
Abstract
Maynard-Smith’s game-theoretical framework suggests that conflicts should be resolved through an exchange of agonistic displays which can be as effective as physical attack in securing contested resources. Hence, participants use signaling tactics that are governed by game-theoretical competition rules to maximize their benefit from the interaction. In rock hyraxes, males initiate singing in response to conspecific male songs, possibly to self-advertise and to facilitate comparison of performance by receivers. We hypothesized that hyrax counter-singing strategy can be explained by resident-intruder model and its outcome is determined by the identity and ability of the participants. Natural hyrax singing events and playback-induced events were classified according to the participants’ residence status—resident or bachelor (i.e., non-resident). As predicted by the resident-intruder model, resident listeners tended to reply to conspecific males at high rates (≈40 %), while bachelors showed low reply rates (17 %) to resident initiators. Unexpectedly, the lowest-ranking bachelors showed a tendency to reply to the highest-ranking residents. One possible explanation for this counter-intuitive behavior might be that weaker individuals exploit a situation in which it is unremunerated for a stronger opponent to retaliate (i.e., Napoleon strategy) and seize a self-advertisement opportunity in front of a large audience attending to the high-ranking males’ signal. Overall, we show that male hyraxes employ different counter-singing tactics depending on the residence and rank differences between the participants. This choice of competition tactics can be based on opponents’ individual recognition or/and on direct decoding of opponents’ traits encoded in the song.
Significance statement
In nature, conflicts are usually resolved through signaling rather than by actual fighting. Rock hyrax males often participate in vocal competitions, possibly to reduce the need for physical confrontations. During our study on the dynamics of male hyrax vocal competition, we discovered that hyraxes employ mixed counter-singing strategy dependent on the residence status and social rank differences between the participants. A paradoxical “Napoleon-complex” tactic was observed in interactions between males, when low ranking bachelors were found to interact and challenge the highest-ranking residents. On the other hand, residents were found to interact with opponents regardless of residency and/or rank. On top of the cognitive abilities required to individually recognize an opponent and possibly decode its vocal signals, our results suggest that male hyraxes compare own individual traits to those of the opponent in a mutual signal assessment interaction.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
In any signaling contest, two types of information are important to the receivers: the signaler’s willingness to escalate (aggressiveness) and its ability to win a fight (i.e., resource-holding potential, RHP (Hurd 2006; Searcy and Beecher 2009)). According to the resident-intruder model (Maynard-Smith and Parker 1976; Kokko et al. 2006), individuals that monopolize resources (residents) are expected to be more aggressive in defending their assets (Svensson et al. 2012), because of the motivational asymmetry between them and intruder individuals (Maynard-Smith and Parker 1976; Yack et al. 2014). Residents gain a higher pay-off for successfully defending their resource due to the time and energy invested in establishing and defending the resource in the past (Bergman et al. 2010), and they consequentially suffer a greater loss than intruders if failing to do so. This makes residents’ tactics predictably aggressive and sometimes creates a “prior-ownership” effect, a situation in which other individuals respect ownership and residents are seldom contested (Grafen 1987; Russ et al. 2010).
In contrast, intruders have a choice between “challenge” and “do not challenge” tactics in confrontation with a resident male. They are expected to carefully assess their opponent before making a decision on how to proceed in a potentially costly contest (Lindstrom 2001; Morrell et al. 2005) and are more likely to escalate against a weaker resident whom they have higher chances to beat (Aragon et al. 2006).
Another issue that is often considered in management of competition strategy is the familiarity with the opponent. The “dear enemy” effect (Getty 1987) predicts lesser aggression towards neighbors than towards strangers or neighbors appearing in an unexpected locations (Bolyard and Rowland 2000). The differences in aggression levels are due to higher potential losses to individuals establishing their ground in a completely new area (Getty 1987) and due to the need to learn about the new opponent (Getty 1989). However, behaviors opposite to “dear enemy” were reported in several species (Schradin et al. 2010), and Temeles (1994) suggested that local individuals’ behavior is determined by the relative levels of threat presented by neighbors and strangers.
Male-to-male vocal displays have been recognized as a competition category in numerous mammalian species (e.g., baboons (Papio cynocephalus ursinus) (Fischer et al. 2004), red deer (Cervus elaphus) (Reby et al. 2005), and arctic foxes (Alopex lagopus) (Frommolt et al. 2003)). Males compete with one another mostly for resources (food, mates, or territory) (Shuster and Wade 2003) and, as in other competition types, vocal displays require strategic decisions based on opponent assessment and prediction of its behavioral response (Jennings et al. 2012). For example, male red deer alter their vocalization according to the challenger’s call in order to emphasize their own quality and physical ability (Behr et al. 2009), and male baboons might retreat from vocal competition when outranked by a stronger opponent (Kitchen et al. 2003).
