
Abstract
Position sense at the human elbow joint has traditionally been measured in blindfolded subjects using a forearm matching task. Here we compare position errors in a matching task with errors generated when the subject uses a pointer to indicate the position of a hidden arm. Evidence from muscle vibration during forearm matching supports a role for muscle spindles in position sense. We have recently shown using vibration, as well as muscle conditioning, which takes advantage of muscle’s thixotropic property, that position errors generated in a forearm pointing task were not consistent with a role by muscle spindles. In the present study we have used a form of muscle conditioning, where elbow muscles are co-contracted at the test angle, to further explore differences in position sense measured by matching and pointing. For fourteen subjects, in a matching task where the reference arm had elbow flexor and extensor muscles contracted at the test angle and the indicator arm had its flexors conditioned at 90°, matching errors lay in the direction of flexion by 6.2°. After the same conditioning of the reference arm and extension conditioning of the indicator at 0°, matching errors lay in the direction of extension (5.7°). These errors were consistent with predictions based on a role by muscle spindles in determining forearm matching outcomes. In the pointing task subjects moved a pointer to align it with the perceived position of the hidden arm. After conditioning of the reference arm as before, pointing errors all lay in a more extended direction than the actual position of the arm by 2.9°–7.3°, a distribution not consistent with a role by muscle spindles. We propose that in pointing muscle spindles do not play the major role in signalling limb position that they do in matching, but that other sources of sensory input should be given consideration, including afferents from skin and joint.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The proprioceptive senses, the senses signalling the body’s own actions, are sometimes referred to collectively as the sixth sense. This is a mysterious sense because it acts largely unconsciously. It includes the sense of limb position and movement, the sense of force or of effort and the sense of balance. The subject of the present study is the sense of limb position. Where is my arm? I know where it is, but there is no identifiable sensation that I can attribute to its adopted position.
We have been studying the sensory neural basis of position sense for a number of years (Proske and Gandevia 2012). The work has been based on the landmark observations by Goodwin et al. (1972), who showed that vibration of elbow flexor muscles of one arm led to sensations of movement and displaced position of the arm in the direction of elbow extension as indicated in tracking movements by the other arm. As a result, the final position the arm appeared to adopt was more extended than its actual position. Similarly, vibration of elbow extensors led to sensations of movement into flexion. At the time it was known from animal experiments that in a passive muscle the muscle spindles were exquisitely sensitive to vibration (Brown et al. 1967). By implication it was assumed that muscle spindles were responsible for the movement illusions generated by vibration. This assumption was subsequently proved correct when direct recordings were made from human spindles during muscle vibration (Roll et al. 1989). It is now generally accepted that muscle spindles are responsible for the vibration illusion, the primary endings contributing to both the senses of limb movement and position, while the secondary endings signal limb position (Matthews 1988).
Our own past experiments sought additional supporting evidence for a role for muscle spindles in position sense. We brought a new technique to bear on the problem. Both the ordinary muscle fibres of a muscle and the intrafusal muscle fibres of muscle spindles exhibit a contraction history-dependent property called thixotropy which influences the passive tension and stiffness in resting muscle fibres. It has been shown in animal experiments that spindle discharge rates can be raised or lowered as a consequence of thixotropic conditioning and in parallel experiments on human subjects, similar conditioning produced reproducible errors in limb position sense (Gregory et al. 1988). For reviews, see Proske et al. (1993, 2014).
The original observations of Goodwin et al. (1972) were all made using a forearm matching task. One elbow muscle was vibrated, and the blindfolded subject was required to track the perceived movement of the forearm with their other arm. Similar matching experiments, with or without vibration, have now been carried out on the fingers (Ferrell and Smith 1988), hands (Walsh et al. 2013), knee (Givoni et al. 2007) and ankle (Boisgontier and Nougier 2013), and the results are generally supportive of the findings of Goodwin et al.
When a blindfolded subject carries out a forearm position matching task, the experimenter moves one arm, the reference arm, to the test angle and the subject moves their other arm, the indicator arm, into a matching position. That is, the subject moves their indicator arm until the sensations generated by the two arms feel the same. It is a sensation matching task.
When we ask ourselves “where is my arm?” we typically do not locate it by moving our other arm into a matching position. Yet we know where our arm is and we are able to point to its position. That conclusion has recently led us to revisit the question of position sense at the elbow, but this time we have used two different methods of measurement of position sense: a conventional forearm matching task and a position pointing task (Tsay et al. 2016). In the pointing task the reference arm is hidden behind a screen and the subject is required to move a pointer until this aligns with the perceived position of the hidden arm.
We posed the question: do muscle spindles contribute to position sense in the same way in matching and pointing tasks? In a matching task spindle signals from both arms contribute to achieve accurate alignment of the forearms (White and Proske 2009; Izumizaki et al. 2010; Tsuge et al. 2012). In a pointing task the relevant afferent information is coming from only one arm, so here the role of spindles was likely to be different. We recently explored this possibility using both thixotropic conditioning and muscle vibration (Tsay et al. 2016). It was concluded that for position sense measured by pointing, muscle spindles did not play the prominent role that they did in matching. Such a conclusion departs from the conventional view of the sensory origins of proprioception and deserves to be tested further by experiment.
The idea that the origin of afferent signals might be different in matching and pointing tasks is not new. Velay et al. (1989) concluded that matching and pointing did not test the same position sense. They proposed that basic position sense was coded by proprioceptors in terms of angular position in intrapersonal space, but that another form of coding was required to provide continuous knowledge of limb location in extrapersonal space. They speculated about a central transformation process which converted one sense to the other.
