
Abstract
A novel antimicrobial peptide named NP-6 was identified in our previous work. Here, the mechanisms of the peptide against Escherichia coli (E. coli) were further investigated, as well as the peptide’s resistance to temperature, pH, salinity, and enzymes. The transmission electron microscopy (TEM), confocal laser scanning microcopy (CLSM), and flow cytometric (FCM) analysis, combined with measurement of released K+, were performed to evaluate the effect of NP-6 E. coli cell membrane. The influence of NP-6 on bacterial DNA/RNA and enzyme was also investigated. The leakage of K+ demonstrated that NP-6 could increase the permeability of E. coli cell membrane. The ATP leakage, FCM, and CLSM assays suggested that NP-6 caused the disintegration of bacterial cell membrane. The TEM observation indicated that NP-6 could cause the formation of empty cells and debris. Besides, the DNA-binding assay indicated that NP-6 could bind with bacterial genomic DNA in a way that ethidium bromide (EB) did, and suppress the migration of DNA/RNA in gel retardation. Additionally, NP-6 could also affect the activity of β-galactosidase. Finally, the effect of different surroundings such as heating, pH, ions, and protease on the antimicrobial activity of NP-6 against E. coli was also investigated. Results showed that the peptide was heat stable in the range of 60~100 °C and performed well at pH 6.0~8.0. However, the antimicrobial activity of NP-6 decreased significantly in the presence of Mg2+/Ca2+, and after incubation with trypsin/proteinase K. The results will provide a theoretical support in the further application of NP-6.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Antimicrobial peptides (AMPs) play an important role in the immune system of all living organisms, and have attracted a great deal of attention due to the emergence of bacterial resistance. Nowadays, more than 2,500 natural AMPs have been isolated from animals, plants, bacteria, and fungi (Wang et al. 2016). Generally, they share characteristics such as hydrophobic and always positively charged (Drin and Antonny 2010), which in turn associates with their activity (Hancock and Sahl 2006). Other characteristics including size, primary sequence, conformation, and structure are also essential for antimicrobial activity and action mode of AMPs (Guilhelmelli et al. 2013).
The mechanism is affected by the varying composition and structure of AMPs (Li et al. 2014). The classic, namely, membrane damage effect, has been widely accepted as key point (Nguyen et al. 2011; Hancock 2011; Malmsten 2016). It is generally believed that the positively charged AMPs firstly bind with the bacterial cell membrane which is composed of negatively charged lipids, leading to penetration of the cell membrane (Li et al. 2012). However, a large number of papers’ results indicated that AMPs have intracellular targets (Aerts et al. 2008; Guilhelmelli et al. 2013; Mahlapuu et al. 2016). In this case, the death-inducing mechanisms may involve in the interaction with DNA/RNA, then negatively impact protein synthesis and activity of intracellular enzyme, or inhibit the formation of cell wall/membrane (Ageitos et al. 2017).
Sichuan pepper (Zanthoxylum bungeanum Maxim) is an aromatic tree and shrub plant belonging to the genus Zanthoxylum of the family Rutaceae (Tang et al. 2014a). The pericarps of Z. bungeanum Maxim are normally used as a spice in Chinese kitchen, as well as a folk medicine for treatment of vomiting, toothache, stomachache, abdominal pain, and infected wounds (Xia et al. 2011). So far, more than 140 compounds have been identified from the plant, and some biological activities such as anti-inflammatory, antioxidant, and antimicrobial effects have been proved (Zhang et al. 2017). However, being a by-product of Z. bungeanum Maxim production, Z. bungeanum Maxim seeds which are rich in oil, protein, melanin, and dietary fiber are often discard. To the best of our knowledge, reports referring to the bioactive compounds in Z. bungeanum Maxim seeds are limited. Besides, the topics of these reports often focus on Z. bungeanum Maxim seed oil which is rich in polyunsaturated fatty acids (Tang et al. 2014a; Yang et al. 2014; Xia et al. 2011). Actually, Z. bungeanum Maxim seeds are also a good resource of protein. Previous report had proven that the hydrolysate of Z. bungeanum Maxim seed protein exhibited inhibitory activity against angiotensin-I-converting enzyme (ACE) (Wu et al. 2016). Jiang et al. (2015) reported that two antimicrobial fractions of Z. bungeanum Maxim seed protein hydrolysate were active against E. coli and S. aureus. So more investigation should be conducted to fully explore the bioactive compounds in Z. bungeanum Maxim seeds. More recently, there have been literatures reporting the medicinal characters of Z. bungeanum Maxim seed oil (Tang et al. 2014a; Yang et al. 2014), but limited attention has been paid to the protein/peptides.
In our previous work, it was found that a peptide (NP-6) being a fragment of glyceraldehyde 3-phosphate dehydrogenase (GAPDH) of Z. bungeanum Maxim seed protein exhibited antimicrobial activity (Hou et al. 2019). In the present work, the mechanisms of NP-6 against Escherichia coli were investigated, which were involved in damage effects of bacteria cell membrane, interaction with genomic DNA/RNA, and effects on activities of intracellular enzymes. Meanwhile, peptide resistance to temperature, pH, salinity, and enzymes was determined by monitoring the changes in antimicrobial activity against E. coli after being heated at various temperature, exposed in solutions with different pH and monovalent/divalent cations, and incubated with different protease. Results of the work will help a lot in clarifying the action mode of NP-6 and considering the application of the peptide in a suitable way.
