
Abstract
Visceral leishmaniasis (VL) caused by Leishmania donovani is a life-threatening disease involving uncontrolled parasitization of vital organs. Drugs to treat leishmaniasis have one or more limitations or insufficiencies in the long run. A safe and efficacious vaccine to control this disease is needed. Killed antigens that could be safer as vaccines have shown limited efficacy in clinical trials. Immunogenic enhancement with appropriate adjuvants may thus be required to elicit protective immunity based on antibodies and effector T-cell functions. Therefore, it is essential to search for adjuvant to enhance the immunogenicity of killed vaccines and to induce protection against leishmaniasis. So, the aim of the present study was to compare the effectiveness of four adjuvants, i.e. alum, saponin, monophosphoryl lipid A, cationic liposome in combination with Killed Leishmania donovani (KLD) antigen against murine VL. Animals were immunized subcutaneously thrice at an interval of 2 weeks with a final volume of 100 μl per dose. Challenge infection was given 2 weeks after last booster. Mice were sacrificed 15 days after last immunization and on 30, 60 and 90 post-infection/challenge days. The protective efficacy of vaccines was revealed by significant reduction in parasite burden and enhanced DTH responses in comparison with the infected controls. Immunized animals also generated significant levels of Th1 cytokines and increased production of IgG2a, thus indicating the generation of a protective Th1 response. All the adjuvants imparted significant protection, but liposomal formulation was most effective followed by KLD + MPL-A, KLD + saponin, KLD + alum and KLD antigen.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Visceral leishmaniasis (VL) is a vector-borne parasitic disease, which results from systemic infection with Leishmania donovani and L. infantum in the Old World and L. chagasi in the New World. L. chagasi clearly belongs to the L. donovani complex but it has been synonymized with L. infantum on the basis of RAPD and DNA sequence analysis, which suggests the introduction of L. chagasi in New World [1]. Parasites of the genus Leishmania target preferentially macrophages throughout the viscera and are usually found in spleen, liver and bone marrow [2]. Current treatment relies on handful of drugs. It is unlikely that chemotherapy alone will enable disease eradication, and hence, there is an urgency to develop an effective vaccine. The acquisition of immunity following Leishmania infection gives a hope that prevention of leishmaniasis through prophylactic vaccination is feasible.
Killed antigens composed of parasite fractions or whole killed parasites reached phase 3 clinical trials, but showed a limited prophylactic efficacy [3]. Most of the prophylactic immunizations in past have been carried out with autoclaved L. major promastigotes. Vaccination with alum-absorbed preparation (alum-ALM) along with BCG has been shown to be more immunogenic, protective and produced a strong and long-lasting Th1 response in mice, monkeys and dogs against VL [4–6] but it did not produce conclusive results in humans [7–9]. In spite of the availability of numerous candidate antigens against leishmaniasis, limited studies are available about the use of formalin-killed promastigotes. Recently, formalin-killed promastigotes of L. major, along with Montanide ISA 720 (MISA) adjuvant, has been reported to be an effective antigen candidate against murine cutaneous leishmaniasis [10].
It seems that major limiting factor for the development of killed vaccines is lack of a suitable adjuvant [11, 12]. The accessibility to hundreds of adjuvants has encouraged a need for identifying rational standards for selection of adjuvant formulation based on safety and sound immunological principles for human vaccines. Aluminium salt-based adjuvants, referred generically as ‘alum’, are among few of the licensed adjuvants approved for human use [13]. In relation to leishmaniasis, alum has been used in combination with ALM + BCG for immunization of patients with VL [14] and with ALM + SSG + BCG in PKDL patients [15]. Monophosphoryl Lipid A (MPL-A) is a nontoxic derivative of lipopolysaccharide of Salmonella minnesota, a gram-negative bacteria. It activates innate immunity through toll-like receptor (TLR4), enhances phagocytosis, upregulates major histocompatibility complex (MHC) expression and induces a cascade of cytokines [9, 16]. MPL is approved for human use and currently used in several human vaccines including malaria, Epstein-Barr virus, herpes simplex, hepatitis and leishmaniasis [9, 17, 18]. Saponins are chemically a heterogeneous group of sterol glycosides and triterpene glycosides. Saponin-based adjuvants have the ability to modulate the cell-mediated immune system as well as to enhance antibody production and have the advantage that only a low dose is needed for adjuvant activity [19]. QS21 fraction of saponin in combination with FML (Leishmune®) has been registered in Brazil for immunization in dogs and induced significant protection in various trials [20–22]. Liposomes are versatile and robust delivery systems known for induction of humoral and cell-mediated immune responses. The potential of liposomes to assimilate broad range of antigens (diphtheria, hepatitis A, influenza, tetanus toxoid and leishmanial proteins) exhibits the versatility of these vesicles as an antigen delivery system [23].
The encouraging findings of above studies prompted us to investigate the immunoprophylactic potential of formalin-fixed Killed Leishmania donovani (KLD) antigen along with different adjuvants against murine VL.
Materials and methods
Parasite culture and preparation of different vaccine formulations
Promastigotes of L. donovani strain MHOM/IN/80/Dd8 were used for the present study and maintained in modified Novy, McNeal and Nicolle’s (NNN) medium by serial subcultures in Minimum Essential Medium (MEM) after every 48–72 h. For the preparation of vaccination antigen, stationary-phase promastigotes were harvested, counted in Neubauer’s chamber and then resuspended in 1 ml sterile PBS (pH 7.2) at a concentration of 108 parasites. Promastigotes were killed by suspending in 0.1 % formalin overnight, and then, they were washed thrice with PBS. About 0.1 ml of this suspension containing 107 parasites was used for immunization of mice. Each animal was immunized with KLD antigen alone or mixed with respective dose of adjuvant in 100 µl of PBS. For each animal, adjuvant dosage was 100 µg saponin and 40 µg of MPL-A. For preparation of KLD + alum vaccine, KLD antigen was dissolved in 4 ml of PBS. It was then added to 1 ml of 10 % potash alum (aluminium potassium sulphate). After that, 10 % Na2Co3 solution (0.3–0.5 ml) was added gradually to bring the pH of solution between 5.8 and 6.8. The precipitates so formed were shaken well and then injected into mice. For preparation of positively charged liposomes, commercially available kit from Sigma-Aldrich (USA) was used. About 63 µmol of phosphatidylcholine, 9 µmol of cholesterol and 18 µmol of stearylamine in the ratio of (7:1:2) were used for the preparation of positively charged liposomes. Encapsulation of antigen in liposomes was carried out by the method described by Afrin and Ali [24]. The protein content entrapped in the liposome was estimated by the method described by Lowry et al. [25], with bovine serum albumin as the standard, in the presence of 0.8 % sodium dodecyl sulphate and appropriate blanks. The protein content was found to be 46.6 µg.