In this study, we used the highly social and vocal rock hyrax (Procavia capensis) as a model system to investigate the role of counter singing. Our aim was to determine if counter singing dynamics in hyrax society is associated with different social tactics and whether it could be explained by the resident-intruder model. Hyraxes live in groups consisting mainly of one adult, resident male, a 5–20 females and their offspring (Barocas et al. 2011; Koren and Geffen 2011). Group-born males disperse when they reach sexual maturity. They often remain in the same area as bachelors (i.e., non-resident males), but do not stably associate with any group (Barocas et al. 2011), and their hierarchal rank is set according to the outcome of agonistic interactions with neighboring males (Koren et al. 2008). In general, hyrax males are not territorial and do not monopolize food resources. During the mating season, the bachelor males can be seen courting and mating with females from different groups, despite the resident males’ aggression towards them. However, only resident males were observed mate guarding older and experienced females (Bar Ziv et al. 2016). Thus, we suggest that from the residents’ point of view, bachelors are considered intruders that challenge ownership of the group resource, which is a better access to quality females (Bar Ziv et al. 2016).
Acoustic communication is a major information transfer channel in the rock hyrax (Fourie 1977). Adult males produce elaborate and complex songs that, while being energetically cheap (Ilany et al. 2013), bear individual signatures (Koren and Geffen 2011) as well as transfer honest information about body weight, size, hormonal levels, and social rank of the singer (Koren and Geffen 2009). Male hyraxes tend to sing in response to other male songs, possibly to announce their presence and advertise their abilities to the audience (Ilany et al. 2011) as a part of intrasexual competition (Koren and Geffen 2009).
Hyrax males can be aggressive towards each other, and close encounters between them may result in chases or fights that can lead to injuries or death (Barocas et al. 2011). On the other hand, counter-singing sessions are performed while the opponents are spatially separated and usually end without any physical contact. Counter-singing displays are not random events of simultaneous singing since songs performed in counter-singing sessions were found to be more elaborate in comparison to songs performed without an audience (Demartsev et al. 2014). Thus, we suggest that hyrax-counter singing is most likely a mutual signaling between males, which enables them to advertise relative social and physical traits (Koren and Geffen 2009) without the need for actual fighting.
Following this assumption, we hypothesized that hyrax counter-singing strategies should depend on the participants’ identity, familiarity, and individual traits such as body weight, residency status, social rank, and tenure on site. Due to the clear and stable division of hyrax males into residents and bachelors (i.e., non-residents/intruders), we predicted that their counter-singing tactics would be consistent with the resident-intruder model and show a strong ownership effect, in which resident males are seldom challenged and always defend their resources. We anticipated that bachelors would evade counter-singing against much heavier and stronger residents to avoid the possibility of a costly physical confrontation. Finally, we hypothesized that, if hyrax males can assess competitors’ quality from attending to their songs, the reply pattern observed during singing events induced by the playbacks of hyrax songs will be similar to the pattern observed in natural counter-singing events.
Materials and methods
Field protocol
Our study is part of a long-term project conducted at the Ein Gedi Nature Reserve, Israel (31° 28′ N, 35° 24′ E) since 1999. At two study sites, we monitored several mixed-sex hyrax groups and bachelor males observed in defined areas. Field procedures followed previously published protocols (Koren et al. 2008; Demartsev et al. 2014).
Briefly, throughout 15 field seasons (March to August; 2000–2014), we conducted field observation sessions during hyrax morning activity hours. Hyraxes were trapped using live box traps (Tomahawk Live Trap Co, Tomahawk, WI, USA) baited with cabbage and kohlrabi. Trapped animals were anesthetized by intramuscular injection of ketamine hydrochloride (0.1 ml/kg), weighed, measured, and individually marked with an electronic subcutaneous transponder (DataMars SA), ear tags, and/or a lightweight-tagged collar (5 g). Following anesthesia recovery (at least 120 min), the animals were released at the location of capture. Recaptures were released immediately after weighing. All animals were handled in shaded places to prevent overheating. Over the course of the study, more than 400 hyraxes were individually marked and no adverse effects by the transponders or the collars were observed.
Vocalization and behavioral recording
Hyrax activities were observed daily using 10X42 binoculars (Monarch, Nikon) and a telescope with up to X75 zoom magnification (Fieldscope ED82, Nikon). It was not possible to record behavioral data blindly because our study involved observation and identification of marked animals in the field. On each observation day, all movements, social interactions, feeding events, and vocalizations of a focal social group and of the bachelor males in its vicinity were recorded (Barocas et al. 2011). When singing was observed, the singer’s identity and singing context were noted, and the singing individual was defined as initiator. All hyrax males that were visible and identifiable during the singing session were noted and defined as listeners. If no vocal response to the initiation song was noted, event was classified as “no reply.” The interaction was defined as counter-singing and classified as “reply” if a singing response, by at least one hyrax male, was observed during or within 2.5 min after the initiation of a singing session (Demartsev et al. 2014). The first male that replied to the initiator’s singing was defined as a responder; all additional replies beyond the first one were disregarded in this analysis. Singing events were selected for analysis according to the following criteria: (1) the initiator was visually identified; (2) at least one additional male was observed and identified in the area during the singing session; (3) we had body weight and/or social rank records for both participants in the relevant field season.
