
Abstract
Bioturbation of reef sediments aerates the upper sediment layers and releases organic material to benthic communities. Despite being the larger and more conspicuous bioturbators on coral reefs, the value of holothurians (sea cucumbers) to reef ecosystems is less often attributed to their ecosystem services than their value for fisheries. This may be because they are considered to have an insignificant effect on reef health relative to other animals. Here, we ground-truthed remote sensing data obtained from drone and satellite imagery to estimate the bioturbation rates of holothurians across the 19 km2 Heron Island Reef in Queensland, Australia. Ex situ bioturbation rates of the most abundant holothurian, Holothuria atra, were assessed during 24-h feeding experiments. Using density measurements of holothurians across reef flat zones in a 27,000 m2 map produced from drone imagery, we extrapolated bioturbation across the reef using satellite remote sensing data. Individual H. atra were estimated to produce approximately 14 kg of bioturbated sediment per year. On a reef scale (excluding the reef lagoon) and accounting for varying densities of holothurians across different reef zones, total bioturbation from holothurians at Heron Reef was estimated at over 64,000 metric tonnes per year, slightly more than the mass of five Eiffel Towers. These results highlight the scale of structural and biochemical impacts that holothurians have on reef flats and their importance to ecosystem functioning and services. Management of these animals on reefs is imperative as overharvesting would likely cause substantial negative effects on sedimentary ecosystems and their biogeochemistry in corals reefs.
Graphical abstract

Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Holothurians, commonly known as sea cucumbers, are globally one of the most conspicuous organisms associated with coral reefs. These animals are exploited in bêche-de-mer fisheries, and their high value in Asian markets has encouraged global overfishing and associated declines (Conand 2004; Anderson et al. 2011; Purcell et al. 2013). Such declines are concerning as holothurian populations can take decades to recover from over-exploitation (Purcell 2010). Issues around over-exploitation of wild populations have led to research targeted at developing holothurian aquaculture (Han et al. 2016) or to approaches for improved management of their fisheries (Friedman et al. 2011; Plagányi et al. 2015). The intrinsic value of holothurians to the reef ecosystem itself, however, is less often acknowledged as a reason for concern or management. As bioturbators, holothurians offer ecosystem services that increase local productivity and may mitigate some of the impacts of climate change.
Bioturbation of sediments by holothurians releases nutrients trapped in the sediments to benthic ecosystems (Uthicke 2001a). While high densities of holothurians can reduce microalgal production (Uthicke 1999), availability of nutrients, such as ammonium released by holothurians feeding at natural densities, can enhance the growth of benthic algae (Uthicke 2001b), increasing the gross productivity of benthic reef communities (Uthicke and Klumpp 1998). This may be increasingly important as coral reefs degrade and shift to more algal-dominated systems (Hughes et al. 2003). Sediment digestion by holothurians may be responsible for up to 50% of the dissolution of calcium carbonate in reef systems (Schneider et al. 2011), an important process as the majority of calcium carbonate on coral reefs is stored in sediment (Gattuso et al. 1998). As suggested by others (Schneider et al. 2011, 2013; Wolfe et al. 2018), this process may also facilitate the growth of scleractinian corals, which are critical reef builders. Holothurian bioturbation also reduces stratification and nutrification of sediments (İşgören-Emiroğlu and Günay 2007) and can directly increase oxygen levels in the sediment (Hammond 1982). The ecological role of holothurians as bioturbators in reef environments is thus pivotal in facilitating the availability of nutrients and oxygen for other organisms. The scale of this bioturbation of sediments by holothurians, however, remains unclear for coral reefs.
Understanding the importance of holothurians to the benthic systems of coral reefs first necessitates a quantitative analysis of their bioturbation rates and an understanding of the scale at which bioturbation occurs in the system. Holothurians are unevenly distributed in reef systems (Tuya et al. 2006), which makes it difficult to determine their density at a reef scale. While previous studies have attempted to quantify the scale of bioturbation by holothurians in reef systems (Uthicke 1999; Wolfe and Byrne 2017; Hammond et al. 2020), these extrapolations were based on relatively small transect areas (100 m2) and small holothurian sample sizes (n = 12) that may not represent the wider heterogeneity of patterns found on reefs. Those studies that quantified distribution and abundance of holothurians typically used line transects or manta surveys (Uthicke and Benzie 2001a; Guzman and Guevara 2002; Friedman et al. 2011), which also have limitations.
The recent wide adoption of drones or unoccupied aerial vehicles (UAVs) (Anderson and Gaston 2013) for marine research provides a means to accurately map the distribution and behavior of many organisms in shallow aquatic environments (Raoult and Gaston 2018; Raoult et al. 2018), including holothurians that typically have high contrast against pale sediments. However, drones have never been used to assess holothurian abundance and densities, so there may be concerns such an approach would not reflect data obtained via other methods.
This study estimated the bioturbation rate of holothurians across a reef flat using drones and upscaled the estimation to an entire reef scale (minus lagoon) using geomorphic zones classified from satellite imagery. To achieve this, we assessed holothurian bioturbation rates ex situ for the dominant holothurian in Heron Reef. A proof-of-concept study was done to assess the accuracy of drones to measure holothurian abundance against traditional in-water line transect methods. Drone imagery was then digitized to determine holothurian densities in different reef geomorphic zones. Density patterns and their associated bioturbation rates were then extrapolated to the entire reef using classified satellite imagery. The methods developed here and the results from this study will elucidate the scale of effect that holothurians have on coral reefs and facilitate more accurate estimations of their ecological impacts and loss from fisheries.