Chemicals and reagents
IgG1 and IgG2a (Serotec), MEM (Hi-Media), alum, saponin, liposomes, monophosphoryl lipid A, Primers (Sigma-Aldrich, USA), Maxima SYBR Green qPCR Master Mix (Thermo Scientific), Cytokine ELISA kits (Diaclone, France), 96-well ELISA plates (Tarson).
Immunization and challenge infection of experimental animal groups
Female BALB/c mice, weighing 20–25 g, were acquired from the Institute of Microbial Technology, Chandigarh, India, and then maintained in the central animal house of Panjab University, Chandigarh. They were housed in clean cages and fed with water and mouse feed ad libitum. Experimental groups consisted of 4–6-week-old inbred BALB/c mice. Three immunizations were carried out at an interval of 2 weeks. Animals immunized with PBS only and challenged with 107 promastigotes of L. donovani served as infected controls. Normal control mice received only PBS. Two weeks after last booster dose, mice of control and immunized groups were challenged with 1 × 107 promastigotes. Various parasitological and immunological studies were carried out 15 days after last immunization and on 30, 60 and 90 post-infection/challenge days (p.c.d.)
Ethical clearance
Experiments were carried out according to the guidelines of the Committee for the purpose of Control and Supervision of Experiments on Animals (CPCSEA, Registration No. 45/1999/CPCSEA). The ethical clearance for conducting the experiments was obtained from the Institutional Animal Ethics Committee (IAEC), Panjab University, Chandigarh, India (Approval No. IAEC 284-295/3.09.2012). Mice were handled according to their guidelines.
Assessment of parasite burden
Six animals from each group were killed on different post-challenge days. Liver and spleen of all animals were removed and weighed. Impression smears were made on clean glass slides. The smears were air dried, fixed in methanol and stained with Giemsa. The parasite load was assessed in terms of Leishman Donovan Units (LDU) by the method prescribed by Bradley and Kirkley [26] and calculated as follows:
For real-time PCR analysis, DNA was isolated from liver and spleen (100 mg) using commercially available Genomic DNA mini spin kit (Cat# RKN 32, Chromous Biotech) according to manufacturer’s instructions. Eluted DNA was quantified, analysed and stored at −20 °C till further use. A real-time PCR was performed using Maxima SYBR Green Master Mix to amplify kinetoplast DNA of L. donovani using specific primers (Table 1). Primers were designed using Primer 3 input software. The 10-µl reaction contained 50 ng of genomic DNA (2 µl), 5 µl of Maxima SYBR Green qPCR Mastermix, 0.12 µl of each primer and 2.76 µl of nuclease-free water. Negative control tubes that received 2 µl of water instead of DNA extract were included in each PCR run to detect any amplicon contamination. Times and temperatures of cycling programme are shown in Table 2. The efficacy of vaccine formulation was determined in terms of fold change decrease in parasitic load as compared to infected controls.
Delayed-type hypersensitivity (DTH) responses to leishmanin
DTH response is a classic hallmark of cell-mediated immunity. The induction of such measurable immunological parameter may have important predictive value with regard to resistance to infection. For evaluation of DTH responses, 48 h before the day of killing, mice were challenged in the right foot pad with a subcutaneous injection of leishmanin (prepared in PBS) and left foot pad with PBS (leishmanin was prepared by harvesting stationary-phase promastigotes from modified NNN medium and washed thrice with PBS. The final pellet was then suspended in 5 ml PBS containing 0.5 % phenol and kept at room temperature for 10 min. The phenol was then removed by washings with PBS, and the final concentration was adjusted to 2 × 108 promastigotes/ml). After 48 h, the thickness of right and left foot pad was measured using a pair of vernier callipers. Results were expressed as mean ± SD of percentage increase in the thickness of the right footpad as compared to the left footpad of mice [27].
ELISA for parasite-specific IgG1 and IgG2a isotypes
Conventional ELISA was performed to determine the levels of serum immunoglobulin G (IgG) isotype antibodies (IgG1 and IgG2a) by the method described by Ravindran et al. [28]. Briefly, 96-well plates were coated with crude antigen (5 μg/well) and incubated overnight at 4 °C. Serum samples were added at twofold serial dilutions. It was followed by addition of isotype-specific HRP-conjugated secondary antibodies (rabbit anti-mouse IgG1 or IgG2a) after which the H2O2 substrate and DAB chromogen were added. Absorbance was read on an ELISA plate reader (Lisa Plus, India) at 450 nm.
Cytokine assays
Cytokines play an important role in shaping the host immune response to Leishmania infection and leading the progression towards protective or non-protective immunity during infection. For the evaluation of cytokine levels, cell suspensions from spleen of normal controls, infected controls and immunized mice were prepared and cultured in 96-well plates in RPMI-1640 containing 20 mM NaHCO3, 10 mM HEPES, 10 U/ml of penicillin, 100 µg/ml streptomycin and 2 mM l-glutamine and 10 % FCS. Cells were stimulated with 50 μg/ml of crude antigen of L. donovani [29]. Cells were incubated at 37 °C in a humified chamber containing 5 % CO2 for 72 h and supernatants were collected and stored at −20 °C [30]. This supernatant was assayed for the concentration of IL-12, IL-4, IL-10 and IFN-γ by commercially available cytokine ELISA kits from Diaclone, France according to the manufacturer’s instructions.
