
Abstract
The treatment for trichomoniasis, based on 5′-nitroimidazol agents, has been presenting failures related to allergic reactions, side effects, and the emergence of resistant isolates. There are no alternative drugs approved for the treatment of these cases; thus, the search for new active molecules is necessary. In this scenario, chalcones have been extensively studied for their promising biological activities. Here, we presented the synthesis of three hydroxychalcones (3a, b, and c), in vitro and in silico analyses against Trichomonas vaginalis. The in vitro biological evaluation showed that hydroxychalcone 3c presented anti-T. vaginalis activity, with complete death in 12 h of incubation at minimum inhibitory concentration (MIC) of 100 μM. 3c showed a dose-dependent cytotoxicity against mammalian VERO cell line, but the association of 3c at 12.5 μM and metronidazole (MTZ) at 40 μM showed 95.31% activity against T. vaginalis trophozoites after 24 h of exposure and did not affect the VERO cell growth, appearing to be a good alternative. In silico analysis by molecular docking showed that 3c could inhibit the activity of TvMGL (methionine gamma-lyase), TvLDH (lactate dehydrogenase), and TvPNP (purine nucleoside phosphorylase) affecting the T. vaginalis survival and also suggesting a different mechanism of action from MTZ. Therefore, these results propose that hydroxychalcones are promising anti-T. vaginalis agents and must be considered for further investigations regarding trichomoniasis treatment.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Trichomoniasis is the most common and prevalent non-viral sexually transmitted infection (STI) in the world. Based on estimates of the World Health Organization, trichomoniasis affects more than 276 million people every year worldwide (WHO 2012; Leitsch 2016). The treatment for human trichomoniasis is currently based on 5′-nitroimidazol agents, more specifically metronidazole (MTZ) and tinidazole (TNZ), which are the only drugs approved and recommended by the Food and Drug Administration (FDA-EUA n.d). However, this course of treatment has been showing some failures, which can be attributed to allergic reactions, side effects, and the emergence of resistant isolates to 5′-nitroimidazoles (Paulish-Miller et al. 2014). There are no alternative drugs approved for the treatment of such cases, establishing a dependence on a single therapeutic class, which is problematic. Thus, the search for alternative treatments to 5′-nitroimidazoles is necessary and has been frequently studied. In this context, synthetic compounds represent an important tool for the drug discovery (Bala and Chhonker 2018; de Brum Vieira et al. 2015).
In the discovery process for new antiparasitic drugs, the design and synthesis of novel compounds with high specificity is particularly important for the future development of clinically useful drugs. Thus, Trichomonas vaginalis proteins, such as lactate dehydrogenase (LDH), methionine gamma-lyase (MGL), and purine nucleoside phosphorylase (PNP), may serve as drug targets to predict potential inhibitors of T. vaginalis, because they differ from human proteins and are important for T. vaginalis survival (Setzer et al. 2017).
The use of the standard scaffold of chalcones (1,3-diphenyl-2-propen-1-one) enables a multiplicity of substitutions because of their easy synthesis, simple chemistry, and straightforward hydrogen atom manipulation, and such changes, e.g., addition of different chemical groups/radicals, can offer novel biological activities (Gomes et al. 2017; Wong 1968). Chalcones are α,β-unsaturated aromatic ketones reported as open-chain precursors for biosynthesis of flavonoids and isoflavonoids and widely distributed in nature, as part of foods and beverages. Its biological properties have been extensively studied, presenting promising activities such as antitumoral (Mahapatra et al. 2015), antifungal (Palanco et al. 2017), antioxidant, analgesic, anti-inflammatory (Abdellatif et al. 2015), antiparasitic (Borsari et al. 2017), and more importantly anti-T. vaginalis (Trein et al. 2019).
To indicate a new alternative for trichomoniasis treatment, we describe here the synthesis and antiparasitic activity of three chalcone analogues, more specifically hydroxychalcones, alone and in association with metronidazole. In addition, analyses of the biochemical effects against trophozoites of T. vaginalis, cytotoxicity against VERO cells, and molecular docking with T. vaginalis enzymes were performed to indicate a more in-depth profile on the behavior of these compounds.
Materials and methods
Chemistry: synthesis and identification of hydroxychalcones
All reagents and solvents employed in the synthesis of chalcones were of analytical grade and purchased from Sigma-Aldrich Co. (St. Louis, MO, USA). Reactions were monitored by thin-layer chromatography (TLC) using hexane/ethyl acetate and observed under ultraviolet light. Hydroxychalcones 3a, 3b, and 3c were synthesized by Claisen-Schmidt condensation based on a previously described procedure (Fig. 1) (Wang et al. 2014). Briefly, an aqueous solution of sodium hydroxide 40% (2.0 mL) was added to a round bottom flask containing 15 mL of ethanol and 1.0 mmol of 2′-hydroxyacetophenone, in an ice bath. After 30 min, 1.2 mmol of the aromatic aldehyde was added and the mixture was stirred at room temperature until completion. Diluted hydrochloric acid was then used to neutralize the reaction mixture; the precipitate was filtered by gravity filtration and washed with cold water. The dried crude solid was purified by recrystallization with hot ethanol. Chalcones 3b and 3c were reported before by our group (Da Silva et al. 2018a, b), and products 3a–c were confirmed by their melting points (Fisatom 430 apparatus), infrared spectroscopy (FTIR-Prestige 21), and mass spectrometry (GC-MS-QP 2010 SE Standard Gas Chromatograph-Mass Spectrometer with an AOC-20 automatic injector, RTx-5MS column, using helium as carrier gas). The data related to the identification of synthetic hydroxychalcones is displayed in Table 1.

