
Abstract
Three experiments were designed to determine the level of cooperation or interference observed from the forces generated in one limb on the forces exhibited by the contralateral limb when one or both limbs were producing a constant force (Experiment 1), one limb was producing a dynamic force while the other limb was producing a constant force (Experiment 2), and both limbs were producing dynamic force patterns (Experiment 3). The results for both Experiments 1 and 2 showed relatively strong positive time series cross correlations between the left and right limb forces indicating increases or decreases in the forces generated by one limb resulted in corresponding changes in the forces produced by the homologous muscles of the contralateral limb. Experiment 3 required participants to coordinate 1:1 and 1:2 rhythmical bimanual force production tasks when provided Lissajous feedback. The results indicated very effective performance of both bimanual coordination patterns. However, identifiable influences of right limb forces on the left limb force time series were observed in the 1:2 coordination pattern but not in the 1:1 pattern. The results of all three experiments support the notion that neural crosstalk is partially responsible for the stabilities and instabilities associated with bimanual coordination.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The ability to coordinate actions between the limbs is important for many activities of daily living. Tasks such as tying shoes, buttoning a shirt, slicing bread, and driving a car involve some type of coordination between the limbs. However, the role of each limb is dependent upon the task goal. Sometimes the task may require the limbs to produce mirror actions (symmetric bimanual coordination), such as when you clap your hands or row a boat. Sometimes the task may require different relationships between the limbs (asymmetric bimanual coordination). For example, the task may require one limb to act as a stabilizer while the other limb performs a specific movement such as opening a bottle or slicing bread. Still other tasks may require the limbs to produce fundamentally different actions, such as tying your shoes or buttoning your shirt. Although these examples are relatively easy for most individuals to produce, other coordination patterns between the limbs can be quite difficult. For example, playing the piano or guitar may require complicated, disparate actions for each hand, which underscore the difficulty that can be associated with certain asymmetric coordination patterns.
Research has traditionally indicated that tasks which require asymmetric coordination patterns are less stable and more difficult to perform than mirror symmetric coordination patterns (e.g., Carson, Byblow, Abernethy, & Summers, 1996; Carson, Riek, Smethurst, Lison Parraga, & Byblow, 2000; Cohen, 1971; Kelso, 1995; Riek, Carson, & Byblow, 1992; Scholz & Kelso, 1989; Semjen, Summers, & Cattaert, 1995; Summers, Maeder, Hiraga, & Alexander, 2008; Temporade, Zanone, Monno, & Laurent, 1999). To account for stability differences between symmetric and asymmetric bimanual coordination patterns and to help explain why producing some asymmetric coordination patterns are difficult, researchers often refer to a coalition of constraints that include neural crosstalk (see Swinnen, 2002; Swinnen & Wenderoth, 2004 for reviews).
Neural crosstalk is defined as a mirror image command sent to the homologous muscles of the contralateral limb (Cattaert, Semjen, & Summers, 1999; Swinnen, 2002). According to the crosstalk model, two independent motor plans exist for each limb and some fraction of the force command for one limb is diverted to the other limb (Cattaert et al., 1999). As such, neural crosstalk may result in the same commands dispatched to each limb (Cattaert et al., 1999). This idea is supported by an abundance of literature that has demonstrated an assimilation effect whereby both limbs tend to produce similar amplitudes (e.g., Heuer, Kleinsorge, Spijkers, & Steglich, 2001; Sherwood, 1994; Spijkers & Heuer, 1995), directions (e.g., Franz, Eliassen, Ivry, & Gazzaniga, 1996; Swinnen, Dounskaia, & Duysens, 2002; Swinnen, Dounskaia, Levin, & Duysens, 2001), frequencies (e.g., Peper, Beek, & van Wieringen, 1995b, c; Treffner & Turvey, 1993), or forces (e.g., Diedrichsen, Hazeltine, Nurss, & Ivry, 2003; Heuer, Spijkers, Steglich, & Kleinsorge, 2002; Steglich, Heuer, Spijkers, & Kleinsorge, 1999) despite task goals requiring disparate activation patterns for the two limbs. Because neural crosstalk conveys the same information to both limbs via cortical and subcortical pathways, it is believed that symmetrical actions are stabilized when complementary contralateral and ipsilateral signals are integrated while asymmetric actions can suffer from ongoing interference due to conflicting information or partial intermingling of signals controlling the two arms (Cardoso de Oliveira, 2002; Kagerer, Summers, & Semjen, 2003; Maki, Wong, Sugiura, Ozaki, & Sadato, 2008; Marteniuk, MacKenzie, & Baba, 1984). However, a clear understanding of how and when crosstalk facilitates or inhibits coordinated actions between the limbs has yet to be proposed.
It has been hypothesized that the effect of neural crosstalk is partially dependent on force levels, with higher forces resulting in stronger crosstalk effects and lower forces in weaker ones (Heuer et al., 2001, p. 69). Therefore, to determine the influence of crosstalk on the stability characteristics associated with bimanual coordination, we believe it is necessary to explore whether the forces generated in one limb affect the forces exhibited by the homologous muscles of the contralateral limb and if so, how and when this influence affects the coordination dynamics (e.g., facilitate or inhibit, cooperate or compete). Bimanual tasks which require the production and coordination of forces may provide further insight into the constraints acting on the system by allowing for the detection and characterization of neural crosstalk at the behavioral level.
Several research studies have used force tasks to investigate bimanual coordination. However, it is important to note that these tasks often involved discrete force pulses (e.g., Diedrichsen et al., 2003; Steglich et al., 1999; Therrien, Lyons, & Balasubramaniam, 2013), or manipulated the force requirements of the task by loading a lever system (e.g., Murian, Deschamps, & Temprado, 2008; Temprado, Chardenon, & Laurent, 2001). For example, Steglich et al. (1999) required participants to generate bimanual isometric force pulses of different magnitudes by matching target force levels presented as two vertical bars on a screen. Peak forces produced by the left and right limb were compared and correlated to determine the degree of coupling between the limbs. The results indicated a strong assimilation effect and high correlations between the mean peak forces for each limb. Note, however, these results were likely due to interference at the planning level (i.e., specification of two different force targets) rather than crosstalk at the execution level (i.e., how forces generated in one limb influence the force produced by the contralateral limb) (Diedrichsen et al., 2003; Spijkers & Heuer, 1995; Steglich et al., 1999). Similar results have been observed for amplitudes of bimanual reversal movements (Heuer et al., 2001). Rather than using discrete force pulses or simple limb/lever movements with load our experiments required the production of a pattern of forces throughout the test period so forces could be continuously monitored and maintained throughout a trial thereby allowing us to determine how forces generated in one limb influenced forces produced in the contralateral limb.
Although several previous experiments have used continuous bimanual force production tasks, it is important to note that these investigations have typically focused on the force sharing pattern between two or more effectors, such as multi-finger tasks, in the production of force by providing feedback that was the result of the sum of the forces produced (e.g., Hu, Loncharich, & Newell, 2011; Hu & Newell, 2011a, b; Latash, 2010; Shapkova, Shapkova, Goodman, Zatsiorskly, & Latash, 2008). For example, Hu and Newell (2011a) required participants to produce a constant force with both fingers so that the weighted sum of force output matched the target line. The study examined how different weighted coefficients imposed on each finger force regulated bimanual force coordination patterns. The results indicated greater performance error when the coefficients were unequal (asymmetric) compared to equal coefficients (symmetric). Using the same task design, Hu and Newell (2011b) also investigated the interactive effects of visual information and task-related force parameters (i.e., force level, force direction, and homologous muscle pairs) on coordination patterns in bimanual isometric force production. The results indicated that the coordination of bimanual isometric force is specified by the interaction of visual information and force level. Yet, research investigating the production of continuous bimanual force coordination has been primarily limited to feedback information that is the result of the sum of forces produced by each limb.
Recently, however, Therrien et al. (2013) compared bimanual force production when the visual feedback was summed (task goal was shared between the limbs) and when visual feedback was provided for each limb (each limb was responsible for the target force level simultaneously). The results indicated differences between the two types of bimanual tasks investigated. That is, when the task goal was shared, the force series produced by each hand were not correlated. However, when the bimanual task required each hand to reach different target force levels, the results indicated asymmetries in the degree of force overproduction between the hands following the removal of visual feedback. It is important to note, however, that the bimanual task involved the coordination of peak forces rather than continuous forces. It is not clear, whether the production of more continuous forces exhibit the same pattern of findings when each limb is responsible for maintaining the target force level rather than sharing the target force goal.
Therefore, the current investigation was designed to determine how and when the forces generated in one limb influenced the forces produced in the contralateral limb. However, given the wide variety of coordination patterns between the limbs necessary for activities of daily living, we investigated how the forces generated in one limb influenced the force produced in the homologous muscles of the contralateral limb in 3 different bimanual force tasks. More specifically, the three experiments were designed to determine how the forces generated in one limb influenced the forces produced by the contralateral limb when both limbs were producing symmetric constant forces (Experiment 1), one limb was producing a dynamic force pattern while the contralateral limb was producing a constant “stabilizing” force (Experiment 2), and both limbs were producing dynamic force patterns (Experiment 3).
To determine the influence the forces generated in one limb had on the forces produced in the contralateral limb, time series cross correlations were performed (Experiments 1 and 2). Correlations between left and right limb forces have been reported in the literature and interpreted as indicators of crosstalk effects (Heuer et al., 2001). Positive time series cross correlations would indicate an increase in force by one limb resulted in a corresponding increase in force by the contralateral limb whereas negative correlations would indicate that an increase in force by one limb are compensated for by a corresponding decrease in force by the other limb. Positive correlations would be consistent with the notion of neural crosstalk.
In addition, force harmonicity (H) was calculated (Experiment 3) to quantify perturbations in the force–time series. This value quantifies the harmonic nature of the forces produced by each limb for each half cycle similar to research that has quantified the harmonic nature of limb motion during reciprocal aiming tasks (Buchanan, Park, & Shea, 2006; Guiard, 1993, 1997). Harmonicity (H) is typically used to examine reciprocal aiming tasks to provide an estimate of the nature of the limb’s movement based upon an analysis of the inflection points in the acceleration trace (Guiard, 1993). Recently, however, this measure has been used to quantify the deflections in the force time series (Kennedy, Boyle, Rhee, & Shea, 2014). Perturbations in the force–time series in one limb that can be associated with force production in the contralateral limb would be consistent with the notion of neural crosstalk.
