
Abstract
Plant diversity can significantly affect the grassland productivity and its stability. However, it remains unclear how plant diversity affects the spatial stability of natural grassland productivity, especially in alpine regions that are sensitive to climate change. We analyzed the interaction between plant (species richness and productivity, etc.) and climatic factors (precipitation, temperature, and moisture index, etc.) of alpine natural grassland on the Qinghai-Tibetan Plateau. In addition, we tested the relationship between plant diversity and spatial stability of grassland productivity. Results showed that an increase in plant diversity significantly enhanced community productivity and its standard deviation, while reducing the coefficient of variation in productivity. The influence of plant diversity on productivity and the reciprocal of productivity variability coefficient was not affected by vegetation types. The absolute values of the regression slopes between climate factors and productivity in alpine meadow communities with higher plant diversity were smaller than those in alpine meadow communities with lower plant diversity. In other words, alpine meadow communities with higher plant diversity exhibited a weaker response to climatic factors in terms of productivity, whereas those with lower plant diversity showed a stronger response. Our results indicate that high plant diversity buffers the impact of ambient pressure (e.g., precipitation, temperature) on alpine meadow productivity, and significantly enhanced the spatial stability of grassland productivity. This finding provides a theoretical basis for maintaining the stability of grassland ecosystems and scientifically managing alpine grasslands under the continuous climate change.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The impact of plant diversity on ecosystem productivity is fundamental to understanding the global extinction crisis and its effects on the functioning of natural ecosystems (Liang et al. 2016). Over the past 3 decades, ecologists have extensively explored the consequences of plant diversity on ecosystem productivity and stability (Loreau & Hector 2001; Loreau & de Mazancourt 2013; Leps et al. 2018; Hautier et al. 2020). Although a positive correlation between productivity and plant diversity is the most common phenomenon in species diversity experiments (Balvanera et al. 2006; Cardinale et al. 2007; Isbell et al. 2015; Grace et al. 2016; Duffy et al. 2017; Wang et al. 2019a, b), this relationship is affected by the external environmental factors (Steudel et al. 2012; Liang et al. 2016). In the context of global trends in biodiversity loss in ecosystems (Allan et al. 2015; Vellend et al. 2017; Linders et al. 2019), the relationship between the stability of productivity and plant diversity in grassland ecosystems remains a hot topic in ecological research (Cardinale et al. 2007; Grace et al. 2016; Duffy et al. 2017; Wang et al. 2019a, b).
Interactions among species have been widely used to explain the positive effects of species diversity on grassland productivity and ecosystem function because beneficial interactions among species can lead to complementarity effects among species (Loreau & Hector et al., 2001; Cardinale et al. 2007). For example, Loreau & Hector (2001) has demonstrated that mixed planting has higher productivity than single planting due to niche complementarity in mixed planting system. Compensatory dynamics are usually caused by differences in resource acquisition and utilization among species with different traits and advantages (Barry et al. 2019), which leads to the possibility that the compensatory dynamics may increase with the increase of plant diversity (Loreau & de Mazancourt 2013; Pires et al. 2018). Therefore, many ecologists have realized that plant diversity plays a crucial role in maintaining ecosystem stability facing with the external disturbances and ambient pressure (Balvanera et al. 2006; Cardinale et al. 2012; Steudel et al. 2012).
The reduction of plant diversity resulting in biotic homogenization may affect community stability by altering the spatial similarity of community composition (Olden et al. 2004; Tilman et al. 2014). Many experimental and theoretical studies have shown that a decrease in local plant diversity can impair the production function and stability of ecosystem (Solar et al. 2015; Wang et al. 2021), but these studies are based on small-scale anthropogenic disturbance and control of ecosystems (Worm et al. 2006; Weigelt et al. 2008; Cardinale et al. 2013; Wang et al. 2019a). The ecological structure and biophysical processes of natural grasslands are more complex than those of controlled experimental grasslands. However, it remains unclear whether these small-scale results can be extrapolated to the effect of plant diversity changes on spatial stability of productivity in natural grasslands (Gonzalez et al. 2020; Wang et al. 2021). Recent reports have begun to address this issue at the spatial scales (Liang et al. 2016, 2021; Wang & Loreau, 2016; Wilcox et al. 2017; Zhang et al. 2018; Patrick et al. 2021; Wang et al. 2021; Liang et al. 2022) and have shown that a reduction in beta diversity at the spatial scale can impair ecosystem functioning (Hautier et al. 2018; Liang et al. 2022). However, it remains unclear how plant diversity affects the spatial stability of productivity in natural grassland, especially in climatically sensitive alpine regions.