Materials and methods
Materials
The peptide NP-6 was chemically synthesized according to the standard fluorenylmethyloxycarbonyl (Fmoc) methodology at GL Biochemistry Corporation (Shanghai, China), followed by further purification with RP-HPLC system and characterization with ESI-MS (Waters, Madrid, Spain), yielding final purity of the peptide as more than 95%. Propidium iodide (PI) was purchased from Sigma (St. Louis, MO, USA). SYTO 9 (S34854) was purchased from Molecular Probes (Invitrogen, Barcelona, Spain). Pepsin (25000 U/mg), trypsin (2500 U/mg), proteinase K (40 U/mg), ethidium bromide (EB), and o-nitrophenyl-β-d-galactopyranoside (ONPG) were purchased from Sangon Biotech Co., Ltd. (Shanghai, China). DNA extraction kit was purchased from TransGen Biotech Co., Ltd. (Beijing, China). One-stop RNA electrophoresis pack was purchased from Shanghai Biya Biotechnology Co., Ltd. (Shanghai, China). The T-ATPase was purchased from Jiancheng Bioengineering Institute (Nangjing, China). Enhanced ATP Assay Kit and BCA protein assay kit were obtained from Beyotime Biotechnology (Shanghai, China). All other chemicals were of analytical grade.
Strains and growth conditions
Escherichia coli (E. coli, ATCC 25922) were obtained from China Center of Industrial Culture Collection (Beijing, China), cultured in sterilized nutrient broth (NB, peptone, 10 g/l, yeast extract, 5 g/l, NaCl, 10 g/l) at 37 °C.
Leakage of K+
E. coli was cultured in NB media at 37 °C for 16~18 h; the cells were centrifuged, washed twice with deionized water, and diluted to 106 CFU/ml. NP-6 solution was added to a finial concentration of 1 × MIC (64 μg/ml) and the mixture were incubated at 37 °C for various times, with deionized water–treated cells as control. Samples were centrifuged at 13400×g for 3 min and the amount of released K+ in the supernatants was measured on an atomic absorption spectrometer (Spectr AA 220 FS, Varian, USA) according to a previous description (Lu et al. 2011).
ATP leakage
The assessment of ATP leakage was performed according to Gottschalk (2017). The bacterial strain E. coli was incubated in NB overnight at 37 °C and grown to mid-exponential phase. Five milliliters of bacterial cells was harvested by centrifugation at 2000×g for 10 min. After being washed twice in 50 mM HEPES buffer, bacterial cells were resuspended in the same buffer (1 ml) and then energized with 0.2% (w/v) glucose at 37 °C for 10 min. Cell suspension was treated with NP-6 (finial concentration 64 μg/ml and 1 mg/ml) for 0, 15, 30, 45, and 60 min. The samples were then centrifuged at 2000×g for 10 min, and the supernatant was collected for estimating the leakage of ATP from the cells by using Beyotime Enhanced ATP Assay Kit.
Flow cytometry
Log-phased cells of E. coli were harvested, washed, and resuspended in PBS. Solutions of NP-6 were added to a final concentration of 1.0 mg/ml, and the mixture was incubated at 37 °C for 1 h, with PBS treatment as a control. Then cells were centrifuged and washed by PBS, followed by being incubated with PI (50 μg/ml solution in dd H2O) at 4 °C for 30 min in darkness (Sik et al. 2006). The samples were centrifuged at 13400×g for 3 min, washed three times with PBS, and then resuspended in the same buffer. Analysis was performed using a FACSCalibur flow cytometer (Becton Dickinson, USA).
Confocal laser scanning microcopy
The assay was performed using a method according to Qian et al. (2011) with some modification. A working solution containing two dyes was employed to stain E. coli in the study. 1.5 μl SYTO 9 (5 mM solution in DMSO) was mixed with the same volume of PI (50 μg/ml solution in ddH2O), then 1 ml PBS was added to the mixture and the solution was prepared fresh daily. E. coli cells treated by NP-6 (1.0 mg/ml) were incubated with the working solution for 30 min in the dark at 4 °C; cells without treatment of NP-6 were set as control. The stained E. coli was observed under a SP2 confocal microscope (Leica, German) after washing three times with PBS. Images were collected at an excitation/emission wavelength of 485/498 nm for SYTO 9 signal and 535/617 nm for PI signal.
Transmission electron microscopy
Log-phased cells of E. coli were treated by NP-6 (1.0 mg/ml) for 1 h at 37 °C, followed by being centrifuged and wash twice with phosphate-buffered saline (PBS, 0.01 M, pH 7.4). PBS-treated cells were set as the control. The collected cells were firstly fixed with 2.5% (v/v) glutaraldehyde for 3 h, then post-fixed with 1% osmium tetroxide for another 2 h; both were conducted at 4 °C (Shi et al. 2016). Cells were then dehydrated by a series of ethanol and embedded in resin. The cut sections of samples were stained with uranyl acetate and lead citrate. Analysis were conducted by a transmission electron microscopy system (Tecnai G2 F20 S-TWIN, USA).
Extraction of bacterial DNA/RNA
The DNA of E. coli was prepared using a DNA extraction kit (TransGen Biotech, Beingjing). Total RNA was purified using a one-stop RNA electrophoresis pack (Biya Biotech, Shanghai).
Competitive binding of NP-6 and EB with bacterial DNA
The assay was performed as described by Li et al. (2013) with little modification. DNA of E. coli was firstly dissolved in TE buffer (10 mM, pH 8.0) to a final concentration of 50 μg/ml, then 15 μl EB solution was added and the mixture was incubated at 37 °C for 10 min in the dark. Various concentrations of NP-6 (0, 1.0, 2.0, and 4.0 mg/ml) were added to the mixture. After being incubated in the dark at 37 °C for 30 min, the fluorescence of the solution was measured using a Lumina Fluorescence Spectrophotometer (Thermo Fisher Scientific, MA, USA). The excitation wavelength was set at 535 nm, and the spectra were recorded at 550~750 nm.