Synthesis of hydroxychalcones 3a–3c
VERO cell line culture
Mammalian VERO cell line, obtained from the Bank of Cells of Rio de Janeiro (BCRJ, University of Rio de Janeiro), was cultured in monolayer in RPMI 1640 medium (Vitrocell Embriolife, Campinas, Brazil), supplemented with 10% fetal bovine serum (Gibco, Grand Island, NY, USA), 1% l-glutamine, and 1% penicillin/streptomycin, and kept in a CO2 incubator with humidified air (5% CO2 at 37 °C).
Cytotoxicity assay against mammalian VERO cell line
To evaluate the cytotoxicity of hydroxychalcones, VERO cells were seeded in 96-well microtiter plates (Cral®) at a density of 2 × 104 cells per well in a volume of 100 μL and incubated at 37 °C in a humidified air atmosphere and 5% CO2 for 24 h. When cells reached > 80% confluence, the cells were treated with hydroxychalcone, previously diluted in 0.6% dimethyl sulfoxide (DMSO), at concentrations varying from 12.5 to 200 μM and in association with MTZ. Untreated cells were used as negative controls. After 24 h of incubation in the CO2 incubator with humidified air (5% CO2 at 37 °C), viability of the cell lines was evaluated by the MTT 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide method. Differences in viability after treatment were measured by spectrophotometry through a plate reader (Mindray) at 492 nm. All assays were performed independently at least three times in quadruplicate, and results were expressed as the percentage of cell growth inhibition in comparison with the negative control.
Trichomonas vaginalis culture
T. vaginalis 30236 isolate, obtained from American Type Culture Collection (ATCC), which is normally susceptible to metronidazole, was used in this study. Trophozoites were axenically cultured in vitro in a trypticase-yeast extract-maltose (TYM) medium without agar (pH 6.0), supplemented with 10% sterile heat-inactivated bovine serum and 5 mg/mL streptomycin, and then incubated at 37 °C (Diamond 1957).
Anti-T. vaginalis assays
Anti-T. vaginalis activity, minimum inhibitory concentration (MIC), association with metronidazole, and kinetic growth assays were performed following the methodology of Sena-Lopes et al. (2017). Initially, for the anti-T. vaginalis assays, cultures with viability equal or higher than 95% were used after being analyzed through observation of motility, morphology, and trypan blue dye exclusion (0.4%) assay under a light microscope at × 400 magnification. The activity of hydroxychalcones was screened in vitro against T. vaginalis in 96-well microtiter plates (Cral®). A volume of 150 μL of trophozoite solution was seeded at an initial density of 2.6 × 105 trophozoites/mL and incubated with 50 μL of a solution containing TYM medium and hydroxychalcones at a final concentration of 100 μM, previously diluted in 0.6% DMSO. Three controls were applied in each assay: a negative control containing only trophozoites, a 0.6% DMSO control, and a positive control containing 100 μM of MTZ (Sigma-Aldrich, St. Louis, USA). The microtiter plates were incubated at 37 °C with 5% CO2 for 24 h. After that, a preparation containing trophozoites and trypan blue (0.4%) at a ratio of 1:1 was counted in a Neubauer chamber, and trophozoite motility and morphology was analyzed by light microscopy while viability was assessed through trypan blue dye exclusion (0.4%) assay. Only the compounds that reduced the viability of parasites by 100% were used in the following experiments, from now on referred as active hydroxychalcone(s).
The MIC values against T. vaginalis were established in the same conditions as described above with variations on the concentrations of the active hydroxychalcone. After MIC determination, confirmation was performed by transferring the culture solution from MIC wells and from the concentrations directly below and above, as well as controls, to tubes containing fresh TYM medium which were then reincubated at 37 °C with 5% CO2. Trophozoites were counted in a Neubauer chamber every 24 h during 96 h to confirm MIC, and the viability was assessed by trypan blue dye exclusion (0.4%) assay.
The effect of the association between the active hydroxychalcone and MTZ was evaluated following the methodology of Hübner et al. (2016), established in 96-well microtiter plates where trophozoites were treated considering the association of the following incubation conditions: 40 μM or 20 μM of MTZ + active hydroxychalcone at non-cytotoxic concentrations for the VERO cell line.
A kinetic growth curve was established to obtain a more accurate profile for hydroxychalcone activity against T. vaginalis. Trophozoite viability was observed by light microscopy after incubation at respective MIC or in association with MTZ. Growth analysis was performed at 1, 6, 12, 24, 48, 72, and 96 h by trypan blue dye exclusion (0.4%) assay.
For all assays, untreated parasites were used as negative controls. Plates were incubated at 37 °C with 5% CO2 for 24 h. After that, trophozoite motility and morphology was analyzed by light microscopy counted in a Neubauer chamber, while viability was assessed through trypan blue dye exclusion (0.4%) assay. All assays were performed independently at least three times in triplicate, and results were expressed as the percentage of viable trophozoites in comparison with the negative control.