The Tibetan Plateau provides important services for human survival and development, but the region is highly climate sensitive and more susceptible to environmental changes caused by climate change. It has been reported that the interannual variations in precipitation and the rate of climate warming on the Qinghai-Tibetan Plateau (Increase by 0.4 °C per decade) are higher than the global average (Chen et al., 2013; Shen et al., 2015). Therefore, studying the effects of plant diversity on grassland productivity and its stability at the spatial scale in alpine regions are of great significance for understanding the effect of plant diversity on grassland ecosystem functioning under the background of climate change. Previous studies focused on the effects of precipitation and temperature changes on various variables in alpine grassland ecosystems, such as productivity, grassland quality, litter, etc. (Klein et al. 2007; Haynes et al. 2014). Nevertheless, few researches have focused on the effects of plant diversity on grassland productivity stability at the spatial scales and along climate gradients. Hence, this study tested two hypotheses: (H1) plant diversity enhances the spatial stability of grassland productivity; (H2) higher plant diversity buffers the impact of the spatial-scale climate changes on grassland productivity. The results of this study can provide management and theoretical basis for the health and sustainable development of grassland ecosystems in the future rapid climate change.
Materials and methods
Study site
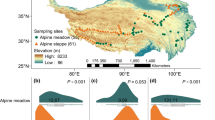
The 117 natural grasslands (sampling sites) surveyed and sampled in this study are located in the Qinghai-Tibetan Plateau (26°00′− 39°47′N, 73°19′− 104°47′E) (Fig. 1), which has the largest natural alpine grasslands in China, covering an area of 1.28 × 106 km2. The main types of alpine grasslands in this region are alpine meadow (AM) and alpine steppe (AS), with areas of 5.82 × 105 km2 and 3.74 × 105 km2, respectively. AS is mainly composed of perennial grasses and Carex ssp. (Cyperaceae), whereas AM consists mainly Kobresia and Carex ssp. The Qinghai-Tibetan Plateau is a typical alpine climate, characterized by long winter and short summer. From the southeast to the northwest, the annual average temperature decreases significantly from 20℃ to – 6 °C. Meanwhile, mean annual precipitation in most areas of the Qinghai-Tibetan Plateau also decreases from the southeast to the northwest (Qin et al. 2018). The mean annual precipitation (MAP) is about 400 mm, mainly in the growing period of plants (from May to September).

Location of sample sites in Qinghai-Tibetan Plateau for alpine grasslands (n = 117). Blue and red points represent sample sites of alpine steppe and alpine meadow, respectively. The diameters of the points are directly proportional to species richness, meaning that larger diameter points correspond to higher species richness. The gray areas are the non-investigated land, including swamp and non-grassland. The map is displayed using ArcGIS 10.2
Investigation of plant community
To exclude the influences of livestock grazing on the experimental results, we selected samples with the same grazing intensity (moderate grazing) and management approach (cool-season grazing). In this study, the vegetation characteristics (species richness, aboveground biomass of each species) of 117 sites from alpine meadow (AM) and alpine steppe (AS) were investigated during the peak-growing season of 2014. Among these sites, 56 were from AM, while 61 were from AS. We randomly assigned three quadrats (1 m × 1 m) to each site (100 m2), each of which was at least 5 m apart. Then we collected the aboveground part of each species and recorded the total number of species to calculate the aboveground net primary productivity (ANPP) and plant diversity of the quadrat. Species richness (SR) is universally used as an indicator of plant diversity in alpine grassland ecosystems (Wang et al. 2019b). The total dry mass of each living species within the quadrat is considered as an estimate of ANPP (Chen et al. 2018) as the sampling was conducted during the peak-growing season (late July to August).
Collection of meteorological data
The meteorological data for the 117 sites in this study were acquired from the Resources and Environmental Sciences and Data Center (RESDC) of Chinese Academy of Sciences (CAS) (http://www.resdc.cn/). The kriging interpolation method was used to carry out spatial interpolation on the data of 109 meteorological observation stations within the study areas, and then mean annual precipitation (MAP), mean annual temperature (MAT), annual accumulated temperature (ACT), and moisture index (MI) were extracted.
Statistical analyses
Spatial stability is represented by the reciprocal of spatial variability. The variability of ecosystem productivity is commonly quantified as coefficient of variation (CV), which is calculated as the ratio of standard deviation (SD) to the mean of productivity. This study used the reciprocal of CV as an indicator to assess the spatial stability of community productivity, which implies that a higher reciprocal of CV corresponds to greater stability of community productivity. The reciprocal of CV for productivity was calculated using the productivity data from three quadrats at the same site. Correlation analysis and linear regression analysis were used to analyze the relationships between ANPP, SD, the reciprocal of CV, and species richness (SR). Covariance analysis was employed to test the effect of grassland type, SR, and the interaction between grassland type and SR on ANPP, SD, and the reciprocal of CV.
Multiple regression analysis was employed to analyze the relationship between climatic factors and ANPP along species richness gradient. To eliminate the influence of vegetation types on the results, we divided the quadrats into alpine meadow and alpine steppe. In the alpine meadow, there is only 1 quadrat with 32 species, while the remaining 164 quadrats have species numbers that are less than or equal to 28. The species richness was divided into three equal intervals, creating three gradients of richness: low species richness (1–9 species), medium species richness (10–18 species), and high species richness (19–28 species). In the alpine steppe, only 3 quadrats have more than 20 species (specifically, 21, 26, and 27), while the species numbers in the remaining 164 quadrats are all less than or equal to 17. The species richness was divided into two equal intervals, creating two gradients of richness: low species richness (1–8 species) and medium species richness (9–17 species). Based on the estimated values of regression slope, their standard deviations, and sample sizes, the T-statistic (T) and degrees of freedom (df) were calculated using the following equation (Moore et al. 1993). Then a t test was conducted using the T-statistic and df to assess whether there are significant differences in the absolute values of regression slope between different levels of species richness.
where |Slope1| and |Slope2| are the absolute values of regression slope 1 and regression slope 2, respectively; SD1 and SD2 are the standard deviations of regression slope 1 and regression slope 2, respectively; N1 and N2 are the sample sizes of regression 1 and regression 2, respectively.