DNA/RNA gel retardation
Gel retardation assays were conducted according to a previous literature (Shi et al. 2016) with some modification. Varying concentrations of NP-6 (0, 0.1, 0.2, 0.4, 0.8, 1.0, 2.0, and 4.0 mg/ml) were added to the DNA solution, and the mixture was incubated at 37 °C for 30 min. After that, 2 μl binding buffer (5% glycerol, 10 mM Tris-HCl (pH 8.0), 1 mM EDTA, 1 mM dl-dithiothreitol, 20 mM KCl, and 50 μg/ml albumin) was added and the reaction mixture was subjected to gel electrophoresis on a 1% agarose gel. For RNA binding, various concentrations of NP-6 (0, 0.1, 0.2, 0.4, 0.8, 1.0, 2.0, and 4.0 mg/ml, dissolved in diethyl pyrocarbonate (DEPC)–treated water) were incubated with bacterial RNA for 10 min, and the formaldehyde denaturing gel electrophoresis was conducted using an one-stop RNA electrophoresis kit according to the manufacturer’s recommendations.
Effect of NP-6 on the activity of T-ATPase
The assay was performed using a previous method (Shi et al. 2016) with minor modification. E. coli was incubated with NP-6 (0.5, 1.0, and 2.0 mg/ml) at 37 °C for 1 h, with PBS as a control. The culture was centrifuged at 13400×g for 5 min, washed three times with PBS and resuspended in the same buffer. Then samples were disrupted by sonication at 300 W for 5 min in an ice bath. The bacterial total protein content was determined using a BCA protein assay kit. The T-ATPase level was measured using a commercial assay kit according to the recommendations of the manufacturer. The samples were scanned by a microplate reader (Dinex Technologies Inc., Chantilly, USA) at 636 nm.
Effect of NP-6 on the activity of β-galactosidase
Lactose can be decomposed into one glucose and one galactose and is used as substrate for β-galactosidase determination. ONPG and lactose have similar structures, and also can be decomposed by β-galactosidase into yellowish O-nitrophenol (ONP). The amount of ONP released can be measured to determine the activity of β-galactosidase. Log-phased cells of E. coli were collected, centrifuged, washed, and resuspended in M9 media, incubated for 8 h at 37 °C. Then cells were centrifuged at 13400×g for 5 min, washed three times with PBS, and resuspended in the same buffer. After that, NP-6 was added to a various concentrations of 0.5, 1.0, and 2.0 mg/ml, followed by being incubated at 37 °C for 1 h, with PBS-treated cells as the control. The culture was then centrifuged, washed, resuspended in PBS, and finally disrupted by sonication at 300 W for 5 min in an ice bath. Samples were further centrifuged and 20 μl of the bacterial suspension mixed with 80 μl ONPG (5 mM) was added to the well of a standard 96-well plate. The plates were incubated at 37 °C for 10 min and the reaction was stopped by adding 100 μl 0.5 M cold Na2CO3 and scanned by a microplate reader (Dinex Technologies Inc., Chantilly, USA) at 420 nm (Kragol et al. 2001). Determination of protein content was done using BCA protein assay kit.
One unit of β-galactosidase was defined as the hydrolysis to produce 1 μmol ONP per minute and labeled as 1 U.
Resistance of NP-6 to temperature, pH, salinity, and enzymes
The surrounding sensitivity analysis was performed as described (Tang et al. 2015) with minor modification. The recovered antimicrobial activities after treatment with different conditions (heat, pH, ions, and enzymes) were measured by monitoring the total numbers of colony. For the heating treatment, NP-6 was dissolved in distilled deionized water (dd H2O) to a final concentration of 1.0 mg/ml, then heated at 60, 70, 80, 90, and 100 °C for 30 min in a water bath. For the acid/alkali treatment, NP-6 was dissolved in dd H2O with different pH values (2.0, 4.0, 6.0, 8.0, and 10.0) for 15 min and then applied to pH 7.0. For ion sensitivity analysis, the peptide was exposed to different concentrations (50 mM, 100 mM, and 150 mM) of K+, Na+, Mg2+, and Ca2+. For enzyme sensitivity test, NP-6 was exposed in pepsin, trypsin, and proteinase K for 15, 30, and 60 min, followed by being heated at 100 °C for 5 min to terminate the reactions. NP-6 without any treatment was set as a control. The inhibition rate was calculated as follows:
where Ncontrol is the number of bacteria in untreated samples, Ntreated is the number of bacteria in treated samples.
Statistical analysis
All the tests were repeated at least three times. Data analysis was performed using the software of SPSS (version 19.0, IBM SPSS, USA). Results were expressed as the means ± standard deviation. The significance of the differences between the treatments and the respective controls were determined based on Student’s t test, and a value of p < 0.05 was considered statistically significant.
Results
Potassium efflux
In order to characterize the effect of NP-6 on permeability of E. coli cell membrane, the amount of released potassium from peptide-treated cells were determined, with untreated cells as a control. As shown in Fig. 1, after 1 h of treatment with 1 × MIC NP-6, the efflux of potassium was immediate. By comparison, the amount of released K+ of untreated E. coli cells was kept in a low level. The result could be the evidence that NP-6 increased the permeability of E. coli cell membrane.

Leakage of K+ from the peptide-treated E. coli cells at various times. The control was processed without NP-6
Detection of ATP leakage
Extracellular ATP levels were determined as indexes of cell lysis (shown in Fig. 2). In the presence of 1 × MIC NP-6, the concentration of extracellular ATP was maintained at a low level, but the increase of released ATP was observed after 2 h. Meanwhile, treatment with NP-6 (1 mg/ml) caused a significant increase in the concentration of extracellular ATP after 15 min. In contrast, concentration of extracellular ATP in untreated cells was maintained at approximately 9 nM. These results clearly indicated that NP-6 could disrupt the integrality of bacterial cell membrane and cause the release of large molecules.

