome growth or soil nutrients between soils in burned and unburned landscapes 10 years after wildfires. However, increases in red brome densities post-fire may be from competition release due to the loss of native plant cover (as observed for surviving native shrubs) since exotic grasses compete with native annuals especially during years of high productivity (Brooks 2000; Horn et al. 2015b).
Soil influences on red brome
Spatial variability in soil texture and nutrient availability likely influences fuel continuities of red brome. Positive correlations of mineral N content with red brome growth supports other studies showing N limitation of red brome in the Mojave Desert (Brooks 2003). Gravel content was the most important soil factor influencing red brome success in our study (Table 1). Coarser-textured surface soils have been shown to decrease emergence of red brome (Abella et al. 2011b), a patterns consistent with our data (Table 1). Soil gravel content was significantly higher in shrub interspaces (Fig. 3a–d), and these interspaces maintained significantly shorter grasses and produced fewer seeds compared to soil from beneath shrub canopies (Figs. 3, 5). High gravel context can drastically reduce the water holding capacity of soils (Young and Nobel 1986; Paruelo et al. 1988; Brakensiek and Rawls 1994). An example of this is the development of desert pavement, which reduces water infiltration and eliminates the presence of most vegetation (Young et al. 2004). We suggest that soils with high gravel context will have lower red brome fuel loads, potentially creating natural fire breaks across soil texture gradients.
Significance and recommendations
The potential and likely emergence of an invasive-plant-driven fire regime, and its long-term negative impacts of altering landscape cover type, has made the need for mitigation imperative in order to preserve native plant communities of arid ecosystems. Positive response of red brome to fire (Horn and St. Clair 2017), the negative effects of fire on native plant communities of deserts (Callison et al. 1985; Lei 1999; Engel and Abella 2011), and the prolonged recovery rates of native perennials, make prevention of the initial fire a primary goal in preventing the establishment of invasive grass-driven fire regimes. Soil surveys can be used to identify soil conditions where the growth of red brome and the potential for wildfire is high (Brooks 1999a; Brooks et al. 2004b). The risk of fire in arid shrublands increases when moisture arrives early in the fall (Fig. 5), is sustained throughout the winter, and where the soil texture of inter-shrub spaces is fine enough to maintain growth and increased biomass of fine fuels. As precipitation increases, the potential for coarser-textured soils in shrub inter-spaces to have fire-carrying fuel loads increases as well (Fig. 5). Additionally, previously burned areas can be at increased risk for subsequent burning due to high fine fuel loads (Brooks 2012; Horn and St. Clair 2017). There is evidence that post-fire revegetation of native plants may compete with red brome and limit its abundance on post-fire landscapes (Jurand and Abella 2013). Based on our data we suggest that anomalous fall precipitation events will further increase the likelihood of expansive fires. Active mitigation against such fires is necessary in order for native perennial restoration and preservation efforts to succeed (Webb et al. 2009; Brooks 2012).
References
Abella SR (2010) Disturbance and plant succession in the Mojave and Sonoran Deserts of the American Southwest. Int J Environ Res Public Health 7:1248–1284. doi:10.3390/ijerph7041248
Abella SR, Craig DJ, Chiquoine LP et al (2011a) Relationships of Native desert plants with red brome (Bromus rubens): toward identifying invasion-reducing species. Invasive Plant Sci Manag 4:115–124. doi:10.1614/IPSM-D-10-00013.1
Abella SR, Lee AC, Suazo AA (2011b) Effects of burial depth and substrate on the emergence of Bromus rubens and Brassica tournefortii. Bull South Calif Acad Sci 110:17–24. doi:10.3160/0038-3872-110.1.17
Abella SR, Embrey TM, Schmid SM, Prengaman KA (2012) Biophysical correlates with the distribution of the invasive annual red brome (Bromus rubens) on a Mojave Desert Landscape. Invasive Plant Sci Manag 5:47–56. doi:10.1614/IPSM-D-11-00030.1