Molecular docking
The binding modes of active hydroxychalcone with T. vaginalis methionine gamma-lyase (TvMGL; PDB: 1E5E), lactate dehydrogenase (TvLDH; PDB 5A1T), and purine nucleoside phosphorylase (TvPNP; PDB 1Z36) were predicted using the software AutoDock Vina 1.1.2 (Trott and Olson 2009). The crystal structure of TvMGL, TvLDH, and TvPNP was retrieved from the Protein Data Bank (http://www.pdb.org/pdb/) and prepared by Chimera 1.5.3, including removal of ligands (Pettersen et al. 2004). A grid box size covering the residues in the active site of the proteins was implemented by AutoDock Tools 1.5.6 (Morris et al. 2009). The active hydroxychalcone was built and optimized in the software Avogadro 1.1.1. (Hanwell et al. 2012). Docking poses of the investigated compound were visualized using Accelrys Discovery Studio 3.5.
In silico drug-likeness evaluation
The active hydroxychalcone was studied by applying the Lipinski’s rule of five, in which the criteria, established by Lipinski, to evaluate the theoretical oral bioavailability were as follows: partition coefficient (miLogP) ≤ 5.0, molecular weight ≤ 500, hydrogen bond acceptors ≤ 10, and donors ≤ 5. The compounds that violate one or more rules demonstrate an unattractive oral bioavailability. In addition, physicochemical property analyses of active hydroxychalcone were performed on software ADMET Predictor with parameters of 1 mg/kg of 3c administered orally in humans (Lipinski 2004).
Statistical analysis
Statistical analysis was carried out by one-way analysis of variance (ANOVA) using a probability value of p < 0.05 using the GraphPad Prism 8.2.0 software. For screening and MIC assays, Tukey’s post-test was conducted to allow multiple comparisons between all treatments. For association with metronidazole and cytotoxicity assay, Dunnett’s post-test was conducted to identify significant differences between the negative control and the means of different treatments. For the kinetic growth curve, two-way ANOVA was conducted followed by Bonferroni post-tests. The IC50 and CC50 values were assessed through a non-linear regression model.
Results
Synthesis and identification of hydroxychalcones
Regarding the synthesis of the hydroxychalcones, it can be seen in Table 1 that three products were obtained in moderate yields (32–60%). Infrared and mass spectra of products 3a, 3b, and 3c are displayed in Figs. 2 and 3 and were in agreement with the proposed structures. Infrared spectra showed absorption bands related to the carbonyl (1635 cm−1) and hydroxyl groups (3300 cm−1). The molecular ions were identified for all molecules, matching their exact mass, as demonstrated in the mass spectra, indicating that the target products were obtained satisfactorily.

Mass spectrum of hydroxychalcones 3a (A), 3b (B), and 3c (C)

Infrared spectrum of hydroxychalcones 3a (A), 3b (B), and 3c (C)
Cytotoxicity assay against mammalian VERO cell line
The MTT assay showed that 3c decreased cell viability in a dose-dependent manner. The CC50 value was established as 118.9 μM, and a significant cell growth inhibition was observed in VERO cells after treatment with 3c at MIC concentration, which inhibited 47.7% of growth. However, from the concentration of 25 μM and below, there was no inhibition of cell growth. Thus, the association of 3c at 12.5 μM and MTZ at 40 μM did not inhibit cell growth after 24 h of exposure, showing no significant difference when compared with the negative control (NC) (Fig. 4).

Cytotoxicity effect of hydroxychalcone 3c alone at 6.25–200 μM and of 3c (12.5 μM) in association with MTZ (40 μM) against VERO cell line. Cell proliferation was investigated by MTT assay. Data are expressed as means ± SD from three independent experiments, analyzed by one-way ANOVA followed by Dunnett’s multiple comparison test. (*) represents a significant difference when compared with the negative control (NC). For all p < 0.05
Anti-T. vaginalis assays
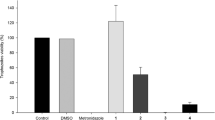
Analysis of the data obtained in the evaluation of anti-T. vaginalis assay showed that hydroxychalcones 3a, 3b, and 3c at 100 μM, after 24 h of exposure, induced 12%, 54%, and 95% of trophozoite death, respectively, showing the potential antiparasitic activity of all three compounds. Nonetheless, at tested concentration, 3c was the most potent without showing a significant statistical difference from the positive control (MTZ at 100 μM), as indicated by letter “b” in Fig. 5A. As expected, the positive control was stained with trypan blue (0.4%) and exhibited negative motility, reducing 100% viability of trophozoites. In addition, the control group DMSO (0.6%) showed positive motility, did not stain with trypan blue (0.4%), and did not present significant statistical difference from the negative control (untreated trophozoites), as indicated by letter “a” (Fig. 5A).