Concentration of the released ATP from the peptide-treated E. coli cells at various times. The control was processed without NP-6
Flow cytometric analysis
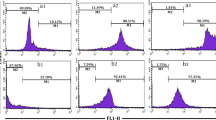
The effects of NP-6 on E. coli cell membrane were further investigated by flow cytometer with PI as a probe. PI is a DNA-intercalating fluorescent dye that can penetrate into impaired cells, but does not enter into intact cells. It is often used as a probe to investigate the integrity of cell membrane. This fluorescent probe will enter into the cytoplasm if the bacterial cell membrane is damaged. As shown in Fig. 3, only 6.68% of untreated E. coli cells showed PI fluorescence signal. By contrast, 91.52% peptide-treated cells were labeled fluorescence, indicating that the bacterial cell membrane integrity was impaired.

The membrane damage effect of NP-6 on E. coli cells by flow cytometry analysis, as measured by an increase of fluorescent intensity of PI (50 μg/ml). The control was processed without NP-6
CLSM images
The above results were confirmed by confocal laser scanning microcopy (CLSM) with SYTO 9 and PI as fluorescent probes. SYTO 9 is a kind of green fluorescent nuclear acid stain; it was proved to stain cells whether alive or dead. While PI is a red-fluorescent nuclear dyes, it is commonly used to detect dead cells due to its non-penetrability to alive cells. As was observed in Fig. 4, green dots on the left are all bacterial cells including alive and dead ones, red dots in the middle are dead cells, yellow dots on the right are dead cells, and green dots are alive ones. It was obvious that most of the untreated cells were alive while an overwhelming fraction of the peptide-treated E. coli cells were dead. The CLSM images were another evidence for the disintegrity of bacterial cell membrane.

Confocal images of E. coli treated and untreated by NP-6. The control was processed without NP-6
TEM observation
Transmission electron microscopy (TEM) was performed to investigate the ultrastructural changes of E. coli cells. As was illustrated in Fig. 5, in the absence of NP-6, the E. coli cells maintained an intact, smooth cell membrane and presented a dense internal organization. Upon exposure to NP-6 for 1 h, cell wall and membrane disruption were clearly observed and the leaked cytoplasmic materials were found around the cells. The result of cell membrane disruption followed by the treatment of NP-6 was consistent with the results obtained from CLSM and flow cytometry (FCM).

Transmission electron microscopy images of E. coli treated and untreated by NP-6. The control was processed without NP-6
DNA/RNA binding
The result of competitive binding of NP-6 and EB with bacterial genomic DNA is shown in Fig. 6a. It was found that the fluorescence of the free EB is the lowest, while bacterial DNA without treatment of NP-6 showed the highest fluorescence. With the increasing concentration of NP-6, a remarkable fluorescence decline of the EB-DNA system was observed. Lepecq and Paoletti (1967) pointed that the fluorescence could be quenched when there was a reagent that executed a similar reaction. The result indicated that NP-6 could bound to E. coli genomic DNA. In addition, EB molecules inserting into the bacterial DNA were replaced by NP-6, suggesting a stronger binding affinity of NP-6 than EB.

Effect of NP-6 on bacterial nucleic acid. a Competitive binding of NP-6 and EB with E. coli genomic DNA, and the fluorescence spectra were measured from 550 to 750 nm. b A gel retardation assay was conducted after various concentrations of NP-6 were incubated with E. coli genomic DNA. Bands 1–8 were for the concentrations of 0, 0.1, 0.2, 0.4, 0.8, 1.0, 2.0, and 4.0 mg/ml, respectively. c A gel retardation analysis of the binding activity of NP-6 to E. coli RNA. Various concentrations of NP-6 were incubated with RNA. Bands 1–8 were for the concentrations of 0, 0.1, 0.2, 0.4, 0.8, 1.0, 2.0, and 4.0 mg/ml, respectively.
A gel retardation assay was performed to further determine the DNA-binding activity of NP-6 (shown in Fig. 6b). It was observed that the lower the peptide concentration, the brighter the band of the peptide-DNA system. The band was almost trapped in the spotting hole when the peptide concentration was 0.2 mg/ml. The observations suggested that NP-6 had DNA-binding activity.
A gel retardation assay was conducted to estimate the RNA-binding activity of NP-6. As shown in Fig. 6c, the bacterial RNA was suppressed by NP-6 in a concentration-dependent manner. Meanwhile, a degradation of 23S rRNA was observed, and all the peptide-treated samples only had a band of 16S rRNA. The migration of RNA was almost completely suppressed when the concentration of NP-6 was 1.0 mg/ml. The results demonstrated that NP-6 had the ability to interact with E. coli RNA.
Effect of NP-6 on the activities of T-ATPase and β-galactosidase
In order to learn whether NP-6 can inhibit the activity of bacterial intracellular enzyme, the activity of T-ATPase and β-galactosidase in bacterial cells treated/untreated by NP-6 was determined. The results are shown in Fig. 7. It was observed that NP-6 did not significantly inhibit the T-ATPase in the range of 0~2.0 mg/ml, indicating that NP-6 had no effect on the T-ATPase of E. coli. However, it was also observed that NP-6 strongly inhibited the β-galactosidase activity of E. coli in a dose-dependent manner (8.72–1.24 U/mg). By the concentration of 2.0 mg/ml, the β-galactosidase activity of E. coli had been suppressed in a very low level compared with the samples without treatment of NP-6.

Effect of NP-6 on the activity of intracellular T-ATPase and β-galactosidase of E. coli
Resistance of NP-6 to temperature, pH, salinity, and enzymes
The surrounding sensitivity assays were performed to investigate the potential value of NP-6 as a natural preservative agent used in food. As shown in Fig. 8a, heating treatment in the range of 60~100 °C had little effect on the antimicrobial activity of NP-6 against E. coli. After heating for 30 min at 100 °C, the retained activity of NP-6 was more than 90%, indicating that the peptide was heat stable at 60~100 °C.

