
Abstract
Spatial heterogeneity of climatic and edaphic gradients can substantially affect the grassland productivity function. However, few studies have tested the importance of species richness and evenness on regulating grassland productivity across spatial-scale climatic and edaphic changes. This study examines the complex mechanisms by which species richness and evenness regulate productivity in alpine meadow and steppe. We used field survey data to explore above-ground productivity formation and sensitivity to spatial-scale climatic and edaphic response of alpine grassland based on species richness and evenness. Results showed that the growing season solar radiation was the main driving factor of above-ground biomass and was strongly negatively correlated with above-ground biomass. Furthermore, compared with alpine steppe, above-ground biomass in alpine meadow was more responsive to climatic variables, but less responsive to soil variables. Unexpectedly, we found that the regulation patterns of species richness and evenness on above-ground biomass were different in both habitats by a structural equation model analysis. Our study demonstrated that species evenness and richness were both important in co-regulating above-ground biomass in alpine meadow, whereas species richness mattered more than species evenness in regulating above-ground biomass in alpine steppe. Our results offer further support for species richness and evenness co-regulating grassland productivity across spatial-scale climatic and edaphic gradients, which helps maintain the benefits of plant diversity and alpine grassland ecosystems.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Global climate is now rapidly changing toward warmer temperatures, which will have alarming consequences for biodiversity and terrestrial ecosystem functioning (Walther et al. 2002; Pugnaire et al. 2019). Above-ground biomass is an essential integrator of terrestrial ecosystem functioning, and affects terrestrial ecosystem carbon cycling feedback to climate change (Cao and Woodward 1998; Liu et al. 2018). Climate can directly affect plant biomass by regulating physiological and environmental determinants of plant growth, e.g., climate-driven shifts in temperature and water availability affect plant production via the photosynthetic rates and respiratory rates of plants (Chu et al. 2016). Indirect effects of climate on plant biomass can be mediated by altering community structure and composition (i.e., altering plant interspecific relationships) (Yang et al. 2011; Wang et al. 2012). Climate can also drive abiotic soil conditions such as soil temperature and moisture to alter soil processes, especially those related to soil fertility, further regulating the effects of soil variables on vegetation growth and plant biomass (Liu et al. 2009; Kardol et al. 2010b). For example, warming reduces soil moisture and increases soil temperature by increasing evapotranspiration, which in turn affects soil nutrient utilization (Niu et al. 2008). Not only does climate influence soil, but also soil also can influence climate. Soils that are wetter or denser hold heat and stabilize the plant productivity from climate change more so than drier or looser soils (Alekseev et al. 2018). Although the feedback between climatic and edaphic is not fully understood, the integrated effects of climatic and edaphic variables may explain directional change of above-ground plant biomass in a future warmer world.
Terrestrial ecosystems may respond to climatic and edaphic variables in different magnitudes and directions. Warming can enhance the vegetation activity by extending the length of the growing season and intensifying the maximum rates of the productivity (Shen et al. 2015), or induce water stress and suppress plant growth by increasing evapotranspiration (Niu et al. 2008). Increased precipitation is typically positively correlated with terrestrial plant biomass due to relief of water and nutrients stress, especially in arid ecosystems (Petrie et al. 2018). Edaphic characteristics with spatiotemporal heterogeneity also are particularly important in influencing the formation and distribution of terrestrial plant community biomass, as soil factors determine nutrient availability and soil water availability (García-Palacios et al. 2012; Ulrich et al. 2014). Nutrient-rich and water-use-efficient soils can promote faster plant development and increase plant biomass, which also increases interspecific competition for available resources and further leads to vegetation death and biomass reduction (Palmquist et al. 2021). While climatic and edaphic variables play different roles in terrestrial ecosystems, uncertainty remains about the sensitivity (mainly referring to strength and direction of responses) of ecosystem function (i.e., plant biomass) to climate and soil responses, then the main drivers of above-ground biomass in terrestrial ecosystem still needs further investigation.
Previous studies have suggested that plant community properties, especially species evenness (equitability of relative abundance among species) and species richness (number of species), also affect key ecosystem processes, such as biomass production (Kardol et al. 2010a; Dorji et al. 2014). Even though there is considerable debate about the shape of the relationships between species richness and plant biomass (e.g., Mittelbach et al. 2001; Sandau et al. 2019), the consensus is that ecosystems with high species richness commonly have higher biomass and productivity (van Ruijven and Berendse 2005; Venail et al. 2015). Species richness has been widely used as the only measurement of species diversity in many studies (Maestre et al. 2012; Chen et al. 2018), and few studies have investigated the effects on plant biomass of the second component of species diversity: species evenness, which has been shown to be equally or more important than species richness (Stirling and Wilsey 2001; Mulder et al. 2004). Species evenness also can indirectly affect terrestrial ecosystem processes through shifting in species richness (Wilsey and Polley 2004). The small-scale and highly controlled diversity experiments are difficult to alter evenness and usually use mixed seeds to maximize species evenness (Smith and Knapp 2003; Mulder et al. 2004), and such diversity-controlled experiments do not reflect the actual distribution patterns of species in natural assembled communities (Kardol et al. 2018). In natural systems, variation in diversity is non-randomly distributed across space and time and driven by various environmental factors (i.e., climate and soil factors), which in turn can affect ecosystem functioning, such as productivity (van der Plas 2019). Regional or large-scale studies of species evenness and richness in natural communities are needed to further advance our understanding of the roles of species evenness and richness in maintaining above-ground plant biomass in terrestrial ecosystems.