(A) Anti-T. vaginalis activity of hydroxychalcones 3a, 3b, and 3c at 100 μM confirmed by trypan blue dye exclusion assay (0.4%) after 24 h of exposure. (B) Determination of 3c MIC after T. vaginalis 30236 isolate treatment at 50, 60, 70, 80, 90, and 100 μM after 24 h of exposure. (C) Anti-T. vaginalis effects of associations of different concentrations of 3c and MTZ. (D) Kinetic growth curve of T. vaginalis 30236 isolate after treatment with 3c alone and in association with MTZ at the period of 1, 6, 12, 24, 48, 72, and 96 h. Vehicle for solubilization at 0.6% (DMSO), metronidazole at 100 μM (MTZ), negative control (NC). Data are expressed as means ± SD from three independent experiments: (A, B) analyzed by one-way ANOVA followed by Tukey’s multiple comparison test, (C) analyzed by one-way ANOVA followed by Dunnett’s multiple comparison test, and (D) analyzed by two-way ANOVA followed by Bonferroni post-tests. Different letters (a–g) indicate a significant difference between treatments, and (*) represents a significant difference when compared with the negative control (NC). For all p < 0.05
The IC50 was established as 50.64 μM, and as a result of MIC assay, the ideal concentration for complete trophozoite death was established as 100 μM, which showed a significant difference from the positive control (MTZ at 100 μM) as indicated by letter “b” (Fig. 5B). The effect of 3c association with MTZ against T. vaginalis trophozoites showed that hydroxychalcone 3c at 12.5 μM in association with MTZ at 40 μM reduced the trophozoite viability by 98% after 24 h of incubation (Fig. 5C). The kinetic growth curve showed that time of exposure affect trophozoite growth after treatment with 3c alone and in association with MTZ. 3c alone at MIC concentration was able to reduce the trophozoite growth by 86%, and 3c at 12.5 μM in association with MTZ at 40 μM reduced trophozoite growth by 89.2%, both at 12 h of exposure (5D) with complete death characterized at 24 h exposure (Fig. 5A, C).
Molecular docking
As depicted in Fig. 6A, B, the binding mode of the compound 3c with the active site of TvMGL involves conventional hydrogen bonds with THR114 (2.65 Å) and LYS209 (3.14 Å). In addition, pi-donor hydrogen bonds, pi-alkyl, pi-anion, and van der Waals forces contribute to the binding mode of compound 3c with TvMGL, yielding binding free energy (ΔGbind) of − 7.8 kcal/mol. A likely binding mode of the compound 3c in the active site of TvLDH is depicted in Fig. 6C, D (ΔGbind of − 7.4 kcal/mol). A hydrogen bond is formed between compound 3c and THR239 residue (2.40 Å), while non-covalent interactions with other amino acid residues help to maintain the binding mode of the compound 3c with TvLDH. The best mode of compound 3c with TvPNP is represented in Fig. 6E, F, with a docking score of − 7.8 kcal/mol. ASP204 (2.40 Å) is involved in a hydrogen bond with compound 3c, while pi-alkyl and pi-sigma interactions and van der Waals forces are formed between 3c and other amino acid residues in the active site of TvPNP. In addition, the 3c physicochemical properties showed that 3c does not violate any of the 5 Lipinski rules, meaning it could have favorable properties for administration in humans and may be used as candidate to the study and development of a new antiparasitic agent (Table 2).

Predicted binding mode of compound 3c in the active site of TvMGL (A and B), TvLDH (C and D), and TvPNP (E and F)
Discussion
The treatment of trichomoniasis relies on a single class of drugs, the 5′-nitroimidazoles, which are the most prescribed and effective drug against T. vaginalis. But, the development of resistant T. vaginalis isolates across the globe has made the research and development of alternatives to metronidazole treatment extremely necessary (Bala and Chhonker 2018).
In the development of new drugs, chalcones and chalcone derivatives possess a large number of different biological activities, which are highly appreciated in many areas, because they are bioactive against virtually all eukaryotes and some prokaryotes, and their molecular targets are numerous (Zhou 2015). During the last decades, numerous investigations have been carried out on the pharmacological activities of natural and synthetic chalcones. Some studies show that chalcones tested against T. vaginalis are usually associated with other chemical class, such as a study by Anthwal et al. (2014) that shows that metronidazole-chalcone conjugates exhibited anti-T. vaginalis activity against MTZ-susceptible and MTZ-resistant T. vaginalis strains and another by Singh et al. (2016) that described a series of chalconyl blended triazole allied silatranes displaying significant activity against T. vaginalis.
Another alternative is the addition of substituents in aromatic rings, developing chalcone derivatives that might present different biological activities. In this manner, a recent study showed the potential activity of 3′-aminochalcone against T. vaginalis (Trein et al. 2019 ), and here we present the potential activity of another type of chalcone derivate, the hydroxychalcones. Until now, there is no data reported about hydroxychalcones with activity against T. vaginalis trophozoites; however, Borsari et al. (2017) discovered methoxylated 2′-hydroxychalcones as potent anti-Trypanosoma brucei agents, showing antiprotozoal activity. Thus, in this study, we evaluate the use of three hydroxychalcones for in vitro potential against T. vaginalis.
All synthetic products tested in this study are chalcones containing hydroxyl groups in their structure; specifically, all of them possess a hydroxyl group at the –ortho position of ring A (derived from the reagent 2′-hydroxyacetophenone). The different results determined in the anti-T. vaginalis assay demonstrated that although being from the same subclass of compounds, the structural differences influenced their antiparasitic potential. Hydroxychalcone 3a is derived from cinnamaldehyde, which itself has been reported to exhibit anthelmintic activity against Dactylogyrus intermedius (Ling et al. 2015) and, because of that, has a central 5-carbon linker and an additional unsaturation when compared with 3b and 3c. However, product 3a did not exert anti-T. vaginalis activity at the tested concentration so that the additional unsaturation does not seem to have an effect against this strain, or it possibly stabilizes the hydroxyl group in a way that prevents an inhibitory interaction with T. vaginalis cells. This is, nonetheless, an interesting result because, as shown by Rezk et al. (2002), a certain biological activity can differ significantly for a starting reagent and its derived synthetic product.