Recovered antimicrobial activity of NP-6 against E. coli after treatment by different conditions. a Heated at 60, 70, 80, 90, and 100 °C for 15 min; b exposed to solutions with different pH values; c in the presence of K+, Na+, Mg2+, and Ca2+; d incubation with pepsin, trypsin, and proteinase K for different times
When the pH was 6.0 and 8.0, the antimicrobial activity of NP-6 against E. coli was more than 90%, but it decreased when the pH was lower than 6.0 or higher than 8.0 (Fig. 8b). As the pH decreased to 2.0, the antimicrobial activity of NP-6 against E. coli reduced to 34.25%, and it reduced to 44.85% when the pH increased to 10.0. The results suggested that NP-6 had the strongest antimicrobial activity at physical pH, and acidic and basic conditions could inhibit its activity against E. coli.
As shown in Fig. 8c, the antimicrobial activity of NP-6 against E. coli was not significantly affected by Na+ and K+, but it was greatly suppressed in the presence of Mg2+ and Ca2+. The residual activity decreased to 15.96% and 20.85% when the concentration of Mg2+ and Ca2+ was 150 mM, and it almost lost its antimicrobial activity against E. coli. The results indicated that divalent ions had a stronger inhibitory effect on the antimicrobial activity of NP-6 than the monovalent ones.
Figure 8d showed the changes in antimicrobial activity of NP-6 after incubation with different enzymes in a time series. It was observed that all the three enzymes could cause decrease in antimicrobial activity of NP-6. The decrease caused by pepsin was slight, and the residual activity of NP-6 against E. coli was more than 70% after incubation for 60 min with pepsin. However, the antimicrobial activity of NP-6 was greatly suppressed after incubation with trypsin and proteinase K, and it was almost lost after long-time hydrolysis with the two enzymes. The antimicrobial activity of NP-6 reduced to 16.00% after incubation for 60 min with proteinase K, while it was 20.10% after incubation with trypsin for the same time. The results suggested that NP-6 was sensitive to the hydrolysis of trypsin and proteinase K, but was resistant to pepsin in a certain extent.
Discussion
We had previously reported that a novel antimicrobial peptide NP-6 derived from pepsin hydrolysis of Z. bungeanum Maxim seeds protein was active against some Gram-positive and Gram-negative bacteria (Hou et al. 2019). In the present work, the mechanisms of NP-6 against E. coli were further investigated, as well as the resistance of the peptide to temperature, pH, salinity, and enzymes. All the results will help a lot in the further application of NP-6 whether as a bio-preservative in food industry or as a therapy agent in medical science. It was said that the negatively charged bacterial cell surface was supposed to be the receptors of the positively charged AMPs (Hirakura et al. 2002).
Many AMPs exhibit their antimicrobial activities by targeting bacterial cell membrane firstly (da Costa et al. 2015, Nguyen et al. 2011, Anderson et al. 2004). Researchers have employed numerous methodology to investigate the effect of AMPs on bacterial cell membrane (Tao et al. 2011; Tang et al. 2014b; Rončević et al. 2019). Determining the amount of K+ released from the peptide-treated cells is a classical method for investigating the permeability of AMPs to bacterial cell membranes (Du et al. 2018). The amount of K+ released to the medium from peptide-treated cells was drastically increased after 1 h and remained stable in 2 h. This might be the evidence that NP-6 increased the permeability of the E. coli cell membrane. Similar results were obtained for bifidocin A (Liu et al. 2016), peptide F1 (Miao et al. 2016), and APP (Li et al. 2016). ATP levels in the medium were measured as indexes of cell lysis. When the peptide was in low concentration (1 × MIC, 64 μg/ml), the extracellular ATP contents of peptide-treated cells increased after 2 h. However, significant increase in K+ efflux (p < 0.05) was observed after only 1 h. The result suggested that the later release of ATP might be due to the early efflux of K+ as the cells consume available ATP in a futile attempt to re-accumulate K+ (Auliffe et al. 1998; Liu et al. 2016). Additionally, significant increase in the amount of ATP (p < 0.05) was observed within 15 min after incubation with NP-6 at a concentration of 1 mg/ml. These results suggested that NP-6 could increase the permeability of E. coli cell membrane at low concentration, thus induced the efflux of K+. However, it could also cause the bacterial cell membrane disintegration in high concentration followed by the release of large molecules such as ATP. The FCM, CLSM, and TEM results were also evidence for the point that NP-6 could cause the disruption of E. coli cell membrane.
Like many other AMPs, despite membrane-disruption effect, NP-6 have intracellular targets as well. In our study, the DNA/RNA-binding assays suggested that the peptide could interact with bacterial DNA/RNA and suppress its migration in vitro. Moreover, the decrease in fluorescence of the EB-DNA system indicated that NP-6 could compete with EB in binding to bacterial DNA, and replace some EB molecular embedded into the bacterial DNA. A similar result was found by Li et al. (2013). In fact, many other AMPs, such as melittin, indolicidin, and NKLP 27, are reported to have the ability to interact with bacterial genomic DNA (Hsu et al. 2005; Zhang et al. 2014; Shi et al. 2016). Besides, it was found that NP-6 could also inhibit the activities of β-galactosidase. Kragol et al. (2001) found that the antimicrobial peptide pyrrhocoricin strongly inhibited the ATPase and β-galactosidase of DnaK, thus prevented the chaperone-assisted protein folding. All these results suggested that NP-6 might exhibit multiple action modes to exert its antimicrobial activity.