Statistical analysis
The data comparisons were tested for significance using Sigma plot (12.5) software. All the data were analysed using two-way analysis of variance (ANOVA). Post hoc test was used for multiple comparisons using Holm–Sidak method. p values of <0.05 and <0.001 were considered significant. Results were expressed as mean ± SD of one of three independent experiments.
Results
Assessment of parasite burden
Quantification of parasite load was done for all groups on 30, 60 and 90 post-challenge days. Parasite load was calculated by studying giemsa-stained impression smears of liver and spleen. As compared to infected controls, we observed a trend of decreased parasite burden in immunized groups (p < 0.001). The combination of liposome-encapsulated KLD antigen imparted maximum protection with 86–93.7 % reduction in LDU from 30 to 90 post-challenge day. Groups immunized with KLD + MPL-A and KLD + saponin also conferred good protection with 83.4–92.8 % and 79.9–88.8 % parasite reduction, respectively. Immunization with KLD + alum resulted in 73.78–82.9 % reduction in parasite load. Between the immunized groups, the difference in LDU was found to be significant (p < 0.05, p < 0.001) on all post-challenge days (Fig. 1a). Furthermore, immunization with KLD antigen also significantly (p < 0.001) reduced the parasite burden (66.3–71.7 %, respectively) as compared to infected controls. In VL, the spleen acts as a reservoir for parasitic persistence, which is further associated with failure to clear the disease. We therefore wanted to observe the effect of immunization with different vaccine formulations on the splenic persistence of L. donovani following challenge infection. The splenic parasite burden was also found to be significantly reduced in all the immunized groups as compared to infected controls (Fig. 1b).

Estimation of hepatic and splenic (a, b) parasite load in all groups of BALB/c mice. The data are presented as mean ± SD of six mice per group. α* p value: infected controls versus KLD; KLD + alum; KLD + saponin; KLD + MPL-A; liposome-encapsulated KLD. !^ p value: KLD versus KLD + alum; KLD + saponin; KLD + MPL-A; liposome-encapsulated KLD. *^ p value <0.001, α! p value <0.05, NS nonsignificant
To confirm the accuracy of parasite load calculated by giemsa-stained slides, we also did qPCR analysis at 90th post-challenge day. Real-time PCR analysis also displayed similar trend of decrease in parasite burden for all groups of immunized animals (Fig. 2a). Maximum protection was achieved with the use of liposome-encapsulated KLD antigen with 13.69-fold decrease in hepatic parasite burden as compared to infected controls. This group was followed by group immunized with KLD + MPL-A (12.82), KLD + saponin (7.12), KLD + alum (5.5) and KLD alone (3.89). Similar trend of decrease in parasite burden was also seen in spleen. Since spleen is a site of persistent infection, so parasite number decreased slowly as compared to liver. However, the parasite load declined significantly when compared with infected controls at 90th post-challenge day (Fig. 2b).

Estimation of fold change decrease in hepatic and splenic (a, b) parasite burden in all groups of BALB/c mice. The data are presented as mean ± SD of six mice per group. α* p value: infected controls versus KLD; KLD + alum; KLD + saponin; KLD + MPL-A; liposome-encapsulated KLD. !^ p value: KLD versus KLD + alum; KLD + saponin; KLD + MPL-A; liposome-encapsulated KLD. *^ p value <0.001, α! p value <0.05, NS nonsignificant
Assessment of DTH responses
To evaluate cell-mediated immune responses, we monitored the delayed-type hypersensitivity (DTH) responses in mice after 15 days post-immunization and at 30, 60, 90 post-infection/challenge days. On day 15 post-immunization, all the animals displayed elevated DTH responses (p < 0.001) in comparison with the control mice. DTH responses were further found to increase from 30 to 90 post-infection/challenge days. A profound DTH response was induced by immunization of mice with KLD alone. Adjuvants used along with KLD antigen for immunization further heightened the DTH responses. Peak DTH response was observed in group of animals immunized with liposome-encapsulated KLD and KLD + MPL-A suggesting the generation of protective cell-mediated immunity. A significant increase (p < 0.001) in footpad thickness was observed in immunized mice as compared to infected control mice on all post-challenge days (Fig. 3).

DTH responses (a, b) in BALB/c mice following immunization and L. donovani challenge infection. The data are presented as mean ± SD of six mice per group. α* p value: Control mice versus KLD; KLD + alum; KLD + saponin; KLD + MPL-A; liposome-encapsulated KLD. !^ p value: KLD versus KLD + alum; KLD + saponin; KLD + MPL-A; liposome-encapsulated KLD. *^ p value <0.001, α! p value <0.05, NS nonsignificant
Humoral immune responses
Serum samples of immunized and control mice were analysed for detection of IgG1 and IgG2a antibodies by ELISA. The relative production of these isotypes can thus be used as a marker for the induction of Th1-like and Th2-like immune responses. At 15th day post-vaccination, all the immunized groups displayed significantly higher levels of parasite-specific IgG isotypes, i.e. IgG1 and IgG2a in comparison with control mice (p < 0.001) that persisted even after challenge infection. Least IgG1 levels were observed in the animals immunized with liposome-encapsulated KLD antigen. IgG1 levels in immunized mice were lesser as compared to the infected control animals. The difference was significant (p < 0.001) on all post-challenge days. IgG2a antibody levels indicate the generation of Th1 type of immune response, which is protective. Immunization of mice with KLD antigen alone significantly raised the level of IgG2a as compared to infected controls (p < 0.001). Inclusion of adjuvants to KLD antigen further enhanced the IgG2a production (p < 0.001). Peak absorbance values were observed in animals immunized with a combination of KLD + MPL-A followed by groups immunized with combination of liposome-encapsulated KLD, KLD + saponin and KLD + alum with IgG2a/IgG1 ratio 1.91, 1.9, 1.48 and 1.36, respectively (Fig. 4). All immunized groups maintained a trend of high IgG2a/IgG1 ratio till day 90 post-challenge, thus reflecting stimulation of a Th1-biased immune response.