Methods
Study site
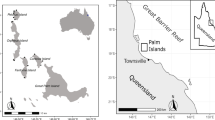
Field surveys were conducted at Heron Reef on the southern end of the Great Barrier Reef (S 23.4423°, E 151.9107°) in September 2016 and February 2019 (Fig. 1). Heron Reef falls under different management zones including a Conservation Park, a Marine National Park, a Public Appreciation Area and a Scientific Research Zone (GBRMPA 2003). Unlike the northern two-thirds of the Great Barrier Reef, this reef was relatively unscathed in the 2016 and 2017 global bleaching events (Hughes et al. 2017, 2018), and the condition of the reef was generally considered healthy at the time of sampling. Holothurian densities inhabiting Heron Reef are considered representative of healthy reef environments in the Pacific, given the reef’s protected status and the lack of broad-scale impacts on this reef.

Map of Heron Reef a, b showing inner and outer reef flat geomorphic zones. c shows the placement of the eight 30-m-long snorkel and drone transects used for the proof of concept study c and the drone survey site (red outline) with the seven virtual transects used to speed digitizing. d shows imagery from one of the proof of concept transects. e shows holothurians digitized from the drone imagery
Bioturbation experiment
Holothuria atra is the most common species of holothurian on Heron Reef flat (Williamson et al. 2017). This species is often found adjacent to reef bommies, on open sediment in shallow reef flat environments (Raoult et al. 2016) and in the lagoon (Madin et al. 2019). It is considered responsible for a substantial portion of bioturbation across Heron Reef. Feeding rates of H. atra are considered fairly constant over days to seasons (Uthicke 1999; Mangion et al. 2004). To assess the productivity of H. atra, a 24-h feeding experiment was run using flow-through outdoor aquaria at Heron Island Research Station. Individual flow-through aquaria (300 × 300 × 300 mm) were set up to receive a constant flow rate (1 L min−1) of sand-filtered seawater pumped directly from the adjacent reef. Twenty-seven holothurians were then collected from various locations on the inner to outer reef flat from the southern side of the island within the Scientific Research Zone, along with ~ 2 kg of the sediment on which they resided. This area is representative of similar habitat, which comprises 58% of Heron Reef (Fig. 1). Individuals were gently transferred from the reef to the aquaria in buckets filled with seawater to reduce stress that may affect feeding rates. Collectors were careful not to handle the holothurians more than necessary. Each 2 kg of sediment was carefully placed in an aquarium so that the upper surface of the sediment remained as upright as possible and allowed to settle for 10 min. After this, the holothurian associated with that sediment was carefully added. A light shade cloth was placed over all aquaria to mimic light penetration at their natural depth.
Individuals were held for 24 h and their fecal pellets collected every three hours as per Uthicke et al. (1999). Pellets were dried at 60 °C for 24 h and then weighed to the nearest milligram. The total amount of dry fecal matter produced after 24 h was assessed by adding all fecal collections per individual and averaging the data.
Proof-of-concept drone-based aerial holothurian counts
A proof-of-concept study was done to validate the use of drone imagery for holothurian counts. To assess the difference between holothurian counts acquired in water via snorkel compared to drone imagery, eight 30-m transect tapes approximately 50 m apart were deployed. Tapes were oriented perpendicular to the observed geomorphic zonation, with four validation transects each in the inner and outer reef flat zones (Fig. 1c). Two observers snorkelled along each validation transect and counted the number of holothurians within a one-meter distance either side of each transect tape. The observers then changed sides and travelled back down the transect, repeating the counts. The average of the two counts per observer was used.
Using a pre-determined flight path, we flew a DJI Phantom 4 Pro with a standard RGB camera over the survey area containing the eight validation transect tapes. We used a flight altitude of 20 m to ensure each transect tape was visible, and to achieve the spatial detail required to identify as many holothurians as possible. We used an overlap of 85% and sidelap between flight lines of 75%, taking care to fly at low tide in the afternoon to avoid sunglint and specular reflection at the water’s surface (Joyce et al. 2019).
Orthomosaics of the region were created using Pix 4D from the resultant photos. Mosaics of the eight transect tapes were then manually digitized with a 1-m buffer either side of the transect to extract imagery co-incident with the in-water counts. Three observers then independently and manually digitized the number of holothurians within each image transect. The difference (if any) between in-water counts and drone counts was then assessed with a linear mixed model using the lme4 package (Bates et al. 2014) in R V. 3.4.4 (Team 2013). The model was designed with holothurian density (individuals per m2) as the response variable, the method (drone or in-water) as the independent variable with an interaction with geomorphic zone and transect nested in zone as a random factor. Including observer as a random factor would have been ideal (accounting for between-observer variation), but as there were only two in-water observers, this was not possible. Moreover, some variation between observers for both methods is to be expected and should not prevent comparison between the two methods. If any significant effects were identified, the scale of the effect was determined using least-square means using the package emmeans (Lenth et al. 2018).