Alpine grassland ecosystems are among the most sensitive ecosystems to global climate change, because high-elevation alpine zones are seeing disproportionately rapid warming (Chen et al. 2013; Grimm et al. 2013). The Qinghai-Tibetan Plateau hosts the biggest alpine grassland ecosystem in the world, which has a warming rate of about twice (0.3–0.4℃ per decade) the global average and its temperature will continue to increase in this century (Hansen et al. 2010; Shen et al. 2015). Climate warming can promote earlier shifts in phenological events, enhance vegetation growth rate and shorten growth period in alpine grasslands (Wang et al. 2020). Climate-change-induced shifts in alpine plant growth patterns will further alter alpine biome characteristics and biomass production (Grimm et al. 2013). Soil nutrients and water availability are also key factors regulating the response of alpine grassland productivity to climate change (Fay et al. 2015; Winkler et al. 2016). In high-elevation alpine ecosystems with water shortages, warming will limit soil moisture and release soil nutrients (i.e., C and N) into the atmosphere due to the removal of low-temperature restrictions, which negatively affects plant growth and further threatens alpine grassland productivity (Li et al. 2018; Gao et al. 2021). The unique climate and vegetation growth patterns make the Tibetan Plateau an ideal region for investigating climatic and soil factors influencing the spatial patterns of above-ground biomass in alpine grassland ecosystems.
Here, we report on a unique regional-scale observational data set of above-ground productivity and species diversity of alpine grassland (alpine meadow and alpine steppe) in climatic and soil variables (i.e., temperature, precipitation, soil water content, and soil nutrients) on the Qinghai-Tibetan Plateau. Specifically, this study aimed to: (1) explore the main climatic and soil drivers of primary production of alpine meadow and steppe; and (2) further determine the contributions of species richness and evenness to regulation of grassland productivity across regional-scale climatic and edaphic gradients. Understanding the sensitivity of alpine grassland productivity to the climatic and edaphic gradients, as well as the role of species richness and evenness in alpine grassland ecosystems, is critical for developing adaptive strategies to protect vulnerable grassland ecosystems and for better predicting the responses of different alpine grasslands to on-going climate change.
Materials and methods
Study sites
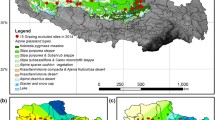
The 116 study sites were located at eastern, southern, and central Qinghai-Tibetan Plateau (26°00′–39°47′N, 73°19′–104°47′E) in Autonomous Region and Qinghai Province, China (Fig. 1a). The Qinghai-Tibetan Plateau has an alpine climate, which is characterized by a short summer and a long winter. The mean altitude is over 4000 m. The annual mean temperature and mean precipitation are 4 ℃ and 400 mm, respectively. The precipitation falls mainly in the rainy season from May to September, considered as the growing season. A significantly decreasing gradient of temperature and precipitation from the southeast to the northwest has been confirmed in the Qinghai-Tibetan Plateau (Tian et al. 2014; Cui et al. 2021). In addition, alpine meadow and alpine steppe are the two dominant grassland types across the Qinghai-Tibetan Plateau. The alpine meadow is characterized by cold and wet climatic conditions, composed by cold-tolerant perennial herbs with dense grass and low grass layers, while alpine steppe is characterized by cold and arid climatic conditions, composed by cold and dry perennial grasses, with sparse vegetation (Zhang et al. 2017; Duan et al. 2021). The vegetation of alpine meadow mainly consists of Kobresia humilis, Carex scabrirostris, Poa pratensis, Potentilla nivea, and Lancea tibetica. The alpine steppe is dominated by Stipa purpurea, Festuca ovina, Leontopodium nanum, Koeleria argentea, and Oxytropis microphylla. The classification of grassland types was based on the Classification Standards of Grassland Types and Classification System of Grassland Types in China. Since the 1990s, China has carried out grazing forbidden and exclusion practices to ensure the sustainable development of grasslands. All sites were in natural grasslands, not grazed or mowed.

a Map of the study region. Locations of sampling sites in alpine meadow (green dots, n = 55) and in alpine steppe (orange triangles, n = 61) on the Qinghai-Tibetan Plateau. The image was generated by ArcGIS software (version 10.5). Data distribution plots of b species richness, c species evenness, and d above-ground biomass in alpine meadow (green) and alpine steppe (orange). Boxplots denote the median (black) and its 95% confidence intervals (whiskers), and the interquartile ranges (gray shadow)
Field measurement
In this survey, we captured a substantial range of vegetation characteristics in 116 sites (55 in the alpine meadows and 61 in the alpine steppes) across a 2131-km east–west and 920-km south-north grassland transects during the peak growing season of 2014 (Fig. 1a). Latitude and longitude of each site were recorded. We determined representative sampling sites based on the basic data of grassland surveys in China and the distribution of typical local grassland communities. We selected a plot (size of 10 m × 10 m) at each sampling site with an interval of 50–100 km, and then randomly placed three quadrats (50 cm × 50 cm) along the diagonal of each plot. The data of each plot were calculated by the average of the three quadrats for subsequent calculations. Within each quadrat, we recorded the height, cover, and density of all the plant species, as well as harvesting the above-ground part, which was then weighed as the species above-ground biomass after being dried at 65 ℃ for 48 h. The number of plant species in each quadrat was recorded as species richness. Species evenness was calculated as:
where S is the total number of species in the community, Pi is the proportion of individuals of species i. We estimated this proportion as Pi = ni / N, where ni is the number of individuals of the ith species, N is the total number of individuals of all the species. ln is the natural logarithm.
Acquisition of meteorological and soil data
For each site, the meteorological data were collected from the European Centre for Medium-Range Weather Forecasts (https://www.ecmwf.int/) with the themes “ERA5 monthly averaged data on single levels from 1979 to present” (Hersbach et al. 2019) and “Essential climate variables for assessment of climate variability from 1979 to present”. The gridded climate data included precipitation, air temperature, solar radiation, soil temperature (0–28 cm depth), and soil water content (0-7 cm depth) during the growing seasons (May to September) from 2004 to 2013. Soil data were obtained from the China soil properties data set (Shangguan et al. 2013). The gridded soil data included soil pH, soil available nitrogen, total nitrogen, available phosphorus, total phosphorus, and soil organic matter at 0–30 cm depth during the growing seasons. All gridded data was extracted using ArcGIS 10.2 software (ESRI, Redlands, CA, USA).