Experiment 1
Constant bimanual isometric forces
Mirror symmetrical force production requires the activation of homologous muscles; therefore the signals controlling each arm are always congruent (Maki et al., 2008). If the control signals directed to one limb are also sent to the contralateral limb during symmetrical force production the force commands from one limb should facilitate, or at least not interfere with the force produced in the contralateral limb. As such, neural crosstalk during mirror symmetric tasks likely stabilizes the coordinated action between the limbs (Maki et al., 2008).
Experiment 1 was designed to determine if the forces generated in one limb influenced the forces produced by the contralateral limb when both limbs were producing mirror symmetric constant forces. The production of unimanual forces was used as a control. Two different force levels (10 and 15 N) were investigated. We removed visual feedback from one limb to determine the influence force produced by one limb (with feedback) had on the contralateral limb (without feedback). The removal of visual feedback also allowed for the comparison of instructed versus uninstructed force production in the unimanual (control) tasks. That is, during bimanual performance of the task, participants were instructed to produce the same goal force with both limbs while in the unimanual task participants were only instructed to produce the goal force in one limb while the contralateral (uninstructed) limb remained in contact with the force transducer. However, we monitored force production for both limbs. As such, we were able to examine if there were unintended changes in force production in the contralateral limb that were associated with voluntary force production in the instructed limb.
To determine the influence the forces generated in one limb had on the forces produced in the contralateral limb, time series cross correlations were performed. We predicted that when forces changed in one limb related changes in force would be observed in the contralateral limb resulting in strong positive correlations between the limbs.
Methods
Participants
Self-declared right-handed undergraduate students (N = 9; age = 20.6 years ± 1.2; 5 males and 4 females) volunteered to participate in the experiment after reading and signing a consent form approved by the local Institutional Review Board for the ethical treatment of experimental participants. The participants had no prior experience with the experimental task and were not aware of the specific purpose of the study.
Apparatus
The apparatus consisted of two static force measurement systems, each of which included a force transducer calibrated with known weights and an amplifier that converted the physical force exerted against the transducer into a voltage representing the instantaneous value of the applied force. The voltages were recorded using an AD converter (DAS16) installed on the computer programmed to sample at 200 Hz. Targets and applied forces were displayed in a 1.64 × 1.23 m image projected on the wall in front of the participant (Fig. 1a). One force transducer was attached to an adjustable metal frame mounted on the left side of a standard padded treatment table and the other on the right side.

Illustration of the position of the participant and the display (a). Examples of the unimanual time series (b, f, j, n) and scatterplots (c, g, k, o). Note that in the unimanual condition the participant was only instructed to produce force with one limb. Although the participant was instructed to not produce force with the other limb any forces produced with the uninstructed limb were recorded. Examples of bimanual times series (d, h, l, p) and scatterplot (e, i, m, q). In the bimanual condition the participant was instructed to produce the same goal force with both limbs
Procedure
Participants were positioned in a supine position on the treatment table with their head elevated with a pillow to view a projected image on the wall located 2 m away. The projected image was used to display the target force levels and the force produced by the arms. The force transducers were adjusted so that the participant’s arm (just below the wrist) contacted the force transducer comfortably while the elbow was in contact with the table and the lower arm was at a 90° angle to the upper arm. This position allowed for the isolation of force produced by the limbs without the effects of gravity and mechanical/elastic properties associated with the upper limbs (Hill, 1970). In Fig. 1a, the force transducer set ups for the left and the right limb tasks are illustrated. To signal the start of each trial, the target lines for that trial were presented in the display projected onto the wall in front of them. Participants were then instructed to exert force against the force transducer in an attempt to produce continuous force that matched as closely as possible the target lines displayed on the computer monitor. In the unimanual task, participants were instructed to produce force with only one limb while resting the other limb against the force transducer. In the bimanual task participants were instructed to produce the same goal force with both limbs. Participants were reminded that their response should be continuous and that they should attempt to reduce the differences between the target lines and their response over practice. Participants were required to perform the unimanual and bimanual tasks at two different force levels (10 and 15 N). Participants performed six 20 s trials for each task. In each trial the online display of the actual forces for one limb was removed during the second half of each trial. In 50 % of the trials visual feedback was removed for the dominant limb, while in the other half visual feedback was removed for the non-dominant limb. Visual feedback was removed from only one limb in each trial to determine the influence force produced in one limb (with feedback) had on the contralateral limb (without feedback).
Measures and data analysis
All data reduction was performed using MATLAB. The analog signals representing the limbs’ force were filtered (second order dual pass Butterworth with a cutoff frequency of 10 Hz).
The last 10 s of the test trials were subjected to analysis. Mean force and the coefficient of variation (CV) of force were used to evaluate accuracy and stability of left and right limb performance. CV is a measure of relative stability, which quantifies total variability [standard deviation (STD)] as a function of the mean force output [CV = ((STD/mean force) × 100)]. Low CV scores indicate stable force production while high CV scores indicate less stable performance. In addition, the time series cross correlations of the left and right limb forces were computed for each task to determine crosstalk effects (Heuer et al., 2001).
Mean force and CV of force were analyzed in separate task (unimanual, bimanual) × limb (left, right) × feedback limb (left, right) × goal force (10, 15 N) analyses of variance (ANOVA) with repeated measures on all factors. Correlations were analyzed in a feedback limb (left, right) × goal force (10, 15 N) ANOVA with repeated measures on all factors. Simple main effects analysis was used to further investigate significant interactions and effect size (η 2 p —partial eta squares) was reported for significant effects.
Results
Figure 1 provides examples of the left and right limb forces produced by one participant during the last 10 s of the unimanual (left panels) and bimanual (right panels) tasks. The corresponding scatterplots, linear regression lines, and time series cross correlations of the left and right limb force time series are also presented (Fig. 1c, e, g, i, k, m, o, q). Note the positive time series cross correlations in both the unimanual and bimanual tasks. Figure 2 depicts mean force (a–c, j–l), CV force (d–f, m–o), and time series cross correlations (g–i, p–r) by limb and feedback conditions. The left panels are from the 10 N goal force trials and the right panels for the 15 N goal force trials. Note, the above baseline force production in the uninstructed limb (Fig. 2b, c, k, l) and the associated positive correlations (Fig. 2h, i, q, r).

Mean force, CV of force, and mean time series cross correlations for Experiment 1 are presented for the 10 N (a–i) and 15 N (j–r) goal force bimanual conditions by limb and feedback conditions. Error bars represent standard errors
Bimanual and unimanual conditions
Force As expected the analysis detected a main effect of goal force, F(1,141) = 1,006.66, p < 0.01, η 2 p = 0.88, with greater forces produced in response to the 15 N goal than for the 10 N goal. The main effect of limb, F(1,141) = 5.30, p < 0.05, η 2 p = 0.15, was also significant. In addition, there were significant goal force × limb, F(1,141) = 4.34, p < 0.05, η 2 p = 0.14, goal force × feedback limb, F(1,141) = 4.18, p < 0.05, η 2 p = 0.14, limb × feedback limb, F(1,141) = 5.83, p < 0.05, η 2 p = 0.15, and task × limb × feedback limb, F(1,141) = 6.41, p < 0.05, η 2 p = 0.16, interactions. Simple main effects analysis of the task × limb × feedback limb interaction indicated that for the unimanual tasks the instructed (left, right) limb produced higher forces than that produced by the uninstructed limb, while both limbs produced similar forces in the bimanual task. The simple main effects analysis also indicated that when the limb was instructed to produce the target force the right limb produced higher forces when not provided feedback than the left limb when not provided feedback. When provided feedback the limbs produced similar forces. No differences were detected for the forces produced by the uninstructed limb (Fig. 2a–c, j–l). All other main effects and interactions were not significance.
Coefficient of variation (CV) The analysis detected a main effect of task, F(1,141) = 35.15, p < 0.05, η 2 p = 0.2. In addition, there was a significant task × limb × feedback limb, F(1,141) = 7.46, p < 0.05, η 2 p = 0.08, interaction. Simple main effects analysis indicated that for the bimanual task the CV of force was lower for the limb provided feedback than the limb where feedback was withdrawn. However, for the unimanual task the CV of force was higher for the uninstructed limb, which was not required to produce a force, than for the instructed limb that was required to produce 10 or 15 N goal force (Fig. 2d–f, m–o). These results indicated that more force fluctuations (e.g., adjustments, hesitations, and/or perturbations) were present in the force time series for the limb without feedback compared to the limb with feedback and in the force time series for the uninstructed limb when compared to the instructed limb. All other main effects and interactions were not significance.
Time series cross correlations The analysis of the time series cross correlations between left and right limb forces indicated significant time series cross correlations in 92 % (33 of 36) of the trials. The analysis of the time series cross correlations failed to detect any main effects and interaction. Interestingly, the consistent positive correlations between the forces produced by the limbs were found regardless of the goal force requirements (10 or 15 N) or whether one (unimanual task) or both (bimanual task) limbs were instructed to produce a specific force (Fig. 2g–i, p–r).
Discussion
The purpose of Experiment 1 was to investigate the unimanual and bimanual production of constant forces. Mean forces in the two limbs were determined to document that the participants were producing the goal pattern of force and CV of force was analyzed to determine if force variability (stability) for the limbs was influenced by the force produced in the contralateral limb. Time series cross correlations were also performed to determine if the forces produced by one limb were influenced by the forces produced in the contralateral limb. The neural crosstalk model would predict positive correlations between the forces produced by the two limbs.
Participants were quite effective in producing the instructed forces especially for the limb that was provided feedback throughout the trial. The analysis of the time series cross correlations indicated moderate, but significant positive correlations. Positive correlations indicate an increase or decrease in the force generated by one limb resulted in a corresponding increase or decrease in the force produced by the contralateral limb. This phenomenon is consistent with the notion of crosstalk. These results are also consistent with Diedrichsen et al. (2003), who reported positive correlations in their force task. However, it is important to note that the task used by Diedrichsen et al. involved discrete force pulses whereas our tasks involved the production of continuous forces. Using continuous forces, the forces could be monitored throughout a trial thereby allowing us to examine the time series cross correlations within a trial rather than the changes from trial to trial.