As a complementary analysis to the simple regression models, we constructed a Bayesian model to eliminate the influence of climatic factors on the relationship between species richness and the spatial stability of ANPP. To exclude the effect of vegetation types on the results, we grouped sites into alpine meadow and alpine steppe, and repeated the Bayesian model for each vegetation type. The model is:
where \({CV}_{ANPP}\) is the coefficient of variation of aboveground net primary productivity; SR represents species richness. MI, MAP, MAT and ACT represent moisture index, mean annual precipitation, mean annual temperature, annual accumulated temperature, respectively. We used Markov Chain Monte Carlo (MCMC) sampling techniques in R 4.3.1 to fit all the Bayesian models.
We examined the linkages between climatic factors, SR, and ANPP at levels of quadrats using the structural equation models (SEM). Due to data loss in 3 quadrats, there are currently a total of 348 quadrat data available. We constructed a structural equation model using 348 quadrat data from alpine grasslands. Then we constructed separate structural equation models using quadrat data from alpine meadows (165 quadrats) and alpine steppe (183 quadrats). IBM SPSS Amos 24.0 was used for the SEM analysis. The SEM was tested and evaluated using five fit indices (Chi-square, P, CMIN/DF, GFI, NFI). A distribution map of sampling points was drawn using ArcGIS 10.2. IBM SPSS Statistics 24.0 was used for the covariance analysis. Other data analysis and graph plotting were implemented using R 4.3.1 (R Core Team 2023).
Results
Effects of species richness on spatial stability of ANPP
Although there were both positive and negative relationships between ANPP and species richness within the same site, they exhibited an overall positive relationship (Fig. 2, P < 0.001). There was a consistent positive relationship between SD, the reciprocal of CV, and species richness in natural grasslands (Table 1, P < 0.05). We further analyzed the effect of species richness on ANPP at both the quadrat and site levels (Fig. 2; Fig. 3). There was a consistent positive linear relationship between species richness and ANPP in natural grasslands, independent of variations in vegetation types (Fig. 2; Fig. 3). When treating species richness as a covariate, vegetation types did not have a significant impact on both the quadrat-level and site-level ANPP (Table 2, P = 0.14; Table 3, P = 0.27). Furthermore, the interaction between vegetation types and species richness had no significant effect on ANPP (Table 2, P = 0.14; Table 3, P = 0.47).

Relationships between species richness (SR) and aboveground net primary productivity (ANPP) in natural grasslands at the quadrat level. The colored short lines represent the fitted relationship between ANPP and SR in the same site. The gray line represents the overall fitting relationship between ANPP and SR in the same site

Relationships between aboveground net primary productivity (ANPP), standard deviation (SD), and species richness (SR) at the site level. The points represent the ANPP and standard deviation of ANPP at each site
Through simple linear regression analysis, we found a positive linear relationship between species richness and standard deviation, which was not influenced by variations in vegetation types (Fig. 3; Table 3). In addition, we found that an increase in species richness significantly increased the reciprocal of the coefficient of variation (Fig. 4). Similar results were also found that an increase in species richness significantly increased the reciprocal of the coefficient of variation in both alpine meadow and alpine steppe (Fig. 4, P < 0.01). When species richness was treated as a covariate, neither vegetation types nor the interaction between vegetation types and species richness had a significant impact on the standard deviation and the reciprocal of the coefficient of variation (Table 3, P > 0.05).

Relationships between species richness (SR) and the reciprocal of the coefficient of variation (reciprocal CV) in natural grasslands at the site level. The points represent the reciprocal of the coefficient of variation at each site
We then tested whether species richness could increase the spatial stability of ANPP. The results showed that the regression slope between species richness and the spatial stability of ANPP was positive, indicating a positive effect of species richness on the spatial stability of ANPP after excluding the influence of climate factors (Fig. 5). While there were differences in the effects of climate factors on the spatial stability of ANPP between alpine meadow and alpine steppe, species richness exhibited a positive effect on the spatial stability of ANPP in both vegetation types (Fig. 5). Therefore, our results explicitly indicated that the spatial stability of ANPP increased with increasing of species richness.

Bayesian models for the relationships between climatic factors, species richness (SR), and spatial stability of aboveground net primary productivity (ANPP) in alpine grasslands, alpine meadow, and alpine steppe. The points represent the mean of the fitted slopes between the reciprocal of the coefficient of variation and the influencing factors. The lines represent the 95% credible interval of the fitted slopes between the reciprocal of the coefficient of variation and the influencing factors. MAP, mean annual precipitation; MAT, mean annual temperature; ACT, annual accumulated temperature; MI, moisture index
Effects of climatic factors and species richness on ANPP
The structural equation model showed that the four climatic factors (MAT, MAP, ACT, and MI) and species richness directly or indirectly affected ANPP (Fig. 6). More than 67 percent of the variance in ANPP could be explained by MAT, ACT, MI, and species richness, indicating a great impact of climatic factors and species richness on ANPP (Fig. 6a). While there were differences in the effects of climate factors on ANPP between alpine meadow and alpine steppe, species richness had a significant positive effect on ANPP (Fig. 6). Overall, more than 55 percent of the variance in ANPP could be explained by climatic factors and species richness (Fig. 6b, c).