Antibody level (a, b—IgG1, c, d—IgG2a) in serum samples of BALB/c mice following immunization and L. donovani challenge infection. The data are presented as mean ± SD of six mice per group. α* p value: Control mice versus KLD; KLD + alum; KLD + saponin; KLD + MPL-A; liposome-encapsulated KLD. !^ p value: KLD versus KLD + alum; KLD + saponin; KLD + MPL-A; liposome-encapsulated KLD. *^ p value <0.001, α! p value <0.05, NS nonsignificant
Determination of vaccine-induced cytokine production
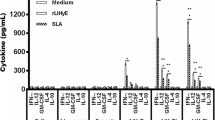
To get a further insight into the protective efficacy of different vaccine formulations, we cultured splenocytes on 15 day post-immunization as well as on 30, 60, 90 post-infection/challenge days. Immune responses (Th1 or Th2) induced by different vaccine formulations were assessed by quantifying the cytokine levels (IFN-γ, IL-12, IL-4 and IL-10) in splenocyte cultures of all groups of animals. On 15th day immunization, we found that splenocytes from immunized groups secreted significantly higher cytokines as compared to normal control mice. It was interesting to note that level of Th1 cytokines in immunized groups was high as compared to Th2 cytokines.
We also wanted to assess the cytokine production after infection, and we found that immunized animals maintained high levels of Th1 cytokines over infected controls. Mice immunized with liposome-encapsulated KLD antigen elicited highest concentration of Th1 specific cytokines, i.e. IFN-γ and IL-12 (Fig. 5) followed by those immunized with a combination of KLD + MPL-A, KLD + saponin and KLD + alum. Among immunized groups, lowest levels of Th1 cytokines were found in mice immunized with KLD alone. In sum, we could say that high IFN-γ and IL-12 levels correlated with protective immunity.

IFN-γ and IL-12 concentration (a, b—IFN-γ, c, d—IL12) in BALB/c mice following immunization and L. donovani challenge infection. The data are presented as mean ± SD of six mice per group. α* p value: Control mice versus KLD; KLD + alum; KLD + saponin; KLD + MPL-A; liposome-encapsulated KLD. !^ p value: KLD versus KLD + alum; KLD + saponin; KLD + MPL-A; liposome-encapsulated KLD. *^ p value <0.001, α! p value <0.05, NS nonsignificant
On the contrary, maximum levels of Th2-regulated cytokines, i.e. IL-10 and IL-4 (Fig. 6), were observed in the infected controls. Levels of IL-10 (p < 0·05) and IL-4 (p < 0·001) were significantly lesser in immunized animals as compared to the infected controls except for mice immunized with KLD antigen where the difference was insignificant on 90th p.c.d (IL-10). Among immunized groups, peak levels of IL-4 and IL-10 were seen in groups of mice immunized with KLD antigen alone. Addition of adjuvants significantly downregulated IL-4 and IL-10 levels except for KLD + alum where the difference was insignificant on all post-infection/challenge days (p.c.d). Minimum level of Th2 cytokines was observed in mice immunized with liposome-encapsulated KLD antigen.

IL-10 and IL-4 concentration (a, b—IL10, c, d—IL4) in BALB/c mice following immunization and L. donovani challenge infection. The data are presented as mean ± SD of 6 mice per group. α* p value: Control mice versus KLD; KLD + alum; KLD + saponin; KLD + MPL-A; liposome-encapsulated KLD. !^ p value: KLD versus KLD + alum; KLD + saponin; KLD + MPL-A; liposome-encapsulated KLD. *^ p value <0.001, α! p value <0.05, NS nonsignificant
We also checked IFN-γ/IL-10 ratio and found that highest IFN-γ/IL-10 ratio was found in liposome-encapsulated KLD antigen (4.61) followed by group immunized with KLD + MPL-A, KLD + saponin, KLD + alum and KLD alone with IFN- γ/IL-10 ratio 3.74, 2.57, 1.9, 1.86 respectively. The ratio implicated that a strong Th1 bias may be an important correlate of protection observed within these groups. Lowest IFN-γ/IL-10 (0.81) ratio was observed in infected controls suggesting bias towards nonprotective Th2 immune response (Fig. 7).

IFN-γ and IL-10 ratio in BALB/c mice following immunization and L. donovani challenge infection
Discussion
Vaccination seems to be the most cost-effective way of controlling infectious diseases. Therefore, the study and designing of safe and effective vaccine regimens for clinical use remains critical. Killed vaccines offer the advantage of being safe and have been tested in clinical trials but they lack immunogenicity. Development of an effective vaccine depends not only upon selection of appropriate vaccine candidates but also on choosing the right adjuvant or delivery vehicle. Thus, a successful anti-leishmanial vaccine will need to be assessed with human-compatible adjuvants. So the main goal of the present study was to evaluate the immunoprophylactic efficacy of different adjuvants in combination with KLD antigen. Our findings clearly indicate that all the adjuvants used in the study exerted adjuvant effects against L. donovani challenge infection. However, the extent of protection varied according to the type of adjuvant used.
Quantification of parasite load was done in all the groups of mice on different post-infection/challenge days. L. donovani infection is described by distinct organ-specific immune interactions, where liver is the site of infectious resolution. Since infection is self-limiting in BALB/c mice, maximum hepatic parasite burden was seen on 30 days post-infection/challenge in all groups of mice. After that, the parasite load declined significantly in all groups of animals. Among immunized groups, we found that mice immunized with liposome-encapsulated KLD antigen showed maximum reduction in parasite load (93.7 %). Our results are in agreement with the study of Shargh et al. [31] where cationic liposomes containing soluble Leishmania antigen + CpG ODNs showed significantly lower parasite burden. The antigenicity of KLD antigen in combination with MPL-A was also studied, and it was found that mice immunized with KLD + MPL-A showed a significant reduction in the parasite load. Results are in accordance with study of Nagill and Kaur [32] where 78 kDa antigen along with an adjuvant (MPL-A) induced 92 % protection against L. donovani infection. Several studies have demonstrated that this adjuvant either directly or indirectly stimulates the production of the T helper cell type 1 (Th1) cytokines IL-2 and IFN-γ [33], which plays a major role in elimination of parasites. Mice immunized with KLD + saponin also resulted in reduction in parasite burden. Saponin as an adjuvant can affect the generation of immune response at several levels. These may include mobilization of antigen-presenting cells to the site of injection, enhancing efficient processing of antigen, prompting the cytokine response including IFN-γ and other co-stimulatory signals needed for an optimal immune response [34, 35]. These studies underline the immunostimulatory potential of saponin that may have led to the enhanced level of protection found in our study. So, there is much hope for a saponin-adjuvanted killed leishmanial vaccine in veterinary and clinical research. Significant level of protection was also conferred by mice immunized with KLD + alum. This is in agreement with another study which showed that vaccination with alum-precipitated autoclaved L. major + BCG conferred significant protection in mice [36].