The muscle synergy literature (see Latash, 2010, 2012 for reviews), however, has demonstrated coupling effects whereby decreases in force by one effector are compensated for by corresponding increases in force by the other effector. This error compensation feature results in the stabilization of overall force production with negative correlations observed between the corresponding forces. Previous researchers that have reported negative correlation between two or more effectors responsible for the production of force have often employed the uncontrolled manifold (UCM) framework to analyze muscle synergies. However, this type of analysis was not appropriate for our examination of bimanual force control. First, the UCM framework is concerned with the sharing pattern between two or more effectors, such as multi-finger tasks, in the production of force. For example, Shapkova et al. (2008) used a task (Experiment 2) that involved pressing on a force sensor with multiple fingers to match a target line. The feedback provided was the result of the sum of the forces produced by each finger. In the present experiment, each limb was responsible for producing the target force level and as such we were not concerned with force sharing between the limbs. More specifically, feedback was provided for the force produced by each limb (first 10 s of each trial) and not the sum of the forces of the two limbs. Second, the UCM hypothesis framework examines motor output variability across large sets of trials. This means the results reflect the change in the effectors contribution to the total force from trial to trial rather than the dynamic interchange of forces within a trial. Because we were concerned with the mechanisms responsible for stability characteristics associated with symmetrical and asymmetrical force control, a within trial examination of the force times series from the two limbs was more appropriate for our research questions. Therefore, we used a time series cross correlation analysis to determine how one limb influenced the other limb during the production of force across a trial and to determine the degree to which the two limbs were constrained to act as a single functional unit.
The results indicated significant positive correlations between the forces produced by the left and right limb regardless of the task. Significant (p < 0.05) positive correlations were found in 92 % of the unimanual and bimanual trials with the average time series cross correlation for various conditions ranging from 0.24 to 0.46. Note for the unimanual tasks that participants were instructed to produce force with only one limb, but we monitored the forces of both limbs. Interestingly, the uninstructed limb produced minimal forces that appeared to be entrained to the forces produced by the instructed limb. In fact, the CV of force was larger in the uninstructed limb, where minimal forces were produced than in the instructed limb where 10 or 15 N of force were maintained. This indicated that a wider range of force fluctuations (e.g., adjustments, hesitations, and/or perturbations) occurred in the uninstructed limb than would be expected given the low levels of force maintained. Presumably the additional variability was a result of the forces produced in the contralateral limb. These results are consistent with predictions based on cortical and subcortical neural crosstalk (e.g., Cattaert et al., 1999). That is, the forces produced by one limb resulted in subtle but complementary changes in the forces produced in the homologous muscle of the contralateral limb.
Experiment 2
Constant and dynamic bimanual isometric forces
The results from Experiment 1 indicated subtle but consistent evidence of interplay between the forces generated with one limb and the forces produced by the homologous muscles of the contralateral limb, regardless of whether the contralateral limb was instructed to produce a similar force or were uninstructed. Experiment 2 was designed to determine if the production of a dynamic pattern of force produced by tracking a signwave template in the display with one limb influenced the maintenance of a constant force with the contralateral limb. The bimanual force task is consistent with activities of daily living that require one limb to perform a dynamic activity while the contralateral limb provides a supporting and/or stabilizing role, such as opening a bottle or slicing bread.
Participants were asked to produce a pattern of force with one limb using a template placed in the display while the other limb manipulated a constant force. The template was constructed from a sine wave with an amplitude twice that of the constant force. Thus, the average forces produced by the two limbs over the course of a trial should be similar but the force variability would be substantially different.
Visual feedback was removed from the limb performing the constant force to determine the influence force generated in the dynamic limb (with feedback) had on the contralateral limb (without feedback). Based upon the results of Experiment 1 and the notion that a mirror image command is also dispatched to the contralateral limb, we anticipated that the active manipulation of force with one limb would result in a changing pattern of force in the limb required to maintain a constant force. More specifically, we predicted that increases and decreases in force generated in the dynamic limb would produce similar increases and decreases in the contralateral limb performing the constant force, resulting in positive correlations between the forces produced by the two limbs.
Methods
Participants
The same right-handed undergraduate students (N = 9; age = 20.6 years ± 1.2; 5 males and 4 females) in Experiment 1 volunteered to participate in the experiment after reading and signing a consent form approved by the local Institutional Review Board for the ethical treatment of experimental participants. The participants were not aware of the specific purpose of the study.
Apparatus
The same apparatus as used in Experiment 1 was used for Experiment 2.
Procedure
The same procedure as Experiment 1 was used for Experiment 2 except participants were instructed to exert force against the force transducer with one limb in an attempt to produce a constant force and a continuous pattern of force that followed the pattern illustrated by the target waveform presented in the display with the contralateral limb. Participants were reminded that their response should be continuous and that they should attempt to reduce the differences between the target line and their response over practice. Then participants were tested in a bimanual task in which one limb was required to produce the constant force while the other limb concurrently produced the sinusoidal pattern of force. Participants performed six 20 s trials for each practice and test conditions at two different force levels. In each trial the online display of the actual forces for the limb performing the constant force task was removed for the second half (10 s) of the trial.
Measures and data analysis
The initial data reduction and filtering was conducted as described in Experiment 1 with the last 10 s of each test trial subjected to analysis. Mean force and root mean square error (RMSE) of force were used to evaluate accuracy and stability of left and right limb performance. RMSE was used to determine the difference between the force time series and the goal force pattern (constant or sine wave). Time series cross correlations between the left and right limb force times series were calculated to determine the influences of the forces produced in one limb on the forces produced in the contralateral limb.
Mean force and RMSE of force were analyzed in separate limb (left, right) × task (constant, sine wave) × goal force (10, 15 N) ANOVA with repeated measures on all factors. Correlations were analyzed in a limb (Left sine wave, right sine wave) × goal force (10, 15 N) ANOVA with repeated measures on both factors. Simple main effects analysis was used to further investigate significant interactions and effect size (η 2 p —partial eta squares) was reported for significant effects.
Results
Examples of the forces produced on the test trials during the last 10 s of the bimanual task are provided in Fig. 3a, d, g, j. Scatterplots of the left and right limb forces, linear regression lines, time series cross correlations (Fig. 3b, e, h, k), and the normalized force times series are also presented (Fig. 3c, f, i, l). Note that the pattern of force produced by the limb producing the constant force appears to increase or decrease coincident with the increase or decrease in force produced in the dynamic limb. This is particularly apparent when the force is normalized (Fig. 3c, f, i, l). Figure 4 provides the mean forces (a, b), RMSE of force (c, d), and the time series cross correlations (e, f), for the 10 N (left panels) and 15 N (right panels) goal force, respectively. Note the positive correlations between the force produced by each limb (Fig. 4e, f).

Examples of the left and right limb forces for one participant in the 15 N (a, d) and 10 N (g, j) bimanual conditions in Experiment 2. Note that feedback was withdrawn during the last 10 s for right limb in a and g. Feedback was withdrawn for left limb in d and j. Scatterplots and times series cross correlations of the left and right limb forces time series (b, e, h, k) and normalized force times series for the same left and right limb forces for the various conditions are also presented (c, f, i, l)

Mean force, RMSE of force, and time series cross correlations in Experiment 2 are presented for the 10 N (a, c, e) and 15 N (b, d, f) goal force bimanual conditions by feedback conditions. Note that the limb/task notation includes limb (l, r) and task (C constant goal force, S sine wave goal force) for force and RMSE. The limb/task notation for the times series cross correlations indicates the limb that maintained a constant force. Error bars represent standard errors
Force The analysis indicated main effects of goal force, F(1,63) = 182.44, p < 0.05, η 2 p = 0.74, with greater forces for the 15 N goal force than for the 10 N. In addition, there was a goal force × task interaction, F(1,63) = 4.06, p < 0.05, η 2 p = 0.16, with moderately higher forces in the sine task than in the constant force task (Fig. 4a, b). All other main effects and interactions were not significant.
RMSE force The analysis indicated main effects of goal force, F(1,63) = 10.05, p < 0.05, η 2 p = 0.15, with the RMSE for the 15 N goal force moderately higher than for the 10 N goal force. The main effect of task, F(1,63) = 69.90, p < 0.01, η 2 p = 0.99, was also significant with substantially greater RMSE scores in the sine task than in the constant force task (Fig. 4c, d). All other main effects and interactions were not significant.
Time series cross correlations The time series cross correlations were significant (p < 0.01) for all participants on all trials with positive correlations (mean 0.618, SEM 0.032) observed between the forces produced by the limbs (Fig. 4e, f). The analysis of the time series cross correlations failed to detect any main effects and interaction.
Discussion
The purpose of Experiment 2 was to investigate the bimanual production of forces where one limb was required to produce a dynamic pattern of force while the homologous muscles of the contralateral limb were required to produce a constant force. Given the findings from Experiment 1 and the notion of neural crosstalk, we anticipated the changing pattern of forces in one limb would have a more profound effect on the contralateral limb than the constant forces used in Experiment 1. In Experiment 1 the commands to both limbs were congruent (Maki et al., 2008) while in Experiment 2 the commands to each limb were in conflict. When conflicting activation patterns are required for the left and right limbs, an individual must inhibit or compensate for the crosstalk that is dispatched to the contralateral limb (Barral, Debu, & Rival, 2006; Barral, Debu, & Rival, 2010). If the inhibition or compensation for the crosstalk fails, the interference may challenge the stability of the coordination dynamics. In theory, therefore a failure to inhibit, suppress, or otherwise compensate for, the neural crosstalk may result in unwanted perturbations to the coordinated action (Houweling, Beek, & Daffertshofer, 2010). As such, the commands required to increase and decrease the forces in the limb producing the dynamic pattern of forces could trigger via neural crosstalk a greater impact of this limb on the limb attempting to maintain a constant force.
In Experiment 2 participants were quite effective in producing the instructed forces with the two limbs producing similar average forces over the last 10 s of the test trial. As expected, RMSE of force increased as the goal force was increased and RMSE of force was substantially larger for the limb producing the sine wave pattern of force than the limb producing the constant force. As in Experiment 1, the analysis indicated significant positive correlations between the force time series produced by the left and right limb regardless of the task. In fact, significant positive correlations were found in 100 % of test trials with the average correlation for specific conditions ranging from r = 0.59 to 0.66. Positive correlations indicated that as force increased or decreased in one limb, similar increases and decreases occurred in the contralateral limb. This relationship is best illustrated in Fig. 3c, f, i, l where the forces are normalized.