Estimating holothurian density across a large ‘reef-scale’ drone survey
The proof of concept showed that holothurians could be counted using drone-based aerial surveys to provide an (albeit conservative) estimate of abundance (see results). To apply the method over a larger area, an Aeronavics Bot Solo drone fitted with a Sony a7R DSLR camera (36 megapixel, pancake lens) was used to survey a typical cross-reef study site on the southern reef at Heron Island at an altitude of 60 m (Fig. 1c). Pix 4D was used to mosaic drone images and create an orthomosaic covering an area of 2.73 ha with a ground sampling distance (pixel size) of 0.8 cm.
The study site incorporated two geomorphic zones: the inner reef flat and the outer reef flat. The geomorphic zones were defined based on the classification of Dove satellite imagery acquired in January 2018, available in the Allen Coral Atlas (Kennedy et al. 2020). Given that geomorphic zones are defined primarily by their location and level of exposure to physical processes (Hopley et al. 2007), it is unlikely that the zones have changed in the 15 months between our 2016 drone survey and the capture of the satellite imagery used to define them here. We were unable to sample the lagoon or reef crest zones as they were beyond the range of the drone from the island and we did not have access to a boat at the time of survey.
ArcGIS 10.5.1 was used to digitize each visible holothurian in the orthomosaic. This was manual process is time-consuming, so we also assessed whether accurate counts could be achieved by digitizing only a subset of the imagery. To do this, “virtual transects” were placed across the drone orthomosaic. The study area was divided into seven 2-m-wide ‘virtual transects’ 10 m apart running the length of the study area (~ 350 m) (Fig. 1c). The number of holothurians within each geomorphic zone along these transects was calculated and divided by the area of each geomorphic zone in the transect to give a density of holothurians per m2 in each geomorphic zone. These results were compared to those obtained by manually digitizing holothurians across the entire study area, and the difference was negligible (Supplementary 1).
Upscaling holothurian densities and bioturbation rates across Heron Island Reef
Holothurian densities recorded in inner and outer reef flat geomorphic zones of the ‘reef-scale’ drone study area were upscaled using the total area of these zones across Heron Reef (Kennedy et al. 2020). This accounted for a total area of 1682 ha, or 57% of Heron Reef. Bioturbation rates based on the 24-h experiments for H. atra were calculated for densities of holothurians in each of the two geomorphic zones within the entire reef using the following formula:
where \(B\) represents the total holothurian bioturbation rate across Heron Reef in kg per year, \(A\) the area of a geomorphic zone \(n\) in m2, \(D\) the density of holothurians per m2 in geomorphic zone \(n\), and \(P\) the mean annual holothurian bioturbation rate in kg year−1. To account for any biases identified with drone counts relative to in-water counts, the densities of holothurians identified in the drone survey were scaled using the mean differences identified with the linear mixed model. To produce a conservative estimate for total bioturbation rates that includes the uncertainty identified in most of these values, total bioturbation across the reef was estimated in a Monte-Carlo-Markov-Chain framework using a custom R script (Supplementary 2) with 106 runs. These measures of uncertainty included standard deviations around the mean counts of holothurians in the larger survey (estimated from the coefficient of variation of the in-water counts), the standard deviation in bioturbation rates from the feeding experiments, and the standard deviation around the mean difference between drone and in-water counts for the reef flat.
Results
Bioturbation experiment
H. atra produced 38.24 ± 18.82 g (mean ± SD) over the 24-h period. Upscaling to an annual production of dry fecal matter per individual, one H. atra on Heron Reef was estimated (mean ± SD) to produce 13.96 ± 6.87 kg yr−1.
Proof-of-concept of drone-based aerial holothurian counts
A total of 29 paired in-water counts by two observers along the same transect at the same time were conducted. The highest number of holothurians counted on any transect was 37, and the lowest was zero. Paired in-water counts showed good, but not perfect, alignment between observers, with less than 8% discrepancy between counts (mean difference between observers 1.4).
When comparing the two methods (in-water and drone), densities of holothurians counted from drone imagery were significantly lower (df = 173, F = 25.8, p < 0.001) by 0.07 ± 0.01 (estimate ± S.E.) holothurians per m2 than those in-water (Fig. 2). Both methods showed that the inner reef flat had significantly higher holothurian densities (df = 16, F = 9.2, p = 0.023), approximately three times more than those measured in the outer reef flat. Tukey’s HSD post hoc tests found significant differences between drone and in-water densities counted in the inner reef flat (estimate = − 0.13 ± 0.02, df = 73, t = − 6.4, p < 0.001) but not between both methods in the outer reef flat (df = 73, t = − 0.8, p = 0.82). The marginal R2 for this model was 0.55, with the conditional R2 (including the variance explained by the random factor) of 0.87, suggesting our model explained nearly 90% of the variation in our data.

Comparison of snorkeler-based transects to drone counts across the same transects
Estimates of holothurian density across a large ‘reef scale’ drone survey
The total area surveyed in the reef scale drone orthomosaic was 27,348 m2. The spatial pattern of holothurian density was similar in the larger drone survey area to that found in the proof-of-concept study and the paired in-water survey transects (Fig. 2), with holothurian densities approximately 40% higher in the inner reef (0.2 per m2) relative to the outer reef (0.14 per m2) (Table 1).
Upscaling holothurian densities and bioturbation rates across Heron Island Reef
The total area of the two geomorphic zones across Heron Reef was 16.8 km2, as determined from satellite imagery (Table 1). Excluding the reef lagoon that was not surveyed, the mean rate of total holothurian bioturbation across Heron Reef as determined from Monte-Carlo-Markov-Chain was estimated at over 63,970 ± 4168 metric tons per year (mean ± S.E) (Fig. 3).