Male group membership was defined by generating association based on social networks as described in Barocas et al. (2011). Bachelors were defined as adult males that are not steadily associated with any of the mixed-sex social groups, whereas residents were defined as adult males that are regularly observed as part of a mixed-sex social group, shared sleeping dens and seen interacting with group females beyond mating context.
Social rank
All agonistic interactions between identified individuals were recorded following our previously published protocol (Koren et al. 2008). Only interactions that involved clear agonistic display by one individual and resulted in an evasive action being taken by the recipient were considered in the social hierarchy analysis. A matrix of encounters was prepared for each research site using all pair-wise agonistic interactions observed during each field season. Social hierarchy was determined using the David’s score procedure (Gammell et al. 2003). Social rank scores were normally distributed.
For analysis of body weight effect on the outcome of fighting, we used 138 male-male agonistic interactions. The interactions were classified according to the participants’ residency status combination (bachelor/bachelor or resident/bachelor). We have not observed any resident/resident interactions. For each interaction class (e.g., resident/ bachelor), the win/lose ratio of the heavier males was calculated and tested against the win/lose ratio expected by random.
Playback experiments
Long-range vocalizations were recorded from a distance of 10–50 m with a Sennheiser ME 67 shotgun microphone (frequency response 50–20,000 Hz ±2.5 dB) powered by a Sennheiser K6 module, covered with a Sennheiser MZW70-1 blimp windscreen (Sennheiser Electronic GmbH & Co. K. G., Wedemark, Germany). The microphone was hand-held using an MZS20-1 shock-mount with a pistol grip. Vocalizations were recorded in mono (Tascam HD-P2 digital audio recorder; TASCAM Corporation, Montebello, CA, USA), with a sampling frequency of 48 kHz and a sampling width of 24 bits (Ilany et al. 2011).
Eleven previously recorded songs from 11 different adult males (Table S1b) were played (average 9.09 ± 0.87 playbacks per song) via a FoxPro Scorpion X1B speaker using a TX200 remote control (FOXPRO Inc., Lewistown, PA, USA). The speaker was placed before dawn in a concealed spot in an area frequently visited by the focal hyraxes. After a male hyrax was observed and identified within a 5–30-m radius of the speaker, one of the songs was played. All songs selected for playback experiments were 1.5–2 min in length, which is the average hyrax song duration. The volume of playbacks was normalized to natural SPL using a graphic sound meter built into a Tascam HD-P2 audio recorder. Natural intensity levels of hyrax songs (about 80 dB) were measured in a previous study by attaching miniaturized audio recorders to individual males (Ilany et al. 2013).
Playback tracks were evenly rotated throughout playback sessions. To control for individual recognition of the song performer, we included both local (recorded at the experiment site) and foreign (recorded at minimal distance of 3 km from the experiment site) songs in each experimental playback set. No male received a playback of his own recording. To avoid desensitization and to minimize disturbance to hyrax daily routine, we restricted the number of performed playbacks to two per day with a minimum interval of 30 min between consecutive trials. We recorded behavioral responses of all visible individuals found in ~40 m radius around the speaker, 5 min before, during, and continuously for 5 more min after each trial.
Statistical analysis
We tested the effect of five potentially important variables on reply rate (ratio of reply to total singing events, Fig. 1). (1) The difference in body weight between the initiator and the responder/listener (∆BW). Body weight usually serves as a good RHP indicator and was previously found to be correlated with male song features (Koren and Geffen 2009). (2) The difference in social rank (i.e., David’s score) between initiator and responder/listener (∆SR). (3) Male tenure (MT), expressed as the number of years the initiator and responder/listener shared the same locality. (4) Male residency status combination (MRSC), expressed as one of four possibilities of the initiator and responder/listener in a given singing event (Bachelor-Bachelor, Resident-Resident, Bachelor-Resident, Resident-Bachelor). (5) Whether the singing event took place during or out of the annual mating season. Mating season was determined between the first and the last mating attempt observed in a given year.

Frequency distribution of reply to male rock hyrax natural songs by dyad type. The residency state of the initiator and the male responded or present nearby are indicated by the top and bottom lines, respectively, and by abbreviation. Sample size is indicated above bars. Horizontal lines denote significant differences (P = 0.02) in reply frequency between dyad types (GEE, sequential Bonferroni correction)
In the analysis of the playback data, we excluded social rank because this measure cannot be evaluated for males that never interact, a situation that occurs when strangers were used as the initiators in playbacks. Furthermore, for the playback data analysis, we added an additional binary nominal term called acquaintance (Acq), which defines whether the playback was from a local male (i.e., possibly known to the listener/responder) or a stranger (i.e., a male outside the gorge occupied by the listener/responder).