Statistical analyses
We used data distribution plots and t test to compare species richness, species evenness, and above-ground biomass differences between alpine meadow and alpine steppe. The relationships between climatic variables (growing season precipitation, temperature, and solar radiation), soil variables (soil temperature, water content, pH, available nitrogen, total nitrogen, available phosphorus, total phosphorus, and organic matter), with species richness, evenness, and above-ground biomass were evaluated using linear regressions in both alpine meadow and steppe. The t-test analysis was performed using SPSS version 22.0 software (SPSS Inc., Chicago, IL, USA). The data distribution plots and the regression analyses were fitted using the ggplot2 package in R version 3.6.1 software (R Core Team 2019). We further used the variation partitioning analysis (VPA) to estimate the contribution of climate and soil variables to the variation in species richness, evenness, and above-ground biomass in alpine meadow and steppe. This procedure was performed using the “vegan” package in R software. We standardized all data before VPA analysis.
We used structural equation model (SEM) to explain the direct and indirect effects of climate variables, soil variables, species richness, and evenness on above-ground biomass in both alpine meadow and steppe. Prior to the construction of the final SEM, a priori model was built to explore the rationale behind these potential effects (Figure S4). We conducted principal component analysis (PCA) to create a multivariate functional index before SEM construction, and then introduced the first principal component (PC1) of climate and soil variables as new variables to represent the combined group properties into the subsequent SEM analysis (Fig S5 and 4). Considering the sensitivity of the Qinghai-Tibetan Plateau to climate change, and climate variables mainly directly affect the grassland biomass by altering soil water and heat factors. Therefore, we developed new SEMs for the effects of individual climate variables on above-ground biomass indirectly through soil water content and soil temperature (Fig. S6 and 7). The model adequacy was determined by using a Pearson's chi-Square test (χ2), comparative fit index (CFI), and root mean square error of approximation (RMSEA). For SEMs, we used the AMOS 22.0 statistical tool (Amos Development Corporation, Chicago, IL, USA). We also used linear regressions to test the relationships between species richness and evenness with above-ground biomass, and created three-dimensional surfaces using SigmaPlot software to visualize the effects of species richness and evenness on above-ground biomass.
Results
Above-ground productivity and species diversity across climatic and edaphic gradients
Species richness and above-ground biomass were significantly higher in alpine meadow than in alpine steppe (P < 0.001; Fig. 1b, d), but the distribution of species evenness in the two types of grasslands was similar (P = 0.053; Fig. 1c). Additionally, in alpine meadow, the relationships (positive or negative) between species richness and above-ground biomass with climate and soil variables were consistent with those in alpine steppe (Fig. S1, S2, and S3). But the relationships between species evenness and soil nutrient indicators in alpine meadow were different from those in alpine steppe (Fig. S2 and S3). Specifically, species richness and above-ground biomass were significantly negatively related to growing season solar radiation (P < 0.001; Fig. S1c and i) and positively related to growing season precipitation and growing season temperature (Fig. S1a, b, g, and h) in both alpine meadow and alpine steppe. Species evenness in alpine meadow and steppe decreased with increasing growing season precipitation (Fig. S1d), but increased slightly with increasing growing season temperature and solar radiation (Fig. S1e and f).
Soil temperature, soil water content, and soil nutrient indicators were generally positively, while soil pH was negatively, correlated with species richness and above-ground biomass in both alpine meadow and alpine steppe (Fig. S2a-e, k–o and Fig. S3a-c, g-i). Furthermore, species evenness in alpine meadow and steppe increased with increasing soil temperature, but decreased with increasing soil water content (Fig. S2f and g). Additionally, soil available nitrogen, available phosphorus, organic matter, total nitrogen, and total phosphorus were significantly negatively correlated with species evenness of alpine meadow, but did not show any relationship with evenness of alpine steppe (Fig. S2h-j and Fig. S1e-f).
Overall, climatic variables, especially growing season solar radiation and growing season temperature, had stronger effects on species richness and above-ground biomass in alpine meadow (explained 18% and 26% of variation, respectively) than alpine steppe (explained 9% and 21% of variation, respectively; Fig. 2a). But climatic variable had weaker effects on species evenness in alpine meadow (explained 6% of variation) than alpine steppe (explained 9% of variation; Figs. 2a and 3). The effect patterns of soil variables on species richness, above-ground biomass, and species evenness were opposite to the effect patterns of climatic variables (Figs. 2b and 3).

Comparing the explanatory power of a climate and b soil variables for changes in above-ground biomass, species richness, and species evenness in alpine meadow (green) and alpine steppe (orange). Numbers along the dots indicate the percentage change in explanatory, calculated by multiplying the numbers of the bars in Fig. 3 by the R2 of each model

Effects of soil (pink) and climate (purple) variables on (a), (d) species richness, (b), (e) species evenness, and (c), (f) above-ground biomass in both alpine meadow and alpine steppe. For each variable in the model, dots represent the standardized effect size and lines represent standard deviation. R.2 represents the proportion of variance explained by model. The arrows in the bars from strong explanatory factors to weak explanatory factors (see Fig. 2). *P < 0.05; **P < 0.01
Direct and indirect effects of climatic and edaphic variables regulate grassland productivity in alpine meadow and alpine steppe
Our structural equation models (SEM) explained 59% of the variation in above-ground biomass of alpine meadow, and 70% of the variation in above-ground biomass of alpine steppe (Fig. 4a, b). In alpine meadow, climate variables positively affect above-ground biomass indirectly through species richness and soil variables (Fig. 4a, c). But in the alpine steppe, climate variables positively affect above-ground biomass only directly and indirectly through species richness (Fig. 4b, d). Most strikingly, soil variables (i.e., soil water content, temperature, and nutrient indicators) indirectly decreased above-ground biomass in alpine meadow via shifts in species evenness, and had no significant indirect effects on above-ground biomass via shifts in species evenness in alpine steppe (Fig. 4a and b). Furthermore, in alpine meadow, species evenness had a significant negative direct effect on above-ground biomass, while there was no relationship between species evenness and above-ground biomass in alpine steppe (Fig. 4a, b).