Posterior density distribution of modeled total bioturbation per year by holothurians across the inner and outer reef geomorphic zones of Heron Reef
Discussion
This research shows that bioturbation by holothurians is a substantial contributor to sediment reworking on Heron Island Reef. Excluding the reef lagoon where holothurians also occur, holothurians were found to produce a conservative estimate of over 64,000 metric tonnes of bioturbated sediment per year across Heron Reef, or approximately 3800 tonnes km−2 y−1. Since holothurian densities are very sensitive to overfishing and population recovery is slow (Uthicke and Benzie 2001a; Uthicke et al. 2004), overexploitation of holothurians is likely to have long-term effects on coral reef sediment communities and the amount of organic carbon available in the water column for nearby organisms. In the current context of anthropogenic pressures on coral reefs (Bellwood et al. 2019), use of remote sensing techniques offers a means to rapidly assess densities of holothurians in shallow reef habitats to facilitate more accurate and targeted management decisions.
Measured bioturbation rates
Our results suggest the total amount of sediment bioturbated by holothurians is over 3800 tonnes km−2 y−1. This value is lower than the 4600 tonnes km−2 y−1 estimated to be bioturbated by H. atra by Uthicke (1999). Uthicke (1999) and others (Yamanouchi 1939) documented bioturbation rates of 67 and 86 g day−1 per individual for H. atra, respectively. Our study, and that of Klinger et al. (1994), documented bioturbation rates of 38 and 11 g day−1 per individual, respectively. Differences in these rates could be due to a selection of larger H. atra in the Uthicke (1999) and Yamanouchi (1939) studies, but this is difficult to tell as accurately weighing and measuring holothurians is problematic due to their ability to extend their bodies and hold varying amounts of fluid. Alternatively, our lower bioturbation rate may have been due to differences in our experimental design. While other studies often allow an acclimation period of approximately 4 h in aquaria prior to the start of their bioturbation experiments for the animals to settle (e.g., Uthicke 1999), our measure of bioturbation started as soon as the animals entered the aquaria. We did not run the experiment over longer timescales due to concerns that the holothurians would consume all the palatable sediment and adjust their feeding rates accordingly. However, this could have caused a reduction in bioturbation in the first couple of hours. Our study thus provides a very conservative bioturbation rate for H. atra over the 24-h experiment.
Environmental parameters such as in water temperature between studies could also partly explain differences in bioturbation rates. Mean seawater temperatures were reported as 27 °C for Uthicke (1999)’s Lizard Island study and 29 °C for Yamanouchi’s (1939) Palao Island study but 24 °C at Heron Island for this study and Klinger et al.’s (1994) research. Water temperature affects holothurian metabolism and overall performance, including bioturbation rates (Fraser et al. 2004; Wheeling et al. 2007; Schiell and Knott 2010). Differences in bioturbation rates between studies could also be caused by differences in organic matter in the sediments that make the sediments more or less palatable (Hammond et al. 2020). Rates of bioturbation do not only differ within studies on H. atra but also between species and over seasons (Wolfe and Byrne 2017) and potentially between years (Schiell and Knott 2010). Such variability in rates of bioturbation means that it is difficult to make a ubiquitous and conclusive statement on the amount of sediment reworked by holothurians across coral reefs. We advocate that studies should continue to assess bioturbation on a reef to reef and species-specific basis to account for such variability. Regardless of the mechanism, we are confident in our bioturbation rates for H. atra in this study due to the high number of individuals assessed and the continual monitoring of feeding rates for 24 h. Bioturbation rates from this study could be considered conservative and highlight the necessity for assessing rates at specific sites before extrapolation.
Monitoring holothurian populations with drones
Assessing the abundance and diversity of holothurians from drone imagery produced results lower than those recorded in situ with traditional methods. If the average in-water transect counts were upscaled, the total count of holothurians would have been 5,132,738 across the inner and outer reef flats at Heron Reef (Table 1). This is 41% higher than the total counted using the drone method (3,033,733 individuals, noting that these counts are not directly comparable as the reef-scale drone survey was conducted at a different time to the in-water transects).
The lower detection of holothurians by drone is contrary to other studies assessing count data of fauna via drones that generally find drones to detect higher numbers of organisms (Hodgson et al. 2018). However, drones cannot survey under coral or rock overhangs like a snorkeler can and the proclivity of some holothurians to coat themselves in sediment also hinders their detection in drone imagery. These factors likely led to the under-counts of holothurians by drones relative to snorkelers in the proof of concept transects. Distortion effects of the water column can also make visibility challenging in drone imagery but can be mitigated by flying at low altitude, using polarizing filters, choosing the lowest possible tide and selecting calm weather conditions to avoid ripples. Nevertheless, in shallow reefs, drones can cover much larger areas than traditional in-water monitoring and produce data that are re-examinable (Joyce et al. 2019). Holothurian monitoring programs could use this approach to produce assessments covering larger areas at a faster rate than traditional approaches. provided the under-estimation of holothurian abundance from drones is accounted for as we have here using a Monte-Carlo-Markov-Chain framework to incorporate measures of uncertainty in the detection of holothurians from drone imagery and the bioturbation rate. The most time-consuming element of the method is the manual digitization of each holothurian from the imagery. Our ‘virtual transects’ yielded almost identical densities to those calculated by digitizing holothurians across the entire area and substantially reduced the time required to digitize (Supplementary 1). This method is thus recommended for future studies though care should be taken to ensure sufficient ‘virtual transects’ are used. There is also the possibility of automating counts. For example, species identification and distribution assessments could be undertaken using machine learning (Dujon and Schofield 2019; Lyons et al. 2019). This would substantially accelerate the processing time and allow coverage of even larger areas. Moreover, with continual improvements in flight endurance and camera resolution (Crutsinger et al. 2016), we predict that the areas that drones can survey for holothurian monitoring will increase, allowing larger and more rapid mapping of holothurian densities.