Allen PS, Meyer SE, Beckstead J (1995) Patterns of seed after-ripening in Bromus tectorum L. J Exp Bot 46:1737–1744. doi:10.1093/jxb/46.11.1737
Allen EB, Steers RJ, Dickens SJ (2011) Impacts of fire and invasive species on desert soil ecology. Rangel Ecol Manag 64:450–462. doi:10.2111/REM-D-09-00159.1
Austin AT, Yahdjian L, Stark JM et al (2004) Water pulses and biogeochemical cycles in arid and semiarid ecosystems. Oecologia 141:221–235. doi:10.1007/s00442-004-1519-1
Balch JK, Bradley BA, D’Antonio CM, Gómez-Dans J (2013) Introduced annual grass increases regional fire activity across the arid western USA (1980–2009). Glob Change Biol 19:173–183. doi:10.1111/gcb.12046
Beatley JC (1974) Effects of rainfall and temperature on the distribution and behavior of Larrea tridentata (creosote-bush) in the Mojave Desert of Nevada. Ecology 55:245–261. doi:10.2307/1935214
Bestelmeyer BT, Brown JR, Havstad KM et al (2003) Development and use of State-and-Transition models for rangelands. J Range Manage 56:114–126. doi:10.2307/4003894
Bond W, Keeley J (2005) Fire as a global “herbivore”: the ecology and evolution of flammable ecosystems. Trends Ecol Evol 20:387–394. doi:10.1016/j.tree.2005.04.025
Brakensiek DL, Rawls WJ (1994) Soil containing rock fragments: effects on infiltration. CATENA 23:99–110. doi:10.1016/0341-8162(94)90056-6
Brittingham S, Walker RL (2000) Facilitation of Yucca brevifolia recruitment by Mojave Desert shrubs. West North Am Nat 60:374–383
Brooks ML (1999a) Habitat invasibility and dominance by alien annual plants in the western Mojave Desert. Biol Invasions 1:325–337. doi:10.1023/A:1010057726056
Brooks ML (1999b) Alien annual grasses and fire in the Mojave Desert. Madrono 46:13–19
Brooks ML (2000) Competition between alien annual grasses and native annual plants in the Mojave Desert. Am Midl Nat 144:92–108
Brooks ML (2003) Effects of increased soil nitrogen on the dominance of alien annual plants in the Mojave Desert. J Appl Ecol 40:344–353. doi:10.1046/j.1365-2664.2003.00789.x
Brooks ML (2012) Effects of high fire frequency in creosote bush scrub vegetation of the Mojave Desert. Int J Wildland Fire 21:61–68
Brooks ML, Berry KH (2006) Dominance and environmental correlates of alien annual plants in the Mojave Desert, USA. J Arid Environ 67(Supplement):100–124. doi:10.1016/j.jaridenv.2006.09.021
Brooks ML, Matchett JR (2006) Spatial and temporal patterns of wildfires in the Mojave Desert, 1980–2004. J Arid Environ 67:148–164
Brooks ML, D’Antonio CM, Richardson DM et al (2004a) Effects of invasive alien plants on fire regimes. BioScience 54:677. doi:10.1641/0006-3568(2004)054[0677:EOIAPO]2.0.CO;2
Brooks ML, Matchett J, Wallace C, Esque T (2004b) Fuels mapping and fire hazard assessment in a desert ecosystem. Arid Lands Newsl 55:1–5
Brooks ML et al (2009) Spatial and temporal distribution of nonnative plants in upland areas of the Mojave Desert. In: Webb RH, Fenstermaker LF, Heaton JS (eds) The Mojave Desert: ecosystem processes and sustainability. University of Nevada Press, Reno, pp 101–124
Bukowski BE, Baker WL (2012) Historical fire regimes, reconstructed from land-survey data, led to complexity and fluctuation in sagebrush landscapes. Ecol Appl 23:546–564. doi:10.1890/12-0844.1
Bykova O, Sage RF (2012) Winter cold tolerance and the geographic range separation of Bromus tectorum and Bromus rubens, two severe invasive species in North America. Glob Change Biol 18:3654–3663. doi:10.1111/gcb.12003
Callison J, Brotherson JD, Bowns JE (1985) The effects of fire on the blackbrush [Coleogyne ramosissima] community of southwestern Utah. J Range Manag 38:535–538
Chambers JC, Germino MJ, Belnap J et al (2016) Plant community resistance to invasion by Bromus species: the roles of community attributes, Bromus interactions with plant communities, and Bromus traits. In: Germino MJ, Chambers JC, Brown CS (eds) Exotic brome-grasses in arid and semiarid ecosystems of the western US. Springer, Berlin, pp 275–304
D’Antonio CM, Vitousek PM (1992) Biological invasions by exotic grasses, the grass/fire cycle, and global change. Annu Rev Ecol Syst 23:63–87. doi:10.2307/2097282
Drezner TD (2006) Regeneration of Carnegiea gigantea (Cactaceae) since 1850 in three populations in the northern Sonoran Desert. Acta Oecol 29:178–186. doi:10.1016/j.actao.2005.09.005