This relationship, however, was not influenced by the limb (left or right) producing the force. This result is inconsistent with previous research that has indicated specialized roles for the dominant and non-dominant limb in bimanual tasks. More specifically, previous research has consistently indicated that the right limb is specialized for performing the dynamic role, while the non-dominant limb is specialized for the supporting and/or stabilizing role (e.g., Guiard, 1987; Sainburg, 2010; Stone, Bryant, & Gonzalez, 2013). In addition, previous research has indicated that neural crosstalk from the dominant left hemisphere exerts a stronger influence on the non-dominant left limb than the non-dominant right hemisphere on the dominant right limb (Kagerer et al., 2003; Maki et al., 2008). As such, it would be expected for the left limb to be less accurate than the right limb, especially when performing the dynamic pattern of force. Yet, there were no differences in accuracy between the forces produced by the left and right limbs.
The results were, however, consistent with predictions based on cortical and subcortical neural crosstalk (e.g., Cattaert et al., 1999). That is, the forces produced with a muscle set in one limb resulted in subtle but complementary changes in the forces produced in the homologous muscles of the contralateral limb. Although it is sometimes contentious to make cross experiment comparisons because of the differences across experiments, a dependent same t test was conducted on the average time series cross correlations for each participant in Experiment 1 and 2. Note that a dependent sample t test was used because the same participants were utilized in Experiment 1 and 2. This analysis indicated time series cross correlations in Experiment 2 were significantly higher, t(8) = 12.79, p < 0.01, than in Experiment 1.
Experiment 3
Rhythmical 1:1 and 1:2 bimanual forces
The results from Experiment 1 and 2 indicated consistently significant time series cross correlations between the forces produced by the two limbs. Furthermore, the results of Experiment 2 indicated that the dynamic pattern of forces produced by one limb systematically impacted the maintenance of constant force by the contralateral limb. That is, as the forces increased for the limb producing the sine wave pattern small but consistent increases in force were observed in contralateral limb. Likewise, as forces decreased for the limb producing the sine wave pattern small but consistent decreases in force were observed in the contralateral limb. This resulted in significant time series cross correlations between the forces produced by the two limbs. Experiment 3 was designed to investigate the bimanual production and coordination of force when both limbs were producing different patterns of dynamic forces. Specifically, Experiment 3 was designed to determine participants’ ability to coordinate a 1:2 rhythmical bimanual force patterns. A 1:1 force pattern was used as a control. In the 1:1 task the commands to both limbs were congruent (Maki et al., 2008) while in the 1:2 the commands to each limb were in conflict (Summers et al., 1993).
Because the 1:2 coordination task required the simultaneous production of two conflicting motor sequences (Summers et al., 1993), it may allow for a clearer picture of how and when neural crosstalk influences bimanual coordination. The coordination of 1:2 bimanual patterns has been investigated extensively using a wide variety of tasks such as: tapping (e.g., Deutsch, 1983; Semjen & Summers, 2002; Serrien, 2009), pendulums (e.g., Sternad, Turvey, & Saltzman, 1999a, b, c), circling/dial rotation (e.g., Beets et al., 2014; Boyles, Panzer, & Shea, 2012; Sisti et al., 2011) flexion–extension movements (e.g., Levin, Suy, Huybrechts, Vangheluwe, & Swinnen, 2004; Puttemans, Wenderoth, & Swinnen, 2005), pronation–supination movements (e.g., Byblow, Bysouth-Young, Summers, & Carson, 1998; Summers, Davis, & Byblow, 2002) and lever movements (e.g., Kovacs, Buchanan, & Shea, 2010a, b; Serrien & Swinnen, 1997; Swinnen, Dounskaia, Walter, & Serrien, 1997). While it has been recognized that 1:2 discrete tapping patterns are relatively easy to perform (e.g., Deutsch, 1983; Kovacs, Buchanan, & Shea, 1995a; Summers et al., 1993), continuous 1:2 bimanual tasks appear to pose difficult challenges for the CNS (e.g., Puttemans et al., 2005; Summers et al., 2002; Swinnen et al., 1997). However, these difficulties appear to be circumvented, or at least minimized, when integrated feedback (e.g., Lissajous displays) is provided (e.g., Boyles et al., 2012; Hessler, Gonzales, & Amazeen, 2010; Kovacs et al. 2010b).
The Lissajous displays used by Kovacs and colleagues, for example, provided a goal template along with on-line integrated visual information regarding the position of the two limbs as a single point in one plane. Lissajous plots have been used in a number of bimanual coordination experiments and the overall results indicated that this type of feedback allowed participants to rapidly tune-in a wide variety (e.g., 1:2; 2:1, 3:2, 4:3, 5:3, 90° relative phase) of bimanual coordination patterns (Boyles et al., 2012; Kovacs et al., 2010a, b; Kovacs & Shea, 2011; Swinnen et al., 1997) with only a few minutes of practice. It believed that Lissajous plots facilitate the successful performance of complex polyrhythms because the integrated information greatly reduces the perceptual and attentional difficulties associated with the task (Kovacs et al., 2009, 2010a, b; Kovacs & Shea, 2011). It is important to note, however, that most of the previous research using integrated feedback information to coordinate multifrequency coordination patterns has involved bimanual coordination tasks performed in near frictionless environments.
Only recently have participants been required to coordinate a pattern of rhythmical bimanual forces when provided Lissajous information to reduce attentional demands (Kennedy et al., 2014). The results indicated that integrated feedback (e.g., Lissajous plots) allowed for the successful performance of a complex multifrequency force coordination task similar to previous bimanual multifrequency research conducted in near frictionless environments (Boyles et al., 2012; Kovacs et al., 2010a, b; Kovacs & Shea, 2010, 2011).
The task in the present experiment was to rhythmically produce a pattern of continuous isometric forces on a left-side force transducer with the left arm that was coordinated with the pattern of continuous isometric forces produced on a right-sided force transducer with the right arm in a 1:1 or 1:2 coordination pattern. The purpose of the experiment was to determine participants’ ability to coordinate a 1:1 or 1:2 continuous bimanual force production tasks and to determine if the forces generated by one limb influenced the forces produced by the contralateral limb. We predicted that individuals would be able to effectively perform the force coordination tasks when provided Lissajous displays. We examined the 1:1 (symmetric) force coordination pattern to have baseline measure of performance from which we could evaluate the performance of the 1:2 (asymmetric) task. Based on the finding of Experiment 1 and 2 and the notion of neural crosstalk, we predicted that in the 1:2 task the forces produced by one limb would result in subtle perturbations to the pattern of forces in the contralateral limb. We also predicted that these types of perturbations would not be observed in the 1:1 task because the complimentary nature of neural crosstalk should facilitate the symmetric pattern (Maki et al., 2008) whereas neural crosstalk may challenge the stability of the asymmetric pattern, resulting in unwanted perturbations to the coordinated action (Houweling et al., 2010).
Methods
Participants
Self-declared right-handed undergraduate students (N = 16; age = 21.3 years ± 1.9; 7 males and 9 females) with little or no musical experience volunteered to participate in the experiment after reading and signing a consent form approved by the local Institutional Review Board for the ethical treatment of experimental participants. The participants had no prior experience with the experimental task and were not aware of the specific purpose of the study.
Apparatus
The same apparatus as used in Experiments 1 and 2 was used for Experiment 3.
Procedure
Prior to entering the testing room participants were randomly assigned to either a 1:1 or 1:2 bimanual coordination task. The same procedure as Experiment 1 and 2 was used in Experiment 3 except participants were instructed to rhythmically produce a pattern of isometric forces on the left-side transducer with the left arm that was coordinated with the pattern of isometric forces produced on the right-sided force transducer with the right arm in a 1:1 or 1:2 frequency ratio using the display information to guide their performance. Lissajous displays provided integrated visual information regarding the position of the two limbs as a single point (cursor) in one plane. The Lissajous displays incorporated a goal template and a cursor indicating the forces produced with both limbs. The template illustrated the specific pattern of force requirements needed to produce the goal coordination pattern of 1:1 or 1:2 (see Fig. 5b, d). The cursor moved from left to right as force was increased with the right arm and from bottom to top as force was increased by the left. The maximum force required to produce the pattern illustrated by the template was ≈15 N. The 1:1 bimanual force coordination pattern required participants to produce a pattern of force (peak ≈15 N) with both the right and left limb simultaneously while the 1:2 bimanual force coordination pattern required the right limb to produce two patterns of force (peak ≈15 N) for each pattern produced by the left limb. Participants were required to perform 14 trials. Each trial was 20 s with a 10 s rest period between trials. After any trial in which the average frequency of the right limb was below 1.0 Hz, the experimenter encouraged the participants to increase the speed with which they produced the patterns of force without disrupting the goal pattern.

Illustration depicting the goal 1:1 (a) and 1:2 (c) left and right limb times series (a) and resulting Lissajous plot for the 1:1 (b) and 1:2 (d) bimanual coordination tasks in Experiment 3. Sample performance and resulting Lissajous for one participant in the 1:1 (e, f) and 1:2 (g, h) coordination tasks. In addition, the force and force velocity time series traces and velocity/force plots resulting from the right (i, j) and left (m, n) limb forces in the 1:1 task and the right (k, l) and left (o, p) limb forces in the 1:2 task are provided (Color figure online)
Measures and data analysis
All data reduction was performed using MATLAB. The analog signals representing the limbs’ force were filtered (second order dual pass Butterworth with a cutoff frequency of 10 Hz). Force velocity and acceleration signals were computed using a 3-point difference algorithm before performing the next differentiation. Note, that coordinating rhythmical patterns of dynamic force with both limbs required both force and timing goals. As such, the analysis focused on both unimanual force performance and bimanual force performance of the required frequency ratio.
Bimanual measures Inter-peak intervals for the right and left limbs were used to determine point estimates of mean cycle duration and were used to compute a frequency ratio of left limb cycle duration to right limb cycle duration. This measure provided a temporal measure of goal attainment that was independent of limb coordination tendencies and actual limb force trajectories. To examine the continuous spatial–temporal coordination of the limb forces, continuous phase angles for the two limbs were also computed. The phase angle (θ i ) for each limb (i = r, l) was computed for each sample of the force time series as follows (Kelso, Scholz, & Schoner, 1986):
with X i representing the normalized force of the right and left limbs and dX i /dt the instantaneous normalized force velocities for the right and left limbs. Next, the individual phase angles θ i were unwrapped by finding absolute jumps greater than 2π and adding appropriate multiples of 2π to each data point following the jump. After the unwrapping, regression analyses of the continuous relative phase angles for the right and left limbs were conducted to determine the slope of unwrapped phase angles across the trial. The ratio of the right limb slope to left limb slope provided a second measure of bimanual goal attainment.