Structural equation models developed to relate spatial-scale climate variables, soil variables, and species richness, species evenness, and above-ground biomass in a alpine meadow and b alpine steppe. Black and red arrows indicate positive and negative effects, respectively. Dashed arrows indicate non-significant effects. The thickness of the solid arrows indicates the magnitude of the standardized path coefficients, which are listed beside each arrow. R.2 represents the proportion of variance explained for each dependent variable. The black and red arrows following the climate and soil variables indicate a positive or negative relationship between the variables and the first component from the principal components analysis (PCA), respectively (see Fig. S5). *P < 0.05; **P < 0.01; ***P < 0.001. c, d Standardized total effects (direct plus indirect effects) derived from the structural equation models depicted. Numbers along the bars are standardized total effect values. The acronyms for climate and soil variables are defined in Table S1
The SEM of individual climate variables showed that growing season solar radiation was a key driver of above-ground biomass in both alpine meadow and alpine steppe, with direct effects as well as indirect effects via shifts in species richness (Fig. S7). The SEM further demonstrated that growing season solar radiation and species evenness had overall negative effects (i.e., the sum of direct and indirect effects) on above-ground biomass in both alpine meadow and steppe, while growing season temperature, soil water content, soil properties (i.e., soil pH and nutrient indicators), and species richness had overall positive effects (Fig. S7b and d). Moreover, in alpine meadow, species evenness could indirectly affect above-ground biomass through shifts in species richness, while the indirect effects of species evenness through species richness on above-ground biomass in alpine steppe were not significant (Fig. S7a and c).
Effects of species richness and evenness on grassland productivity formation
To further explore the importance of plant species evenness and richness to above-ground biomass, we analyzed the relationships between species evenness, species richness, and above-ground biomass in both alpine meadow and steppe. Results showed that species richness was positively, while species evenness was negatively, correlated with above-ground biomass in both alpine meadow and alpine steppe (Fig. S8a and b). Additionally, for alpine meadow, species evenness and richness co-maintained above-ground biomass, and lower species evenness and richness had negative effects on above-ground biomass (Fig. S8c). For alpine steppe, the link between species evenness and above-ground biomass shifted from negative to positive as species richness increased, and species richness was related to higher above-ground biomass at low species evenness sites (Fig. S8d).
Discussion
Our results showed that above-ground biomass had positive associations with growing season precipitation, growing season temperature, soil water content, and soil nutrient indicators (i.e., available nitrogen, total nitrogen, available phosphorus, and soil organic matter), but negative associations with growing season solar radiation and soil pH in both alpine meadow and alpine steppe. The growing season solar radiation was a strong driver of above-ground biomass decline in alpine meadow and steppe. Moreover, above-ground biomass in alpine meadow was more responsive to climatic variables than alpine steppe, but less responsive to soil variables. The inconsistent responses of the two grasslands to climatic and edaphic variables may be due to the differences in species composition and species diversity distributions caused by growth environments, i.e., alpine meadow is wetter and warmer than alpine steppe, and meadow can hold more species (Perring 1960; Grime et al. 2008). Furthermore, soil variables indirectly decreased above-ground biomass via shifts in species evenness in alpine meadow, and had no significant indirect effects on above-ground biomass via shifts in species evenness in alpine steppe (Fig. 4). Species evenness also could indirectly affect above-ground biomass in alpine meadow through shifts in species richness, while the indirect effects of species evenness through species richness in alpine steppe were not significant. This implies that the ability of species evenness and richness to regulate above-ground biomass may differ between alpine meadow and steppe. These differences were expressed as species richness and evenness co-determined above-ground biomass in alpine meadow, but species richness rather than evenness primarily determined above-ground biomass in alpine steppe. Elucidating the patterns of species evenness and richness indirectly regulating grassland biomass in different grassland ecosystems under climate and soil changes holds the key to better managing changes in alpine grassland ecosystem functioning under future climate scenarios.
The unique environmental conditions of alpine grassland ecosystems, such as high elevation, low temperatures, and poor soil nutrient availability, strongly affected alpine plant developmental process and productivity in response to climate warming (Shen et al. 2015). Recent research on the effects of climate warming on plant growth in alpine grassland ecosystems has shown inconsistent results. For example, experimental studies on the central Qinghai-Tibetan Plateau indicated that warming stimulated plant growth in alpine meadows, but reduced growth in alpine steppes (Ganjurjav et al. 2016). However, a 9-year ground survey revealed that increasing temperature had a negative effect on plant growth in both alpine meadow and steppe on the Qinghai-Tibetan Plateau (Ganjurjav et al. 2018). In our current study, we found a consistent positive relationship between growing season temperature and soil temperature with species richness and above-ground biomass in both alpine meadow and steppe. Considering that rising temperatures was proved to promote earlier vegetation phenology (such as green-up dates) and faster growth (Wang et al. 2020), and warming is generally expected to alleviate low-temperature constraints on plant growth in alpine regions (Park et al. 2019), our results suggest that increasing temperature might be conducive to plant growth of alpine meadow and steppe. Precipitation is also a critical climatic determinant of plant growth and community biomass over much of the globe (O'Connor et al. 2001; Knapp et al. 2017). Our results were consistent with previous studies (Yang et al. 2009; Sun et al. 2013) showing that growing season precipitation and soil water content were positively related to plant species richness and above-ground biomass in both alpine meadow and steppe. This finding further supported that water availability is an important control of alpine plant production.
It should be noted that growing season solar radiation had a negative effect (standardized total effects < 0) on species richness and above-ground biomass in alpine meadow and steppe in this study, which predicted that the growing season solar radiation was the main driver of above-ground biomass declines. This was probably because that light, as a direct source of photosynthesis, was an essential prerequisite for plant growth and development (Sun et al. 2019). Appropriate illumination intensity promotes photosynthesis rate and improves the yield and quality of vegetation (Chang et al. 2008). However, the high radiation intensity in the study area on the Qinghai-Tibetan Plateau makes it easy for plant to reach the light saturation point (Liu et al. 2012), thus reduces the activity of photosynthesis enzymes, damages plant growth hormones, leads to photoinhibition of photosynthesis, and ultimately reduce above-ground biomass. Another possible reason for the decline in above-ground biomass is that increased solar radiation will accelerate the rise of soil temperature and evapotranspiration, reduce soil moisture, and inhibit plant growth (Liu et al. 2020).