We have recently introduced a new method of thixotropic muscle conditioning, which we have called test angle co-conditioning (Tsay et al. 2014, 2015). Here the reference arm is brought to the test angle, elbow flexors and elbow extensors are both contracted isometrically, and the subject is then asked to match position of the arm with the indicator arm which has been either flexion or extension conditioned. In trials with such a conditioning procedure large matching errors were observed (Tsay et al. 2014).
In the present study we have resorted to the same method of test angle conditioning in an attempt to further explore the recently observed differences in the sensory afferent basis of position sense measured in pointing and matching tasks. Given that test angle conditioning produces large errors in a matching task, it was hoped that the same method applied to a pointing task would reveal new differences in position sense measured in this way.
Methods
A total of 23 subjects were recruited for this study. They included 9 males and 14 females with an average age of 24.5 (±0.6) years. Subjects gave informed, written consent prior to participating in the study, which was approved by the Monash University Human Research Ethics Committee. The ethical aspects of the experiments conformed to the Declaration of Helsinki.
The apparatus
Position sense was measured in two ways. For the matching task the blindfolded subject sat at a table and placed both forearms on lightweight paddles attached to a custom-built piece of apparatus for measuring forearm position sense (Allen and Proske 2006). The forearms, palms facing upward, were strapped to the paddles by Velcro straps placed just below the crease of the wrist and 5 cm in width. In order to minimise potential differences in skin sensation between the two arms, equal tension from the strapping was checked before proceeding with the experiment. One arm was designated the reference arm (the arm placed at the target angle by the experimenter), while the other arm was the indicator arm (the arm moved by the subject to match the perceived position of the reference arm).
In the pointing task the two arms were separated by a screen. The screen consisted of a large, removable board that obscured subjects’ vision of their reference arm, including the shoulder. The reference arm was strapped in, as before, its position set by the experimenter. In a pointing trial the subject was required to push or pull a lever at the base of the pointer paddle to move the pointer upwards or downwards until it was perceived to be aligned with the hidden reference arm (Tsay et al. 2016, Fig. 1).
Forearm and pointer paddle angles were measured using potentiometers located at the hinges of each paddle. When an arm was strapped to a paddle, the paddle hinges were co-linear with the elbow joint (Allen and Proske 2006, Fig. 1). The potentiometers provided a continuous voltage output proportional to the angle of each paddle, where 0° indicated that the forearm was horizontal and 90° indicated that it was vertical. Calibration of the potentiometers was checked before commencement of an experimental series with each subject.
Muscle activity of the reference arm was measured using surface electromyogram (EMG). Here we did not analyse the EMG signal as such, but used it to provide feedback, indicating whether or not the subject had remained relaxed as the experimenter moved the reference arm to the test angle. A pair of Ag–AgCl electrodes with an adhesive base and solid gel contact points (3 M Health Care, London, Ontario, Canada) were placed approximately 2.5 cm apart over the surface of the biceps brachii and triceps brachii. A reference electrode was placed on the collar bone. EMG output was connected to an audio amplifier for biofeedback. Position, force and EMG signals were acquired using a MacLab 4/s data acquisition module running Chart software (AD Instruments, Castle Hill, NSW, Australia) on a Macintosh computer.
Measuring position errors
Position errors between the two paddles were calculated using the formula:
The convention was used that 0° represented a fully extended arm with the forearm horizontal, 90° a flexed arm with the forearm in the vertical position. A positive error was an error in the direction of extension relative to the position of the reference arm, and a negative error, an error in the direction of flexion.
Reference and indicator arms were randomly assigned for each set of trials to reduce any biases from matching with a dominant or non-dominant arm (Goble et al. 2006). During a matching trial the reference arm was placed by the experimenter at an angle of 45° and the blindfolded subject was asked to match its perceived position with their indicator arm. In a pointing trial the reference arm was positioned in the same way, but the subject indicated its perceived position by moving the pointer paddle to align it with the hidden arm.
During movement of the reference arm to the test angle by the experimenter subjects were asked to remain relaxed. This was monitored with auditory feedback of EMG. Throughout these experiments, once the reference arm had been placed at the test angle, the subject maintained its position voluntarily. All of the matching by the indicator arm was also done voluntarily by the blindfolded subject. In both matching and pointing trials subjects were therefore required to generate mild contractions sufficient to support the reference arm against gravity and in the matching task for movement of the indicator into the matching position. These conditions were chosen to keep both matching and pointing processes close to what subjects might do in everyday life.
Muscle conditioning
Muscle thixotropy is a property of resting skeletal muscle, a contraction history-dependent change in passive tension and stiffness dependent on the presence of long-term stable cross-bridges between actin and myosin in sarcomeres. The thixotropic state of a muscle can significantly alter the responses of muscle spindles and therefore exerts a strong influence on the measurement of position sense (Proske et al. 2014). In order to control for thixotropy-related position errors it is necessary at the start of each matching trial to put elbow muscles of both arms into a defined state. This is called muscle conditioning. In the pointing trial it was only necessary to condition the reference arm.
We have previously applied two forms of conditioning to both arms, called flexion conditioning and extension conditioning (Allen et al. 2007). In the present study such conditioning was applied only to the indicator arm in the matching trials. For flexion conditioning (FC) the forearm was moved into a vertical position (90°) and the blindfolded subject was asked to generate a 2 s, approximately half-maximum contraction, flexing the arm towards the body. Once arm muscles had relaxed, the subject moved the arm in the direction of extension into a matching position with the reference arm which had been isometrically conditioned at the test angle. For extension conditioning (EC) the indicator arm was moved into full extension (0°) and the subject was asked to push down onto the supporting table to generate a half-maximum contraction in elbow extensors. Again, once arm muscles had relaxed, the subject moved the indicator arm in the direction of flexion to adopt a matching position with the isometrically conditioned reference arm.