Besides, NP-6 was a fragment of glycerol-3-phosphate dehydrogenase (GAPDH) through the protein that is generally associated with its glycolytic role (Branco et al. 2017). Actually, GAPDH also plays roles in many other different subcellular locations, apoptosis, and nuclear pathways (Silva et al. 2011; Nakajima et al. 2009). Besides, there have been a few publications reporting the antifungal/antibacterial activity of fragments of GAPDH. One was found secreted by Saccharomyces cerevisiae (Branco et al. 2017), and the other three were identified from the skin of yellow tuna (Seo et al. 2012), skipjack tuna (Seo et al. 2014), and human placental tissue (Wagener et al. 2013). All these observations indicated that GAPDH plays a significant role in defense system, further investigations are supposed to be carried out, to explore its extra function. Moreover, GAPDH-derived AMPs had not been identified except one.
It appeared that NP-6 was heat stable in the range of 60~100 °C. The reason that NP-6 exhibits extreme properties may be attributed to the small molecule weight and simple secondary structure. That is to say extreme environmental conditions such as temperature have little effect on the peptide’s structure because it is too simple, or it is easy for the peptide to recover its original structure in the absence of extreme conditions. The resistance of AMPs to high temperature has been reported by many literatures (Du et al. 2018; Kumar et al. 2018; Mahlapuu et al. 2016). This feature also makes AMPs have a wider range of applications. Meanwhile, the peptide performed well in a pH range of 6.0~8.0, and the antimicrobial activity decreased significantly when the pH value was higher than 8.0 or lower than 6.0. This may attribute to the different charged state of the peptide in solutions with different pH values, which would further have an effect on the interaction between NP-6 and E. coli cell membrane (Tang et al. 2015). Meanwhile, the antimicrobial activity of NP-6 was significantly decreased in the presence of Mg2+ and Ca2+. Tang et al. (2015) also declared that the divalent ions had a stronger inhibitory effect than the monovalent ones on the antimicrobial activity. Finally, the protease digestion assay indicated that the antimicrobial activity of NP-6 was significantly affected by trypsin and proteinase K, indicating that the peptide was sensitive to the hydrolysis of protease. A similar case was declared by Seo et al. (2014) who found that SJGAP almost lost all of its activity after treatment of trypsin.
In conclusion, all these results suggested that NP-6 could increase the permeability of E. coli cell membrane, cause the disruption of the bacterial cell membrane, bind with bacterial DNA/RNA, and inhibit the activity of β-galactosidase. But it is still unclear which way is the leading role and more assays should be conducted to further investigate it. What’s more, NP-6 was heat stable in the range of 60~100 °C and performed well in a pH range of 6.0~8.0. However, the peptide was sensitive to Mg2+ and Ca2+, as well as trypsin and proteinase K. All these results suggested that NP-6 could be a potential biological preservative for the agricultural and food industry.
References
Aerts AM, Franåois IEJA, Cammue BPA, Thevissen K (2008) Review the mode of antifungal action of plant , insect and human defensins. Cell Mol Life Sci 65:2069–2079. https://doi.org/10.1007/s00018-008-8035-0
Ageitos JM, Sánchez-pérez A, Calo-mata P, Villa TG (2017) Antimicrobial peptides ( AMPs ): ancient compounds that represent novel weapons in the fight against bacteria. Biochem Pharmacol 133:117–138. https://doi.org/10.1016/j.bcp.2016.09.018
Anderson RC, Hancock REW, Yu PL (2004) Antimicrobial activity and bacterial-membrane interaction of ovine-derived Cathelicidins. Antimicrob Agents Chemother 48:673–676. https://doi.org/10.1128/AAC.48.2.673
Auliffe OMC, Ryan MP, Ross RP, Hill C, Breeuwer P, Abee T (1998) Lacticin 3147 , a broad-spectrum bacteriocin which selectively dissipates the membrane potential. Appl Environ Microbiol 64:439–445
Branco P, Francisco D, Monteiro M, Almeida MG, Caldeira J, Arneborg N, Prista C, Albergaria H (2017) Antimicrobial properties and death-inducing mechanisms of saccharomycin, a biocide secreted by Saccharomyces cerevisiae. Appl Microbiol Biotechnol 101:159–171. https://doi.org/10.1007/s00253-016-7755-6
da Costa JP, Cova M, Ferreira R, Vitorino R (2015) Antimicrobial peptides: an alternative for innovative medicines? Appl Microbiol Biotechnol 99:2023–2040. https://doi.org/10.1007/s00253-015-6375-x
Drin G, Antonny B (2010) Amphipathic helices and membrane curvature. FEBS Lett 584:1840–1847. https://doi.org/10.1016/j.febslet.2009.10.022
Du H, Yang J, Lu Z, Bie X, Zhao H, Zhang C, Lu F (2018) Purification , characterization , and mode of action of plantaricin GZ1-27 , a novel bacteriocin against Bacillus cereus. J Agric Food Chem 66:4716–4724. https://doi.org/10.1021/acs.jafc.8b01124
Gottschalk S (2017) The interaction of antimicrobial peptides with the membrane and intracellular targets of Staphylococcus aureus investigated by ATP leakage, DNA-binding analysis, and the expression of a LexA-controlled gene, recA. In: Antimicrobial Peptides. Humana Press, New York, pp 297–305