We used a logistic regression under the framework of the generalized estimating equations (GEE) to test for the effect of the above four variables on the probability of reply to the song of the initiator. GEE is an extension of generalized linear models (GLM) for correlated data (i.e., mixed model). We set initiator and listener identities as a random effect in the GEE analyses. The GEE approach results in estimates of model parameters that are robust regardless of correlation structure between observations (repeated measures within the male subjects). We selected the robust estimator (sandwich estimator) because it consistently estimates the covariance even if the working correlation matrix is misspecified. The Wald χ 2 metric was used for testing the significance of each effect. Multiple comparisons were conducted using the sequential Bonferroni correction. GEE model fitting was performed using SPSS (version 22, SPSS Inc.).
Results
Agonistic matches
To test whether body weight affects the probability of winning a physical conflict, we examined the outcome of 34 agonistic matches between resident and bachelor males and 104 matches between two bachelor males. Resident males won all matches against solitary males even though in 20 cases (59 %) the bachelor male was heavier. This winning rate was significantly higher than expected from body weight differences between the competitors (binomial probability, P < 0.0001). In contrast, 62.5 % of matches between bachelor males were won by the heavier male, a winning rate that is significantly higher than expected by random (P = 0.003). Although male social rank increased with body weight (r 2 = 0.681, F 1,102 = 8.3, P = 0.005), we observed heavy males that were socially low ranked.
Counter-singing
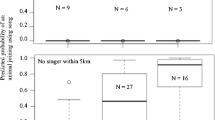
To examine the factors that drive acoustic competition and counter-singing tactics in hyraxes, 261 natural singing events were separated into four dyad types based on the participant’s residency status combination and classified as “reply” or “no reply” according to listeners’ response (Fig. 1 and Table S1a). The distribution of singing events within Resident/Bachelor (RB) and Bachelor/Resident (BR) dyads shows a significant difference in the reply frequency between them (GEE; sequential Bonferroni correction; n = 114; P = 0.02; Fig. 1). Given the rarity of the Resident/Resident (RR) dyad in our observations (about 5 %, Fig. 1), we repeated the above analysis without the RR category. This setup showed that the BR reply rate is significantly higher than RB (GEE; sequential Bonferroni correction; n = 251; P = 0.01) and tended to be higher than Bachelor/Bachelor (BB) (P = 0.07).
Determinants of reply probability
To further understand male hyrax counter-singing, we evaluated the effects of mating season, tenure on site, residency, relative body weight, and relative social rank (David’s score) between the initiator and listener, on singing reply probability (Table 1). Mating season, body weight, and tenure did not have any significant effect on listeners’ reply rate.
As mentioned above, the probability that a bachelor initiator would receive a reply from a resident listener (BR) was significantly higher (46 %) than vice versa (RB; 19 %; Fig. 1, Table 1). This trend was intensified when the difference in social rank between the initiator and responder/listener was incorporated (Fig. 2, Fig. S1). The reply marginal mean of RB was significantly different from those observed for RR (P = 0.037), BB (<0.001), and BR (<0.001). The RB dyad was the only combination that showed a significant rise in reply rate as a function of increase in social rank difference between initiator and listener (model β = 1.1 ± 0.2, Wald χ 2 = 38.6, P < 0.001; Fig. 2). Bachelor listeners tended to reply at a higher rate to resident initiators that were at the highest social rank relative to their own (Fig. 2). Social rank did not affect the reply rate in the other counter-singing dyads (P ≥ 0.365 in all other dyads). The above difference was evident by the significant interaction between residency status and social rank difference (P < 0.001; Table 1).

The probability of replying to a male hyrax natural song as a function of the interaction between Δ social rank (the difference in social rank between the singer and the listener/responder) and residency status combination. The four functions represent the residency state of the initial singer and the male responded or nearby stander. BB denotes bachelor initiator to bachelor listener, RR denotes resident initiator to resident listener, RB denotes resident initiator to bachelor listener, and BR denotes bachelor initiator to resident listener. Shaded areas represent 95 % confidence intervals for the counter singing dyad that showed a significant trend. Parameters were estimated using GEE
Playback experiments
A set of 100 playback experiments, conducted in our research area between the years 2008 and 2014, was analyzed similarly to natural singing events and categorized according to the four residency status dyads (Table S1b). The number of experiments in each dyad type was as follows: RR—10 (5 males), RB—11 (8 males), BR—35 (8 males), BB—44 (21 males). Twenty-three percent (23 %) of the playback experiments resulted in a singing response by a target male. Playback reply rates were similar to rates of natural singing events used in the analysis (27 %) and to the general observed reply rates for hyrax singing (25 %) (Ilany et al. 2011). We evaluated the effect of mating season, body weight difference between initiator and listener, and acquaintance. The only significant effect we detected was for acquaintance (Acq, Table 1). Males tended to reply at a higher rate to familiar individuals (30 %) relative to strangers (17 %, P < 0.038, n = 100). Residency seemed to have no effect on reply rates in playback experiments.