Therefore, in a matching trial muscles of the reference arm had been co-conditioned, while muscles of the indicator arm had been either flexion or extension conditioned. For flexion conditioning it left indicator flexor muscles taut after the conditioning contraction, and when the indicator arm was extended from 90° to the test angle (45°), they were stretched by the movement and therefore kept in a taut state. The same movement slackened the extensors whose passive tension therefore fell to low levels. This meant that after flexion conditioning flexor spindles were sensitised, while extensor spindles were de-sensitised. As a consequence, the proprioceptive signal coming from the indicator arm was biased in favour of elbow flexors. Similarly, after extension conditioning the signal was biased in favour of elbow extensors.
Experiment 1: Co-conditioning at the test angle
We wanted to measure position sense under conditions where such a bias in the origin of the proprioceptive signal from flexion or extension conditioning was not present. An additional form of conditioning was therefore used with the reference arm that left its elbow flexors and extensors in the same sensitised state (Tsay et al. 2014). To do that, isometric contractions of both antagonists were carried out at the test angle (co-conditioning).
Two conditioning sequences were used: in one the arm was placed at the test angle (45°) by the experimenter and the subject was asked to generate a half-maximum contraction of elbow extensors, followed by a half-maximum contraction of elbow flexors (Ref: EC + FC, Fig. 1). Alternatively, the reverse sequence was used, beginning with a flexor contraction followed by an extensor contraction (Ref: FC + EC, Fig. 1). During the contractions the paddle supporting the arm was held fixed in position at the test angle, so there was no accompanying movement.

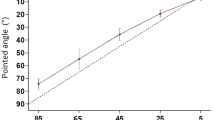
Position errors measured in matching and pointing tasks after co-conditioning of the reference arm. Mean matching errors (left-hand panel) and pointing errors (right-hand panel) after co-conditioning the reference arm at 45° with the sequence EC + FC (filled circles) and FC + EC (open circles). In the matching trials the indicator arm was either flexion conditioned at 90° (filled circle) or extension conditioned at 0° (open circle). In the pointing task the position of the hidden arm was indicated with a pointer. In this and subsequent figures the convention is used that errors by the indicator arm, or the pointer, in the direction of extension are given a positive value. Errors in the direction of flexion are given a negative value. Dotted line zero error. Asterisk indicates significant difference
In the matching trials, for the reference conditioning sequence EC + FC, at the same time as performing the flexor contraction of the reference arm, the subject also carried out flexion conditioning of their indicator arm while it was held at 90°. The conditioning was done in this way, finishing up with flexion conditioning of both arms, to minimise any differences between the arms in adaptation effects of afferent responses. For the reference sequence FC + EC the same procedure was carried out, but the indicator arm was extension conditioned by holding it at 0° and carrying out an extensor contraction. Once it had relaxed, the indicator arm was moved by the subject into a matching position. So the elbow of the reference arm had undergone both extension and flexion contractions at the test angle, while the indicator had been either flexion conditioned (filled circle, Matching, Fig. 1) or extension conditioned (open circle, Matching, Fig. 1).
In the pointing task, only the reference arm was conditioned, using the same two sequences of test angle extensor and flexor contractions: EC + FC (filled circle, pointing, Fig. 1) and FC + EC (open circle, pointing, Fig. 1). When the reference conditioning had finished with a flexor contraction, the subject moved the pointer paddle downwards from a starting angle of 90°, into a position where they considered it to be aligned with the hidden reference arm. When reference conditioning had finished with an extensor contraction, they moved the pointer upwards from an initial position of 0°. The pointing was done in this way to make the direction of movement of the pointer similar to that for the indicator arm in the matching task.
Experiment 2: Experimenter-move versus self-move
A question that arose during the pointing experiments was whether signals coming from the arm moving the pointer to indicate location of the reference arm contributed in some way to the position errors. It was therefore decided to do a series of control experiments where in addition to trials in which the subject moved the pointer themselves, a further set was carried out where the pointer was moved into alignment by the experimenter, under instructions from the subject. During these trials the subject was asked to place the pointing arm in their lap, unmoving.
Experiment 3: Matching with the indicator under visual control
In the pointing trials the subject could see the pointer as they moved it into the perceived position of the hidden reference arm, so the pointing was done under visual control, even though it was non-informative vision. In contrast, the matching trials had all been done with the subject blindfolded. It was therefore felt necessary to do an additional series of control matching trials where the subject could see their indicator arm (experiment 3). Here the reference arm was always co-conditioned, and in the matching trials the indicator arm was flexion or extension conditioned, as before. However, in addition to the blindfolded matching trials, a second set of matching trials was done where the subject moved their indicator arm into a matching position with the arm in full view. The reference arm remained hidden.
Statistical analysis
In experiment 1 subjects carried out a series of 5 trials with each conditioning sequence. For matching there were 10 trials: 5 where the reference arm was isometrically conditioned and the indicator flexion conditioned and 5 where the reference was isometrically conditioned and the indicator extension conditioned. For pointing there were also 10 trials: 5 trials where the reference arm was isometrically conditioned, beginning with an extension contraction (EC + FC) and 5 trials where the isometric contraction began with a flexor contraction (FC + EC). This made for a total of 20 trials. For 8 subjects an additional 10 trials were carried out where the experimenter rather than the subject moved the pointer. The results from these trials were compared with 10 trials where the subject moved the pointer themselves (experiment 2). A new cohort of 9 subjects did two series of matching trials: one under visual control, the other blindfolded, as well as the pointing trials, making for a total of 30 trials (experiment 3).
In each experimental trial conditions were randomised.
The effect of conditioning on position errors for each task, matching and pointing, was analysed using a two-tailed paired t-test. Further, separate, repeated-measures ANOVA were used to test for the effects on arm position errors of self-moved versus experimenter-moved placement of the indicator, as well as for the effects of vision on pointing and matching trials. If significance was found, a Bonferroni post hoc test was used to determine which pointing trials were significantly different.