Structural equation model shows the linkages between mean annual precipitation (MAP) and temperature (MAT), annual accumulated temperature (ACT), moisture index (MI), species richness (SR), and aboveground net primary productivity (ANPP) at the quadrat level (N = 348 quadrats). a, b, and c represent alpine grassland, alpine meadow, and alpine steppe, respectively. Due to data loss in 3 quadrats, there are currently a total of 348 quadrat data available. The number of quadrats included in the structural equation model for alpine grassland, alpine meadow, and alpine steppe are 348, 165, and 183, respectively. All retained arrows are significant (P < 0.05). Red and blue one-way arrows represent negative and positive effects, respectively. Regression coefficients (above arrows) and variance explanatory rates (top right corner of the MI, species richness and ANPP boxes) of the fitted model are also marked in the model
The results showed that only the regression slope between the moisture index and ANPP showed a significant difference between the moderate and high species richness communities, while the slopes between other gradients were not significant (Table 4). When shifting from low species richness communities to medium species richness communities in alpine meadow and alpine steppe, there was no consistent trend of increasing or decreasing absolute values in the regression slopes between various climate factors (MAP, ACT, MAT, and MI) and ANPP (Table 5). However, the absolute values of the regression slopes between climate factors and ANPP in high species richness (19–28 species) alpine meadow communities were smaller than those in medium (10–18 species) and low (1–9 species) species richness alpine meadow communities (Table 5). This suggested that in alpine meadow communities with higher species richness, ANPP exhibited a weaker response to climatic factors, whereas in alpine meadow communities with lower species richness, ANPP showed a more violent response to climatic factors.
Discussion
The species loss greatly affects the functioning and services of ecosystems (Baldwin et al. 2014; Garcia et al. 2014; Allan et al. 2015; Seddon et al. 2016; Vellend et al. 2017; Komatsu et al. 2019). By analyzing data from 117 sites in natural grasslands, the results showed that the reciprocal of the coefficient of variation increased with increasing plant diversity. This indicates that plant diversity enhanced the spatial stability of productivity in alpine grasslands (confirming H1). Furthermore, we also found that, relative to alpine meadow communities with low plant diversity, productivity in high plant diversity communities exhibited a relatively weaker response to climatic factors. This may be because the complementarity effect generated by species with different functional traits maximizes the overall utilization of resources, thereby enhancing the stability of grassland ecosystems (Matias et al. 2013; Wang and Loreau 2014; Liang et al. 2021). The decrease in plant diversity also reduces the asynchrony among species in plant communities, potentially affecting the stability of productivity in grassland ecosystems (Olden et al. 2004; Wang and Loreau 2014; Hector et al. 2010; Isbell et al. 2015). Therefore, understanding the effect of plant diversity on the stability of annual net primary productivity is of far-reaching significance for predicting the changes in grassland ecosystems under future climatic conditions.
Plant diversity affects grassland productivity mainly through complementarity effect and selection effect (Loreau et al. 2001, 2003; Cardinale et al. 2007; Wang et al. 2019b). The complementarity effect is mainly reflected in the resource allocation or compensation among species, while the selection effect is mainly caused by interspecific competition (Wang et al. 2019b). The dominant complementarity effect leads to an increase in ANPP in the fewer-species grassland communities, thereby resulting in a strong positive effect of plant diversity on ANNP (Loreau & Hector 2001; Cardinale et al. 2007; Chen et al. 2018). However, in the richer-species grassland communities, complementarity effects weaken and interspecific competition intensifies, resulting in a weaker positive effect of plant diversity on ANPP (Loreau & Hector 2001; Wang et al. 2019b). The strength of the relationship between ANPP and plant diversity largely depends on the gradient of plant diversity in the community. These changing interactions lead to the existence of a certain peak in community productivity, rather than an unrestricted increase with increasing species diversity. In this study, there was a strong linear relationship between community productivity and plant diversity, which may be attributed to the relatively low number of sites occurrences exceeding 25 species. Within the range of plant diversity investigated in this study, the community productivity has not yet reached its peak.
Although this study did not provide a complete curve depicting the relationship between community productivity and plant diversity, it did find that plant diversity can enhance the spatial stability of community productivity. This is because species with different functional traits exhibit differences in resource utilization, thus avoiding interspecific competition. For example, different species can acquire nutrients and water from different soil layers due to variations in root distribution (Violle et al. 2012). Under conditions of spatial heterogeneity in factors such as soil nutrients and water availability, communities with high plant diversity can enhance the spatial stability of productivity due to their higher asynchrony (Wang and Loreau 2014; Wang et al. 2019a; Liang et al. 2021; Schnabel et al. 2021). Therefore, natural grassland communities with higher plant diversity exhibit higher productivity, lower variation of productivity, and higher spatial stability due to complementarity effect among species.