Discussion
Determinants of reply probability
Our analysis of natural counter-singing patterns showed that a male hyrax’ choice of a counter-singing tactic is dependent on both relative residency status and social rank of the participants. Other tested factors (relative body size, tenure on site, and season) had no effect on reply rate variation.
In BR and RR dyads, resident males employed a straightforward “always defend”-like tactic and maintained high reply rates, regardless of their own and their opponent’s rank. Signaling tactics of bachelor males in the presence of a resident male (RB dyad) follow a different pattern, as bachelors have a higher tendency to reply to resident initiators of a higher relative rank. BB dyads interactions showed no significant dependency on any of the tested parameters, neither contesters’ relative body weight nor their social rank affected the listener reply rates, which was nearly equal to the population average.
Our results from the playback experiments did not reproduce the results from natural singing events. We could not statistically differentiate between the four singing dyads and participants’ body weight, and residency had no effect on the listeners’ reply rate. The failure to reproduce natural counter-singing tactics in playback experiments could be due to the considerably smaller sample size and to the generally lower reply rates to songs of unfamiliar males used in the playback experiments. Male hyraxes replied more readily to familiar males than to strangers in contradiction to the “dear enemy” effect (Getty 1987). We believe that male hyrax relative tolerance towards unfamiliar individuals could be explained by the fact that strangers, randomly passing through the area, are perceived as a relatively low threat. Therefore, according to Temeles (1994), they should be more tolerated by local males in comparison to established neighborly competition. Another option is that reliable evaluation of an unfamiliar male requires familiarization with its signal. In this case, an individual may avoid challenging a stranger at first encounter since it could be risky due to possible mis-assessment (Ydenberg et al. 1988). In both cases, we would expect the reply levels to rise if the stranger remains in the area and local males perceive its presence as a higher threat and familiarize themselves with his signal.
Resident males reply tactics
In our previous work, we showed that male songs convey the singers’ social rank (Koren and Geffen 2009), which in our case is based on the outcome of physical confrontations with other males. Our current results show that, in fights between bachelor males, heavier males have a higher winning probability. However, in a physical confrontation between bachelor and resident males, resident usually wins regardless of the differences in body weight. Taken together, these matching trends suggest that ownership-based motivational asymmetry (Maynard-Smith and Parker 1976) affects the outcome of the interactions.
Our results on hyrax counter-singing tactics in BR dyads fit this paradigm and are in line with the resident-intruder model. Residents gain a higher pay-off when winning an aggressive interaction and are expected to always defend against any intruder (Bergman et al. 2010). Therefore, high rates of reply in BR and RR dyads (>40 %, Fig. 1), independent of the difference in the opponent’s social rank (Fig. 2) and consequently its RHP, were expected. A similar strategy has been reported in males of a scincid lizard (Niveoscincus microlepidotus) (Olsson and Shine 2000) and in the tawny dragon (Ctenophorus decresii) (Umbers et al. 2012), where residency was found to predict fighting initiation.
Bachelor males reply tactics
Bachelor males’ reply rates in RB dyads suggest a tendency to respect ownership as residents were replied to in only in 19 % of singing events (Fig. 1). However, when social rank was considered, the results did not fit our initial predictions, as we expected bachelors to avoid counter-singing with high-ranking residents. It might be disadvantageous to signal an inferior ability to a highly motivated and potentially aggressive resident as well as to other potential male and female audience (Morrell et al. 2005; Svensson et al. 2012). However, bachelor males seem to employ a counter-intuitive singing strategy of challenging residents of a higher social rank (Fig. 2). Such puzzling behavior has been previously described in group-living cichlid fish (Neolamprologus pulcher) (Reddon et al. 2013), green anole lizard (Anolis carolinensis) (Jenssen et al. 2005), and velvet swimming crab (Necora puber) (Smith et al. 1994). If the behavior observed here indeed constitutes a deliberate tactic, it can become stable under one of two game-theoretical models; The “Napoleon strategy” suggested by Morrell et al. (2005) or the “Desperado effect” described by Grafen (1987). Thus, we discuss both models below.
Under the Napoleon strategy, weaker individuals could exploit stronger opponents’ tolerance towards subordinate contestants (Miyai et al. 2011) and profit from signaling against them. This strategy can become stable if resources are of relatively low value or are freely available. Since the payoff from winning cheap resources is small, stronger individual can benefit from retreating without aggression (Morrell et al. 2005) or at least from delaying the escalation of the conflict as much as possible (Just and Morris 2003). Another prerequisite of Napoleon strategy is the existence of cheap and honest signals (Morrell et al. 2005). Multiple repeated interactions between individuals facilitate acquaintance and can eliminate the need for exaggerated and costly display (Michod and Hasson 1990).