Pooled data from each experiment were shown as mean ± standard error of the mean (SEM).
Results
In previous experiments we had shown that distributions for position errors measured in matching trials were significantly different from the distributions in pointing trials under conditions where the arms had been either flexion or extension conditioned (Tsay et al. 2016). In the present experiments the aim was to further explore this difference, using co-conditioning at the test angle (Tsay et al. 2014). This additional method of conditioning was selected because it was predicted to produce rather large differences in outcomes for matching and pointing trials.
Experiment 1: Co-conditioning at the test angle
Matching
For the co-conditioning sequence EC + FC of the reference arm and with flexion conditioning of the indicator arm, in the matching trials (filled circle, left-hand panel, Fig. 1), the pooled data for the 14 subjects gave a mean error of −6.2° (±1.3°). That is, the blindfolded subject tended to indicate the position of their reference arm as more flexed than its actual position. Here the error was somewhat smaller than expected, based on similar recent experiments (Tsay et al. 2015).
The conditioning order was now reversed. Instead of the isometric conditioning starting with a contraction of extensors, it began with a contraction of flexors to give the sequence for the reference arm, FC + EC. Here the indicator arm was always extension conditioned. The reason for choosing such a reversal of the conditioning sequence was that in previous matching experiments it had led to a reversal of the direction of the errors. The pooled data for the 14 subjects (open circle, left-hand panel, Fig. 1) gave a mean error of +5.8° (±1.7°); that is, subjects perceived their reference arm to be more extended than its true position. The errors into extension were as had been predicted, in a direction opposite to that following flexion conditioning of the indicator, although their size was smaller than had been anticipated from previous experiments (Tsay et al. 2015).
Pointing
The experiment was repeated, but this time only the reference arm was conditioned. Position of the hidden arm was indicated by the subject moving the pointer paddle to a position where the subject felt it was in alignment with the reference arm. So this was measuring position sense by pointing. Data for pointing trials are shown in the right-hand panel in Fig. 1. For the conditioning sequence EC + FC of the reference arm (filled circle, right-hand panel, Fig. 1) the mean error for the 14 subjects was +7.2° (±1.1°). That is, subjects felt that the pointer paddle was accurately aligned with the hidden reference arm when in fact its position was more extended than the reference arm by an average of 7.2°. Therefore, using the same conditioning of the reference arm as in matching (EC + FC), the pointing trials produced errors differing from the matching errors by 13.4° (6.2° + 7.2°).
In interpreting this result it should be kept in mind that matching trials involve proprioceptive signals coming from both arms while in pointing trials presumably only a signal from the reference arm was involved. We have previously provided evidence that co-conditioning of elbow antagonists at the test angle led to a difference signal coming from the conditioned arm that lay close to zero relative to the signal coming from the indicator arm (Tsay et al. 2015). We therefore predicted that in the present experiments conditioning the reference arm in this way would similarly generate a difference signal close to zero. So in the pointing trials the observed errors were in the opposite direction from the errors in the matching trials and, in addition, they did not lie close to zero, as had been predicted.
The experiment was then repeated but using pointing trials, with the reference arm conditioned with the reverse sequence, FC + EC (open circle, right-hand panel, Fig. 1). The pooled data for the 14 subjects yielded a mean error of +2.9° (±1.0°). This compared with an expected error of 0° (Tsay et al. 2015). So again the outcome of the pointing trials differed from expectations.
When the distributions of errors using the two forms of conditioning shown in Fig. 1 were compared, statistical analysis, using paired t-tests, showed that they were significantly different (p < 0.05) for both pointing (t(13) = 2.709, p < 0.01) and matching tasks (t(13) = −6.341, p < 0.05). A significant difference was expected for the errors in the matching tasks, given that two forms of conditioning were used, but they were unexpected for the pointing tasks since here the hypothesis was that the distribution of errors was independent of the form of conditioning (see “Discussion”).
Experiment 2: Experimenter-move versus self-move
The data presented so far suggested that muscle conditioning had different effects on forearm position errors measured in pointing and matching trials. This conclusion was in line with other recent observations (Tsay et al. 2016). It is known that in forearm matching trials signals from both arms make a contribution (White and Proske 2009; Hakuta et al. 2014; Tsay et al. 2014). In the search for an explanation of the pointing results, the possibility was considered that proprioceptive activity associated with the arm moving the indicator paddle to align it with the perceived position of the hidden reference arm influenced, in some way, the measured values.
To put this idea to the test, for 8 of the 14 subjects pointing errors were measured both with the subject aligning the pointer themselves and with the alignment carried out by the experimenter. Here the experimenter adjusted the position of the pointer until the subject declared accurate alignment with the unseen arm. Throughout the trial the subject’s other arm rested on their lap, unmoving.
Results for the 8 subjects are shown in Fig. 2 for the same forms of conditioning of the reference arm as had been used in the previous trials (EC + FC and FC + EC). Notice that the general trend of the data with errors lying in the direction of extension was the same as for pointing errors observed in the earlier trials (Fig. 1). When the pointer paddle was moved by the experimenter, position errors lay a little further in the direction of extension than when the subject moved the pointer themselves. For co-contraction conditioning of the reference arm, EC + FC, self-moved pointing produced an error of +6.7° (±1.6°), while for experimenter-moved pointing the error was +9.0° (±1.3°). Reversing the isometric conditioning to FC + EC yielded a self-moved pointing error of +4.7° (±0.8°), while experimenter-moved error gave +5.2° (±2.3°).