Hydroxychalcones 3b and 3c have a 3-carbon linker, but 3c has another hydroxyl group at the –meta position of ring B (derived from the 3-hydroxybenzaldehyde). Borsari et al. (2017) investigated the antiparasitic potential of chalcones from the same class reported herein (2′-hydroxychalcones) but containing also methoxy substituents in the aromatic rings. Most of the methoxylated 2′-hydroxychalcones with an antiparasitic activity against Trypanosoma brucei were also substituted in the –meta position of ring B, which could indicate an important pattern of substitution associated with a pharmacological potential. It seems that the additional hydroxyl substituent present at chalcone 3c contributed for its higher potential, while the length of the conjugated bonds (i.e., 3a) did not seem to play a crucial role for the antiparasitic activity. Other synthetic hydroxychalcones with different substitution patterns can be obtained for a better understanding of the structural motifs required for an anti-T. vaginalis activity.
To evaluate the cytotoxicity of hydroxychalcone 3c, we used mammalian VERO cell line. After 24 h of exposure, the CC50 value was established as 118.9 μM and 3c at MIC concentration showed a cell growth inhibition of 47.7%, which is significantly high. No other previous report was found with the evaluation of hydroxychalcone 3c in VERO cells, although two research articles performed toxicity assessments of molecules with the same structures of 3b and 3c. Lee et al. (2014) investigated the in vivo toxicities of some synthetic chalcones in zebrafish embryos, in which compounds equal to 3b and 3c (original codes: 1a and 1d, respectively) did not show significant differences from the untreated control in concentrations of up to 5 ppm (≈ 20 μM). At a concentration of 3 ppm of 3c (≈ 12.5 μM considering its molar weight of 240.25), the survival rate was kept near 100% for 60 h post-fertilization. These results are in agreement with our in vitro assays, as the cell viability starts to decrease at concentrations above 25 μM. Forejtníková et al. (2005) demonstrated that hydroxychalcones, including 3b and 3c, display in vitro chemoprotective and toxic potentials depending on the tested concentration. In their report, chalcone equal to 3c (original code: chalcone 2′,3-diOH; Table 1) inhibited 10% of CYP1A-dependent EROD activity and its IC50 cytotoxicity in rat liver epithelial WB-F344 cells was higher than 50 μM.
Considering 3c cytotoxicity on VERO cells, the pharmacological effects of chalcone 3c are dose-dependent and could be optimized to avoid adverse effects. Other synthetic approaches could be attempted to enhance the antiparasitic activity of chalcone 3c and decrease its cytotoxicity effect, such as hydrogenation of its double bond to form a dihydrochalcone (Forejtníková et al. 2005), or cyclization of 3c to yield a chromenone (Badavath et al. 2016). Other possibility is the cytotoxicity evaluation in other cell lines, to get a better understanding of this cytotoxic effect, and alternatives such as nanoencapsulation and topical application have many advantages for drug delivery, because they can increase compound’s interaction with tissues and cells, bioavailability, and drug targeting consequently resulting in increased efficacy and decreased drug adverse effects (Bouchemal et al. 2017; Frank et al. 2015). Thereby, these approaches can allow the use of lower concentrations of treatment.
Due to the limited treatment options for trichomoniasis, the association of different chemical classes is a good alternative and allows the use of lower doses of the constituents, a situation that may reduce adverse reactions and overcome the resistant strain problem (Tallarida 2011). To confirm if the use of lower concentrations of 3c could be applied in the treatment of trichomoniasis, we chose to test the effect of 3c in association with MTZ in different concentrations determined from 3c MIC value (100 μM) and MTZ MIC value (80 μM) against T. vaginalis ATCC 30236 trophozoites, assessed under laboratory conditions (data not shown).
The best association was the use of 3c at 12.5 μM and MTZ at 40 μM that reduced trophozoite viability by 89.2% in 12 h of exposure and led to complete death in 24 h, similar to anti-T. vaginalis assay results for 3c alone at MIC concentration. The results demonstrate that the interaction between both compounds reduces the concentration required to cause parasite death. Since 3c concentration of 12.5 μM did not affect the growth of mammalian VERO cells, this is a good alternative to reduce the possible side effects caused by cytotoxicity. In addition, this association also allows the use of a lower concentration of metronidazole. In cases of resistance to this drug, it is necessary to increase the treatment concentration, increasing the possibility of adverse effects such as headache, nausea, diarrhea, and a metallic taste, and consequently decrease or impair treatment adhesion by the patient (Kissinger 2015).
Molecular docking analyses with three T. vaginalis enzymes were performed to investigate the possible molecular mechanism responsible for the anti-T vaginalis activity of compound 3c. TvMGL is a pyridoxal-5-phosphate-dependent enzyme that catalyzes an α,γ-elimination reaction on l-methionine to generate ammonia, methanethiol, and α-oxobutyrate (Lockwood and Coombs 1991). Since mammals do not have MGL, developing drugs targeting this enzyme is a rational strategy to reduce the toxicity following anti-T vaginalis treatment. TvLDH plays an important role in T. vaginalis metabolism via generation of NAD+ required for glycolysis through the reduction of pyruvate to lactate (Steindel et al. 2016). TvPNP is important for T. vaginalis survival, since these protozoa lack de novo synthesis of purine nucleosides, relying on the functions of TvPNP and nucleoside kinase (Rinaldo-Matthis et al. 2007). This metabolic feature makes the purine salvage pathway a key target for antiparasitic chemotherapy, reinforcing the promising anti-T. vaginalis action of 3c.