Importantly, we found that compared with above-ground biomass in alpine steppe, above-ground biomass in alpine meadow showed stronger correlations with spatial-scale climatic variables, soil temperature, soil water content, and soil pH, while weaker correlations with soil nutrient indicators. This might be due to meteorological, topographic, and soil factors leading to differences in the spatial patterns of plant composition and productivity in alpine meadow and steppe (Sun et al. 2013). In detail, the habitat for alpine meadow was wetter and warmer than the alpine steppe, and the species richness and above-ground biomass of alpine meadow were also higher (Sun et al. 2020). Diversity has been shown to be the basis of productivity, resistance to invasion, stability, and other desirable community properties (Harrison et al. 2015). Substantial evidences also suggests that high plant richness could stabilize grassland ecosystem functioning (e.g., productivity) in a fluctuating environment (Yachi and Loreau 1999; Wang et al. 2019). It might be assumed that alpine meadow with high diversity responds to changing climatic variables faster than the low-diversity alpine steppe. Due to the long-term dry and cold environment of alpine steppe (Peng et al. 2020), this type of alpine grassland might have developed adaptability and resistance to low soil moisture and temperature (Thompson and Fronhofer 2019). Thus, alpine steppe was less sensitive to changes in soil water content, soil temperature, and soil pH than alpine meadow in the current survey. Additionally, the reason that alpine steppe was more sensitive to changes in soil nutrients, while this was not the case for alpine meadow, was probably the unique alpine sod layer containing high content of soil material and sufficient nutrients of the latter one (Su et al. 2018). Collectively, our study highlights the different impacts of spatial-scale climatic and edaphic variables on different grassland types, and suggests that future research should adopt appropriate conservation measures for different grassland types.
Results of the structural equation models analysis revealed that climate variable, especially growing season solar radiation, played a vital role in alpine grasslands biomass. Climate variables exhibited a positive effect on above-ground biomass through its direct effects and the indirect effects mediated by soil variables (i.e., soil water content, temperature, and nutrient indicators) and species diversity in both alpine meadow and steppe. For example, high precipitation and temperature levels may alleviate low temperature limitations of alpine grasslands on the Qinghai-Tibetan Plateau and increase soil moisture (Chen et al. 2018), and therefore, promote species diversity and plant biomass. Remarkably, the indirect effects of soil variables (i.e., soil water content, temperature, and soil nutrients) on above-ground biomass were significantly negative in alpine meadow while no significant in alpine steppe, mediated by shifts in species evenness (Fig. 4). This result suggests that species evenness is more important in alpine meadow than in alpine steppe for indirectly regulating the effects of climate and soil variables on above-ground biomass. According to previous research, variation in community evenness was largely driven by spatial changes in dominant species (Cerabolini et al. 2010). Increasing soil water and nutrient resources (i.e., N, P, and organic matter) probably promoted vegetation growth and increased species evenness in alpine meadow (Orwin et al. 2014). Subsequently, the competition of species for resources gradually became stronger, and the slow-growing species became the dominant species, leading to a decrease in plant biomass (Loreau et al. 2003; Mulder et al. 2004).
Additionally, we found a significant negative relationship between species evenness and above-ground biomass in alpine meadow, but did not detect any significant relationships between species evenness and above-ground biomass in alpine steppe by the SEMs (Fig. 4 and S7). This further supports the above results that species evenness is more crucial in regulating above-ground biomass in alpine meadow than in alpine steppe. Our analysis also showed that species richness and evenness co-regulated above-ground biomass in alpine meadow, but species richness played a dominant role rather than species evenness in alpine steppe (Fig. 4, S7, and S8). At the regional scale, the role of species diversity might be influenced by plant community attributes. For example, as a measure of species distribution and species abundance in plant communities, species evenness was been shown to be equal or more important than species richness (Maestre et al. 2012). Moreover, we found that species evenness could affect above-ground biomass indirectly via affecting species richness in alpine meadow, but not in alpine steppe (Fig. S7a and c). As a result, species evenness could directly and indirectly affect biomass via shifts in species richness, thus realizing the co-effect patterns of species evenness and richness on plant biomass found in the current alpine meadow study. In alpine steppe, as species richness increased, the relationships between species evenness and above-ground biomass shifted from negative to positive. Since above-ground biomass was positively correlated with species richness and negatively correlated with species evenness, these two effects might offset with each other, resulting in the ultimate species richness-determined above-ground biomass of alpine steppe. However, the potential mechanism for the influence of species evenness and richness on different grassland ecosystems was poorly understood. Future studies are necessary to consider the combined effects of species richness and evenness and which index can driver plant biomass more effectively for different grassland ecosystems.
Conclusion
Our results showed that growing season solar radiation was the primary predictor of above-ground biomass decline in both alpine meadow and steppe under spatial-scale climatic heterogeneity, and the negative effects of solar radiation on above-ground biomass were weaker than the total positive effects of growing season precipitation and temperature. Furthermore, the response of alpine meadow productivity to spatial-scale climatic gradient was stronger than that of alpine steppe, while the response of alpine meadow productivity to spatial-scale edaphic variables was weaker than that of alpine steppe. We, therefore, suggest that the negative effect of spatial-scale climatic gradient, especially of solar radiation should be particularly considered when conserving alpine meadow productivity functioning. Results of the structural equation models further showed that species evenness and richness co-regulated alpine meadow productivity at the regional scale, with a direct negative effect of the former and a direct positive effect of the latter. However, species richness was the main driver rather than specie evenness in alpine steppe productivity formation and sensitivity. Remarkably, species evenness also participates to contribute to buffer the effects of spatial-scale climatic gradient on alpine grassland productivity as species richness do and, therefore, warrant further study. Collectively, our study highlights the importance of conserving species diversity and evenness to maintain the balance and stability of alpine grassland productivity under future climate change.
Data availability
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Code availability
Not applicable.