Errors in a pointing task when the pointer is moved by the subject or the experimenter. Mean pointing errors (±SEM) for a group of 8 subjects with the reference arm co-conditioned at the test angle with two forms of conditioning (FC + EC and EC + FC). Filled circles, values when the subject moved the pointer themselves; open circles, when the pointer was moved by the experimenter. Error displays as in Fig. 1. Asterisk indicates significant difference
A repeated-measures ANOVA between subjects showed that these differences were significant F(7, 49) = 2.384, p = 0.04. Bonferroni post hoc tests showed significant differences (p < 0.05) in values for experimenter-moved versus self-moved pointing for EC + FC.
Experiment 3: Matching with the indicator under visual control
In this experiment two sets of matching trials were carried out: in one the subject remained blindfolded, as before, and in the other they could see their indicator arm. The reference arm always remained hidden from view. The matching with vision was done as a control since in the pointing trials the subject could always see the pointer. The experiment was carried out with a new cohort of 9 subjects. The results are shown in Fig. 3.

Position errors in matching and pointing tasks where the matching was done with or without visual control. Mean matching and pointing errors (±SEM) for a group of 9 subjects where pointing (crosses) was done as before, but matching was done blindfolded (filled circles) or under visual control (open circles). Error displays as in Fig. 1. Asterisks indicate significant differences
For blindfolded matching with the reference arm conditioned EC + FC and the indicator FC, the mean error was −13.4° (±3.2°). In the trials where the subject could see their indicator arm the mean error was −8.6° (±2.5°). So matching under visual control reduced the errors by 4.8°. In the pointing trials where the reference arm was conditioned EC + FC, position errors were +5.4° (±2.1°). The pointing result was therefore similar to previous values (Fig. 1, 2).
The experiment was now repeated using the reverse conditioning sequence FC + EC for the reference arm and EC for the indicator arm. Blindfolded matching gave a mean error of +7.9° (±2.0°). When matching was repeated with the indicator arm visible, the error was +6.2° (±2.4°). So here the difference in errors with and without the indicator arm visible was rather smaller, 1.7°. For pointing, using the conditioning sequence FC + EC, the mean error was 5.8° (±3.0°). The result was approximately in line with the earlier finding (Fig. 1, 2).
Statistical analysis using a repeated-measures ANOVA showed a significant difference between values: F(5, 40) = 15.5, p = 0.0. Post hoc analysis (Bonferroni test), with the reference arm conditioned EC + FC, revealed a significant difference between pointing and blindfolded matching (p = 0.01) and between pointing and matching under visual control (p = 0.04). There was no significant difference between the matching values, with and without vision. For conditioning of the reference arm, FC + EC, there were no significant differences between the errors in the matching and pointing tasks, with and without vision.
Discussion
These experiments are submitted in support of the view that there is a task dependency in the afferent origins of proprioceptive signals responsible for generating human position sense. In a forearm position matching task muscle spindles play a prominent role, while in a pointing task spindles are relegated to play only a minor role, if any, in providing the position signal.
Experiment 1: Co-conditioning at the test angle
For the matching trials in experiment 1 the observed distribution of position errors compared favourably with findings from a similar experiment carried out previously (Tsay et al. 2015). In that study we had observed that when the reference arm had elbow muscles conditioned at the test angle and the indicator arm was flexion conditioned, errors lay 9.2° in the direction of flexion. This was rather more than the errors seen in the present study (6.2°), although their direction was the same. When the indicator had been extension conditioned, errors were 7.5° in the direction of extension which again was consistent with observations in the present study (5.8°). We then showed that if slack was introduced in indicator flexor and extensor muscles to effectively lower their levels of spindle afferent activity (Tsay et al. 2015; Fig. 1c, d), matching errors lay close to zero. The result implied that the net spindle discharge rate and therefore the position signal coming from the co-conditioned reference arm was low.
For the pointing trials the most important consideration is that here the afferent information is coming largely from one arm, while in matching trials afferent input from both arms contributes (White and Proske 2009; Izumizaki et al. 2010; Tsuge et al. 2012). We predicted errors for the pointing trials to be lying close to zero. We have explained above that the output from a co-conditioned reference arm is predicted to be close to zero. In the event errors in the pointing trials all lay in the direction of extension.
While the general trend observed in the pointing trials was errors in the direction of extension, there were significant differences in size between the pooled values for conditioning FC + EC and its reverse, EC + FC (Fig. 1). There is no obvious explanation for this result given that two near-identical forms of conditioning had been used. Further, pointing errors for the same two conditioning sequences in the experimenter-move versus self-move experiment (Fig. 2) yielded values that were not significantly different from one another for this cohort of subjects (paired sample t-test, p < 0.05). Hence, we retain the view that in a pointing task all errors are approximately the same no matter what form of conditioning is used.
Experiment 2: Experimenter-move versus self-move
Here we assessed the possibility that one source of the error distribution in the pointing task was the arm moving the pointer. Indeed, it was found that there was a significant difference when the pointer was moved by the experimenter, compared with when it was moved by the subject (Fig. 2). It implied some, if small, influence on the measured errors coming from the pointing arm. However, more importantly, the general trend seen in pointing trials that the distribution of errors was insensitive to the form of muscle conditioning of the reference arm and always lay in the direction of extension remained unchanged (see also Tsay et al. 2015).
Experiment 3: Matching with the indicator under visual control
The differences in errors in the forearm matching task obtained with the subject blindfolded and with the indicator arm in view were relatively small (Fig. 3). They were only significant for the conditioning sequence of the reference arm EC + FC, not for the reverse, FC + EC. More importantly, whether the indicator arm was visible or not did not alter the overall pattern of matching errors, with values lying in the direction of flexion or extension, depending on the form of conditioning of the reference arm. It suggests that vision of the indicator arm exerts only a limited influence on the error distribution. The finding is consistent with our earlier observation that vision of the indicator arm altered only slightly, non-significantly, the angular difference in an arm matching task, even when measured during vibration of the reference arm (Izumizaki et al. 2010). Similarly, Longo (2014) found that hand shape distortions seen in implicit maps drawn by pointing to the unseen hand persisted, but were significantly altered by non-informative vision of the pointing arm. We conclude that in a pointing task vision can influence the position error distribution, but does not alter the general trend of errors lying in the direction of arm extension.