While drone and satellite imagery robustly mapped H. atra density at a reef scale in this study, the biology, demography and ecology of the organisms to be mapped are important considerations in the efficacy of this method. The distribution of holothurians is typically linked with natural sediment features; however, the abundance and movement of holothurians can also be influenced by the life stage of the animal, sediment quantity and quality, light intensity, water temperature and depth (Sloan and von Bodungen 1980; Uthicke and Karez 1999; Dong et al. 2011; Morgan 2011; Navarro et al. 2013; Domínguez-Godino and González-Wangüemert 2020). As such, holothurians may be patchily distributed at scales finer than the geomorphic zones reported in this study and patchiness may be species-specific (Klinger et al. 1994). Knowledge of the ecology and biology of the organism to be mapped is thus important to the choice of scale and interpretation of drone-based population assessments. Drones give the ability to capture continuous high spatial and temporal resolution data over much larger areas than in-water methods and also enable finer-scale distributions of organisms to be monitored over time.
Implications for fisheries management
While H. atra, is the most common species on Heron Reef (Williamson et al. 2017), it is only a low value species in the bêche-de-mer fisheries (Purcell et al. 2010; Eriksson and Byrne 2015). The mapping method presented in this research could easily be translated to higher value commercial species where they occur in shallow reefs. H. leucospilota, H. edulis, S. hermanni and S. variegatus were also able to be identified in the imagery in this study. Many of these species are targeted for traditional fisheries in the Pacific (Drumm and Loneragan 2005; Friedman et al. 2011), with deeper-water species such as H. whitmaei and H. scabra more often targeted by commercial fisheries on the Great Barrier Reef and Pacific islands (Uthicke and Benzie 2001a, b). Our study suggests that if bêche-de-mer fisheries target only a few species, leaving dominant species such as H. atra present on reefs, the repercussions on coral reef ecosystems may not be as serious as the impacts of a broader, indiscriminate fishery. However the differing rates of bioturbation between holothurian species must be taken into account. For example, less abundant species targeted by fisheries such as Thelenota ananas are much larger (3–6 kg) than H. atra (< 100 g) (Purcell et al. 2016) and thus likely to bioturbate comparatively more. Studies should determine broader patterns of bioturbation in holothurians and relate this to the biology and ecology of each species. Changes in total bioturbation from holothurians in a reef could then be estimated and modeled as a result of specific species and population declines.
Implications to ecosystem functioning and reef health
The link between healthy reef systems and bioturbation rates of holothurians is largely unknown. In the context of holothurian overharvesting through bêche-de-mer fisheries, which can reduce holothurian abundances to a quarter of initial numbers for over 50 years (Holland 1994), associated bioturbation of benthic systems without holothurians could decline to less than a quarter of the levels found in a healthy reef systems. Direct extrapolation between total bioturbation, as determined in this study, and the associated benefits of bioturbation (e.g., algal productivity) is, however, not possible from currently available research. To our knowledge, no study has directly examined the link between holothurian bioturbation rates and flow-on benefits to ecosystems. There is evidence that a localized loss of holothurians causes a 63% reduction in O2 sediment penetration (Lee et al. 2017), but it is not clear how bioturbation rates relate to this. Future studies should aim to directly link bioturbation rates to ecosystem flow-on benefits to more accurately predict the effects of declining holothurian populations on coral reefs.
It is well accepted that coral reefs are projected to continue suffering substantial losses of reef structure and functionality from changing ocean conditions, including the dissolution of calcium carbonate due to ocean acidification (Albright et al. 2016; Doney et al. 2009; Johnson et al. 2014; Kornder et al. 2018; Shaw et al. 2015). The positive effects of biogenic buffering on carbonate chemistry by one species of holothurian has recently been documented (Wolfe et al. 2018). The mass of bioturbated sediments produced by holothurians on a reef scale, conservatively estimated here to be slightly higher than the mass of five Eiffel Towers (Castellaro et al. 2016) per year on Heron Reef, highlights the scale of the effect that these organisms may have as biogenic buffers against increasing dissolution of calcium carbonate. We advocate that the functional role of holothurians on coral reefs is highly likely to be more substantial than previously thought and that greater attention needs to be directed to their management and ecology, particularly in relation to overharvesting on reefs already compromised in resilience.