References
Alekseev A, Alekseeva T, Kalinin P, Hajnos M (2018) Soils response to the land use and soil climatic gradients at ecosystem scale: Mineralogical and geochemical data. Soil till Res 180:38–47. https://doi.org/10.1016/j.still.2018.02.008
Cao MK, Woodward FI (1998) Dynamic responses of terrestrial ecosystem carbon cycling to global climate change. Nature 393:249–252. https://doi.org/10.1038/30460
Cerabolini B, Pierce S, Luzzaro A, Ossola A (2010) Species evenness affects ecosystem processes in situ via diversity in the adaptive strategies of dominant species. Plant Ecol 207:333–345. https://doi.org/10.1007/s11258-009-9677-1
Chang XM, Alderson PG, Wright CJ (2008) Solar irradiance level alters the growth of basil (Ocimum basilicum L.) and its content of volatile oils. Environ Exp Bot 63:216–223. https://doi.org/10.1016/j.envexpbot.2007.10.017
Chen H, Zhu QA, Peng CH, Wu N, Wang YF, Fang XQ et al (2013) The impacts of climate change and human activities on biogeochemical cycles on the Qinghai-Tibetan Plateau. Global Change Biol 19:2940–2955. https://doi.org/10.1111/gcb.12277
Chen SP, Wang WT, Xu WT, Wang Y, Wan HW, Chen DM et al (2018) Plant diversity enhances productivity and soil carbon storage. Proc Natl Acad Sci USA 115:4027–4032. https://doi.org/10.1073/pnas.1700298114
Chu CJ, Bartlett M, Wang YS, He FL, Weiner J, Chave J, Sack L (2016) Does climate directly influence NPP globally? Global Change Biol 22:12–24. https://doi.org/10.1111/gcb.13079
Cui T, Li C, Tian FQ (2021) Evaluation of temperature and precipitation simulations in cmip6 models over the Tibetan Plateau. Earth and Space Sci. https://doi.org/10.1029/2020EA001620
Dorji T, Moe SR, Klein JA, Totland Ø (2014) Plant species richness, evenness, and composition along environmental gradients in an alpine meadow grazing ecosystem in central Tibet, China. Arct Antarct Alp Res 46:308–326. https://doi.org/10.1657/1938-4246-46.2.308
Duan HC, Xue X, Wang T, Kang WP, Liao J, Liu SL (2021) Spatial and temporal differences in alpine meadow, alpine steppe and all vegetation of the Qinghai-Tibetan Plateau and their responses to climate change. Remote Sens 13:669. https://doi.org/10.3390/rs13040669
Fay PA, Prober SM, Harpole WS, Knops JMH, Bakker JD, Borer ET et al (2015) Grassland productivity limited by multiple nutrients. Nat Plants 1:15080. https://doi.org/10.1038/NPLANTS.2015.80
Ganjurjav H, Gao QZ, Gornish ES, Schwartz MW, Liang Y, Cao XJ et al (2016) Differential response of alpine steppe and alpine meadow to climate warming in the central Qinghai-Tibetan Plateau. Agr Forest Meteorol 223:233–240. https://doi.org/10.1016/j.agrformet.2016.03.017
Ganjurjav H, Gornish ES, Hu GZ, Wan YF, Li Y, Danjiu LB, Gao QZ (2018) Temperature leads to annual changes of plant community composition in alpine grasslands on the Qinghai-Tibetan Plateau. Environ Monit Assess 190:585. https://doi.org/10.1007/s10661-018-6964-0
Gao WL, Sun WM, Xu XL (2021) Permafrost response to temperature rise in carbon and nutrient cycling: effects from habitat-specific conditions and factors of warming. Ecol Evol 11:16021–16033. https://doi.org/10.1002/ece3.8271
García-Palacios P, Maestre FT, Bardgett RD, de Kroon H (2012) Plant responses to soil heterogeneity and global environmental change. J Ecol 100:1303–1314. https://doi.org/10.1111/j.1365-2745.2012.02014.x
Grime JP, Fridley JD, Askew AP, Thompson K, Hodgson JG, Bennett CR (2008) Long-term resistance to simulated climate change in an infertile grassland. Proc Natl Acad Sci USA 105:10028–10032. https://doi.org/10.1073/pnas.0711567105
Grimm NB, Chapin FS, Bierwagen B, Gonzalez P, Groffman PM, Luo YQ et al (2013) The impacts of climate change on ecosystem structure and function. Front Ecol Environ 11:474–482. https://doi.org/10.1890/120282
Hansen J, Ruedy R, Sato M, Lo K (2010) Global surface temperature change. Rev Geophys 48:1–52. https://doi.org/10.1029/2010RG000345
Harrison SP, Gornish ES, Copeland S (2015) Climate-driven diversity loss in a grassland community. Proc Natl Acad Sci USA 112:8672–8677. https://doi.org/10.1073/pnas.1502074112
Hersbach H, Bell B, Berrisford P, Biavati G, Horányi A, Muñoz Sabater J et al (2019) ERA5 monthly averaged data on single levels from 1979 to present. Copernicus Climate Change Service (C3S) Climate Data Store (CDS). 10:252-266
Kardol P, Campany CE, Souza L, Norby RJ, Weltzin JF, Classen AT (2010a) Climate change effects on plant biomass alter dominance patterns and community evenness in an experimental old-field ecosystem. Global Change Biol 16:2676–2687. https://doi.org/10.1111/j.1365-2486.2010.02162.x
Kardol P, Cregger MA, Campany CE, Classen AT (2010b) Soil ecosystem functioning under climate change: plant species and community effects. Ecology 91:767–781. https://doi.org/10.1890/09-0135.1
Kardol P, Fanin N, Wardle DA (2018) Long-term effects of species loss on community properties across contrasting ecosystems. Nature 557:710–713. https://doi.org/10.1038/s41586-018-0138-7
Knapp AK, Ciais P, Smith MD (2017) Reconciling inconsistencies in precipitation-productivity relationships: implications for climate change. New Phytol 214:41–47. https://doi.org/10.1111/nph.14381
Li CY, Peng F, Xue X, You QG, Lai CM, Zhang WJ, Cheng YX (2018) Productivity and quality of alpine grassland vary with soil water availability under experimental warming. Front Plant Sci 9:1790. https://doi.org/10.3389/fpls.2018.01790