Mechanism
Our interpretations are based on the assumption that spindle resting discharge levels determine limb position (Clark et al. 1985) and that changes in limb position produced by muscle conditioning are a result of alterations in spindle resting discharge (Gregory et al. 1988). We have previously proposed that it is the difference in afferent signal coming from the two antagonists at the elbow that determines the perceived position of the forearm (Proske et al. 2014). When these muscles are conditioned with a voluntary contraction at the test angle, thixotropic considerations dictate that any pre-existing slack is removed in extrafusal and intrafusal muscle fibres of both muscle groups. As a consequence, resting discharge rates in spindles of both muscle groups rise to similarly high levels. When this is the case, the difference signal is low. This leads to generation of a low position signal.
There is evidence from other quarters that supports such an interpretation. Gilhodes et al. (1986) showed that raising spindle discharge rates by similar amounts in both antagonists at the elbow, using vibration, led to abolition of the vibration illusion seen when only one muscle was vibrated. More recently, the technique of co-vibration of antagonists has been used as a method for degrading or masking proprioceptive feedback from an arm (Bock et al. 2007; Brun and Guerraz 2015). Our interpretation of these results is that the similarly high levels of spindle signal in both antagonists during co-vibration lead to a low difference signal which is interpreted centrally as reduced proprioceptive input from the arm. We are therefore proposing that co-conditioning of the antagonists with voluntary contractions at a given test angle leads to a similar outcome as co-vibration. However, there is one distinction. While the vibration can potentially engage a range of receptor types, including muscle, tendon, joint and skin receptors, only striated muscle, including the intrafusal fibres of spindles, exhibits thixotropic properties (Proske et al. 2014). So the effects of a conditioning contraction can be attributed specifically to afferents of muscle spindles.
It is currently believed that both primary and secondary endings of muscle spindles contribute to limb position sense (McCloskey 1973). The question arises: do all afferents of muscle spindles exhibit thixotropic behaviour? In an animal study, the responses of single, identified afferents of muscle spindles, including primary and secondary endings, were tested for thixotropic properties (Proske et al. 1992). It was found that all primary endings exhibited such behaviour, but 16 of 35 secondary endings did not. These afferents had characteristically low conduction velocities, and it was speculated that the lack of a history dependence related to the location of the sensory terminals on the intrafusal fibres. The result means that thixotropic conditioning of human muscle may not engage all spindle afferents in the muscle. It raises the possibility, although unlikely, that in a pointing task there is a contribution to the position signal from spindle secondary endings which are unresponsive to the conditioning.
Accepting that a reference arm with antagonists co-conditioned at the test angle generated a net low proprioceptive signal, why were the errors so large after flexion or extension conditioning of the indicator? We propose that flexion conditioning of the indicator arm at 90°, followed by movement of the arm to the matching position generated in the arm a signal strongly biased in favour of flexor spindles, so the subject was trying to match a weak reference signal with a strong indicator flexor signal. It led the subject to match with the indicator elbow flexed, trying to keep elbow flexor muscles short and therefore their spindle discharge rates low. So position errors lay in the direction of flexion (−6.2°, filled circle, left-hand panel, Fig. 1). The same kind of explanation can be applied to extension conditioning of the indicator where the proprioceptive signal was biased in favour of extensors, so matching errors were predicted to lie in the direction of extension (+5.8°, open circle, left-hand panel, Fig. 1).
The trend in the pointing trials that all of the errors lay in the direction of extension was similar to that seen previously (Tsay et al. 2016). The result suggested that the central processes involved in generating the sensation of limb position in a pointing task were relatively insensitive to changes in spindle afferent input produced by muscle conditioning. In that report we provided additional evidence for our view by showing that the illusions of limb position generated by muscle vibration in matching tasks (Goodwin et al. 1972) were not present in pointing tasks. Since muscle vibration is a powerful stimulus for muscle spindles (Roll et al. 1989), it was concluded that spindle signals played a lesser role in position sense measured in a pointing task compared with a matching task. As a consequence, in a pointing task position errors generated by muscle conditioning no longer conformed to predictions.
The finding of no vibration response in a pointing task (Tsay et al. 2016) remains controversial. Izumizaki et al. (2010) observed a 30 % reduction of the vibration illusion in a pointing task compared with a matching task, while Kammers et al. (2006) found the illusion three times larger in a matching task compared with a reaching task. So it may be that the vibration illusion is not always fully suppressed in a pointing task.
Recently, Kigawa et al. (2015) showed that the size of the vibration illusion generated in one arm could be reduced by vibrating the other arm. Such an outcome is presumably due to a signal difference mechanism operating during forearm matching (Proske et al. 2014). Kigawa et al. showed further that if the second arm was vibrated in an abducted position, the crossed effect on the other arm was no longer present. Yet it was possible to demonstrate a healthy vibration response in the abducted arm itself. Some years ago it was shown that when position sense was measured in a matching task, with both arms abducted, the standard deviation of matching errors was double that seen in a matching task with the arms in front (Gooey et al. 2000). The result suggested that subjects were less sure of the positions of their abducted arms. All of this emphasises the importance of arm posture for the position matching mechanism. When we carry out skilled manipulations with our hands, the hands are in front, forearms aligned. It is under these conditions that high alignment accuracy is required.