References
Albright R, Caldeira L, Hosfelt J, Kwiatkowski L, Maclaren JK, Mason BM, Nebuchina Y, Ninokawa A, Pongratz J, Ricke KL, Rivlin T, Schneider K, Sesboüé SK, Silverman J, Wolfe K, Zhu K, Caldiera K (2016) Reversal of ocean acidification enhances net coral reef calcification. Nature 531:362–365
Anderson K, Gaston KJ (2013) Lightweight unmanned aerial vehicles will revolutionize spatial ecology. Front Ecol Environ 11:138–146
Anderson SC, Flemming JM, Watson R, Lotze HK (2011) Serial exploitation of global sea cucumber fisheries. Fish Fish 12:317–339
Bates D, Mächler M, Bolker B, Walker S (2014) Fitting linear mixed-effects models using lme4. arXiv preprint arXiv:1406.5823
Bellwood DR, Pratchett MS, Morrison TH, Gurney GG, Hughes TP, Alvarez-Romero JG, Day JC, Grantham R, Grech A, Hoey AS, Jones GP, Pandolfi JM, Tebbett SB, Techera E, Weeks R, Cumming GS (2019) Coral reef conservation in the Anthropocene: confrinting spatial mismatches and prioritizing functions. Biol Conserv 236:604–615
Castellaro S, Perricone L, Bartolomei M, Isani S (2016) Dynamic characterization of the Eiffel tower. Eng Struct 126:628–640
Conand C (2004) Present status of world sea cucumber resources and utilization: an international overview. Adv Sea Cucumber Aquac Manag 463:13–23
Crutsinger GM, Short J, Sollenberger R (2016) The future of UAVs in ecology: an insider perspective from the Silicon Valley drone industry. NRC Research Press
Domínguez-Godino JA, González-Wangüemert M (2020) Habitat associations and seasonal abundance patterns of the sea cucumber Holothuria arguinensis at Ria Formosa coastal lagoon (South Portugal). Aquat Ecol 54:337–354
Doney SC, Fabry VJ, Feely RA, Kleypass JA (2009) Ocean acidification: the other CO2 problem. Annual Review of Marine Science. 1. Palo Alto: Annual Reviews 1:169–192
Dong G, Dong S, Tian X, Wang F (2011) Effects of photoperiod on daily activity rhythym of juvenile sea cucumber, Apostichopus japonicus (Selenka). China J Oceanol Limnol 29:1015–1022
Drumm DJ, Loneragan NR (2005) Reproductive biology of Holothuria leucospilota in the Cook Islands and the implications of traditional fishing of gonads on the population. N Z J Mar Freshwater Res 39:141–156
Drumm DJ, Loneragan NR, Ellis N, Skilleter GA (2011) The significance of habitat and exposire to the reef-flat macroinvertebrates of Rarotonga, Cook Islands, for conservation planning. Pac Conservat Biol 17:320–337
Dujon AM, Schofield G (2019) Importance of machine learning for enhancing ecological studies using information-rich imagery. Endanger Spec Res 39:91–104
Eriksson H, Byrne M (2015) The sea cucumber fishery in Australia’s Great Barrier Reef Marine Park follows global patterns of serial exploitation. Fish Fish 16(2):329–341
Fraser KP, Peck LS, Clarke A (2004) Protein synthesis, RNA concentrations, nitrogen excretion, and metabolism vary seasonally in the Antarctic holothurian Heterocucumis steineni (Ludwig 1898). Physiol Biochem Zool 77:556–569
Friedman K, Eriksson H, Tardy E, Pakoa K (2011) Management of sea cucumber stocks: patterns of vulnerability and recovery of sea cucumber stocks impacted by fishing. Fish Fish 12:75–93
Gattuso J-P, Frankignoulle M, Bourge I, Romaine S, Buddemeier RW (1998) Effect of calcium carbonate saturation of seawater on coral calcification. Global Planet Change 18:37–46
Great Barrier Reef Marine Park Authority (2003) Great Barrier Reef Marine Park Zoning Plan 2003. ISBN 1 876945 38 9
Guzman HM, Guevara CA (2002) Population structure, distribution, and abundance of three commercial species of sea cucumber (Echinodermata) in Panama. Caribb J Sci 38:230–238
Hammond LS (1982) Patterns of feeding and activity in deposit-feeding holothurians and echinoids (Echinodermata) from a shallow back-reef lagoon, Discovery Bay, Jamaica. Bull Mar Sci 32:549–571
Hammond AR, Meyers L, Purcell SW (2020) Not so sluggish: movement and sediment turnover of the world’s heaviest holothuroid, Thelenota anax. Mar Biol 167:1–9
Han Q, Keesing JK, Liu D (2016) A review of sea cucumber aquaculture, ranching, and stock enhancement in China. Reviews in Fisheries Science & Aquaculture 24:326–341
Hodgson JC, Mott R, Baylis SM, Pham TT, Wotherspoon S, Kilpatrick AD, Raja Segaran R, Reid I, Terauds A, Koh LP (2018) Drones count wildlife more accurately and precisely than humans. Methods Ecol Evol 9:1160–1167
Holland A (1994) The status of global beche-de-mer fisheries with special reference to the Solomon Islands and the potentials of holothurian culture. Unpublished M Sc Thesis, University of Newcastle, United Kingdom
Hopley D, Smithers SG, Parnell KE (2007) The Geomorphology of the Great Barrier Reef: Development. Cambridge University Press, Diversity and Change