Liu HY, Mi ZR, Lin L, Wang YH, Zhang ZH, Zhang FW et al (2018) Shifting plant species composition in response to climate change stabilizes grassland primary production. Proc Natl Acad Sci USA 115:4051–4056. https://doi.org/10.1073/pnas.1700299114
Liu JD, Liu JM, Linderholm HW, Chen DL, Yu Q, Wu DG, Haginoya S (2012) Observation and calculation of the solar radiation on the Tibetan Plateau. Energ Convers Manage 57:23–32. https://doi.org/10.1016/j.enconman.2011.12.007
Liu LB, Gudmundsson L, Hauser M, Qin DH, Li SC, Seneviratne SI (2020) Soil moisture dominates dryness stress on ecosystem production globally. Nat Commun 11:4892. https://doi.org/10.1038/s41467-020-18631-1
Liu WX, Zhang Z, Wan SQ (2009) Predominant role of water in regulating soil and microbial respiration and their responses to climate change in a semiarid grassland. Global Change Biol 15:184–195. https://doi.org/10.1111/j.1365-2486.2008.01728.x
Loreau M, Mouquet N, Gonzalez A (2003) Biodiversity as spatial insurance in heterogeneous landscapes. Proc Natl Acad Sci USA 100:12765–12770. https://doi.org/10.1073/pnas.2235465100
Maestre FT, Castillo-Monroy AP, Bowker MA, Ochoa-Hueso R (2012) Species richness effects on ecosystem multifunctionality depend on evenness, composition and spatial pattern. J Ecol 100:317–330. https://doi.org/10.1111/j.1365-2745.2011.01918.x
Mittelbach GG, Steiner CF, Scheiner SM, Gross KL, Reynolds HL, Waide RB, Willig MR, Dodson SI, Gough L (2001) What is the observed relationship between species richness and productivity? Ecology 82:2381–2396. https://doi.org/10.1890/0012-9658(2001)082[2381:WITORB]2.0.CO;2
Mulder CPH, Bazeley-White E, Dimitrakopoulos PG, Hector A, Scherer-Lorenzen M, Schmid B (2004) Species evenness and productivity in experimental plant communities. Oikos 107:50–63. https://doi.org/10.1111/j.0030-1299.2004.13110.x
Niu SL, Wu MY, Han Y, Xia JY, Li LH, Wan SQ (2008) Water-mediated responses of ecosystem carbon fluxes to climatic change in a temperate steppe. New Phytol 177:209–219. https://doi.org/10.1111/j.1469-8137.2007.02237.x
O’Connor TG, Haines LM, Snyman HA (2001) Influence of precipitation and species composition on phytomass of a semi-arid African grassland. J Ecol 89:850–860. https://doi.org/10.1046/j.0022-0477.2001.00605.x
Orwin KH, Ostle N, Wilby A, Bardgett RD (2014) Effects of species evenness and dominant species identity on multiple ecosystem functions in model grassland communities. Oecologia 174:979–992. https://doi.org/10.1007/s00442-013-2814-5
Palmquist KA, Schlaepfer DR, Renne RR, Torbit SC, Doherty KE, Remington TE, Watson G, Bradford JB, Lauenroth WK (2021) Divergent climate change effects on widespread dryland plant communities driven by climatic and ecohydrological gradients. Global Change Biol 27:5169–5185. https://doi.org/10.1111/gcb.15776
Park T, Chen C, Macias-Fauria M, Tømmervik H, Choi S, Winkler A et al (2019) Changes in timing of seasonal peak photosynthetic activity in northern ecosystems. Global Change Biol 25:2382–2395. https://doi.org/10.1111/gcb.14638
Peng F, Xue X, Li CY, Lai CM, Sun J, Tsubo M et al (2020) Plant community of alpine steppe shows stronger association with soil properties than alpine meadow alongside degradation. Sci Total Environ 733:139048. https://doi.org/10.1016/j.scitotenv.2020.139048
Perring FH (1960) Climatic gradients of chalk grassland. J Ecol 48:415–442. https://doi.org/10.2307/2257526
Petrie MD, Peters DPC, Yao J, Blair JM, Burruss ND, Collins SL et al (2018) Regional grassland productivity responses to precipitation during multiyear above- and below-average rainfall periods. Global Change Biol 24:1935–1951. https://doi.org/10.1111/gcb.14024
Pugnaire FI, Morillo JA, Peñuelas J, Reich PB, Bardgett RD, Gaxiola A et al (2019) Climate change effects on plant-soil feedbacks and consequences for biodiversity and functioning of terrestrial ecosystems. Sci Adv. https://doi.org/10.1126/sciadv.aaz1834
R Core Team (2019) R: A Language and Environment for Statistical Computing. R for Statistical Computing, Vienna
Sandau N, Naisbit RE, Fabian Y, Bruggisser OT, Kehrli P, Aebi A, Rohr RP, Bersier LF (2019) Understanding negative biodiversity-ecosystem functioning relationship in semi-natural wildflower strips. Oecologia 189:185–197. https://doi.org/10.1007/s00442-018-4305-1
Shangguan W, Dai YJ, Liu BY, Zhu AX, Duan QY, Wu LZ et al (2013) A China dataset of soil properties for land surface modeling. J Adv Model Earth Syst 5:212–224. https://doi.org/10.1002/jame.20026
Shen MG, Piao SL, Jeong SJ, Zhou LM, Zeng ZZ, Ciais P et al (2015) Evaporative cooling over the Tibetan Plateau induced by vegetation growth. Proc Natl Acad Sci USA 112:9299–9304. https://doi.org/10.1073/pnas.1504418112
Smith MD, Knapp AK (2003) Dominant species maintain ecosystem functioning with non-random species loss. Ecol Lett 6:509–517. https://doi.org/10.1046/j.1461-0248.2003.00454.x
Stirling G, Wilsey B (2001) Empirical relationships between species richness, evenness, and proportional diversity. Am Nat 158:286–299. https://doi.org/10.1086/321317
Su PX, Zhou ZJ, Shi R, Xie TT (2018) Variation in basic properties and carbon sequestration capacity of an alpine sod layer along moisture and elevation gradients. Acta Ecol Sini 38:1040–1052. https://doi.org/10.5846/stxb201611222384
Sun J, Cheng GW, Li WP (2013) Meta-analysis of relationships between environmental factors and aboveground biomass in the alpine grassland on the Tibetan Plateau. Biogeosciences 10:1707–1715. https://doi.org/10.5194/bg-10-1707-2013