What might be the reason for the distribution of errors in the direction of extension in the pointing tasks? We have previously proposed that in a matching task it is the difference in afferent signals coming from the two arms that is used to indicate their degree of alignment (Proske et al. 2014). Obviously, in a task such as arm pointing the sense of limb position cannot be generated using such a difference mechanism since afferent signals from only one arm are available. We have postulated that in a pointing task the afferent signals from the reference arm access a central map of the body, the postural schema (Longo et al. 2010) to generate the positional information. One possible factor contributing to the distribution of position errors in a pointing task is a distortion of central somatosensory maps, reflecting regional differences in sensory innervation (Miller et al. 2016). It is conceivable that map distortions of forearm length and its angular location are responsible for errors lying in the direction of extension.
Wider implications
The original experiments establishing a role for muscle spindles in proprioception were by Goodwin et al. (1972). Their findings have been very influential; whenever the subject of proprioceptors is brought up and their role in human position sense is considered, it is tacitly assumed that muscle spindles comprise a major source of the afferent input (see, for example, Goble et al. 2009). The present report is submitted in support of the view that the role of muscle spindles in position sense depends on how position sense is measured. If it is measured in a pointing task, evidence from thixotropic conditioning of muscle, including the present study, and from the effects of muscle vibration (Tsay et al. 2016), does not support a prominent role for muscle spindles. On the other hand, where position sense is measured in a matching task, by placing one limb to align it with the perceived position of the other, the evidence for spindle involvement is strong.
If our conclusion is accepted and muscle spindles do not contribute significant proprioceptive signals in a position pointing task, what source of afferent signal might be responsible for providing the positional information? In the pointing trials the errors all lay 3°–7° in the direction of extension. So subjects were aware of the position of their reference arm, if less precisely than in a two-arm matching task. In an arm movement task concerned with interlimb coupling, Brun and Guerraz (2015) reported that when elbow antagonists of the moving arm were vibrated, a “condition of proprioceptive masking”, subjects reported that they could still feel the passive displacement of the arm. It was concluded that the effect of the vibration was not sufficient to fully suppress conscious movement perception. The authors speculated that other muscle, skin or joint receptors were responsible for the remnant sensation.
These considerations lead to the suggestion that we have two senses of limb position. When both arms are involved in determining limb position and they are relatively closely aligned (±10°, Proske et al. 2014), the spindle mechanism plays a prominent role and it ensures an alignment accuracy of ±2°. If this mechanism is unable to be engaged because the two arms are too far apart, or if the task involves afferent input predominantly from only one arm (pointing), an additional underlying source of position signal is accessed.
Given that subjects were blindfolded in the main experiment, senses such as vision and hearing, which are able to provide spatial information, were not involved. Apart from spindles, other sensory receptors which have been considered to contribute to position sense include skin and joint receptors (Proske and Gandevia 2012). Potential candidates for position sensors in the skin are the slowly adapting Type II, Ruffini endings that respond to skin stretch. We have recently attempted to bias limb position sense at the forearm by stretching skin over the elbow joint (Tsay et al. 2016). This did not produce any detectable effect on position sense measured in either a position matching or pointing task. Such a negative result does not mean, of course, that cutaneous input plays no role, but simply that position sense cannot be significantly modified with the method used. There is, however, evidence that movement sense can be altered by skin stretch (Collins and Prochazka 1996; Collins et al. 2005). Similarly for joint receptors, they are considered principally to be movement detectors, but the presence in many joints of Ruffini endings means they are potentially capable of providing positional information, especially towards the extremes of the range of joint movement (Ferrell and Smith 1988; Fuentes and Bastian 2010). In summary, the jury is still out over the afferent origins of position sense measured in a pointing task. Our evidence suggests spindles do not play a major role, but that other sources of position signals including skin and joint should be given further consideration (Tsay et al. 2016).
One other possibility is that the positional information in a pointing task is generated centrally, perhaps the result of an effort-related signal generated by the subject as they support the weight of their arm. We have tested this possibility, and the evidence does not support a role for a motor command signal in position sense at the elbow (Tsay et al. 2016).
To conclude, we present the observations in this report in support of the view that in tasks measuring position sense by pointing to the hidden limb, the afferents of muscle spindles do not play a prominent role. By contrast, spindles make a major contribution to position sense measured in a forearm matching task. The realisation that the sensory origins of position sense measured by matching and pointing are different offers new insight into the underlying mechanisms. If this view is confirmed, it represents an important advance in our understanding of the processing of spatial information.