Hughes TP, Baird AH, Bellwood DR, Card M, Connolly SR, Folke C, Grosberg R, Hoegh-Guldberg O, Jackson JBC, Kleypass J, Lough JM, Marshall P, Nyström M, Palumbi S, Pandolfi JM, Rosen B, Roughgarden J (2003) Climate change, human impacts, and the resilience of coral reefs. Science 301:929–933
Hughes TP, Kerry JT, Álvarez-Noriega M, Álvarez-Romero JG, Anderson KD, Baird AH, Babcock RC, Beger M, Bellwood DR, Berkelmans R (2017) Global warming and recurrent mass bleaching of corals. Nature 543:373
Hughes T, Kerry J, Simpson T (2018) Large-scale bleaching of corals on the Great Barrier Reef. Ecology 99:501–501
İşgören-Emiroğlu D, Günay D (2007) The effect of sea cucumber Holothuria tubulosa (G., 1788) on nutrient and organic matter contents of bottom sediment of oligotrophic and hypereutrophic shores. Fresenius Environ Bull 16:290–294
Johnson MD, Price NN, Smith JE (2014) Contracting effects of ocean acificiation on tropical fleshy and calcareous algae. PeerJ 2:e411
Joyce K, Duce S, Leahy S, Leon JX, Maier S (2019) Principles and practice of acquiring drone based image data in marine environments. Mar Freshwat Res 70:952–963
Kennedy E, Roelfsema C, Lyons M, Kovacs E, Borrego-Acevedo R, Roe M, Phinn S, Larsen, K, Murray N, Yuwono D, Wolff J, Tudman P (2020) Reef cover: a coral reef classification to guide global habitat mapping from remote sensing. bioRxiv, https://doi.org/https://doi.org/10.1101/2020.09.10.292243 Accessed 31 October 2020
Klinger TS, Johnson CR, Jell J (1994) Sediment utilization, feeding-niche breadth, and feeding-niche overlap of Aspidochirotida (Echinodermata: Holothuroidea) at Heron Island, Great Barrier Reef. In David B, Guille A, Feral J-P, Roux M (eds), Echinoderms Through Time: proceedings of the Eighth International Echinoderm Conference, Dijon, France, 6–10 September 1993, Balkema, Rotterdam, Netherlands, pp. 523–528
Kornder NA, Riegl BM, Figueiredo J (2018) Thresholds and drivers of coral calcification responses to climate change. Glob Change Biol 24:5084–5095
Lee S, Ferse SC, Ford A, Wild C, Mangubhai S (2017) Effect of sea cucumber density on the health of reef-flat sediments. Wildlife Conservation Society
Lenth R, Singmann H, Love J (2018) Emmeans: Estimated marginal means, aka least-squares means. R package version 1
Lyons MB, Brandis KJ, Murray NJ, Wilshire JH, McCann JA, Kingsford RT, Callaghan CT (2019) Monitoring large and complex wildlife aggregations with drones. Methods Ecol Evol 10:1024–1035
Madin E, Precoda K, Harborne A, Atwood TB, Roelfsema CM, Luiz O (2019) Multi-trophic species interactions shape seascape-scale coral reef vegetation patterns. Frontiers in Ecology and Evolution 7:102
Mangion P, Taddei D, Frouin P, Conand C (2004) Feeding rate and impact of sediment reworking by two deposit feeding feeders Holothuria leucospilota and Holothuria atra on a fringing reef (Reunion Island, Indian). Echinoderms: Munchen. In: Heinzeller T & Nebelsick JH (eds). London: Taylor & Francis, pp. 311–317
Morgan AD (2011) Patterns of distribution and abundance of the temperate sea cucumber Australostichopus mollis on a rocky subtidal reef. New Zeal J Zool 38:195–206
Navarro PG, García-Sanz S, Barrio JM, Tuya F (2013) Feeding and movement patterns of the sea cucumber Holothuria sanctori. Mar Biol 160:2957–2966
Plagányi ÉE, Skewes T, Murphy N, Pascual R, Fischer M (2015) Crop rotations in the sea: Increasing returns and reducing risk of collapse in sea cucumber fisheries. Proc Natl Acad Sci 112:6760–6765
Purcell S (2010) Managing Sea Cucumber Fisheries with an Ecosystem Approach. FAO Fisheries and Aquaculture Technical Paper No. 520, FAO, Rome
Purcell SW, Mercier A, Conand C, Hamel JF, Toral-Granda MV, Lovatelli A, Uthicke S (2013) Sea cucumber fisheries: global analysis of stocks, management measures and drivers of overfishing. Fish Fish 14:34–59
Purcell SW, Piddocke TP, Dalton SJ, Wang Y-G (2016) Movement and growth of the coral reef holothuroids Bohadschia argus and Thelenota ananas. Mar Ecol Prog Ser 551:201–214
Raoult V, Gaston T (2018) Rapid biomass and size-frequency estimates of edible jellyfish populations using drones. Fish Res 207:160–164
Raoult V, David PA, Dupont SF, Mathewson CP, O’Neill SJ, Powell NN, Williamson JE (2016) GoPros™ as an underwater photogrammetry tool for citizen science. PeerJ 4:e1960
Raoult V, Tosetto L, Williamson JE (2018) Drone-Based High-Resolution Tracking of Aquatic Vertebrates. Drones 2:37
Schiell GR, Knott B (2010) Aggregations and temporal changes in the activity and bioturbation contribution of the sea cucumber Holothuria whitmaei (Echinodermata: Holothuroidea). Mar Ecol Prog Ser 415:127–139
Schneider K, Silverman J, Woolsey E, Eriksson H, Byrne M, Caldeira K (2011) Potential influence of sea cucumbers on coral reef CaCO3 budget: A case study at One Tree Reef. Journal of Geophysical Research: Biogeosciences 116:G04032