In hyraxes, receiver attention is apparently difficult to attract and males often exploit audience alertness in order to increase their signaling efficiency (Ilany et al. 2011). Signaling by a high quality, resident male might draw greater conspecific attention, so a subsequent signal by weaker male might enjoy a larger audience of listeners than it could reach independently. In addition, as previously suggested by Lotem (1998), an individual’s success in a competition depends on an integration of quality and motivation. As low ranking bachelors may have difficulty in concealing their inferiority in a stable population, their tactics may be to emphasize persistence and motivation in an attempt to compensate for quality. A similar strategy has been reported in black grouse (Tetrao tetrix), where lifetime attendance in male–male competitions was considered an attractive trait even for low-quality males (Kokko et al. 1999).
An alternative explanation for the bachelor tendency to challenge high-ranking residents could be the Desperado effect. This model predicts lower quality males to be aggressive towards stronger opponents as a desperate attempt to obtain access to essential resources (Grafen 1987). A weaker individual who constantly avoids conflicts against stronger opponents may find itself deprived of crucial resources needed for survival or reproduction. Thus, they are forced into a desperate, “nothing to lose” conflicts, disregarding RHP asymmetries and despite the danger of injury or death (Grafen 1987; Elias et al. 2010). This can be a winning tactic, as “daring” to escalate is sometimes enough to overcome a disadvantage in fighting abilities (Barlow et al. 1986). The high value of the contested resource plays a key role in the desperado effect model, implying that it might not fit into the context of our system. In our study site, food and shelter are abundantly available all year round and females are not being strictly monopolized, as all males are observed copulating, regardless of rank and residency (Bar Ziv et al. 2016). In contrast, the Napoleon strategy pre-requisites low resource value and thus offers a better explanation to the bachelor male signaling strategy in a BR dyad.
However, given the fact that at this point we cannot show a direct gain of resources following counter-singing interactions and can only hypothesize on its likelihood, there is a third explanation to be considered. We can speculate that resident-bachelor counter-singing is not a challenge but actually an appeasing signal. A weak bachelor intentionally signals its inferiority to avoid being attacked or chased away by a stronger resident. Similar phenomenon was suggested in black kite (Milvus migrans) (Blas et al. 2013) and demonstrated in ring-necked pheasant (Phasianus colchicussuggest) (Mateos and Carranza 1997). Although we cannot rule out this possibility, an appeasing signal is commonly used at close proximity, whereas counter-singing by male hyraxes is mostly performed from a larger distance (often >50 m). In addition, a weaker individual with appeasing intentions would perform a dumbed-down signal to stress its inferiority, while in our case it is the opposite. Songs performed in counter-singing sessions are longer and more complex (Demartsev et al. 2014). Therefore, at this stage, the counter-singing interaction seems to fit better as an intended tournament and not an appeasing attempt.
However, at this point, we still cannot fully confirm the Napoleon strategy as the only explanation to our results. In order to achieve definite conclusions regarding the function of resident-bachelor counter singing patterns, the payoff to the participants should be further investigated. In addition, monitoring the life-long changes in counter-singing tactics would help better evaluation of social rank differences effect, as well as the relative importance of alternative factors (e.g., familiarity, relatedness, context, etc.).
In BB singing dyads, we could not identify a rank dependent counter-singing strategy among males. Possibly, since there are no clear resource limitations for male hyrax in Ein Gedi, and little if any ownership-based asymmetry exists between bachelors, BB interactions could be subject to different competition rules then BR and RB dyads. Factors like relatedness (Innocent et al. 2011), hormonal levels (Adkins-Regan 2005), personality (Dirienzo and Hedrick 2014), and social environment (Sih et al. 2009) may drive BB counter-singing tactics.
Concluding remarks
Collectively, our current results add to the previous body of work on male hyrax singing and reveal a set of tactics behind hyrax male-male counter-singing. This mixed signaling strategy exhibited by male hyrax is indicative of their cognitive ability to receive information regarding their opponents’ traits, by personal identification of the signaler and possibly by direct decoding of its song. In addition, male hyraxes seem to assess their own rank and residency relative to that of their opponent and to behave accordingly. However, our current results show only a general strategic frame of male hyrax vocal contests. Additional factors such as previous encounters, eavesdropping, and audience composition may also have an impact and potentially reveal a much finer scale of tactical choices made in male hyrax counter-singing. Hence, further research is needed in order to better understand the extent and the rates that hyraxes can perceive information from conspecific vocal signals.