References
Allen TJ, Proske U (2006) Effect of muscle fatigue on the sense of limb position and movement. Exp Brain Res 170:30–38
Allen TJ, Ansems GE, Proske U (2007) Effects of muscle conditioning on position sense at the human forearm during loading or fatigue of elbow flexors and the role of the sense of effort. J Physiol 580(2):423–434
Bock O, Pipereit K, Mierau A (2007) A method to reversibly degrade proprioceptive feedback in research on human motor control. J Neurosci Methods 160(2):246–250
Boisgontier MP, Nougier V (2013) Proprioception: bilateral inputs first. Neurosci Lett 534:96–100
Brown MC, Engberg I, Matthews PB (1967) Fusimotor stimulation and the dynamic sensitivity of the secondary ending of the muscle spindle. J Physiol 189(3):545–550
Brun C, Guerraz M (2015) Anchoring the “floating arm”: use of proprioceptive and mirror visual feedback from one arm to control involuntary displacement of the other arm. Neuroscience 310:268–278
Clark FJ, Burgess RC, Chapin JW, Lipscomb WT (1985) Role of intramuscular receptors in the awareness of limb position. J Neurophysiol 54(6):1529–1540
Collins DF, Prochazka A (1996) Movement illusions evoked by ensemble cutaneous input from the dorsum of the human hand. J Physiol 496(Pt 3):857–871
Collins DF, Refshauge KM, Todd G, Gandevia SC (2005) Cutaneous receptors contribute to kinesthesia at the index finger, elbow, and knee. J Neurophysiol 94(3):1699–1706
Ferrell WR, Smith A (1988) Position sense at the proximal interphalangeal joint of the human index finger. J Physiol 399:49–61
Fuentes CT, Bastian AJ (2010) Where is your arm? Variations in proprioception across space and tasks. J Neurophysiol 103(1):164–171
Gilhodes JC, Roll JP, Tardy-Gervet MF (1986) Perceptual and motor effects of agonist–antagonist muscle vibration in man. Exp Brain Res 61(2):395–402
Givoni NJ, Pham T, Allen TJ, Proske U (2007) The effect of quadriceps muscle fatigue on position matching at the knee. J Physiol 584(1):111–119
Goble DJ, Lewis CA, Brown SH (2006) Upper limb asymmetries in the utilization of proprioceptive feedback. Exp Brain Res 168(1–2):307–311
Goble DJ, Coxon JP, Wenderoth N, Van Impe A, Swinnen SP (2009) Proprioceptive sensibility in the elderly: degeneration, functional consequences and plastic-adaptive processes. Neurosci Biobehav Rev 33(3):271–278
Goodwin GM, McCloskey DI, Matthews PB (1972) The contribution of muscle afferents to kinaesthesia shown by vibration induced illusions of movement and by the effects of paralysing joint afferents. Brain 95(4):705–748
Gooey K, Bradfield O, Talbot J, Morgan DL, Proske U (2000) Effects of body orientation, load and vibration on sensing movement and position at the human elbow joint. Exp Brain Res 133:340–348
Gregory JE, Morgan DL, Proske U (1988) Aftereffects in the responses of cat muscle spindles and errors of limb position sense in man. J Neurophysiol 59(4):1220–1230
Hakuta N, Izumizaki M, Kigawa K, Murai N, Atsumi T, Homma I (2014) Proprioceptive illusions created by vibration of one arm are altered by vibrating the other arm. Exp Brain Res 232(7):2197–2206
Izumizaki M, Tsuge M, Akai L, Proske U, Homma I (2010) The illusion of changed position and movement from vibrating one arm is altered by vision or movement of the other arm. J Physiol 588(15):2789–2800
Kammers MPM, van der Ham IJM, Dijkerman HC (2006) Dissociating body representations in healthy individuals: differential effects of a kinaesthetic illusion on perception and action. Neuropsychologia 44:2430–2436
Kigawa K, Izumizaki M, Tsukada S, Hakuta N (2015) Proprioceptive interaction between the two arms in a single-arm pointing task. PLoS ONE 10(8):e0137031
Longo MR (2014) The effects of immediate vision on implicit hand maps. Exp Brain Res 232(4):1241–1247
Longo MR, Azanon E, Haggard P (2010) More than skin deep: body representation beyond primary somatosensory cortex. Neuropsychologia 48(3):655–668
Matthews PBC (1988) Proprioceptors and their contribution to somatosensory mapping: complex messages require complex processing. Can J Physiol Pharmcol 66:430–438
McCloskey DI (1973) Differences between the sense of movement and position shown by the effects of loading and vibration of muscles in man. Brain Res 61:119–131
Miller LE, Longo MR, Saygin AP (2016) Mental body representations retain homuncular shape distortions: evidence from Weber’s illusion. Conscious Cogn 40:17–25
Proske U, Gandevia SC (2012) The proprioceptive senses: their roles in signaling body shape, body position and movement, and muscle force. Physiol Rev 92(4):1651–1697
Proske U, Morgan DL, Gregory JE (1992) Muscle history dependence of responses to stretch of primary and secondary endings of cat soleus muscle spindles. J Physiol 445:81–95
Proske U, Morgan DL, Gregory JE (1993) Thixotropy in skeletal muscle and in muscle spindles: a review. Prog Neurobiol 41(6):705–721
Proske U, Tsay A, Allen T (2014) Muscle thixotropy as a tool in the study of proprioception. Exp Brain Res 232(11):3397–3412
Roll JP, Vedel JP, Ribot E (1989) Alteration of proprioceptive messages induced by tendon vibration in man: a microneurographic study. Exp Brain Res 76(1):213–222
Tsay A, Savage G, Allen TJ, Proske U (2014) Limb position sense, proprioceptive drift and muscle thixotropy at the human elbow joint. J Physiol 592(Pt 12):2679–2694
Tsay A, Allen TJ, Proske U (2015) Position sense at the human forearm after conditioning elbow muscles with isometric contractions. Exp Brain Res 233(9):2635–2643
Tsay A, Giummarra M, Allen TJ, Proske U (2016) The sensory origins of human position sense. J Physiol 594(4):1037–1049
Tsuge M, Izumizaki M, Kigawa K, Atsumi T, Homma I (2012) Interaction between vibration-evoked proprioceptive illusions and mirror-evoked visual illusions in an arm-matching task. Exp Brain Res 223(4):541–551
Velay JL, Roll J, Paillard J (1989) Elbow position sense in man: contrasting results in matching and pointing. Hum Mov Sci 8:177–193
Walsh LD, Proske U, Allen TJ, Gandevia SC (2013) The contribution of motor commands to position sense differs between elbow and wrist. J Physiol 591(Pt 23):6103–6114
White O, Proske U (2009) Illusions of forearm displacement during vibration of elbow muscles in humans. Exp Brain Res 192(1):113–120
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Tsay, A., Allen, T.J. & Proske, U. Position sense at the human elbow joint measured by arm matching or pointing. Exp Brain Res 234, 2787–2798 (2016). https://doi.org/10.1007/s00221-016-4680-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-016-4680-y