Previous studies have shown that plant diversity can increase the temporal stability of ecosystem processes and properties (Tilman et al. 2006; Kardol et al. 2018; Wang et al. 2019b). This study considers the impact of plant diversity on spatial stability, which is complementary to the previous experiments that only focused on the temporal stability of ecosystems. Using the reciprocal of the coefficient of variation as a stability index, we found a significant positive correlation between plant diversity and spatial stability of productivity. This positive effect was not affected by vegetation types, that is, plant diversity significantly enhanced spatial stability of productivity in both alpine meadow and alpine steppe. Some studies of other natural ecosystems have found that the relationship between plant diversity and ecosystem functioning varies with the external environment changes. For example, Paquette et al. and Jucker & Coomes found that climate and environmental factors would cause changes in the relationship between plant diversity and ecosystem function by analyzing the data of forest and dryland ecosystems, respectively (Paquette et al., 2011; Jucker et al., 2012; Maestre et al., 2012). However, in terms of overall trends, both studies support our hypothesis that species diversity stabilizes ecosystem function at large scales. In addition, we also used Bayesian analysis to eliminate the effect of climate differences on productivity variability across different sites, which further confirms the reliability of the results that increased plant diversity enhances spatial stability of productivity.
Furthermore, we tested the impact of external environmental changes on grassland productivity under different plant diversity gradients. In alpine meadow, high plant diversity communities (19–28 species) exhibit lower sensitivity of productivity to climate factors compared to low plant diversity communities (1–9 species). This indicates that high plant diversity communities can buffer the impact of climate change on productivity, thereby enhancing the stability of ANPP in alpine meadow ecosystems. Our results support the hypothesis (H2) that plant diversity may buffer the impact of ambient disturbances on ANPP at a large scale. In general, ecologists also recognize that more diverse ecosystems may be better adapted to the external environment fluctuations (Isbell et al. 2017; Pires et al. 2018; Wang et al. 2019a, b). One of the main mechanisms behind the stabilizing effect of plant diversity is the asynchronous response of different species to environmental fluctuations, where this response compensates for each other, resulting in higher stability at the community level (Yachi and Loreau 1999; Tilman et al. 2006; Wang and Loreau 2014; Liang et al. 2021). In a variable environment, species with different traits can exhibit varying responses to changes in external pressure. Species asynchrony reflects the differential responses of different species to environmental changes, whereby certain species can maintain and compensate for productivity losses resulting from the decline of other species (Schnabel et al. 2021). By exhibiting asynchronous responses, grassland species can take advantage of favorable environmental conditions. For example, some species may exhibit rapid growth and reproduction during periods of abundant rainfall, while others may have adaptations to survive and reproduce during droughts (Tilman et al. 2006; Hector et al. 2010; Isbell et al. 2015). This variability in response reduces the overall variability in community productivity and functionality, buffering environmental fluctuations and maintaining a more stable ecosystem.
Increasing plant diversity in grassland can be conducive to buffering the impact of external disturbances and ambient pressure on grassland productivity, thus improving the spatial stability of grassland productivity. This result can be attributed to two factors: first, it may be due to variations in the strength of interactions between different species resulting from different diversity gradients; second, it may be due to compensatory effects resulting from asynchrony among species (Loreau et al. 2001, 2003; Cardinale et al. 2007; Tilman et al. 2006; Hector et al. 2010; Wang and Loreau 2014; Isbell et al. 2015; Wang et al. 2019b). Based on the intrinsic link between plant diversity and ecosystem functioning (Yachi and Loreau 1999; Balvanera et al. 2006; Hector et al. 2010; Wang and Loreau 2014; Pires et al. 2016), conserving plant diversity can minimize the impact of ambient pressure on grassland productivity. Therefore, implementing appropriate management measures to increase species diversity can enhance the spatial stability of community productivity and buffer the effects of climate change on community productivity. These findings provide a theoretical basis for the stability and scientific management of alpine grassland ecosystems under the ongoing climate change.
Conclusion
Based on large-scale field surveys and observational data on the Qinghai-Tibet Plateau, this study revealed the relationship between plant diversity and productivity in natural grasslands. The results demonstrated that increasing plant diversity enhanced productivity and standard deviation of productivity in alpine grasslands, but also improved the spatial stability of productivity. This effect of plant diversity on community productivity and spatial stability of productivity is not affected by vegetation types. Furthermore, we found that climatic factor had a stronger impact on productivity in alpine meadow communities with low plant diversity, while their effect on communities with higher plant diversity is weaker. This result indicates that higher plant diversity buffers the impact of climatic factors on productivity of alpine meadow communities. These findings suggest that plant diversity is essential for improving grassland productivity and maintaining productivity stability in natural grasslands.
Data availability
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Code availability
Not applicable.