Guilhelmelli F, Vilela N, Albuquerque P, Derengowski LS (2013) Antibiotic development challenges : the various mechanisms of action of antimicrobial peptides and of bacterial resistance. Front Microbiol 4:1–12. https://doi.org/10.3389/fmicb.2013.00353
Hancock REW (2011) Multifunctional cationic host defence peptides and their clinical applications. Cell Mol Life Sci 68:2161–2176. https://doi.org/10.1007/s00018-011-0710-x
Hancock REW, Sahl H (2006) Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat Biotechnol 24:1551–1557. https://doi.org/10.1038/nbt1267
Hirakura Y, Kobayashi S, Matsuzaki K (2002) Specific interactions of the antimicrobial peptide cyclic h -sheet tachyplesin I with lipopolysaccharides. Biochim Biophys Acta - Biomembr 1562:32–36. https://doi.org/10.1016/S0005-2736(02)00358-9
Hou XY, Li SS, Luo QY, Shen GH, Wu HJ, Li ML, Liu XY, Chen AJ, Ye M, Zhang ZQ (2019) Discovery and identification of antimicrobial peptides in Sichuan pepper (Zanthoxylum bungeanum Maxim) seeds by peptidomics and bioinformatics. Appl Microbiol Biotechnol 103:1–12. https://doi.org/10.1007/s00253-018-09593-y
Hsu C, Chen C, Jou M, Lee AY, Lin Y, Yu Y, Huang W, Wu S (2005) Structural and DNA-binding studies on the bovine antimicrobial peptide , indolicidin : evidence for multiple conformations involved in binding to membranes and DNA. Nucleic Acids Res 33:4053–4064. https://doi.org/10.1093/nar/gki725
Jiang T, Wu H, Dong X, Chen S, Shen G, Zhang Z (2015) Isolation and purification of antimicrobial peptides from prickly ash (Zanthoxylum bungeanum Maxim) seed protein. China Oils Fats 40:20–24
Kragol G, Lovas S, Varadi G, Condie BA, Hoffmann R, Otvos L (2001) The antibacterial peptide pyrrhocoricin inhibits the ATPase actions of DnaK and prevents chaperone-assisted protein folding. Biochemistry 40:3016–3026. https://doi.org/10.1021/bi002656a
Kumar P, Kizhakkedathu JN, Straus SK (2018) Antimicrobial peptides : diversity , mechanism of action and strategies to improve the activity and biocompatibility in vivo. Biomolecules 8:4–28. https://doi.org/10.3390/biom8010004
LePecq JB, Paoletti C (1967) A fluorescent complex between ethidium and nucleic acids bromide. J Mol Biol 27:87–106. https://doi.org/10.1016/0022-2836(67)90353-1
Li Y, Xiang Q, Zhang Q, Huang Y, Su Z (2012) Peptides overview on the recent study of antimicrobial peptides : origins , functions , relative mechanisms and application. Peptides 37:207–215. https://doi.org/10.1016/j.peptides.2012.07.001
Li L, Shi Y, Cheserek MJ (2013) Antibacterial activity and dual mechanisms of peptide analog derived from cell-penetrating peptide against Salmonella typhimurium and Streptococcus pyogenes. Appl Microbiol Biotechnol 97:1711–1723. https://doi.org/10.1007/s00253-012-4352-1
Li W, Tailhades J, O’Brien-Simpson NM, Separovic F, Otvos L, Hossain MA, Wade JD (2014) Proline-rich antimicrobial peptides: potential therapeutics against antibiotic-resistant bacteria. Amino Acids 46:2287–2294
Li L, Sun J, Xia S, Tian X (2016) Mechanism of antifungal activity of antimicrobial peptide APP , a cell-penetrating peptide derivative , against Candida albicans : intracellular DNA binding and cell cycle arrest. Appl Microbiol Biotechnol 100:3245–3253. https://doi.org/10.1007/s00253-015-7265-y
Liu G, Song Z, Yang X, Gao Y, Wang C (2016) Antibacterial mechanism of bifidocin A , a novel broad-spectrum bacteriocin produced by Bifidobacterium animalis BB04. Food Control 62:309–316. https://doi.org/10.1016/j.foodcont.2015.10.033
Lu C, Wang H, Lv W, Lou Z (2011) Antibacterial properties of anthraquinones extracted from rhubarb against Aeromonas hydrophila. Fish Sci 77:375–384. https://doi.org/10.1007/s12562-011-0341-z
Mahlapuu M, Håkansson J, Ringstad L, Björn C (2016) Antimicrobial peptides : an emerging category of therapeutic agents. Front Cell Infect Microbiol 6:1–12. https://doi.org/10.3389/fcimb.2016.00194
Malmsten M (2016) Interactions of antimicrobial peptides with bacterial membranes and membrane components. Curr Top Med Chem 16:16–24
Miao J, Zhou J, Liu G, Chen F, Chen Y, Gao X, Dixon W, Song M, Xiao H, Cao Y (2016) Membrane disruption and DNA binding of Staphylococcus aureus cell induced by a novel antimicrobial peptide produced by Lactobacillus paracasei subsp . tolerans FX-6. Food Control 59:609–613. https://doi.org/10.1016/j.foodcont.2015.06.044
Nakajima H, Amano W, Fukuhara A, Ihara H, Azuma Y, Tajima H (2009) Glyceraldehyde-3-phosphate dehydrogenase aggregate formation participates in oxidative stress-induced cell death. J Biol Chem 284(49):34331–34341. https://doi.org/10.1074/jbc.M109.027698
Nguyen LT, Haney EF, Vogel HJ (2011) The expanding scope of antimicrobial peptide structures and their modes of action. Trends Biotechnol 29:464–472. https://doi.org/10.1016/j.tibtech.2011.05.001
Qian L, Xiao H, Zhao G, He B (2011) Synthesis of modified guanidine-based polymers and their antimicrobial activities revealed by AFM and CLSM. Appl Mater interfaces 3:1895–1901. https://doi.org/10.1021/am200094u