Unimanual measures Inter-peak intervals were computed on a cycle basis with each interval representing the time between two consecutive force peaks (Inter-peak interval = forcepeak i+2 − Forcepeak i ). Inter-peak interval variability was defined as the standard deviation of the inter-peak intervals within a trial. These values were determined for both the left and right limb forces. These measures provided information related to the timing of the goal coordination pattern. A goal value of 1.0 for inter-peak interval would indicate that the interval for the right and left limb was equal while a goal value of 2.0 would indicate that the interval for the left limb was twice that of the right limb.
To quantify the deflections in the force–time series, we examined the force velocity and acceleration profiles associated with the production of force by each limb to determine force harmonicity (H) (Kennedy et al., 2014). This value quantified the harmonic nature of the forces produced by each limb for each half cycle similar to research that has quantified the harmonic nature of limb motion during repetitive aiming tasks (Buchanan et al., 2006; Guiard, 1993, 1997). To compute an index of force H (see Guiard, 1993), non-overlapping windows between a pair of zero crossings in the force trace were defined. Each non-overlapping time window comprised a single force trace reversal. Within each time window, all deflections of the normalized force acceleration trace were identified. When the force acceleration trace reversals were all positive (negative force value) or negative (positive force value) within this window, H was computed as the ratio of minimum to maximum force acceleration reversals. When the minimum force acceleration reversals equaled zero within this time window, H was set to 0. When only one single force acceleration reversal was detected within this window, the value of H was set to 1, indicating a harmonic force was exerted (without any jerks in force acceleration) of the limb. If the acceleration trace crossed from positive to negative (or vice versa) within this window, the value of H was set to 0, indicating an inharmonic force trace was exerted (with a larger jerk in force acceleration). Finally, the individual force harmonicity values of each time window for a trial were averaged yielding a global estimate of force H.
To examine the control of force during the coordination task the peak force and mean force produced were calculated to determine the accuracy of left and right limb force performance. The peak forces produced during the trial were calculated by averaging the absolute peak force per cycle for each participant during the trial. The mean force produced was quantified by averaging the absolute force produced during the trial. Note the goal coordination pattern required a peak force of ≈15 N, and a mean force of ≈7.5 N.
The mean inter-peak interval and phase angle slope ratios were analyzed in separate task (1:1, 1:2) ANOVA. Peak force, mean force, inter-peak interval, standard deviation (STD) of inter-peak interval, phase angle slope and force harmonicity were analyzed in separate task (1:1, 1:2) × limb (left, right) ANOVAs with repeated measure on limb. Simple main effects analysis was used to further investigate significant interactions and effect size (η 2 p —partial eta squares) was reported for significant effects.
Results
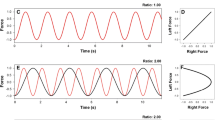
Figure 5 provides an illustration depicting the goal 1:1 (a) and 1:2 (c) left and right limb time series and resulting Lissajous display for the 1:1 (b) and 1:2 (d) bimanual coordination task. Figure 5 also provides sample force and force velocity time series and the resulting Lissajous plots for one participant tested on the 1:1 task (e, f) and one participant tested on the 1:2 (g, h) task. The force and force velocity time series traces and normalized force velocity/normalized force plots resulting from the right (i–l) and left limb (m–p) forces were also provided. In addition, grey bars were included to highlight the point of force initiation (force onset) by the right limb while yellow bars highlighted the release of force (force offset). The placement and width of the grey and yellow bars was determined by identifying peak force velocity in each half cycle and then tracing backwards (force onset) and forwards (force offset) to 5 % of peak force velocity. In the 1:2 task both the grey and yellow bars captured the observed distortions in the left limb (Fig. 5o, red trace). Importantly, these types of distortions in the force time series were not observed in the 1:1 task when both limbs received congruent signals (Maki et al., 2008). In the 1:2 task it appeared that as the right limb initiated force there was a corresponding increase in force in the left limb and when the right limb released force there was a corresponding decrease in force produced by the left limb. In addition, the observed interference in the force produced by the left limb in the 1:2 task continued until peak force velocity was obtained by the right limb (5 K) resulting in observable distortions in both the force and force velocity profiles of the left limb (5O).
Mean data related to timing of the force pattern and force production for participants in the 1:1 and 1:2 tasks are provided in Fig. 6. The blue bars indicate the goal ratio or goal force required by the template. Based upon point estimates of mean cycle duration (a) as well as continuous phase angles for the two limbs (e) participants were able to accurately ‘tune in’ the 1:1 (goal ratio = 1) and 1:2 (goal ratio = 2) coordination pattern. Participants were also able to accurately produce the required maximum (≈15 N) and mean (≈7.5 N) force required by the template (d, g) with both the right and left limb. However, the left limb performance in the 1:2 coordination was significantly slower (b), less stable (c), had a different phase angle slope (f), and was less harmonic (h) than the right limb in the 1:2 pattern and the left limb in the 1:1 pattern.

Mean inter-peak interval ratio (a), inter-peak intervals (b), STD inter-peak intervals (c), peak force (d), phase angle slope ratio (e), phase angle slope (f), force (g), and force harmonicity (f) by task (1:1 and 1:2) in Experiment 3. Note that the goal ratios (blue) are provided in a and e. Error bars represent standard errors (Color figure online)
Bimanual analyses
Inter-peak interval ratio Note that the goal inter-peak ratio for the 1:1 and 1:2 coordination task would be 1.00 and 2.00, respectively, with no variability. The analysis indicated a main effect of task, F(1,14) = 37,921.60, p < 0.01, η 2 p = 0.99, with the ratio for the 1:1 coordination task, as expected, lower than for the 1:2 coordination task (Fig. 6a). This indicates that based on point estimates that the participants were very effective in producing the respective goal coordination pattern.
Phase angle slope ratio As with the inter-peak interval ratio, the goal phase angle slope for the 1:1 and 1:2 coordination task would be 1.00 and 2.00, respectively, with no variability. The analysis indicated a main effect of task, F(1,14) = 20,421.54, p < 0.01, η 2 p = 0.99, with the ratio for the 1:1 coordination task, as expected, lower than for the 1:2 coordination task (Fig. 6e). This indicates that based on regression analysis of the continuous left and right limb phase angle data that the participants were very effective in producing the respective goal coordination pattern.
Unimanual analyses
Inter-peak interval The analysis indicated main effects of limb, F(1,14) = 56.68, p < 0.01, η 2 p = 0.80, and task, F(1,14) = 14.05, p < 0.01, η 2 p = 0.50. In addition, the task × limb interaction, F(1,14) = 56.50, p < 0.01, η 2 p = 0.80, was significant. Simple main effects analysis indicated similar inter-peak intervals for the left and the right limb for the 1:1 task, but slower inter-peak intervals for the left limb than the right limb for the 1:2 task (Fig. 6b). All other main affects and interactions were not significant.
STD inter-peak interval The analysis indicated a main effect of limb, F(1,14) = 9.35, p < 0.01, η 2 p = 0.40. In addition, the task × limb interaction, F(1,14) = 5.77, p < 0.05, η 2 p = 0.29, was significant. Simple main effects analysis indicated similar STD inter-peak intervals for the left and the right limb for the 1:1 task, but larger STD inter-peak intervals for the left limb than the right limb for the 1:2 task (Fig. 6c). All other main affects and interactions were not significant.
Peak force None of the main effects or interaction were significant. Note that participants’ in both conditions and with both limbs were able to produce the maximum force required by the template (≈15 N) (Fig. 6d).
Phase angle slope However, the main effect of limb F(1,14) = 96.04, p < 0.01, η 2 p = 0.87, and the task × limb interaction, F(1,14) = 93.75, p < 0.01, η 2 p = 0.87, were significant. Simple main effects analysis indicated similar slopes for the left and right limbs for the 1:1 coordination task. As expected, the slope for left limb was lower than for the right limb in the 1:2 coordination task (Fig. 6f).
Mean force All main effects and interactions were not significant. Note that participant’s in both tasks and with both limbs were able to produce the required force (≈7.5 N) (Fig. 6g).
Force harmonicity The analysis indicated main effects of limb, F(1,14) = 38.24, p < 0.01, η 2 p = 0.73, and task, F(1,14) = 35.53, p < 0.01, η 2 p = 0.71. In addition, the task × limb interaction, F(1,14) = 35.35, p < 0.01, η 2 p = 0.71, was significant. Simple main effects analysis indicated similar force harmonicity for the left and the right limb for the 1:1 task, but lower force harmonicity for the left (slower moving) limb than the right (faster moving) limb for the 1:2 task (Fig. 6h). This indicates more adjustments, hesitations, and/or perturbations to the left limb than the right in the 1:2 task, but not the 1:1 task. All other main effects and interactions were not significant.
Discussion
Experiment 3 was designed to determine whether individuals can effectively produce bimanual 1:1 and 1:2 force production coordination patterns when provided integrated feedback in the form of Lissajous information and to determine if the force time series for the 1:2 task indicated distortions in the patterns of forces that could be attributed to neural signals sent to the homologous muscles of the contralateral limb. The point estimates using inter-peak interval ratio and the continuous measure using the phase angle slope ratio indicated very effective performance of the 1:2 bimanual coordination task after only 4 min of practice. This result is quite impressive when compared to previous research that has demonstrated tasks which require frequency relationships other than 1:1 are difficult and some ratios are nearly impossible to perform without extensive practice (e.g., Byblow & Goodman, 1994; Peper et al., 1995a, b; Zanone & Kelso, 1992). Research has indicated that continuous 1:2 tasks, such as the one used in the current experiment, pose difficult challenges for the CNS (e.g., Puttemans et al., 2005; Summers et al., 2002; Swinnen et al., 1997).