References
Allan E, Manning P, Alt F, Binkenstein J, Blaser S, Bluethgen N et al (2015) Land use intensification alters ecosystem multifunctionality via loss of biodiversity and changes to functional composition. Ecol Lett 18:1–10. https://doi.org/10.1111/ele.12469
Baldwin AH, Jensen K, Schönfeldt M (2014) Warming increases plant biomass and reduces diversity across continents, latitudes, and species migration scenarios in experimental wetland communities. Glob Change Biol 20:835–850. https://doi.org/10.1111/gcb.12378
Balvanera P, Pfisterer AB, Buchmann N, He JS, Nakashizuka T, Raffaelli D et al (2006) Quantifying the evidence for biodiversity effects on ecosystem functioning and services. Ecol Lett 9:1146–1156. https://doi.org/10.1111/j.1461-0248.2006.00963.x
Barry KE, Mommer L, van Ruijven J, Wirth C, Wright AJ, Bai YF et al (2019) The future of complementarity: disentangling causes from consequences. Trends Ecol Evol 34:167–180. https://doi.org/10.1016/j.tree.2018.10.013
Cardinale BJ, Wright JP, Cadotte MW, Carroll IT, Hector A, Srivastava DS et al (2007) Impacts of plant diversity on biomass production increase through time because of species complementarity. Proc Natl Acad Sci 104:18123–18128. https://doi.org/10.1073/pnas.0709069104
Cardinale BJ, Duffy JE, Gonzalez A, Hooper DU, Perrings C, Venail P et al (2012) Biodiversity loss and its impact on humanity. Nature 486:59–67. https://doi.org/10.1038/nature11148
Cardinale BJ, Gross K, Fritschie K, Flombaum P, Fox JW, Rixen C et al (2013) Biodiversity simultaneously enhances the production and stability of community biomass, but the effects are independent. Ecology 94:1697–1707. https://doi.org/10.1890/12-1334.1
Chen SP, Wang WT, Xu WT, Wang Y, Wan HW, Chen DM et al (2018) Plant diversity enhances productivity and soil carbon storage. Proc Natl Acad Sci 115:4027–4032. https://doi.org/10.1073/pnas.1700298114
Duffy JE, Godwin CM, Cardinale BJ (2017) Biodiversity effects in the wild are common and as strong as key drivers of productivity. Nature 549:261–264. https://doi.org/10.1038/nature23886
Elmendorf SC, Henry GHR, Hollister RD, Fosaa AM, Gould WA, Hermanutz L et al (2015) Experiment, monitoring, and gradient methods used to infer climate change effects on plant communities yield consistent patterns. Proc Natl Acad Sci 112:448–452. https://doi.org/10.1073/pnas.1410088112
Garcia RA, Cabeza M, Rahbek C, Araújo MB (2014) Multiple dimensions of climate change and their implications for biodiversity. Science 344:1247579. https://doi.org/10.1126/science.1247579
Gonzalez A, Germain RM, Srivastava DS, Filotas E, Dee LE, Gravel D et al (2020) Scaling-up biodiversity-ecosystem functioning research. Ecol Lett 23:757–776. https://doi.org/10.1111/ele.13456
Grace JB, Anderson TM, Olff H, Scheiner SM (2010) On the specification of structural equation models for ecological systems. Ecol Monogr 80:67–87. https://doi.org/10.1890/09-0464.1
Grace JB, Anderson TM, Seabloom EW, Borer ET, Adler PB, Harpole WS et al (2016) Integrative modelling reveals mechanisms linking productivity and plant species richness. Nature 529:390–393. https://doi.org/10.1038/nature16524
Hautier Y, Isbell F, Borer ET, Seabloom EW, Harpole WS, Lind WM et al (2018) Local loss and spatial homogenization of plant diversity reduce ecosystem multifunctionality. Nat Ecol Evol 2:50–56. https://doi.org/10.1038/s41559-017-0395-0
Hautier Y, Zhang P, Loreau M, Wilcox KR, Seabloom EW, Borer ET et al (2020) General destabilizing effects of eutrophication on grassland productivity at multiple spatial scales. Nat Commun 11:5375. https://doi.org/10.1038/s41467-020-19252-4
Haynes MA, Kung S, Brandt J, Yang YP, Waller DM (2014) Accelerated climate change and its potential impact on Yak herding livelihoods in the eastern Tibetan plateau. Clim Change 123:147–160. https://doi.org/10.1007/s10584-013-1043-6
Hector A, Hautier Y, Saner P, Wacker L, Bagchi R, Joshi J et al (2010) General stabilizing effects of plant diversity on grassland productivity through population asynchrony and overyielding. Ecology 91:2213–2220. https://doi.org/10.1890/09-1162.1
IPCC (2013) Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge Univ. Press
Isbell F, Craven D, Connolly J, Loreau M, Schmid B, Beierkuhnlein C et al (2015) Biodiversity increases the resistance of ecosystem productivity to climate extremes. Nature 526:574–577. https://doi.org/10.1038/nature15374