The binding mode of compound 3c with the active site of TvMGL, TvLDH, and TvPNP shows that 3c could inhibit the active sites of these enzymes, affecting T. vaginalis survival as evidenced in our in vitro studies. Nonetheless, the activity of these enzymes in the presence of compound 3c should be further investigated. The molecular docking studies are in agreement with the preliminary anti-T. vaginalis screening as they indicate that the additional meta-hydroxyl substituent of chalcone 3c is an important binding site for TvMGL and TvLDH. As discussed before, carbonyl molecules containing a hydroxyl group at the –ortho position of the ring may form intramolecular bonding, which would prevent these groups from forming the hydrogen bonds (Da Silva et al. 2018a, b; Lee et al. 2014). Therefore, for hydroxychalcone 3c, even if that happened, the meta-hydroxy group would still be available for interaction with the evaluated enzymes, which would not be possible for chalcones 3a and 3b. Also, when a hydroxyl group loses its hydrogen, it forms a phenoxy radical with unpaired electrons which could be responsible for the predicted alkyl bonding.
Since MTZ did not present a good binding mode with the tested enzymes (data not shown), we believe that the association of different mechanisms of action of compounds 3c and MTZ is responsible for the parasites’ death. Whereas MTZ mechanism of action is based on drug penetration into the cell wall of T. vaginalis by passive diffusion, activation in the hydrogenosomes, anaerobic reduction of the nitro group by the enzyme pyruvate ferredoxin oxidoreductase (PFOR) forming nitro cytotoxic radicals, inhibition of the synthesis and degradation of DNA strands, and consequently the death of trophozoites (Petrin et al. 1998), here we suggest that 3c mechanism of action may be the inhibition of three important enzymes for T. vaginalis survival.
An effective drug must reach its specific target in sufficient concentration since early stages, such as biologic screenings for identification of a potential molecule (Gajdács 2019). Thus, computer models constitute valid alternatives to perform assessments of absorption, distribution, metabolism, and excretion (ADME) increasingly earlier in the drug development process (Daina et al. 2017). In this work, the drug-likeness was established from structural or physicochemical inspections. The hydroxychalcone 3c showed properties probably compatible with an acceptable pharmacokinetic profile, such as being available for oral administration, being absorbed, and being bioavailable in 24 h. Besides that, the results of the interaction between 3c with cytochromes P450 (CYP), especially with the isoforms CYP1A2, CYP2C9, CYP2E1, allow to infer the occurrence of metabolic biotransformation and consequently lower probability of adverse effects, thus demonstrating the pharmacological potential of these compounds for the development of a new anti-T. vaginalis drug.
Conclusion
Hydroxychalcone 3c demonstrated a potent anti-T. vaginalis activity against ATCC 30236 isolate, and the association of 3c and MTZ in lower concentrations also displayed a good anti-T. vaginalis activity, not affecting VERO cell growth, which indicates that this association may not cause damage to human cells and can be considered a new alternative treatment for trichomoniasis. In addition, enzymes related to T. vaginalis survival could be inhibited by 3c binding, suggesting a molecular mechanism of action different to MTZ, which allows lower doses of both compounds, reducing possible adverse effects caused by high cytotoxicity, and can also prevent cross-resistance development. Together, these results indicate that hydroxychalcones can be considered a good option for development of a new alternative drug for trichomoniasis treatment in association with metronidazole.
References
Abdellatif KRA, Elshemy HAH, Salama SA, Omar HA (2015) Synthesis, characterization and biological evaluation of novel 4′-fluoro-2′-hydroxy-chalcone derivatives as antioxidant, anti-inflammatory and analgesic agents. J Enzyme Inhib Med Chem 30:484–491. https://doi.org/10.3109/14756366.2014.949255
Anthwal A, Rajesh UC, Rawat MSM, Kushwaha B, Maikhuri JP, Sharma VL, Gupta G, Rawat DS (2014) Novel metronidazole-chalcone conjugates with potential to counter drug resistance in Trichomonas vaginalis. Eur J Med Chem 79:89–94. https://doi.org/10.1016/j.ejmech.2014.03.076
Badavath VN, Jadav SS, Pastorino B, de Lamballerie X, Jayaprakash BNS, (2016) Synthesis and antiviral activity of 2-aryl-4H-chromen-4-one derivatives against Chikungunya virus. Lett Drug Des Discov. https://doi.org/10.2174/1570180813666160711163349
Bala V, Chhonker YS (2018) Recent developments in anti-Trichomonas research: an update review. Eur J Med Chem 143:232–243. https://doi.org/10.1016/j.ejmech.2017.11.029