The results of the current experiment, however, is similar to that observed in earlier work with reciprocal and circling motion demonstrating a wide range of bimanual coordination patterns (e.g., 1:2, 4:3; 5:3), can be effectively performed following relatively little practice when salient feedback (e.g., Lissajous information) and attentional distractors (i.e., no metronome and no vision of the limbs) were reduced (Boyles et al., 2012; Kovacs et al., 2010a, b; Kovacs & Shea, 2010, 2011). However, it is important to note that these experiments were conducted in near frictionless environments. The participants’ ability to coordinate this pattern of force with the aid of Lissajous plots is especially informative given that the increased force required to produce the desired bimanual coordination pattern resulted in relatively consistent and identifiable distortions in the left limb force time series that appeared to be related to the forces produced in the right (contralateral) limb. This result is consistent with previous research that has demonstrated the influence of one limbs movement on the contralateral limb occurred at the point of muscle activation (Kennedy, Wang, & Shea, 2013). Furthermore, these distortions resulted in relatively small but identifiable increases or decreases in forces produced by the left limb that coincided with the increased or decreased forces produced by the right limb. These perturbations were especially evident in the force velocity profiles for the left limb and are reflected in the reduced harmonicity values for the left limb. Presumably, if the Lissajous displays were withdrawn these perturbations in the left limb forces could result in unintended phase transitions in the coordination patterns.
The 1:2 results are also consistent with previous research that demonstrated that in right limb dominant individuals, the dominant left hemisphere exerts a stronger influence on the non-dominant left limb than the non-dominant right hemisphere on the dominant right limb (Kagerer et al., 2003; Maki et al., 2008). It should be noted, however, that the influence of the right limb on the left limb forces could be a result of the faster frequency and therefore the greater force velocity changes in the right limb rather than a bias strictly associated with limb dominance. This type of right to left limb influence was not evident in our earlier work when the bimanual coordination task involved movements of the limbs in a relatively frictionless environment (Boyles et al., 2012; Kovacs, Buchanan, & Shea, 2010a, b; Kovacs & Shea, 2010, 2011). However, isolated distortions have been observed in a 1:2 coordination pattern in which participants were required to coordinate hand-held pendulums (Sternad et al., 1999a). Interestingly, the ability to coordinate hand-held pendulums also required increased force production compared to tasks performed in near frictionless environments.
The ability to observe relatively consistent distortions in the force times series for the left limb that could be attributed to the forces produced by the right limb as well as the distortions observed in the displacement trace during the coordination of hand-held pendulums (Sternad et al., 1999a) supports the notion that an increase in the force requirements to produce the desired coordination pattern may allow for a clearer detection and characterization of interference “crosstalk” at the behavioral level.
General discussion
Three experiments were conducted to determine whether the forces generated in one limb affect the forces exhibited by the homologous muscles of the contralateral limb and if so, how and when these forces influenced the coordination dynamics (e.g., facilitate or inhibit, cooperate or compete). The results of Experiment 1 detected significant positive time series cross correlation between the forces produced by each limb. Positive correlations indicate that an increase in force by one limb results in a corresponding increase in force by the contralateral limb while a decrease in force by one limb results in a corresponding decrease in force by the other limb. This relationship between the limbs was observed regardless of which limb was provided feedback. Remarkably, we also observed positive time series cross correlations when the participant was instructed to produce a continuous force with only one limb. However, because the participants contralateral limb was resting against the force transducer we were able to record forces produced by the “uninstructed limb.” What we observed was an increase above baseline and a correlation between these forces with those of the contralateral limb. This result was not expected but the effect was consistent across participants regardless of the limb producing the instructed force, and the magnitude of the associated time series cross correlations in the unimanual conditions were similar to those observed under the bimanual conditions.
In Experiment 2, participants were asked to produce a dynamic pattern of forces with one limb while the contralateral limb produced a constant force. By instructing the participants to track a sine wave pattern of force, we were able to observe whether the correlated activity between the limbs was simply due to a common influence impacting the production of force in both limbs or if the pattern of force produced by one limb was influenced by the pattern of forces produced by the contralateral limb. The results indicated a relatively strong relationship between the forces produced by the limbs with the dynamic pattern of forces produced by one limb systematically altering the forces produced by the contralateral limb. This resulted, as in Experiment 1, in significant time series cross correlations between the forces produced by the two limbs.
Experiment 3 was designed to investigate the bimanual production and coordination of force when both limbs were producing dynamic force patterns. Specifically, Experiment 3 was designed to determine participants’ ability to coordinate 1:1 (symmetric) and 1:2 (asymmetric) rhythmical bimanual force patterns when provided Lissajous feedback. The goal was to not only detect distortions in force during the production of the 1:2 force patterns, but also implicate the forces produced by the contralateral limb to these distortions. The results indicated very effective temporal performance and accurate force performance of the bimanual coordination patterns. However, consistent distortions in the left limb force and force velocity traces were observed for participants in the 1:2 task, but not the 1:1 task. The harmonicity values for the left limb during the 1:2 coordination task also reflected adjustments, hesitations, and/or perturbations in the left limb. Additionally, the results indicated that the observed distortions coincided with the initiation and release of force in the right limb.
Neural crosstalk
The literature on bimanual coordination has brought attention to a coalition of constraints which are manifest when the actions of one limb are different from the actions of the contralateral limb, but not when the actions involve the simultaneous activation of homologous muscles (Swinnen, 2002; Swinnen & Wenderoth, 2004). Indeed, when homologous muscles of the left and right limbs are simultaneously activated to produce mirror image actions there appears to be synergistic and/or coupling effects that stabilize the coordination pattern. However, when the two homologous muscle groups are required to produce different actions the patterns are less stable.
One possible constraint that may in part account for the stability differences and difficulty in producing asymmetric coordination pattern is neural crosstalk (see Swinnen, 2002; Swinnen & Wenderoth, 2004 for reviews). Neural crosstalk is a frequently invoked concept to account for observed interference in bimanual coordination (e.g., Aramaki, Osu, & Sadato, 2010; Buchanan & Ryu, 2012; Cattaert et al., 1999; Kasuga & Nozaki, 2011; Maki et al., 2008; Spijkers & Heuer, 1995; Steglich et al., 1999; Swinnen, 2002; Swinnen & Wenderoth, 2004) and its existence has been implicated in both kinematic (e.g., Cattaert et al., 1999; Kasuga & Nozaki, 2011; Park, Dijkstra, & Sternad, 2013; Spijkers & Heuer, 1995) and neuroimaging (e.g., Aramaki et al. 2006; Aramaki et al., 2010; Houweling et al., 2010; Maki et al., 2008) studies. According to the crosstalk model, two independent motor plans exist for each limb and some fraction of the force command for one limb is diverted to the other limb (Cattaert et al., 1999). Because neural crosstalk conveys the same information to both limbs, it is believed that symmetrical actions are facilitated when the contralateral and ipsilateral signals are integrated while asymmetric actions suffer from perturbations due to conflicting information or partial intermingling of signals controlling the two arms (Kagerer et al., 2003; Marteniuk et al., 1984). The present experiments were designed in an attempt to characterize and quantify this influence using isometric forces.
Indeed, we were able to detect and characterize consistent distortions in the forces produced by one limb that were associated with the forces produced in the contralateral limb. In Experiments 1 and 2, we used time series cross correlation analysis to determine how the forces generated in one limb were related to the forces produced by the contralateral limb and to determine the degree to which the two limbs were constrained to act as a single functional unit. In Experiment 1, 92 % of the unimanual and bimanual trials resulted in significant positive correlations and 100 % of the trials were positively correlated in Experiment 2. A positive correlation between forces indicates that the forces produced in one limb were associated with the forces produced in the contralateral limb. This was even true, when we examined the influence the instructed limb had on the uninstructed limb in the unimanual conditions. The uninstructed limb produced minimal forces that appeared, at least to some degree, entrained to the forces produced by the instructed limb. This result is quite interesting if you consider the body of research examining “mirror movements” during unimanual tasks.
Previous research has identified mirror movements (MM) as involuntary activity of the homologous contralateral limb that are associated with voluntary unilateral actions (e.g., Ridderikhoff, Peper, & Beek, 2005). It is believed that mirror movements occur in the contralateral limb as a result of motor overflow or more specifically, neural crosstalk. Part of the signal to the limb controlling the unimanual action is dispatched to the contralateral limb (Cattaert et al., 1999) causing activity to occur in the uninstructed limb. Typically, MM are detected with electromyographic (EMG) analysis whereby participants are asked to produce a force or movement in one limb and EMG data are collected from the contralateral limb (e.g., Ridderikhoff et al., 2005; Cincotta et al., 2006). MMs have also been detected with force production protocols (e.g., Armatas & Summers, 2001; Armatas, Summers, & Bradshaw, 1996; Uttner et al., 2007). Although we did not collect EMG data the force data from our unimanual trials clearly indicated related forces were produced in the uninstructed contralateral limb. These findings provide observable behavioral evidence that neural crosstalk may be implicated as a contributing factor to the influence one limb has over the other limb.
In Experiment 3 we examined the force velocity and acceleration profiles associated with the production of force by each limb to determine how and when forces generated in one limb influenced the forces produced by the contralateral limb. The results indicated very effective temporal performance of the bimanual coordination patterns. This result was similar to that observed in our earlier work with reciprocal and circling motions, but is especially informative given that the increased forces required to produce the desired bimanual coordination pattern resulted in a consistent and identifiable distortion of the left limb forces that could be attributable to the production of right limb forces. We were not able to detect distortions of the forces produced by the right limb that could be attributable to the forces produced by the left limb. This type of right to left limb influence, which may be attributable to asymmetrical cortical and subcortical crosstalk, was not evident in our earlier work when the bimanual coordination tasks involved movements of the limbs in a relatively frictionless environment or in Experiments 1 and 2 of the current investigation.
Previous research has indicated that the dominant hemisphere exerts a stronger influence on the non-dominant limb than the non-dominant hemisphere does on the dominant limb (Aramaki et al., 2006; Cattaert et al., 1999; Kagerer et al., 2003; Maki et al., 2008). As such, cortical and subcortical crosstalk is expected to be asymmetric (Aramaki et al., 2006). In fact, Cattaert et al. (1999) successfully simulated a neural crosstalk model for bimanual interference predicated on the notion that the left limb receives the attenuated mirror image of the commands sent to the right limb. Their simulation successfully reproduced characteristics associated with asymmetric bimanual circle drawing with increasing movement frequencies resulting in a deterioration of the circular trajectories (Cattaert et al., 1999). Although we did not utilize a circle drawing task, the results of Experiment 3 are consistent with the simulation produced by Cattaert and colleagues. In Experiment 3 we were able to identify distortion of the left limb forces that could be attributable to the production of right limb forces. However, we were not able to detect distortions of the forces produced by the right limb that could be attributable to the forces produced by the left limb. It should be noted, however, that the influence of the right limb on the left limb forces could be a result of the faster frequency and therefore the greater force velocity changes in the right limb rather than a bias strictly associated with limb dominance. The results of Experiments 1 and 2 support such a possibility. In Experiment 1 and 2, we did not detect a limb effect indicating the influence of crosstalk was symmetrical. Although this could possibly be due to the task demands (Krishnan & Jaric, 2010; Monno, Chardenon, Temprado, Zanone, & Laurent, 2000) associated with producing constant forces with one or both limbs compared to producing a pattern of forces with both limbs.