Isbell F, Gonzalez A, Loreau M, Cowles J, Díaz S, Hector A et al (2017) Linking the influence and dependence of people on biodiversity across scales. Nature 546:65–72. https://doi.org/10.1038/nature22899
Kardol P, Fanin N, Wardle DA (2018) Long-term effects of species loss on community properties across contrasting ecosystems. Nature 557:710–713. https://doi.org/10.1038/s41586-018-0138-7
Klein JA, Harte J, Zhao XQ (2007) Experimental warming, not grazing, decreases rangeland quality on the Tibetan plateau. Ecol Appl 17:541–557. https://doi.org/10.1890/05-0685
Komatsu KJ, Avolio M, Lemoine NP, Isbell F, Grman E, Houseman GR et al (2019) Global change effects on plant communities are magnified by time and the number of global change factors imposed. Proc Natl Acad Sci 116:17867–17873. https://doi.org/10.1073/pnas.1819027116
Lange M, Eisenhauer N, Sierra CA, Bessler H, Engels C, Griffiths RI et al (2015) Plant diversity increases soil microbial activity and soil carbon storage. Nat Commun 6:6707. https://doi.org/10.1038/ncomms7707
Leps J, Májeková M, Vítová A, Dolezal J, de Bello F (2018) Stabilizing effects in temporal fluctuations: management, traits, and species richness in high-diversity communities. Ecology 99:360–371. https://doi.org/10.1002/ecy.2065
Liang J, Crowther TW, Picard N, Wiser S, Zhou M, Alberti G et al (2016) Positive biodiversity-productivity relationship predominant in global forests. Science. https://doi.org/10.1126/science.aaf8957
Liang MW, Liang CZ, Hautier Y, Wilcox KR, Wang SP (2021) Grazing-induced biodiversity loss impairs grassland ecosystem stability at multiple scales. Ecol Lett 24:2054–2064. https://doi.org/10.1111/ele.13826
Liang M, Baiser B, Hallett LM, Hautier Y, Jiang L, Loreau M et al (2022) Consistent stabilizing effects of plant diversity across spatial scales and climatic gradients. Nat Ecol Evol 6:1669–1675. https://doi.org/10.1038/s41559-022-01868-y
Linders TEW, Schaffner U, Eschen R, Abebe A, Choge SK, Nigatu L et al (2019) Direct and indirect effects of invasive species: biodiversity loss is a major mechanism by which an invasive tree affects ecosystem functioning. J Ecol 107:2660–2672. https://doi.org/10.1111/1365-2745.13268
Loreau M, de Mazancourt C (2013) Biodiversity and ecosystem stability: a synthesis of underlying mechanisms. Ecol Lett 16:106–115. https://doi.org/10.1111/ele.12073
Loreau M, Hector A (2001) Partitioning selection and complementarity in biodiversity experiments. Nature 412:72–76. https://doi.org/10.1038/35097128
Loreau M, Naeem S, Inchausti P, Bengtsson J, Grime JP, Hector A et al (2001) Biodiversity and ecosystem functioning: current knowledge and future challenges. Science 294:804–808. https://doi.org/10.1046/j.1523-1739.1995.09040742.x
Loreau M, Mouquet N, Gonzalez A (2003) Biodiversity as spatial insurance in heterogeneous landscapes. Proc Natl Acad Sci 100:12765–12770. https://doi.org/10.1073/pnas.2235465100
Ma ZY, Liu HY, Mi ZR, Zhang ZH, Wang YH, Xu W et al (2017) Climate warming reduces the temporal stability of plant community biomass production. Nat Commun 8:15378. https://doi.org/10.1038/ncomms15378
Matias MG, Combe M, Barbera CG, Mouquet N (2013) Ecological strategies shape the insurance potential of biodiversity. Front Microbiol 3:432. https://doi.org/10.3389/fmicb.2012.00432
Moore DS, McCabe GP, Craig BA (1993) Introduction to the Practice of Statistics. W. H. Freeman and Company, New York, United States.
Olden JD, Poff NL, Douglas MR, Douglas ME, Fausch KD (2004) Ecological and evolutionary consequences of biotic homogenization. Trends Ecol Evol 19:18–24. https://doi.org/10.1016/j.tree.2003.09.010
Patrick CJ, McCluney KE, Ruhi A, Gregory A, Sabo J, Thorp JH (2021) Multi-scale biodiversity drives temporal variability in macrosystems. Front Ecol Environ 19:47–56. https://doi.org/10.1002/fee.2297
Pires APF, Marino NAC, Srivastava DS, Farjalla VF (2016) Predicted rainfall changes disrupt trophic interactions in a tropical aquatic ecosystem. Ecology 97:2750–2759. https://doi.org/10.1002/ecy.1501
Pires APF, Srivastava DS, Farjalla VF (2018) Is biodiversity able to buffer ecosystems from climate change? What we know and what we don’t. Bioscience 68:273–280. https://doi.org/10.1093/biosci/biy013
Qin XJ, Sun J, Wang XD (2018) Plant coverage is more sensitive than species diversity in indicating the dynamics of the above-ground biomass along a precipitation gradient on the Tibetan Plateau. Ecol Indic 84:507–514. https://doi.org/10.1016/j.ecolind.2017.09.013
R Core Team (2023) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org/.