References
Adkins-Regan E (2005) Hormones and animal social behavior. Princeton University Press, Princeton
Aragon P, Lopez P, Martin J (2006) Roles of male residence and relative size in the social behavior of Iberian rock lizards, Lacerta monticola. Behav Ecol Sociobiol 59:762–769
Bar Ziv E, Ilany A, Demartsev V, Barocas A, Geffen E, Koren L (2016) Individual, social, and sexual niche traits affect copulation success in a polygynandrous mating system. Behav Ecol Sociobiol 70:901–912
Barlow G, Rogers W, Fraley N (1986) Do Midas cichlids win through prowess or daring? It depends. Behav Ecol Sociobiol 19:1–8
Barocas A, Ilany A, Koren L, Kam M, Geffen E (2011) Variance in centrality within rock hyrax social networks predicts adult longevity. PLoS One 6:e22375
Behr O, Knörnschild M, von Helversen O (2009) Territorial counter-singing in male sac-winged bats (Saccopteryx bilineata): low-frequency songs trigger a stronger response. Behav Ecol Sociobiol 63:433–442
Bergman M, Olofsson M, Wiklund C (2010) Contest outcome in a territorial butterfly: the role of motivation. Proc R Soc Lond B 277:3027–3033
Blas J, Cabezas S, Figuerola J, Lopez L, Tanferna A, Hiraldo F (2013) Carotenoids and skin coloration in a social raptor. J Raptor Res 47:174–184
Bolyard KJ, Rowland WJ (2000) The effects of spatial context and social experience on the territorial aggression of male threespine stickleback. Behaviour 137:845–864
Demartsev V, Kershenbaum A, Ilany A, Barocas A, Bar Ziv E, Koren L, Geffen E (2014) Male hyraxes increase song complexity and duration in the presence of alert individuals. Behav Ecol 25:1451–1458
Dirienzo N, Hedrick AV (2014) Animal personalities and their implications for complex signaling. Curr Zool 60:381–386
Elias DO, Botero CA, Andrade MCB, Mason AC, Kasumovic MM (2010) High resource valuation fuels “desperado” fighting tactics in female jumping spiders. Behav Ecol 21:868–875
Fischer J, Kitchen D, Seyfarth R, Cheney D (2004) Baboon loud calls advertise male quality: acoustic features and their relation to rank, age, and exhaustion. Behav Ecol Sociobiol 56:140–148
Fourie PB (1977) Acoustic communication in the Rock Hyrax (Procavia capensis. Z Tierpsychol 44:194–219
Frommolt K, Goltsman M, MacDonald D (2003) Barking foxes, Alopex lagopus: field experiments in individual recognition in a territorial mammal. Anim Behav 65:509–518
Gammell MP, De Vries H, Jennings DJ, Carlin CM, Hayden TJ (2003) David's score: a more appropriate dominance ranking method than Clutton-Brock et al.'s index. Anim Behav 66:601–605
Getty T (1987) Dear enemies and the prisoners-dilemma—why should territorial neighbors form defensive coalitions. Am Zool 27:327–336
Getty T (1989) Are dear enemies in a war of attrition? Anim Behav 37:337–339
Grafen A (1987) The logic of divisively asymmetric contests—respect for ownership and the desperado effect. Anim Behav 35:462–467
Hurd PL (2006) Resource holding potential, subjective resource value, and game theoretical models of aggressiveness signalling. J Theor Biol 241:639–648
Ilany A, Barocas A, Koren L, Kam M, Geffen E (2011) Do singing rock hyraxes exploit conspecific calls to gain attention? PLoS One 6:e28612
Ilany A, Barocas A, Kam M, Ilany T, Geffen E (2013) The energy cost of singing in wild rock hyrax males: evidence for an index signal. Anim Behav 85:995–1001
Innocent TM, West SA, Sanderson JL, Hyrkkanen N, Reece SE (2011) Lethal combat over limited resources: testing the importance of competitors and kin. Behav Ecol 22:923–931
Jennings DJ, Elwood RW, Carlin CM, Hayden TJ, Gammell MP (2012) Vocal rate as an assessment process during fallow deer contests. Behav Process 91:152–158
Jenssen TA, Decourcy KR, Congdon JD (2005) Assessment in contests of male lizards (Anolis carolinensis): how should smaller males respond when size matters? Anim Behav 69:1325–1336
Just W, Morris MR (2003) The Napoleon Complex: why smaller males pick fights. Evol Ecol 17:509–522
Kitchen DM, Seyfarth RM, Fischer J, Cheney DL (2003) Loud calls as indicators of dominance in male baboons (Papio cynocephalus ursinus. Behav Ecol Sociobiol 53:374–384
Kokko H, Rintamaki PT, Alatalo RV, Hoglund J, Karvonen E, Lundberg A (1999) Female choice selects for lifetime lekking performance in black grouse males. Proc R Soc Lond B 266:2109–2115
Kokko H, Lopez-Sepulcre A, Morrell LJ (2006) From hawks and doves to self-consistent games of territorial behavior. Am Nat 167:901–912
Koren L, Geffen E (2009) Complex call in male rock hyrax (Procavia capensis): a multi-information distributing channel. Behav Ecol Sociobiol 63:581–590
Koren L, Geffen E (2011) Individual identity is communicated through multiple pathways in male rock hyrax (Procavia capensis) songs. Behav Ecol Sociobiol 65:675–684
Koren L, Mokady O, Geffen E (2008) Social status and cortisol levels in singing rock hyraxes. Horm Behav 54:212–216
Lindstrom K (2001) Effects of resource distribution on sexual selection and the cost of reproduction in sandgobies. Am Nat 158:64–74