Borsari C, Santarem N, Torrado J, Olías AI, Corral MJ, Baptista C, Gul S, Wolf M, Kuzikov M, Ellinger B, Witt G, Gribbon P, Reinshagen J, Linciano P, Tait A, Costantino L, Freitas-Junior LH, Moraes CB, Bruno dos Santos P, Alcântara LM, Franco CH, Bertolacini CD, Fontana V, Tejera Nevado P, Clos J, Alunda JM, Cordeiro-da-Silva A, Ferrari S, Costi MP (2017) Methoxylated 2′-hydroxychalcones as antiparasitic hit compounds. Eur J Med Chem 126:1129–1135. https://doi.org/10.1016/j.ejmech.2016.12.017
Bouchemal K, Bories C, Loiseau PM (2017) Crossm of Trichomonas vaginalis infections. Clin Microbiol Rev 30:811–825
Da Silva CC, Pacheco BS, de Freitas SC, Berneira LM, dos Santos MAZ, Pizzuti L, de Pereira CMP (2018a) Hydroxychalcones: synthetic alternatives to enhance oxidative stability of biodiesel. In: Trindade M (ed) Increased biodiesel efficiency: alternatives for production, stabilization, characterization and use of coproduct. Springer International Publishing, Cham, pp 81–110. https://doi.org/10.1007/978-3-319-73552-8_4
Da Silva CC, Pacheco BS, de Freitas SC, Berneira LM, dos Santos MAZ, Pizzuti L, de Pereira CMP (2018b) Hydroxychalcones: synthetic alternatives to enhance oxidative stability of biodiesel. Green Energy Technol:81–110. https://doi.org/10.1007/978-3-319-73552-8_4
Daina A, Michielin O, Zoete V (2017) SwissADME: a free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci Rep 7:42717
de Brum Vieira P, Giordani RB, Macedo AJ, Tasca T (2015) Natural and synthetic compound anti-Trichomonas vaginalis: an update review. Parasitol Res 114:1249–1261. https://doi.org/10.1007/s00436-015-4340-3
Desideri N, Mastromarino P, Conti C (2003) Synthesis and evaluation of antirhinovirus activity of 3-hydroxy and 3-methoxy 2-styrylchromones. Antivir Chem Chemother 14:195–203. https://doi.org/10.1177/095632020301400404
Diamond LS (1957) The establishment of various trichomonads of animals and man in axenic cultures. J Parasitol 43:488–490
FDA-EUA (n.d) Drug approvals and databases. http://www.accessdata.fda.gov/scripts/cder/drugsatfda/index.cfm
Forejtníková H, Lunerová K, Kubínová R, Jankovská D, Marek R, Kareš R, Suchý V, Vondráček J, Machala M (2005) Chemoprotective and toxic potentials of synthetic and natural chalcones and dihydrochalcones in vitro. Toxicology 208:81–93. https://doi.org/10.1016/j.tox.2004.11.011
Frank LA, Contri RV, Beck RCR, Pohlmann AR, Guterres SS (2015) Improving drug biological effects by encapsulation into polymeric nanocapsules. Wiley Interdiscip Rev Nanomed Nanobiotechnol. https://doi.org/10.1002/wnan.1334
Gajdács M (2019) The concept of an ideal antibiotic: implications for drug design. Molecules 24(5):892. https://doi.org/10.3390/molecules24050892
Gomes MN, Muratov EN, Pereira M, Peixoto JC, Rosseto LP, Cravo PVL, Andrade CH, Neves BJ (2017) Chalcone derivatives: promising starting points for drug design. Molecules 22. https://doi.org/10.3390/molecules22081210
Hanwell MD, Curtis DE, Lonie DC, Vandermeersch T, Zurek E, Hutchison GR (2012) Avogadro: an advanced semantic chemical editor, visualization, and analysis platform. J Cheminform 4:17. https://doi.org/10.1186/1758-2946-4-17
Hübner DPG, de Brum Vieira P, Frasson AP, Menezes CB, Senger FR, Santos da Silva GN, Baggio Gnoatto SC, Tasca T (2016) Anti-Trichomonas vaginalis activity of betulinic acid derivatives. Biomed Pharmacother 84:476–484. https://doi.org/10.1016/j.biopha.2016.09.064
Jhala YS, Dulawat SS, Verma BL (2006) Solvent-free improved syntheses of some substituted 1 , 3-diaryl-propenones and 3 , 5-diaryl-6-carbethoxycyclohexenones under microwave irradiation and their antibacterial activity. Indian J Chem 45:466–469
Kamboj RC, Arora R, Sharma G, Kumar D, Sharma C, Joshi R, Aneja KR (2010) Eco-friendly synthesis and antimicrobial activity of chalcones. Der Pharma Chem. 2:157–170. https://doi.org/10.1186/2191-2858-2-20
Kissinger P (2015) Trichomonas vaginalis: a review of epidemiologic, clinical and treatment issues. BMC Infect Dis 15:307. https://doi.org/10.1186/s12879-015-1055-0
Lee YT, Fong TH, Chen HM, Chang CY, Wang YH, Chern CY, Chen YH (2014) Toxicity assessments of chalcone and some synthetic chalcone analogues in a zebrafish model. Molecules 19:641–650. https://doi.org/10.3390/molecules19010641
Leitsch D (2016) Recent advances in the Trichomonas vaginalis field. F1000Res 5:1–7. https://doi.org/10.12688/f1000research.7594.1
Ling F, Jiang C, Liu G, Li M, Wang G (2015) Anthelmintic efficacy of cinnamaldehyde and cinnamic acid from cortex cinnamon essential oil against Dactylogyrus intermedius. Parasitology 142:1744–1750. https://doi.org/10.1017/S0031182015001031
Lipinski CA (2004) Lead-and drug-like compounds: the rule-of-five revolution. Drug Discovery Today: Technologies 1(4):337–341. https://doi.org/10.1016/j.ddtec.2004.11.007