Schnabel F, Liu XJ, Kunz M, Barry KE, Bongers FJ, Bruelheide H et al (2021) Species richness stabilizes productivity via asynchrony and drought-tolerance diversity in a large-scale tree biodiversity experiment. Sci Adv. https://doi.org/10.1126/sciadv.abk1643
Seddon AWR, Macias-Fauria M, Long PR, Benz D, Willis KJ (2016) Sensitivity of global terrestrial ecosystems to climate variability. Nature 531:229–232. https://doi.org/10.1038/nature16986
Shi Z, Xu X, Souza L, Wilcox K, Jiang LF, Liang JY et al (2016) Dual mechanisms regulate ecosystem stability under decade-long warming and hay harvest. Nat Commun 7:11973. https://doi.org/10.1038/ncomms11973
Solar RRC, Barlow J, Ferreira J, Berenguer E, Lees AC, Thomson JR et al (2015) How pervasive is biotic homogenization in human-modified tropical forest landscapes? Ecol Lett 18:1108–1118. https://doi.org/10.1111/ele.12494
Steudel B, Hector A, Friedl T, Löfke C, Lorenz M, Wesche M et al (2012) Biodiversity effects on ecosystem functioning change along environmental stress gradients. Ecol Lett 15:1397–1405. https://doi.org/10.1111/j.1461-0248.2012.01863.x
Tilman D, Reich PB, Knops JMH (2006) Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature 441:629–632. https://doi.org/10.1038/nature04742
Tilman D, Isbell F, Cowles JM (2014) Biodiversity and ecosystem functioning. Annu Rev Ecol Evol S 45:471–493. https://doi.org/10.1146/annurev-ecolsys-120213-091917
Vellend M, Dornelas M, Baeten L, Beauséjoer R, Brown CD, De Frenne P et al (2017) Estimates of local biodiversity change over time stand up to scrutiny. Ecology 98:583–590. https://doi.org/10.1002/ecy.1660
Violle C, Enquist BJ, Mcgill BJ, Jiang L, Albert CH, Hulshof C et al (2012) The return of the variance: intraspecific variability in community ecology. Trends Ecol Evol 27:244–252. https://doi.org/10.1016/j.tree.2011.11.014
Wang SP, Loreau M (2014) Ecosystem stability in space: α, β and γ variability. Ecol Lett 17:891–901. https://doi.org/10.1111/ele.12292
Wang SP, Loreau M (2016) Biodiversity and ecosystem stability across scales in metacommunities. Ecol Lett 19:510–518. https://doi.org/10.1111/ele.12582
Wang SP, Lamy T, Hallett LM, Loreau M (2019a) Stability and synchrony across ecological hierarchies in heterogeneous metacommunities: linking theory to data. Ecography 42:1200–1211. https://doi.org/10.1111/ecog.04290
Wang YF, Cadotte MW, Chen YX, Fraser LH, Zhang YH, Huang FM et al (2019b) Global evidence of positive biodiversity effects on spatial ecosystem stability in natural grasslands. Nat Commun 10:3207. https://doi.org/10.1038/s41467-019-11191-z
Wang SP, Loreau M, de Mazancourt C, Isbell F, Beierkuhnlein C, Connolly J et al (2021) Biotic homogenization destabilizes ecosystem functioning by decreasing spatial asynchrony. Ecology 102:1–10. https://doi.org/10.1002/ecy.3332
Weigelt A, Schumacher J, Roscher C, Schmid B (2008) Does biodiversity increase spatial stability in plant community biomass? Ecol Lett 11:338–347. https://doi.org/10.1111/j.1461-0248.2007.01145.x
Wilcox KR, Tredennick AT, Koerner SE, Grman E, Hallett LM, Avolio ML et al (2017) Asynchrony among local communities stabilises ecosystem function of metacommunities. Ecol Lett 20:1534–1545. https://doi.org/10.1111/ele.12861
Worm B, Barbier EB, Beaumont N, Duffy JE, Folke C, Halpern BS et al (2006) Impacts of biodiversity loss on ocean ecosystem services. Science 314:787–790. https://doi.org/10.1126/science.1132294
Yachi S, Loreau M (1999) Biodiversity and ecosystem productivity in a fluctuating environment: the insurance hypothesis. Proc Natl Acad Sci 96:1463–1468. https://doi.org/10.1073/pnas.96.4.1463
Zhang Y, He N, Loreau M, Pan QM, Han XG (2018) Scale dependence of the diversity–stability relationship in a temperate grassland. J Ecol 106:1277–1285. https://doi.org/10.1111/1365-2745.12903
Acknowledgements
The authors thank Prof. Shaopeng Wang for sharing their valuable insights and comments on this study manuscript draft.
Funding
This research was funded by the National Natural Science Foundation of China (NSFC 32230068, NSFC 41722107), the Strategic Priority Research Program of the Chinese Academy of Sciences (XDB40000000).
Author information
Authors and Affiliations
Contributions
G.L.W. and J.S. conceived the research. J.S. contributed field experimental data. Z.C. and G.L.W. analyzed data. Z.C. and G.L.W. wrote the first draft of the manuscript, and Z.C., G.L.W., and J.S. contributed substantially to revisions.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethics approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Additional information
Communicated by Brian J. Wilsey .
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Cui, Z., Sun, J. & Wu, GL. Plant diversity increases spatial stability of aboveground productivity in alpine grasslands. Oecologia 205, 27–38 (2024). https://doi.org/10.1007/s00442-024-05552-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-024-05552-9