Sun J, Liu BY, You Y, Li WP, Liu M, Shang H, He JS (2019) Solar radiation regulates the leaf nitrogen and phosphorus stoichiometry across alpine meadows of the Tibetan Plateau. Agr Forest Meteorol 271:92–101. https://doi.org/10.1016/j.agrformet.2019.02.041
Sun J, Zhou TC, Liu M, Chen YC, Liu GH, Xu M et al (2020) Water and heat availability are drivers of the aboveground plant carbon accumulation rate in alpine grasslands on the Tibetan Plateau. Global Ecol Biogeogr 29:50–64. https://doi.org/10.1111/geb.13006
Thompson PL, Fronhofer EA (2019) The conflict between adaptation and dispersal for maintaining biodiversity in changing environments. Proc Natl Acad Sci USA 116:21061–21067. https://doi.org/10.1073/pnas.1911796116
Tian L, Zhang YJ, Zhu JT (2014) Decreased surface albedo driven by denser vegetation on the Tibetan Plateau. Environ Res Lett 10:104001. https://doi.org/10.1088/1748-9326/9/10/104001
Ulrich W, Soliveres S, Maestre FT, Gotelli NJ, Quero JL, Delgado-Baquerizo M et al (2014) Climate and soil attributes determine plant species turnover in global drylands. J Biogeogr 41:2307–2319. https://doi.org/10.1111/jbi.12377
van der Plas F (2019) Biodiversity and ecosystem functioning in naturally assembled communities. Biol Rev 94:1220–1245. https://doi.org/10.1111/brv.12499
van Ruijven J, Berendse F (2005) Diversity-productivity relationships: initial effects, long-term patterns, and underlying mechanisms. Proc Natl Acad Sci USA 102:695–700. https://doi.org/10.1073/pnas.0407524102
Venail P, Gross K, Oakley TH, Narwani A, Allan E, Flombaum P et al (2015) Species richness, but not phylogenetic diversity, influences community biomass production and temporal stability in a re-examination of 16 grassland biodiversity studies. Funct Ecol 29:615–626. https://doi.org/10.1111/1365-2435.12432
Walther GR, Post E, Convey P, Menzel A, Parmesan C, Beebee TJC, Fromentin JM, Hoegh-Guldberg O, Bairlein F (2002) Ecological responses to recent climate change. Nature 416:389–395. https://doi.org/10.1038/416389a
Wang H, Liu HY, Cao GM, Ma ZY, Li YK, Zhang FW et al (2020) Alpine grassland plants grow earlier and faster but biomass remains unchanged over 35 years of climate change. Ecol Lett 23:701–710. https://doi.org/10.1111/ele.13474
Wang SP, Duan JC, Xu GP, Wang YF, Zhang ZH, Rui YC et al (2012) Effects of warming and grazing on soil N availability, species composition, and ANPP in an alpine meadow. Ecology 93:2365–2376. https://doi.org/10.2307/41739308
Wang YF, Cadotte MW, Chen YX, Fraser LH, Zhang YH, Huang FM, Luo S, Shi NY, Loreau M (2019) Global evidence of positive biodiversity effects on spatial ecosystem stability in natural grasslands. Nat Commun 10:3207. https://doi.org/10.1038/s41467-019-11191-z
Wilsey BJ, Polley HW (2004) Realistically low species evenness does not alter grassland species-richness-productivity relationships. Ecology 85:2693–2700. https://doi.org/10.1890/04-0245
Winkler DE, Chapin KJ, Kueppers LM (2016) Soil moisture mediates alpine life form and community productivity responses to warming. Ecology 97:1555–1565. https://doi.org/10.1890/15-1197.1
Yachi S, Loreau M (1999) Biodiversity and ecosystem productivity in a fluctuating environment: the insurance hypothesis. Proc Natl Acad Sci USA 96:1463–1468. https://doi.org/10.1073/pnas.96.4.1463
Yang HJ, Wu MY, Liu WX, Zhang Z, Zhang NL, Wan SQ (2011) Community structure and composition in response to climate change in a temperate steppe. Global Change Biol 17:452–465. https://doi.org/10.1111/j.1365-2486.2010.02253.x
Yang YH, Fang JY, Pan YD, Ji CJ (2009) Aboveground biomass in Tibetan grasslands. J Arid Environ 73:91–95. https://doi.org/10.1016/j.jaridenv.2008.09.027
Zhang T, Wang GX, Yang Y, Mao TX, Chen XP (2017) Grassland types and season-dependent response of ecosystem respiration to experimental warming in a permafrost region in the Tibetan Plateau. Agr Forest Meteorol 247:271–279. https://doi.org/10.1016/j.agrformet.2017.08.010
Acknowledgements
We thank the editors and anonymous reviewers for their constructive comments and suggestions, thank work team members for their contributions to the assistance with field work and data collection. We also thank Paul B. Reed, Jonathan A. Walter and Shuqiong Wang for their suggestions and improvement on this manuscript.
Funding
This study was funded by projects of the National Natural Science Foundation of China (NSFC 32230068, NSFC 41722107), the Strategic Priority Research Program of the Chinese Academy of Sciences (XDB40000000), the Natural Science Foundation of Qinghai Province (2020-ZJ-726), and the Open Project of the Qinghai Provincial Key Laboratory of Restoration Ecology in Cold Area (2020-KF-07, 2020-KF-05).
Author information
Authors and Affiliations
Contributions
G.L.W. and J.S. conceived the idea and designed the study; Z.C. and J.S. collected the data and analyzed the data; Z.C., J.Z., J.S., and G.L.W. wrote the manuscript. All authors contributed critically to the draft and gave final approval for publication.
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Ethics approval
Not applicable.
Additional information
Communicated by Brian J. Wilsey.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Cheng, Z., Zhao, J., Sun, J. et al. Determining the role of richness and evenness in alpine grassland productivity across climatic and edaphic gradients. Oecologia 200, 491–502 (2022). https://doi.org/10.1007/s00442-022-05279-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-022-05279-5