When asymmetric patterns are required for the left and right limbs, an individual must inhibit or compensate for the crosstalk that is dispatched to the contralateral limb (Barral et al. 2006, 2010). If the inhibition or compensation for the crosstalk fails, the interference may challenge the stability of the coordination dynamics. In theory, therefore a failure to inhibit, suppress, or otherwise compensate for, the neural crosstalk may result in unwanted perturbations to the coordinated action (Houweling et al., 2010). Indeed, the observed perturbations observed in the left limb (Experiment 3) essentially pushed the force in the direction of the force concurrently produced by the right limb. This is best observed in Fig. 5g where the controlled decrease in force for the right limb essentially depressed the force produced by the left limb and the increase in force of the right limb counteracts the controlled decrease of force for the left limb. These perturbations were observed not only in the force time series for the left limb (Fig. 5g, red trace) but also in the left limb force velocity profiles (Fig. 5o, green trace), and force velocity profile (Fig. 5p).
Detecting and characterizing crosstalk
Consistent with the notion of neural crosstalk, we were able to detect and characterize consistent perturbations in forces time series produced by one limb that were associated with the forces generated by the contralateral limb. More specifically, the results of all three experiments indicated that forces generated in one limb influenced forces produced by the contralateral such that increases in force in one limb corresponded with increases in force produced by the contralateral limb while decreases in force in one limb correspond with decreases in force in the other limb. These types of distortions were not evident in our earlier investigations using reciprocal and circling motion in near frictionless environments (Boyles et al., 2012; Kovacs et al., 2010a, b; Kovacs & Shea, 2010, 2011). The ability to observe relatively consistent distortions in the force time series for one limb that could be attributed to the forces produced by the contralateral limb supports the notion that an increase in the force requirements to produce the desired coordination pattern may allow for a clearer detection and characterization of crosstalk at the behavioral level. The ability to detect and characterize crosstalk in the behavioral data is an important step in understanding constraints acting on the perceptual-motor system.
Summary
In the present study we employed bimanual force production tasks to examine how force generated in one limb influenced the force produced in the contralateral limbs. The results indicated that forces produced by one limb were significantly correlated to the forces produced by the contralateral limb and the influence was stronger when dynamic patterns of force were produced as opposed to when constant forces were maintained. We also observed this form of coupling when the participant was instructed to produce a constant force with only one limb. The results from the multifrequency task indicated consistent and identifiable distortion of the left limb forces that could be attributable to the production of right limb forces. Additionally, the results indicated that the influence the right limb on the left limb occurred when the right limb initiated or released force. The results of all three experiments support the notion that crosstalk is partially responsible for the stabilities and instabilities associated with bimanual coordination.
References
Aramaki, Y., Honda, M., Okada, T., & Sadato, N. (2006). Neural correlates of the spontaneous phase transition during bimanual coordination. Cerebral Cortex, 16, 1338–1348.
Aramaki, Y., Osu, R., & Sadato, N. (2010). Resource-demanding versus cost-effective bimanual interaction in the brain. Experimental Brain Research, 203, 407–418.
Armatas, C. A., & Summers, J. J. (2001). The influence of task characteristics on the intermanual asymmetry of motor overflow. Journal of Clinical Experimental Neuropsycholology, 23, 557–567.
Armatas, C. A., Summers, J. J., & Bradshaw, J. L. (1996). Handedness and performance variability as factors influencing mirror movement occurrence. Journal of Clinical Experimental Neuropsychology, 18, 823–835.
Barral, J., De Pretto, M., Debû, B., & Hauert, C. A. (2010). Activation and inhibition of bimanual movements in school-aged children. Human Physiology, 36, 47–57.
Barral, J., Debû, B., & Rival, C. (2006). Developmental changes in unimanual and bimanual aiming movements. Developmental Neuropsycholgy, 29, 415–429.
Beets, I. A. M, Gooijers, J., Boisgontier, M. P., Pauwels, L., Coxon, J. P., Wittenberg, G., & Swinnen, S. P. (2014). Reduced neural differentiation between feedback conditions after bimanual coordination with and without augmented feedback. Cerebral Cortex. doi:10.1093/cercor/bhu005
Boyles, J., Panzer, S., & Shea, C. H. (2012). Increasingly complex bimanual multi-frequency coordination patterns are equally easy to perform with on-line relative velocity feedback. Experimental Brain Research, 216, 515–525.
Buchanan, J. J., Park, J. H., & Shea, C. H. (2006). Target width scaling in a repetitive aiming task: switching between cyclical and discrete units of action. Experimental Brain Research, 175, 710–725.
Buchanan, J. J., & Ryu, Y. U. (2012). Scaling movement amplitude: adaptation of timing and amplitude control in a bimanual task. Journal of Motor Behavior, 44, 135–147.
Byblow, W. D., Bysouth-Young, D., Summers, J. J., & Carson, R. G. (1998). Performance asymmetries and coupling dynamics in the acquisition of multifrequency bimanual coordination. Psychologoical Research, 61, 56–70.
Byblow, W. D., & Goodman, D. (1994). Performance asymmetries in multifrequency coordination. Human Movement Science, 13, 147–174.
Cardoso de Oliveira, S. (2002). The neuronal basis of bimanual coordination: recent neurophysiological evidence and functional models. Acta Psychologica, 110, 139–159.
Carson, R. G., Byblow, W. D., Abernethy, B., & Summers, J. J. (1996). The contribution of inherent and incidental constraints to intentional switching between patterns of bimanual coordination. Human Movement Science, 15, 565–589.
Carson, R. G., Riek, C. J., Smethurst, J. F., Lison Parraga, J. F., & Byblow, W. D. (2000). Neuromuscular-skeletal constraints upon the dynamics of unimanual and bimanual coordination. Experimental Brain Research, 131, 196–214.
Cattaert, D., Semjen, A., & Summers, J. J. (1999). Simulating a neural cross-talk model for between-hand interference during bimanual circle drawing. Biological Cybernetics, 81, 343–358.
Cincotta, M., Giovannelli, F., Borgheresi, A., Balestrieri, F., Vanni, P., Ragazzoni, A., & Ziemann, U. (2006). Surface electromyography shows increased mirroring in Parkinson’s disease patients without overt mirror movements. Movement Disorders, 21, 1461–1465.
Cohen, L. (1971). Synchronous bimanual movements performed by homologous and non-homologous muscles. Perceptual Motor Skills, 32, 639–644.
Deutsch, D. (1983). The generation of two isochronous sequences in parallel. Perception and Psychophysics, 34, 331–337.
Diedrichsen, J., Hazeltine, E., Nurss, W. K., & Ivry, R. B. (2003). The role of the corpus callosum in the coupling of bimanual isometric force pulses. Journal of Neurophysiology, 4, 2409–2418.
Franz, E. A., Eliassen, J. C., Ivry, R. B., & Gazzaniga, M. S. (1996). Dissociation of spatial and temporal coupling in the bimanual movements of callosotomy patients. Psychological Science, 7, 306–310.
Guiard, Y. (1987). Asymmetric division of labor in human skilled bimanual action: the kinematic chain as a model. Journal of Motor Behavior, 19, 486–517.
Guiard, Y. (1993). On Fitts’s and Hooke’s laws: simple harmonic movement in upper-limb cyclical aiming. Acta Psychologica, 82, 139–159.
Guiard, Y. (1997). Fitts’ law in the discrete vs cyclical paradigm. Human Movement Science, 16, 97–131.
Hessler, E. E., Gonzales, L. M., & Amazeen, P. G. (2010). Displays that facilitate performance of multifrequency ratios during motor-respiratory coordination. Acta Psychologica, 133, 96–105.
Heuer, H., Kleinsorge, T., Spijkers, W., & Steglich, W. (2001). Static and pha-sic cross-talk effects in discrete bimanual reversal movements. Journal of Motor Behavior, 33, 67–85.
Heuer, H., Spijkers, W., Steglich, C., & Kleinsorge, T. (2002). Parametric coupling and generalized decoupling revealed by concurrent and successive isometric contractions of distal muscles. Acta Psychologica, 111, 205–242.
Hill, A. V. (1970). First and last experiments in muscle mechanics. Cambridge: Cambridge University Press.
Houweling, S., Beek, P. J., & Daffertshofer, A. (2010). Spectral changes of interhemispheric crosstalk during movement instabilities. Cerebral Cortex, 20, 2605–2613.
Hu, X., Loncharich, M., & Newell, K. M. (2011). Visual information interacts with neuromuscular factors in the coordination bimanual isometric force. Experimental Brain Research, 209, 129–138.
Hu, X., & Newell, K. M. (2011a). Adaption to bimanual asymmetric weights in isometric force coordination. Neuroscience Letters, 490, 121–125.
Hu, X., & Newell, K. M. (2011b). Visual information gain and task asymmetry interact in bimanual force coordination and control. Experimental Brain Research, 212, 497–504.
Kagerer, F. A., Summers, J. J., & Semjen, A. (2003). Instabilities during antiphase bimanual movements: are ipsilateral pathways involved? Experimental Brain Research, 151, 489–500.
Kasuga, S., & Nozaki, D. (2011). Cross talk in implicit assignment of error information during bimanual visuomotor learning. Journal of Neurophysiology, 106, 1218–1226.
Kelso, J. A. S. (1995). Dynamic patterns: the self-organization of the brain and behavior. Cambridge: MIT Press.
Kelso, J. A. S., Scholz, J. P., & Schoner, G. (1986). Nonequilibrium phase-transitions in coordinated biological motion; critical fluctuations. Physics Letters A, 118, 279–284.
Kennedy, D. M., Boyle, J. B., Rhee, J., & Shea, C. H. (2014). Rhythmical bimanual force production: homologous and non-homologous muscles. Experimental Brain Research. doi:10.1007/s00221-014-4102-y
Kennedy, D. M., Wang, C., & Shea, C. H. (2013). Reacting while moving: influence of right limb movement on left limb reaction. Experimental Brain Research, 230, 143–152.