Schneider K, Silverman J, Kravitz B, Rivlin T, Schneider-Mor A, Barbosa S, Byrne M, Caldeira K (2013) Inorganic carbon turnover caused by digestion of carbonate sands and metabolic activity of holothurians. Estuar Coast Shelf Sci 133:217–223
Shaw EC, Phinn SR, Tilbrook B, Steven A (2015) Natural in situ relations suggest coral reef calcium carbonate production will decline with ocean acidification. Limnol Oceanogr 60:777–788
Sloan NA, von Bodungen B (1980) Distribution and feeding of the sea cucumber Isostichopus badionatus in relation to shelter and sediment criteria of the Bermuda Platform. Mar Ecol Prog Ser 2:14–28
Team RC (2013) R: A language and environment for statistical computing
Tuya F, Hernandez JC, Clemente S (2006) Is there a link between the type of habitat and the patterns of abundance of holothurians in shallow rocky reefs? Hydrobiologia 571:191–199
Uthicke S (1999) Sediment bioturbation and impact of feeding activity of Holothuria (Halodeima) atra and Stichopus chloronotus, two sediment feeding holothurians, at Lizard Island, Great Barrier Reef. Bull Mar Sci 64:129–141
Uthicke S (2001a) Nutrient regeneration by abundant coral reef holothurians. J Exp Mar Biol Ecol 265:153–170
Uthicke S (2001b) Interactions between sediment-feeders and microalgae on coral reefs: grazing losses versus production enhancement. Mar Ecol Prog Ser 210:125–138
Uthicke S, Benzie J (2001a) Effect of bêche-de-mer fishing on densities and size structure of Holothuria nobilis (Echinodermata: Holothuroidea) populations on the Great Barrier Reef. Coral Reefs 19:271–276
Uthicke S, Benzie J (2001b) Restricted gene flow between Holothuria scabra (Echinodermata: Holothuroidea) populations along the north-east coast of Australia and the Solomon Islands. Mar Ecol Prog Ser 216:109–117
Uthicke S, Karez R (1999) Sediment patch selectivity in tropical sea cucumbers (Holothurioidea: Aspidochirotida) analysed with multiple choice experiments. J Exp Mar Biol Ecol 236:69–87
Uthicke S, Klumpp D (1998) Microphytobenthos community production at a near-shore coral reef: seasonal variation and response to ammonium recycled by holothurians. Mar Ecol Prog Ser 169:1–11
Uthicke S, Benzie J, Ballment E (1999) Population genetics of the fissiparous holothurian Stichopus chloronotus (Aspidochirotida) on the Great Barrier Reef, Australia. Coral Reefs 18:123–132
Uthicke S, Welch D, Benzie J (2004) Slow growth and lack of recovery in overfished holothurians on the Great Barrier Reef: evidence from DNA fingerprints and repeated large-scale surveys. Conserv Biol 18:1395–1404
Wheeling RJ, Verde EA, Nestler JR (2007) Diel cycles of activity, metabolism, and ammonium concentration in tropical holothurians. Mar Biol 152:297–305
Williamson JE, Byrnes EE, Clark JA, Connolly DM, Schiller SE, Thompson JA, Tosetto L, Martinelli JC, Raoult V (2017) Ecological impacts and management implications of reef walking on a tropical reef flat community. Mar Pollut Bull 114:742–750
Wolfe K, Byrne M (2017) Biology and ecology of the vulnerable holothuroid, Stichopus herrmanni, on a high-latitude coral reef on the Great Barrier Reef. Coral Reefs 36:1143–1156
Wolfe K, Vidal-Ramirez F, Dove S, Deaker D, Byrne M (2018) Altered sediment biota and lagoon habitat carbonate dynamics due to sea cucumber bioturbation in a high-pCO2 environment. Glob Chang Biol 24:465–480
Yamanouchi T (1939) Ecological and physiological studies on the holothurians in the coral reef of Palao Islands. Palao Tropical Biological Station Studies, Report 25:603–634
Acknowledgements
The authors thank Kristy Brown, Katy Baker and Edith Shum for their assistance with digitizing and Gemma Clifton and Olivia Seeger for assistance in the field. Eva Kovacs and Chris Roelfsema are also thanked for their provision of classified satellite imagery. Funding for this research was provided by the Department of Biological Sciences at Macquarie University, the University of Newcastle, and an Early Career Rising Star Grant from JCU (Duce). Thanks are due to the University of Queensland and the staff at Heron Island Research Station for the use of their facilities. The authors of this manuscript declare that they have no conflicts of interest and that research was conducted under the JCU Limited Impact Research in the GBRMP Memorandum of Understanding between JCU and the GBRMPA.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Topic Editor Morgan S. Pratchett
Supplementary Information
Below is the link to the Supplementary Information.
Rights and permissions
About this article

Cite this article
Williamson, J.E., Duce, S., Joyce, K.E. et al. Putting sea cucumbers on the map: projected holothurian bioturbation rates on a coral reef scale. Coral Reefs 40, 559–569 (2021). https://doi.org/10.1007/s00338-021-02057-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00338-021-02057-2