Successful vaccination of animals and humans is often related to antigen-induced DTH responses in vivo and T-cell stimulation with antigen in vitro [37, 38]. In the present study, results obtained upon vaccination with killed leishmanial antigen in association with different adjuvants demonstrated induction of an appreciable DTH response suggesting the activation of cell-mediated immunity. This measure of cellular immunity did correlate well with enhanced resistance to infection in immunized groups. In our study, immunization of mice with KLD antigen alone significantly raised the DTH responses in comparison with the infected controls. The addition of adjuvants further increased the DTH responses significantly. Since the infection is associated with depressed cell-mediated immune responses, lowest DTH responses were observed in infected controls. The significantly higher DTH response induced by liposome-encapsulated KLD antigen over other adjuvant before and after challenge infection demonstrates elicitation of strong and persistent cell-mediated immunity by this vaccine. Our results are in agreement with the studies, which showed that mice immunized with liposome-encapsulated gp63 [39] induced good DTH response. Further results are in line with a previous study in which mice immunized with recombinant Leishmania antigen plus MPL showed significant elevation of DTH response against L. donovani infection [40].
Since IgG1 and IgG2 responses are strictly T cell dependent, we used them as marker to assess the overall immunogenicity of different vaccine formulations [41]. IgG1 and IgG2 subtypes have been traditionally used as surrogate markers of the Th2 and Th1 type of immune responses, respectively. Serological data showed that mice immunized with different adjuvants in combination with KLD antigen induced strong humoral responses after immunization that persisted after challenge infection. All immunized groups showed almost similar level of IgG1 antibodies. Higher levels of IgG2a antibody were detected in serum samples of immunized animals again pointing towards the control of the infection in these animals. Maximum levels of IgG2a were seen in serum samples of mice immunized with MPLA vaccine followed by liposome-encapsulated KLD antigen, KLD + saponin and KLD + alum. High antibody titres generated by KLD + alum confirm that this adjuvant is a potent inducer of antibodies in vaccines [5]. Further our results are in agreement with study of Tafaghodi et al. [42] where vaccination with ALM + QS (autoclaved L. major + Quillaja saponin) showed highest IgG2a/IgG1 ratio against experimental L. major infection.
Recovery and protection against leishmaniasis result from a strong and specific cellular immune response, followed by the development of long-lasting protection [43]. Studies have shown that control of VL infection in mice requires the development of IL-12 dependent IFN-γ production by splenic cells that drives the immune response towards a protective Th1 type [44–47] with low levels of IL-4 [48]. These inflammatory cytokines are required for the generation of reactive oxygen and nitrogen species by infected macrophages that enables killing of intracellular parasites. Similar trend of cytokine production was seen in our study as splenic T cells of immunized animals produced higher amount of IFN-γ and IL-12 with downregulation of IL-4 and IL-10. Highest level of IFN-γ and IL-12 were produced by liposome-encapsulated KLD antigen followed by group immunized with KLD + MPL-A. These findings may suggest that cationic liposomes and MPL-A are potent inducers of Th1 cytokines and thus are suitable for vaccines intended to use against intracellular pathogens. Moderate levels of IFN-γ and IL-12 were seen in mice immunized with KLD + saponin and KLD + alum. It is in agrement with a study which showed that alum-precipitated proteins can also induce CD8+ T cells to produce Th1-associated IFN-γ [49].
Recent advances have also been made in understanding immunoregulatory mechanisms that suppress parasite-specific CD4+ T-cell responses in human VL patients, and it was found that interleukin-10 produced by CD4+ T cells is a potential, autocrine inhibitor of IFN-γ production and promotes parasite persistence in visceral organs [50]. Our results are in accordance with this study as infected control mice displayed highest levels of interleukin-10. The negative regulatory role of IL-10 in vaccine failure is indeed well established [51]. Interestingly, it was found that after immunization IL-4 was produced in the spleen cells of all vaccinated groups. However, level of IL-4 declined significantly on 90th post-challenge day as compared to the infected controls. The evidences that early IL-4 is needed to drive Th1 differentiation [52] and to enhance IFN-γ secretion [53, 54] imply that early IL-4 production does not hinder the Th1 response. It is in line with observations revealing that IL-4 is crucial for the priming of long-term CD8+ T-cell memory responses [55–57].
Moreover, previous studies have shown that SSG chemotherapy of L. donovani in IL-4−/− [58] or IL-13−/− [59] BALB/c mice compared with wild-type mice resulted in downregulated IFN-γ production and increased IL-10 production. This suggests a requirement for IL-4 and IL-13 in instructing a type-1 response. Another study has also shown that successful immunotherapeutic intervention in the BALB/c mouse using a hybrid cell vaccine was associated not only with downregulation of IL-10 but also with significantly enhanced IL-4 and IL-13 expression as well as IFN-γ production [60] providing further circumstantial evidence for these Th2 cytokines in facilitating the generation of type-1 immunity.