Kovacs, A. J., Buchanan, J. J., & Shea, C. H. (2009). Bimanual 1:1 with 90 degrees continuous relative phase: difficult or easy. Experimental Brain Research, 193, 129–136.
Kovacs, A. J., Buchanan, J. J., & Shea, C. H. (2010a). Impossible is nothing: 5:3 and 4:3 multi-frequency bimanual coordination. Experimental Brain Research, 201, 249–259.
Kovacs, A. J., Buchanan, J. J., & Shea, C. H. (2010b). Perceptual and attentional influences on continuous 2:1 and 3:2 multi-frequency bimanual coordination. Journal of Experimental Psychology: Human Perception and Performance, 36, 936–954.
Kovacs, A. J., & Shea, C. H. (2010). Amplitude differences, spatial assimilation, and integrated feedback in bimanual coordination. Experimental Brain Research, 202, 519–525.
Kovacs, A. J., & Shea, C. H. (2011). The learning of 90 degrees continuous relative phase with and without Lissajous feedback: external and internally generated bimanual coordination. Acta Psychologica, 136, 311–320.
Krishnan, V., & Jaric, S. (2010). Effects of task complexity on coordination of inter-limb and within-limb forces in static bimanual manipulation. Motor Control, 230, 528–544.
Latash, M. L. (2010). Motor synergies and the equilibrium-point hypothesis. Motor Control, 14, 294–322.
Latash, M. L. (2012). The bliss (not the problem) of motor abundance (not redundancy). Experimental Brain Research, 217, 1–5.
Levin, O., Suy, E., Huybrechts, J., Vangheluwe, S., & Swinnen, S. P. (2004). Bimanual coordination involving homologous and heterologous joint combinations: when lower stability is associated with higher flexibility. Behavioral Brain Research, 152, 437–445.
Maki, Y., Wong, K. F. K., Sugiura, M., Ozaki, T., & Sadato, N. (2008). Asymmetric control mechanisms of bimanual coordination: an application of directed connectivity analysis to kinematic and functional MRI data. Neuroimage, 42, 1295–1304.
Marteniuk, R. G., MacKenzie, C. L., & Baba, D. M. (1984). Bimanual movement control: information processing and interaction effects. Quarterly Journal of Experimental Psychology, 37, 335–365.
Monno, A., Chardenon, A., Temprado, J. J., Zanone, P. G., & Laurent, M. (2000). Effects of attention on phase transitions between bimanual coordination patterns: a behavioral and cost analysis in humans. Neuroscience Letters, 283, 93–96.
Murian, A., Deschamps, T., & Temprado, J. J. (2008). Effects of force production and trial duration on bimanual performance and attentional demands in a rhythmic coordination task. Motor Control, 12, 21–37.
Park, S., Dijkstra, T. M. H., & Sternad, D. (2013). Learning to never forget—time scales and specificity of long-term memory of a motor skill. Frontiers in Computational Neuroscience, 7, 1–13.
Peper, C. E., Beek, P. J., & van Wieringen, P. C. W. (1995a). Coupling strength in tapping a 2/3 polyrhythm. Human Movement Science, 14, 217–245.
Peper, C. E., Beek, P. J., & van Wieringen, P. C. W. (1995b). Frequency-induced phase-transitions in bimanual tapping. Biological Cybernetics, 73, 301–309.
Peper, C. E., Beek, P. J., & Vanwieringen, P. C. W. (1995c). Multifrequency coordination in bimanual tapping—asymmetrical coupling and signs of supercriticality. Journal of Experimental Psychology-Human Perception and Performance, 21, 1117–1138.
Puttemans, V., Wenderoth, N., & Swinnen, S. P. (2005). Changes in brain activation during the acquisition of a multifrequency bimanual coordination task: from the cognitive stage to advanced levels of automaticity. The Journal of Neuroscience, 25, 4270–4278.
Ridderikhoff, A., Peper, C. L., & Beek, P. J. (2005). Unraveling interlimb interactions underlying bimanual coordination. Journal Neurophysiology, 94, 3112–3125.
Riek, S., Carson, R. G., & Byblow, W. D. (1992). Spatial and muscular dependencies in bimanual coordination. Journal of Human Movement Studies, 23, 251–265.
Sainburg, R. L. (2010). Lateralization of goal-directed movement. Vision and goal-directed movement (pp. 219–288). Champaign: Human Kinetics.
Scholz, J. P., & Kelso, J. A. S. (1989). A quantitative approach to understanding the formation and change of coordinated movement patterns. Journal of Motor Behavior, 21, 122–144.
Semjen, A., & Summers, J. J. (2002). Timing goals in bimanual coordination. Quarterly Journal of Experimental Psychology A, Human Experimental Psychology, 55, 155–171.
Semjen, A., Summers, J. J., & Cattaert, D. (1995). Hand coordination in bimanual circle drawing. Journal of Experimental Psychology-Human Perception and Performance, 21, 1139–1157.
Serrien, D. (2009). Interactions between new and pre-existing dynamics in bimanual movement control. Experimental Brain Research, 197, 269–278.
Serrien, D., & Swinnen, S. P. (1997). Coordination constraints induced by effector combination under isofrequency and multifrequency conditions. Journal of Experimental Psychology-Human Perception and Performance, 23, 1493–1510.
Shapkova, E. Y., Shapkova, A. L., Goodman, S. R., Zatsiorsky, V. M., & Latash, M. L. (2008). Do synergies decrease force variability? A study of single-finger and multi-finger force production. Experimental Brain Research, 188, 411–425.
Sherwood, D. E. (1994). Hand preference, practice order, and spatial assimilations in rapid bimanual movements. Journal of Motor Behavior, 26, 123–134.
Sisti, H. M., Geurts, M., Clerckx, R., Gooijers, J., Coxon, J. P., Heitger, M. H., & Swinnen, S. P. (2011). Testing multiple coordination constraints with a novel bimanual visuomotor task. PLoS ONE, 6, e23619.
Spijkers, W., & Heuer, H. (1995). Structural constraints on the performance of symmetrical bimanual movements with different amplitudes. Quarterly Journal of Experimental Psychology Section a-Human Experimental Psychology, 48, 716–740.
Steglich, C., Heuer, H., Spijkers, W., & Kleinsorge, T. (1999). Bimanual coupling during the specification of isometric forces. Experimental Brain Research, 129, 302–316.
Sternad, D., Turvey, M. T., & Saltzman, E. L. (1999a). Dynamics of 1:2 coordination: temporal scaling, latent 1:1, and bistability. Journal of Motor Behavior, 31, 236–247.
Sternad, D., Turvey, M. T., & Saltzman, E. L. (1999b). Dynamics of 1:2 coordination: sources of symmetry breaking. Journal of Motor Behavior, 31, 224–235.
Sternad, D., Turvey, M. T., & Saltzman, E. L. (1999c). Dynamics of 1:2 coordination: generalizing relative phase to n:m rhythms. Journal of Motor Behavior, 31, 207–223.
Stone, K. D., Bryant, D. C., & Gonzalez, C. L. R. (2013). Hand use for grasping in a bimanual task: evidence for different roles? Experimental Brain Research, 224, 455–467.
Summers, J. J., Davis, A. S., & Byblow, W. D. (2002). The acquisition of bimanual coordination is mediated by anisotropic coupling between the hands. Human Movement Science, 21, 699–721.
Summers, J. J., Maeder, S., Hiraga, C. Y., & Alexander, J. R. (2008). Coordination dynamics and attentional costs of continuous and discontinuous bimanual circle drawing movements. Human Movement Science, 27, 823–837.
Summers, J. J., Todd, J. A., & Kim, Y. H. (1993). The influence of perceptual and motor factors on bimanual coordination in a polyrhythmic tapping task. Psychological Research, 55, 107–115.
Swinnen, S. P. (2002). Intermanual coordination: from behavioural principles to neural-network interactions. Nature Review: Neuroscience, 3, 348–359.
Swinnen, S. P., Dounskaia, N., & Duysens, J. (2002). Patterns of bimanual interference reveal movement encoding within a radial egocentric reference frame. Journal of Cognitive Neuroscience, 14, 463–471.
Swinnen, S. P., Dounskaia, N., Levin, O., & Duysens, J. (2001). Constraints during bimanual coordination: the role of direction in relation to amplitude and force requirements. Behavioral Brain Research, 123, 201–218.
Swinnen, S. P., Dounskaia, N., Walter, C. B., & Serrien, D. J. (1997). Preferred and induced coordination modes during the acquisition of bimanual movements with a 2:1 frequency ratio. Journal of Experimental Psychology: Human Perception and Performance, 23, 1087–1110.
Swinnen, S. P., & Wenderoth, N. (2004). Two hands, one brain: cognitive neuroscience of bimanual skill. Trends in Cognitive Sciences, 8, 18–25.
Temprado, J. J., Chardenon, A., & Laurent, M. (2001). Interplay of biomechanical and neuromuscular constraints on pattern stability and attentional demands in a bimanual coordination task in human subjects. Neuroscience Letters, 303, 127–131.
Temprado, J. J., Zanone, P. G., Monno, A., & Laurent, M. (1999). Attentional load associated with performing and stabilizing preferred bimanual patterns. Journal of Experimental Psychology: Human Perception and Performance, 25, 1579–1594.
Therrien, A. S., Lyons, J., & Balasubramaniam, R. (2013). Continuous theta-burst stimulation to primary motor cortex reveals asymmetric compensation for sensory attenuation in bimanual repetitive force production. Journal of Neurophysiology, 110, 872–882.
Treffner, P. J., & Turvey, M. T. (1993). Resonance constraints on rhythmic movement. Journal of Experimental Psychology: Human Perception and Performance, 19, 1221–1237.
Uttner, I., Kraft, E., Nowak, D. A., Muller, F., Philipp, J., Zierdt, A., & Hermsdorfer, J. (2007). Mirror movements and the role of handedness: isometric grip forces changes. Motor Control, 11, 16–28.
Zanone, P. G., & Kelso, J. A. S. (1992). The evolution of behavioral attractors with learning: nonequilibrium phase transitions. Journal of Experimental Psychology: Human Perception and Performance, 18, 403–421.
Acknowledgments
This work was supported by student research grants from the Huffines Institute and College of Education and Human Development, Texas A&M University to D. M. Kennedy.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Kennedy, D.M., Boyle, J.B., Wang, C. et al. Bimanual force control: cooperation and interference?. Psychological Research 80, 34–54 (2016). https://doi.org/10.1007/s00426-014-0637-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00426-014-0637-6