Rončević T, Vukičević D, Krce L, Benincase M, Aviani I, Maravić A, Tossi A (2019) Selection and redesign for high selectivity of membrane-active antimicrobial peptides from a dedicated sequence/function database. BBA - Biomembr 1861:827–834. https://doi.org/10.1016/j.bbamem.2019.01.017
Seo J-K, Lee MJ, Go H-J, Park TH, Park NG (2012) Purification and characterization of YFGAP, a GAPDH-related novel antimicrobial peptide, from the skin of yellowfin tuna, Thunnus albacares. Fish Shellfish Immunol 33:743–752. https://doi.org/10.1016/j.fsi.2012.06.023
Seo J, Jeong M, Go H, Jun Y, Gyu N (2014) Antimicrobial function of the GAPDH-related antimicrobial peptide in the skin of skipjack tuna , Katsuwonus pelamis. Fish Shellfish Immunol 36:571–581. https://doi.org/10.1016/j.fsi.2014.01.003
Shi W, Li C, Li M, Zong X, Han D, Chen Y (2016) Antimicrobial peptide melittin against Xanthomonas oryzae pv. oryzae, the bacterial leaf blight pathogen in rice. Appl Microbiol Biotechnol 100:5059–5067. https://doi.org/10.1007/s00253-016-7400-4
Sik W, Ki H, Young K, Am S, Soo Y, Hee I (2006) Antifungal activity of synthetic peptide derived from halocidin, antimicrobial peptide from the tunicate, Halocynthia aurantium. FEBS Lett 580:1490–1496. https://doi.org/10.1016/j.febslet.2006.01.041
Silva A, Almeida B, Sampaio-marques B, Reis MIR, Ohlmeier S, Rodrigues F (2011) Glyceraldehyde-3-phosphate dehydrogenase ( GAPDH ) is a speci fi c substrate of yeast metacaspase. BBA - Mol Cell Res 1813:2044–2049. https://doi.org/10.1016/j.bbamcr.2011.09.010
Tang W, Xie Q, Guan J, Jin S, Zhao Y (2014a) Phytochemical profiles and biological activity evaluation of Zanthoxylum bungeanum Maxim seed against asthma in murine models. J Ethnopharmacol 152:444–450. https://doi.org/10.1016/j.jep.2014.01.013
Tang W, Zhang H, Wang L, Qian H (2014b) New cationic antimicrobial peptide screened from boiled-dried anchovies by immobilized bacterial membrane liposome chromatography. J Agric Food Chem 62:1564–1571. https://doi.org/10.1021/jf4052286
Tang W, Yuan H, Zhang H, Wang L, Qian H, Qi X (2015) An antimicrobial peptide screened from casein hydrolyzate by Saccharomyces cerevisiae cell membrane af fi nity method. Food Control 50:413–422. https://doi.org/10.1016/j.foodcont.2014.09.030
Tao R, Tong Z, Lin Y, Xue Y, Wang W, Kuang R, Wang P, Tian Y, Ni L (2011) Antimicrobial and antibiofilm activity of pleurocidin against cariogenic microorganisms. Peptides 32:1748–1754. https://doi.org/10.1016/j.peptides.2011.06.008
Wagener J, Schneider JJ, Baxmann S, Kalbacher H, Borelli C, Nuding S, Küchler R, Wehkamp J, Kaeser MD, Mailänder-Sanchez D, Braunsdorf C, Hube B, Schild L, Forssmann WG, Korting HC, Liepke C, Schaller M (2013) A peptide derived from the highly conserved protein gapdh is involved in tissue protection by different antifungal strategies and epithelial immunomodulation. J Invest Dermatol 133:144–153. https://doi.org/10.1038/jid.2012.254
Wang G, Li X, Wang Z (2016) APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res 44:D1087–D1093. https://doi.org/10.1093/nar/gkv1278
Wu H, Jiang T, Dong X, Shen G, Li S, Zhang Z (2016) Purification and characterization of angiotensin-I converting enzyme inhibitory peptides from prickly ash (Zanthoxylum bungeanum Maxim) seed protein hydrolysates. Int J Food Eng 12:333–342. https://doi.org/10.1515/ijfe-2015-0091
Xia L, You J, Li G (2011) Compositional and antioxidant activity analysis of Zanthoxylum bungeanum seed oil obtained by supercritical CO2 fluid extraction. J Am Oil Chem Soc 88:23–32. https://doi.org/10.1007/s11746-010-1644-4
Yang Q, Cao W, Zhou X, Cao W, Xie Y, Wang S (2014) Anti-thrombotic effects of α-linolenic acid isolated from Zanthoxylum bungeanum Maxim seeds. BMC Complement Altern Med 14:1–8. https://doi.org/10.1186/1472-6882-14-348
Zhang M, Li M, Sun L (2014) NKLP27 : A teleost NK-lysin peptide that modulates immune response , induces degradation of bacterial DNA , and inhibits bacterial and viral infection. PLoS One 9:e106543. https://doi.org/10.1371/journal.pone.0106543
Zhang M, Wang J, Zhu L, Li T, Jiang W, Zhou J (2017) Zanthoxylum bungeanum Maxim . ( Rutaceae ): a systematic review of its traditional uses , botany , phytochemistry , pharmacology , pharmacokinetics , and toxicology. Int J Mol Sci 18:2172. https://doi.org/10.3390/ijms18102172
Funding
This study received financial assistance from the Scientific Fund of the Application Fundamental Project (2016-JY-0118) and Key Project of Research and Development Program (18ZDYF1175), Science and Technology Department of Sichuan Province.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
This article does not contain any studies with human participants or animals by any of the authors.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Hou, X., Feng, C., Li, S. et al. Mechanism of antimicrobial peptide NP-6 from Sichuan pepper seeds against E. coli and effects of different environmental factors on its activity. Appl Microbiol Biotechnol 103, 6593–6604 (2019). https://doi.org/10.1007/s00253-019-09981-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-019-09981-y