Engel EC, Abella SR (2011) Vegetation recovery in a desert landscape after wildfires: influences of community type, time since fire and contingency effects. J Appl Ecol 48:1401–1410. doi:10.1111/j.1365-2664.2011.02057.x
Esque TC, Kaye JP, Eckert SE et al (2010) Short-term soil inorganic N pulse after experimental fire alters invasive and native annual plant production in a Mojave Desert shrubland. Oecologia 164:253–263. doi:10.1007/s00442-010-1617-1
Ewing SA, Southard RJ, Macalady JL et al (2007) Soil microbial fingerprints, carbon, and nitrogen in a Mojave Desert creosote-bush ecosystem. Soil Sci Soc Am J 71:469–475
Germino MJ, Belnap J, Stark JM et al (2016) Ecosystem impacts of exotic annual invaders in the genus Bromus. In: Germino MJ, Chambers JC, Brown CS (eds) Exotic brome-grasses in arid and semiarid ecosystems of the western US. Springer, Berlin, pp 61–95
Haubensak K, D’Antonio C, Wixon D (2009) Effects of fire and environmental variables on plant structure and composition in grazed salt desert shrublands of the Great Basin (USA). J Arid Environ 73:643–650. doi:10.1016/j.jaridenv.2008.12.020
Havstad KM, Huenneke LF, Schlesinger WH (2006) Structure and function of a Chihuahuan Desert ecosystem: the Jornada Basin long-term ecological research site. Oxford University Press, Oxford
Hooper DU, Johnson L (1999) Nitrogen limitation in dryland ecosystems: responses to geographical and temporal variation in precipitation. Biogeochemistry 46:247–293
Horn KJ, St. Clair SB (2017) Wildfire and exotic grass invasion alter plant productivity in response to climate variability in the Mojave Desert. Landsc Ecol. doi:10.1007/s10980-016-0466-7 (In press)
Horn KJ, Nettles R, St. Clair SB (2015a) Germination response to temperature and moisture to predict distributions of the invasive grass red brome and wildfire. Biol Invasions. doi:10.1007/s10530-015-0841-3
Horn KJ, Wilkinson J, White S, St. Clair SB (2015b) Desert wildfire impacts on plant community function. Plant Ecol 216:1623–1634. doi:10.1007/s11258-015-0546-9
Huenneke LF, Noble I (1996) Ecosystem function of biodiversity in arid ecosystems. In: Mooney HA, International Council of Scientific Unions, United Nations Environment Programme (eds) Functional roles of biodiversity: A global perspective. Wiley, Chichester, New York, pp 99–128
Humphrey RR (1974) Fire in the deserts and desert grasslands of North America. In: Kozlowski TT, Ahlgren CE (eds) Fire and ecosystems. USA Academic Press, New York, pp 365–401
Hunter ME, Omi PN (2006) Response of native and exotic grasses to increased soil nitrogen and recovery in a postfire environment. Restor Ecol 14:587–594
Jurand BS, Abella SR (2013) Soil seed banks of the exotic annual grass Bromus rubens on a burned desert landscape. Rangel Ecol Manag 66:157–163. doi:10.2111/REM-D-12-00106.1
Jurand BS, Abella SR, Suazo AA (2013) Soil seed bank longevity of the exotic annual grass Bromus rubens in the Mojave Desert, USA. J Arid Environ 94:68–75. doi:10.1016/j.jaridenv.2013.03.006
Keeney D, Nelson D (1982) Nitrogen—inorganic forms. In: Page A (ed) Methods of soil analysis part 2. Madison, WI, pp 643–698
Lei SA (1999) Postfire woody vegetation recovery and soil properties in blackbrush (Coleogyne ramosissima Torr.) shrubland ecotones. J Ariz-Nev Acad Sci 32:105–115
Mack RN, Pyke DA (1983) The demography of bromus tectorum: variation in time and space. J Ecol 71:69–93. doi:10.2307/2259964
Meyer SE, Allen PS (2009) Predicting seed dormancy loss and germination timing for Bromus tectorum in a semi-arid environment using hydrothermal time models. Seed Sci Res 19:225–239
Miller DM, Bedford DR, Hughson DL et al (2009) Mapping mojave desert ecosystem properties with surficial geology. In: Webb RH, Fenstermaker LF, Heaton JS et al. (eds) The mojave desert: ecosystem processes and sustainability. University of Nevada Press, Reno, Nevada 89577 USA, pp 225–251
Noy-Meir I (1973) Desert ecosystems: environment and producers. Annu Rev Ecol Syst 4:25–51
Paruelo JM, Aguiar MR, Golluscio RA (1988) Soil water availability in the Patagonian arid steppe: gravel content effect. Arid Soil Res Rehabil 2:67–74. doi:10.1080/15324988809381159
Phillips DL, MacMahon JA (1981) Competition and spacing patterns in desert shrubs. J Ecol 69:97–115. doi:10.2307/2259818