Thus, in the present study, it was found that all adjuvanted killed antigen formulations were immunogenic and imparted significant protection. Protective efficacy of cationic liposomes as an adjuvant was superior when compared to alum and saponin and it was comparable to MPL-A. Liposome vaccine technology has advanced in recent years, and now several vaccines containing liposome-based adjuvants have been approved for human use or have reached late stages of clinical evaluation [61]. In contrast to a report [62] preferring intra-peritoneal route for liposome-encapsulated antigen, our liposomal formulation given through subcutaneous route was effective. Subcutaneous route is always considered as a slow release route in case of antigen presentation, allowing the body to generate enough resources to counter the invading microorganisms. Moreover, it is a compatible route for human vaccination. Our findings correlate very well with these assumptions. High levels of Th1 cytokines and IgG2a antibody were seen in group immunized with KLD + MPL-A. It is well known that MPL acts upon TLR-4, promoting the maturation of APCs. Ultimately, activation of TLR-4 not only triggers innate immunity, but also enhances both humoral and cellular immune responses [63, 64]. Recently, Mutiso et al. [65] indicated good safety levels of a vaccine comprising L. donovani sonicate antigen delivered with either monophosphoryl lipid A or montanide ISA 720 in the vervet monkey model. Saponins are natural glycosides of steroid or triterpene, which are known to activate immune system. From our results, we found that vaccine formulation composed of KLD + saponin also showed promising results. We observed that vaccine formulation comprising KLD + alum also induced protection, although it was low when compared with other adjuvants. Therefore, in order to improve efficacy of this vaccine, it is better to use alum with another Th1-promoting adjuvant.
Conclusion
Most studies have focussed on the use of autoclaved vaccine formulations with or without adjuvants against VL, and few studies are available on the use of formalin-killed antigens. The present study highlights the greater efficacy of formalin-fixed KLD antigen as potential vaccine candidate along with promising role of cationic liposome and MPL-A as adjuvants to enhance stronger immune response against murine VL. Considering the easy preparation of killed antigen vaccines, their lower production cost and promising results, this approach might prove to be useful in controlling leishmaniasis in endemic areas, particularly in developing countries. For future perspective, more studies are required in higher animal models in order to ensure the efficacy of above vaccine formulations.
References
Mauricio IL, Howard MK, Stothard JR, Miles MA (1999) Genomic diversity in the Leishmania donovani complex. Parasitology 119:237–246
Kumar R, Engwerda C (2014) Vaccines to prevent leishmaniasis. Clin Transl Immunol 3:e13. doi:10.1038/cti.2014.4
Noazin S, Khamesipour A, Moulton LH, Tanner M, Nasseri K, Modabber F, Sharifi I, Khalil EA, Bernal ID, Antunes CM, Smith PG (2009) Efficacy of killed whole-parasite vaccines in the prevention of leishmaniasis: a meta-analysis. Vaccine 27:4747–4753
Kenney RT, Sacks DL, Sypek JP, Vilela L, Gam AA, Evans-Davis K (1999) Protective immunity using recombinant human IL-12 and alum as adjuvants in a primate model of cutaneous leishmaniasis. J Immunol 163:4481–4488
Misra A, Dube A, Srivastava B, Sharma P, Srivastava JK, Katiyar JC, Naik S (2001) Successful vaccination against Leishmania donovani infection in Indian langur using alum-precipitated autoclaved Leishmania major with BCG. Vaccine 19:3485–3492
Mohebali M, Khamesipour A, Mobedi I, Zarei Z, Hashemi-Fesharki R (2004) Double-blind randomized efficacy field trial of alum precipitated autoclaved Leishmania major vaccine mixed with BCG against canine visceral leishmaniasis in Meshkin-Shahr district, I.R. Iran. Vaccine 22:4097–4100
Kamil AA, Khalil EAG, Musa AM, Modabber F, Mukhtar MM, Ibrahim ME, Zijlstra EE, Sacks D, Smith PG, Zicker F, El Hassan AM (2003) Alum precipitated autoclaved L. major plus BCG, a candidate vaccine for visceral leishmaniasis: safety, skin delayed hypersensitivity response and dose finding in healthy volunteers. Trans R Soc Trop Med Hyg 97:365–368
Khalil EA, Musa AM, Modabber F, El-Hassan AM (2006) Safety and immunogenicity of a candidate vaccine for visceral leishmaniasis (alum-precipitated autoclaved Leishmania major + BCG) in children: an extended phase II study. Ann Trop Paediatr 26:357–361
Ghalib H, Modabber F (2007) Consultation meeting on t development of therapeutic vaccines for post kala azar dermal leishmaniasis. Kinetoplastid Biol Dis 6:7–20. doi:10.1186/1475-9292-6-7
Mutiso JM, Macharia JC, Mutisya RM, Taracha E (2010) Subcutaneous immunization against Leishmania major—infection in mice: efficacy of formalin-killed promastigotes combined with adjuvants. Rev Inst Med Trop Sao Paulo 52:95–100
Khamesipour A, Rafati S, Davoudi N, Maboudi F, Modabber F (2006) Leishmaniasis vaccine candidates for development: a global overview. Indian J Med Res 123:423–438
Keshavarz Valian H, Khoshabe Abdollah Kenedy L, Nateghi Rostami M, Miramin Mohammadi A, Khamesipour A (2008) Role of Mycobacterium vaccae in the protection induced by first generation Leishmania vaccine against murine model of leishmaniasis. Parasitol Res 103:21–28. doi:10.1007/s00436-008-0921-8
Reed SG, Bertholet S, Coler RN, Friede M (2009) New horizons in adjuvants for vaccine development. Trends Immunol 30:23–32
Khalil EA, Musa AM, Modabber F, El-Hassan AM (2006) Safety and immunogenicity of a candidate vaccine for visceral leishmaniasis (alum-precipitated autoclaved Leishmania major + BCG) in children: an extended phase II study. Ann Trop Paediatr 26:357–361