Lockwood BC, Coombs GH (1991) Purification and characterization of methionine y-lyase from. Biochem. J. 279:675–682. https://doi.org/10.1042/bj2790675
Mahapatra DK, Bharti SK, Asati V (2015) Anti-cancer chalcones: structural and molecular target perspectives. Eur J Med Chem 98:69–114. https://doi.org/10.1016/j.ejmech.2015.05.004
Morris GM, Huey R, Lindstrom W, Sanner MF, Belew RK, Goodsell DS, Olson AJ (2009) AutoDock4 and AutoDockTools4 : automated docking with selective receptor flexibility. J Comput Chem 30:2785-2791 https://doi.org/10.1002/jcc.21256
Palanco AC, Lacorte S, De J, Costa-Orlandi CB, Gullo FP, Strohmayer Lourencetti NM, Gomes PC, Ayusso GM, Dutra LA, Silva B, Da V, Regasini LO, Soares Mendes-Giannini MJ, Fusco-Almeida AM (2017) Activity of 3′-hydroxychalcone against Cryptococcus gattii and toxicity, and efficacy in alternative animal models. Future Microbiol 12:1123–1134. https://doi.org/10.2217/fmb-2017-0062
Paulish-miller TE, Augostini P, Schuyler JA, Smith WL, Mordechai E, Adelson ME, Gygax SE, Secor WE, Hilbert W (2014) Trichomonas vaginalis metronidazole resistance is associated with single nucleotide polymorphisms in the Nitroreductase genes ntr4 Tv and ntr6 Tv. Antimicrob Agents Chemother 58:2938–2943. https://doi.org/10.1128/AAC.02370-13
Petrin D, Delgaty K, Bhatt R, Garber G (1998) Clinical and microbiological aspects of Trichomonas vaginalis. Clin Microbiol Rev 11:300–317
Pettersen EF, Goddard TD, Huang CC, Couch GS, Greenblatt DM, Meng EC, Ferrin TE (2004) UCSF Chimera - a visualization system for exploratory research and analysis. J Comput Chem 25:1605–1612. https://doi.org/10.1002/jcc.20084
Rezk BM, Haenen GRMM, Van der Vijgh WJF, Bast A (2002) The antioxidant activity of phloretin: the disclosure of a new antioxidant pharmacophore in flavonoids. Biochem Biophys Res Commun 295:9–13. https://doi.org/10.1016/S0006-291X(02)00618-6
Rinaldo-Matthis A, Wing C, Ghanem M, Deng H, Wu P, Gupta A, Tyler PC, Evans GB, Furneaux RH, Almo SC, Wang CC, Schramm VL (2007) Inhibition and structure of Trichomonas vaginalis purine nucleoside phosphorylase with picomolar transition state analogues. Biochemistry 46:659–668. https://doi.org/10.1021/bi061515r
Sena-Lopes Â, das Neves RN, Bezerra FSB, de Oliveira Silva MT, Nobre PC, Perin G, Alves D, Savegnago L, Begnini KR, Seixas FK, Collares T, Borsuk S (2017) Antiparasitic activity of 1,3-dioxolanes containing tellurium in Trichomonas vaginalis. Biomed Pharmacother 89:284–287. https://doi.org/10.1016/j.biopha.2017.01.173
Setzer MS, Byler KG, Ogungbe IV, Setzer WN (2017) Natural products as new treatment options for Trichomoniasis : a molecular docking investigation. https://doi.org/10.3390/scipharm85010005
Singh G, Arora A, Mangat SS, Rani S, Kaur H, Goyal K, Sehgal R, Maurya IK, Tewari R, Choquesillo-Lazarte D, Sahoo S, Kaur N (2016) Design, synthesis and biological evaluation of chalconyl blended triazole allied organosilatranes as giardicidal and trichomonacidal agents. Eur J Med Chem 108:287–300. https://doi.org/10.1016/j.ejmech.2015.11.029
Steindel PA, Chen EH, Wirth JD, Theobald DL (2016) Gradual neofunctionalization in the convergent evolution of trichomonad lactate and malate dehydrogenases. Protein Sci 25:1319–1331. https://doi.org/10.1002/pro.2904
Tallarida RJ (2011) Quantitative methods for assessing drug synergism. Genes and Cancer 2:1003–1008. https://doi.org/10.1177/1947601912440575
Trein MR, Rodrigues L, Rigo GV, Aparecida M, Garcia R (2019) Anti-Trichomonas vaginalis activity of chalcone and amino-analogues. Parasitol Res 118:607-615. https://doi.org/10.1007/s00436-018-6164-4
Trott O, Olson AJ (2009) AutoDock Vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J Comput Chem. https://doi.org/10.1002/jcc.21334
Wang F-W, Wang S-Q, Zhao B-X, Miao J-Y (2014) Discovery of 2′-hydroxychalcones as autophagy inducer in A549 lung cancer cells. Org Biomol Chem 12:3062–3070. https://doi.org/10.1039/C3OB42429D
WHO (2012) Global incidence and prevalence of selected curable sexually transmitted infections-2008. World Heal Organ., pp 1–28
Wong E (1968) The role of chalcones and flavanones in flavonoid biosynthesis. Phytochemistry 7:1751–1758
Zhou B (2015) Diverse molecular targets for chalcones with varied bioactivities. Med Chem (Los Angeles) 5:388–404. https://doi.org/10.4172/2161-0444.1000291
Acknowledgments
This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES) - Finance Code 001.
Author information
Authors and Affiliations
Corresponding author
Additional information
Section Editor: Kevin S.W. Tan
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
das Neves, R.N., Sena-Lopes, Â., Alves, M.S.D. et al. 2′-Hydroxychalcones as an alternative treatment for trichomoniasis in association with metronidazole. Parasitol Res 119, 725–736 (2020). https://doi.org/10.1007/s00436-019-06568-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-019-06568-4