Provencher L, Frid L, Czembor C, Morisette JT (2016) State-and-transition models: conceptual versus simulation perspectives, usefulness and breadth of use, and land management applications. In: Germino MJ, Chambers JC, Brown CS (eds) Exotic brome-grasses in arid and semiarid ecosystems of the western US. Springer, Berlin, pp 371–407
Ravi S, D’Odorico P (2009) Post-fire resource redistribution and fertility island dynamics in shrub encroached desert grasslands: a modeling approach. Landsc Ecol 24:325–335. doi:10.1007/s10980-008-9307-7
Reisner MD, Grace JB, Pyke DA, Doescher PS (2013) Conditions favouring Bromus tectorum dominance of endangered sagebrush steppe ecosystems. J Appl Ecol 50:1039–1049. doi:10.1111/1365-2664.12097
Rice PM, McPherson GR, Rew LJ (2008) Fire and nonnative invasive plant in the interrior west bioregion. In: Zouhar K, Smith JK, Sutherland S, Brooks ML (eds) Wildland fire in ecosystems: fire and nonnative invasive plants. US Deparmtent of Agriculture, Forest Service, Rocky Mountain Research Station, Ogden, pp 141–171
Roundy BA, Hardegree SP, Chambers JC, Whittaker A (2007) Prediction of cheatgrass field germination potential using wet thermal accumulation. Rangel Ecol Manag 60:613–623. doi:10.2111/05-206R3.1
Salo LF (2004) Population dynamics of red brome (Bromus madritensis subsp. rubens): times for concern, opportunities for management. J Arid Environ 57:291–296. doi:10.1016/S0140-1963(03)00110-1
Salo LF (2005) Red brome (Bromus rubens subsp. madritensis) in North America: possible modes for early introductions, subsequent spread. Biol Invasions 7:165–180. doi:10.1007/s10530-004-8979-4
Schlesinger WH, Pilmanis AM (1998) Plant-soil interactions in deserts. Biogeochemistry 42:169–187. doi:10.1023/A:1005939924434
Shinneman DJ, Baker WL (2009) Environmental and climatic variables as potential drivers of post-fire cover of cheatgrass (Bromus tectorum) in seeded and unseeded semiarid ecosystems. Int J Wildland Fire 18:191–202
Soil Survey Staff (2011) Web soil survey. Natural Resources Conservation Service, United States Department of Agriculture. http://websoilsurvey.nrcs.usda.gov. Accessed 1 May 2013
St. Clair SB, O’Connor R, Gill R, McMillan B (2016) Biotic resistance and disturbance: rodent consumers regulate post-fire plant invasions and increase plant community diversity. Ecology 97:1700–1711. doi:10.1002/ecy.1391
Vitousek PM, D’Antonio CM, Loope LL et al (1997) Introduced species: a significant component of human-caused global change. N Z J Ecol 21:1–16
Walker LR, Thompson DB, Landau FH (2001) Experimental manipulations of fertile islands and nurse plant effects in the Mojave Desert, USA. West North Am Nat 61:25–35
Webb RH, Belnap J, Thomas KA et al (2009) Natural recovery from severe disturbance in the Mojave Desert. In: Webb RH, Fenstermaker LF, Heaton JS (eds) The Mojave Desert: ecosystem processes and sustainability. University of Nevada Press, Reno, pp 343–377
Westerling AL, Gershunov A, Brown TJ et al (2003) Climate and wildfire in the western United States. Bull Am Meteorol Soc 84:595–604. doi:10.1175/BAMS-84-5-595
Young DR, Nobel PS (1986) Predictions of soil-water potentials in the north-western Sonoran Desert. J Ecol 74:143–154. doi:10.2307/2260355
Young MH, McDonald EV, Caldwell TG et al (2004) Hydraulic properties of a desert soil chronosequence in the Mojave Desert, USA. Vadose Zone J 3:956–963. doi:10.2113/3.3.956
Zuur AF, Ieno EN, Elphick CS (2010) A protocol for data exploration to avoid common statistical problems. Methods Ecol Evol 1:3–14. doi:10.1111/j.2041-210X.2009.00001.x
Acknowledgements
We would like to acknowledge Dr. Zachary Aanderud and Dr. Steven Petersen for their review of this manuscript along with Earl Hansen, Rachel Nettles, Nathan Duncan, Alysa DeFranco, Amanda Wittington and Justin Taylor for assistance with the experiments. We express appreciation for the use of Brigham Young University’s Lytle Ranch Preserve. This research was funded by the Sant Educational Endowment for a Sustainable Environment, the United States Department of Agriculture NIFA award number 2010-04092, and the Bureau of Land Management.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Horn, K.J., Bishop, T.B.B. & St. Clair, S.B. Precipitation timing and soil heterogeneity regulate the growth and seed production of the invasive grass red brome. Biol Invasions 19, 1339–1350 (2017). https://doi.org/10.1007/s10530-016-1348-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-016-1348-2