Musa AM, Khalil EA, Mahgoub FA, Elgawi SH, Modabber F, Elkadaru AE et al (2008) Immunochemotherapy of persistent post-kala-azar dermal leishmaniasis: a novel approach to treatment. Trans Roy Soc Trop Med Hyg 102:58–63
Martin M, Michalek SM, Katz J (2003) Role of innate immune factors in the adjuvant activity of monophosphoryl lipid A. Infect Immun 71:2498–2507
Heppner DG, Gordon DM, Gross M, Wellde B, Leitner W, Krzych U et al (1996) Safety immunogenicity and efficacy of Plasmodium falciparum repeatless circumsporozoite protein vaccine encapsulated in liposomes. J Infect Dis 174:361–366
Stoute JA, Kester KE, Krzych U, Wellde BT, Hall T, White K et al (1998) Long-term efficacy and immune responses following immunization with the RTS, S malaria vaccine. J Infect Dis 178:1139–1144
Oda K, Matsuda H, Murakami T, Katayama S, Ohgitani T, Yoshikawa M (2000) Adjuvant and haemolytic activities of 47 saponins derived from medical and food plants. Biol Chem 381:67–74
Santos FN, Borja-Cabrera GP, Miyashiro LM, Grechi J, Reis AB, Moreira MAB, Martins Filho OA, Menz I, Pessoa LM, Goncalves PR, Palatnik M, Palatnik-de-Sousa CB (2007) Immunotherapy against experimental canine visceral leishmaniasis with the saponin enriched-Leishmune® vaccine. Vaccine 25:6176–6190
Borja-Cabrera GP, Santos FN, Bauer FS, Parra LE, Menz I, Morgado A, Soares IS, Batista LM, Palatnik-De-Sousa CB (2008) Immunogenicity assay of the Leishmune® vaccine against canine visceral leishmaniasis in Brazil. Vaccine 26:4991–4997
Araujo MSS, de Andrade RA, Sathler Avelar R, Teixeira-Carvalho A, Andrade MC, Vianna LR, Mayrink W, Reis AB, Malaguias LC, Mello MN, Martins-Filho OA (2009) T-cell-derived cytokines, nitric oxide production by peripheral blood monocytes and seric anti-Leishmania (Leishmania) chagasi IgG subclass patterns following immunization against canine visceral leishmaniasis using Leishvaccine and Leishmune. Vaccine 27:1008–1017
Felnerova D, Viret JF, Gluck R, Moser C (2004) Liposomes and virosomes as delivery systems for antigens, nucleic acids and drugs. Curr Opin Biotechnol 15:518–529
Afrin F, Ali N (1997) Adjuvanticity and protective immunity elicited by Leishmania donovani antigens encapsulated in positively charged liposomes. Infect Immun 65:2371–2377
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Protein measurement with the folin phenol reagent. J Biol Chem 193:265–275
Bradley DJ, Kirkley J (1977) Regulation of Leishmania population within the host. I. The variable course of Leishmania donovani infections in mice. Clin Exp Immunol 30:119–129
Kaur S, Kaur T, Garg N, Mukherjee S, Raina P, Athokpam V (2008) Effect of dose and route of inoculation on the generation of CD4+ Th1/Th2 type of immune response in murine visceral leishmaniasis. Parasitol Res 103:1413–1419
Ravindran R, Anam K, Bairagi BC, Saha B, Pramanik N, Guha SK et al (2004) Characterization of immunoglobulin G and its subclass response to Indian kala-azar infection before and after chemotherapy. Infect Immun 72:863–870
Kaur T, Thakur A, Kaur S (2013) Protective immunity using MPL-A and autoclaved Leishmania donovani as adjuvants along with a cocktail vaccine against murine model of visceral leishmaniasis. J Parasit Dis 37:231–239. doi:10.1007/s12639-012-0171-7
Park AY, Hondowicz BD, Scott P (2000) IL-12 is required to maintain a Th-1 response during Leishmania major infection. J Immunol 165:896–902
Shargh VH, Jaafari MR, Khamesipour A, Jaafari I, Jalali SA, Abbasi A, Badiee A (2012) Cationic liposomes containing soluble Leishmania antigens (SLA) plus CpG ODNs induce protection against murine model of leishmaniasis. Parasitol Res 111:105–114. doi:10.1007/s00436-011-2806-5
Nagill R, Kaur S (2010) Enhanced efficacy and immunogenicity of 78 kDa antigen formulated in various adjuvants against murine visceral leishmaniasis. Vaccine 28:4002–4012
Gustafson GL, Rhodes MJ (1994) Effects of tumor necrosis factor and dexamethasone on the regulation of interferon-gamma induction by monophosphoryl lipid A. J Immunother Emphasis Tumor Immunol 15:129–133
Caro MR, Ortega N, Buendía AJ, Gallego MC, Del Río L, Cuello F, Salinas J (2003) Relationship between the immune response and protection conferred by new designed inactivated vaccines against ovine enzootic abortion in a mouse model. Vaccine 21:3126–3136
Buendía AJ, Nicolás L, Ortega N, Gallego MC, Martinez CM, Sanchez J, Caro MR, Navarro JA, Salinas J (2007) Characterization of a murine model of intranasal infection suitable for testing vaccines against C. abortus. Vet Immunol Immunopathol 115:76–86
Nateghi Rostami M, Keshavarz H, Khamesipour A (2010) Immune responses of BALB/c mice against an experimental vaccine of alum precipitated autoclaved Leishmania major (Alum-ALM) mixed with BCG or mycobacterium vaccae. Trop Biomed 27:89–102
Howard JG, Liew FY (1984) Mechanisms of acquired immunity in leishmaniasis. Philos Trans R Soc Lond B Biol Sci 307:87–98
Bhowmick S, Ravindran R, Ali N (2007) Leishmanial antigens in liposomes promote protective immunity and provide immunotherapy against visceral leishmaniasis via polarized Th1 response. Vaccine 25:6544–6556
Jaafari MR, Ghafarian A, Farrokh-Gisour A, Samiei A, Kheiri MT, Mahboudi F, Barkhordari F, Khamesipour A, McMaster WR (2006) Immune response and protection assay of recombinant major surface glycoprotein of Leishmania (rgp63) reconstituted with liposomes in BALB/c mice. Vaccine 24:5708–5717
Bhardwaj S, Vasishta RK, Arora SK (2009) Vaccination with a novel recombinant Leishmania antigen plus MPL provides partial protection against L. donovani challenge in experimental model of visceral leishmaniasis. Exp Parasitol 121:29–37
Fujiwara RT, Vale AM, Franca-Silva JC, Costa RT, Quetz JS, Martins-Filho OA et al (2005) Immunogenicity in dogs of three recombinant antigens (TSA, LeIF and LmSTI1) potential vaccine candidates for canine visceral leishmaniasis. Vet Res 36:827–838