Lotem A (1998) Higher levels of begging behavior by small nestlings: a case of a negatively correlated handicap. Isr J Zool 44:29–45
Mateos C, Carranza J (1997) The role of bright plumage in male-male interactions in the ring-necked pheasant. Anim Behav 54:1205–1214
Maynard-Smith J, Parker GA (1976) Logic of asymmetric contests. Anim Behav 24:159–175
Michod RE, Hasson O (1990) On the evolution of reliable indicators of fitness. Am Nat 135:788–808
Miyai CA, Carretero Sanches FH, Costa TM, Colpo KD, Volpato GL, Barreto RE (2011) The correlation between subordinate fish eye colour and received attacks: a negative social feedback mechanism for the reduction of aggression during the formation of dominance hierarchies. Zoology 114:335–339
Morrell LJ, Lindstrom J, Ruxton GD (2005) Why are small males aggressive? Proc R Soc Lond B 272:1235–1241
Olsson M, Shine R (2000) Ownership influences the outcome of male-male contests in the scincid lizard, Niveoscincus microlepidotus. Behav Ecol 11:587–590
Reby D, McComb K, Cargnelutti B, Darwin C, Fitch WT, Clutton-Brock T (2005) Red deer stags use formants as assessment cues during intrasexual agonistic interactions. Proc R Soc Lond B 272:941–947
Reddon AR, Balk D, Balshine S (2013) Probing aggressive motivation during territorial contests in a group-living cichlid fish. Behav Process 92:47–51
Russ BE, Comins JA, Smith R, Hauser MD (2010) Recognizing and respecting claims over resources in free-ranging rhesus monkeys, Macaca mulatta. Anim Behav 80:563–569
Schradin C, Schneider C, Lindholm AK (2010) The nasty neighbour in the striped mouse (Rhabdomys pumilio) steals paternity and elicits aggression. Front Zool 7:19
Searcy WA, Beecher MD (2009) Song as an aggressive signal in songbirds. Anim Behav 78:1281–1292
Shuster SM, Wade MJ (2003) Mating systems and strategies. Princeton University Press, Princeton
Sih A, Hanser SF, McHugh KA (2009) Social network theory: new insights and issues for behavioral ecologists. Behav Ecol Sociobiol 63:975–988
Smith IP, Huntingford FA, Atkinson RJA, Taylor AC (1994) Strategic decisions during agonistic behaviour in the velvet swimming crab (Necora puber L. Anim Behav 47:885–894
Svensson PA, Lehtonen TK, Wong BBM (2012) A high aggression strategy for smaller males. PLoS One 7:e43121
Temeles EJ (1994) The role of neighbors in territorial systems—when are they dear enemies. Anim Behav 47:339–350
Umbers KDL, Osborne L, Keogh JS (2012) The effects of residency and body size on contest initiation and outcome in the territorial dragon, Ctenophorus decresii. PLoS One 7:e47143
Yack JE, Gill S, Drummond-Main C, Sherratt TN (2014) Residency duration and shelter quality influence vibratory signalling displays in a territorial caterpillar. Ethology 120:354–364
Ydenberg RC, Giraldeau LA, Falls JB (1988) Neighbours, strangers, and the asymmetric war of attrition. Anim Behav 36:343–347
Acknowledgments
We wish to thank two anonymous reviewers for their constructive remarks on the earlier versions of this paper. We are grateful to the Ein Gedi Nature Reserve and Ein Gedi Field School staff for their hospitality and logistic support. We appreciate the valuable help in the field received from all field technicians, project students, and guests. We are grateful to the Nature and Park Authority for permission to work in the Ein Gedi Nature Reserve. We thank M. Samuni-Blank for a fruitful discussion on early drafts of the manuscript and N. Paz for her editorial services.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Funding
This study was supported by three grants from the Israel Science Foundation (577/99, 488/05, 461/09).
Conflict of interest
The authors declare that they have no conflict of interests.
Ethical approval
This study was conducted under annual permits from the Israeli Nature and Parks Authority (NPA) for capturing, handling, and sampling the hyraxes at the Ein Gedi Nature Reserve (2000/8871, 2001/8871, 2002/14674, 2003/14674, 2004/17687, 2005/17687, 2007/27210, 2008/31138, 2009/32871, 2010/37520, 2011/38061, 2012/38400, 2013/38803, 2014/40185). All procedures performed in this study involving animals were in accordance with the ethical standards of the NPA. During the whole course of this 15-year field study, no observable long-term stress or interference effects were detected to individual animals or the whole study population. Both general population numbers and the integrity of the specific social groups in the research area remained stable.
Additional information
Communicated by C. Voigt
Rights and permissions
About this article

Cite this article
Demartsev, V., Ilany, A., Barocas, A. et al. A mixed strategy of counter-singing behavior in male rock hyrax vocal competitions. Behav Ecol Sociobiol 70, 2185–2193 (2016). https://doi.org/10.1007/s00265-016-2222-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00265-016-2222-z