Tafaghodi M, Eskandari M, Kharazizadeh M, Khamesipour A, Jaafari MR (2010) Immunization against leishmaniasis by PLGA nanospheres loaded with an experimental autoclaved Leishmania major (ALM) and Quillaja saponins. Trop Biomed 27:639–650
El-On J (2009) Current status and perspectives of the immunotherapy of leishmaniasis. Isr Med Assoc J 11:623–628
Squires KE, Schreiber RD, McElrath MJ, Rubin BY, Anderson SL, Murray HW (1989) Experimental visceral leishmaniasis: role of endogenous IFN-γ in host defense and tissue granulomatous response. J Immunol 143:4244–4249
Miralles GD, Stoeckle MY, McDermott DF, Finkelman FD, Murray HW (1994) Th1 and Th2 cell-associated cytokines in experimental visceral leishmaniasis. Infect Immun 62:1058–1063
Murray HW (1997) Endogenous interleukin-12 regulates acquired resistance in experimental visceral leishmaniasis. J Infect Dis 175:1477–1479
Stober CB, Lange UG, Roberts MT, Alcam A, Blackwell JM (2005) IL-10 from regulatory T cells determines vaccine efficacy in murine Leishmania major infection. J Immunol 175:2517–2524
Solbach W, Laskay T (2000) The host response to Leishmania infection. Adv Immunol 74:275–317
Mohr E, Cunningham AF, Toellner K-M, Bobat S, Coughlan RE, Bird RA, MacLennan IC, Serre K (2010) IFN-gamma produced by CD8 T cells induces T-bet-dependent and -independent class switching in B cells in responses to alum-precipitated protein vaccine. Proc Natl Acad Sci USA 107:17292–17297
Gautam S, Kumar R, Maurya R, Nylen S, Ansari N, Rai M et al (2011) IL-10 neutralization promotes parasite clearance in splenic aspirate cells from patients with visceral leishmaniasis. J Infect Dis 204:1134–1137
Stober CB, Lange UG, Roberts MTM, Alcami A, Blackwell JM (2005) IL-10 from regulatory T cells determines vaccine efficacy in murine Leishmania major infection. J Immunol 175:2517–2524
Kamogawa Y, Minasi LA, Carding SR, Bottomly K, Flavell RA (1993) The relationship of IL-4 and IFN gamma-producing T cells studied by lineage ablation of IL-4 producing cells. Cell 75:985–995
Platzer C, Richter G, Uberla K, Muller W, Blocker H, Diamantstein T et al (1992) Analysis of cytokine mRNA levels in interleukin-4-transgenic mice by quantitative polymerase chain reaction. Eur J Immunol 22:1179–1184
Noble A, Kemeny DM (1995) Interleukin-4 enhances interferon-gamma synthesis but inhibits development of interferon-gamma-producing cells. Immunology 85:357–363
Huang LR, Chen FL, Chen YT, Lin YM, Kung JT (2000) Potent induction of long-term CD8+ T cell memory by short-term IL-4 exposure during T cell receptor stimulation. Proc Natl Acad Sci USA 97:3406–3411
Morrot A, Hafalla JC, Cockburn IA, Carvalho LH, Zavala F (2005) IL-4 receptor expression on CD8+ T cells is required for the development of protective memory responses against liver stages of malaria parasites. J Exp Med 202:551–560
Stager S, Alexander J, Kirby AC, Botto M, Rooijen NV, Smith DF et al (2003) Natural antibodies and complement are endogenous adjuvants for vaccine-induced CD8+ T-cell responses. Nat Med 9:1287–1292
Alexander J, Carter KC, Al-Fasi N, Satoskar A, Brombacher F (2000) Endogenous IL-4 is necessary for effective drug therapy against visceral leishmaniasis. Eur J Immunol 30:2935–2943
McFarlane E, Carter KC, McKenzie AN, Kaye PM, Brombacher F, Alexander J (2011) Endogenous IL-13 plays a crucial role in liver granuloma maturation during Leishmania donovani infection, independent of IL-4Rα-responsive macrophages and neutrophils. J Infect Dis 204:36–43
Basu R, Bhaumik S, Haldar AK, Naskar K, De T, Dana SK, Walden P, Roy S (2007) Hybrid cell vaccination resolves Leishmania donovani infection by eliciting a strong CD8 + cytotoxic T-lymphocyte response with concomitant suppression of interleukin-10 (IL-10) but not IL-4 or IL-13. Infect Immun 75:5956–5966
Watson DS, Endsley AN, Huang L (2012) Design considerations for liposomal vaccines: influence of formulation parameters on antibody and cell-mediated immune responses to liposome associated antigens. Vaccine 30:2256–2272
Bhowmick S, Mazumdar T, Ali N (2009) Vaccination route that induces transforming growth factor beta production fails to elicit protective immunity against Leishmania donovani infection. Infect Immun 77:1514–1523. doi:10.1128/IAI.01739-07
Morefield GL, Hawkins LD, Ishizaka ST, Kissner TL, Ulrich RG (2007) Synthetic Toll-like receptor 4 agonist enhances vaccine efficacy in an experimental model of toxic shock syndrome. Clin Vaccine Immunol 14:1499–1504
Rhee EG, Kelley RP, Agarwal I, Lynch DM, La Porte A, Simmons NL et al (2010) TLR4 ligands augment antigen-specific CD8+ T lymphocyte responses elicited by a viral vaccine vector. J Virol 84:10413–10419. doi:10.1128/JVI.00928-10
Mutiso JM, Macharia JC, Taracha E, Wafula K, Rikoi H, Gicheru MM (2012) Safety and Skin delayed-type hypersensitivity response in vervet monkeys immunized with Leishmania donovani sonicate antigen delivered with adjuvants. Rev Inst Med Trop Sao Paulo 54:37–41
Acknowledgments
The authors acknowledge the support provided by the Department of Science and Technology, India, under INSPIRE Fellowship programme.
Conflict of interest
The authors hereby declare that they have no conflicts of interest.
Ethical standard
The authors hereby declare that the experiments comply with the current laws in India.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Thakur, A., Kaur, H. & Kaur, S. Evaluation of the immunogenicity and protective efficacy of Killed Leishmania donovani antigen along with different adjuvants against experimental visceral leishmaniasis. Med Microbiol Immunol 204, 539–550 (2015). https://doi.org/10.1007/s00430-014-0367-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00430-014-0367-9