
Abstract
Mango (Mangifera indica) is a widely enjoyed tropical fruit that is rich in numerous nutrients, but it is also a common allergenic fruit that can induce anaphylactic shock in some mango-sensitized individuals. To protect allergic consumers, a TaqMan real-time PCR is developed herein by targeting the major mango allergen GAPDH gene to identify mango in foods. To prevent interference by other ingredients in the foods, an internal amplification control (IAC) is established and incorporated into the developed qPCR. Under the optimized assay conditions, the developed assay can detect mango genomic DNA down to 10 pg/μL and effectively differentiate mango from 21 other fruits and vegetables. In the incurred assay, the developed qPCR has a limit of detection (LOD) of 10 μg/g and 330 μg/g of mango powder in juice and cookie, respectively. The assay exhibits a board spectrum, successfully detecting six mango cultivars, and has good accuracy and precision as calculated intra- and inter-assay CV values < 10%. Forty commercial processed foods were simultaneously analyzed using the developed method and a qPCR method that is officially recognized by the Taiwan FDA. The sensitivity and specificity of this assay were estimated to be 85% and 100%, respectively, based on the ingredient labeling of these products. Hence, this developed qPCR is an effective method for screening mango in processed foods.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Food allergy is an immunoreaction with specific symptoms that occur when individuals come into contact with allergenic foods. Food allergies are estimated to affect about 5% of adults and 8% of children globally, making them an important public health problem [1]. Food allergies are correlated with genetic and environmental factors such as gender, ethnic culture, dietary habits, use of medicines, and exposure to allergens [2]. Various countries and geographic areas have different major allergenic foods, to which consumers must be alerted on their packaging. In the USA, the nine major food allergens recognized are peanuts, milk, eggs, fish, crustacean shellfish, soybean, tree nuts, wheat, and sesame [3]. Additionally, the Codex list identifies cereals containing gluten, crustacea, egg, fish, peanut, milk, sesame, and tree nuts as the recommended global priority allergens [4]. Apple, a member of the Rosaceae family, is the most commonly allergenic fruit in Central and Northern Europe and peach, also a member of that family, is the most commonly allergenic fruit in the Mediterranean region; while mango, a member of the Anacardiaceae family, is a major allergenic fruit in East Asia [5,6,7,8].
Mango (Mangifera indica) might have originated in northwestern Myanmar, Bangladesh, or northeastern India. It is often used as a major ingredient in products such as purees, beverages, jams, jellies, ice creams, cakes, pickles, and candies since it tastes sweet and is rich in nutrients, such as β-carotene and vitamin C [9]. However, mango is also an allergenic fruit to which allergenic reactions include immediate hypersensitive reactions and delayed reactions. The former occurs within minutes of consumption, causing symptoms such as anaphylaxis, angioedema, erythema, urticaria, and wheezing dyspnea; the latter occurs within 2 to 6 h, causing such symptoms as contact dermatitis, periorbital edema, eczematous rash, oral allergy syndrome (OAS), and blistering around the lips [10, 11]. The prevalence of mango allergies correlates most strongly with geographic location. For instance, it has reported that 2% of 100 patients with rhinitis, atopic dermatitis, and bronchial asthma in India have been found to be allergic to mango [12]. In Henan Province of China, 4.94% of 15,534 patients with suspected allergies were diagnosed with mango allergy [13]. In Japan, among 59 adolescents with Pollen-food allergy syndrome, 13.6% were determined to have mango allergy [14].
A mango allergy prevalence of 18.5% in 2086 food-allergic people in Taiwan has been estimated, making mango the most allergenic fruit in the country [15]. Furthermore, a recent study estimated a prevalence of mango allergy of 3.6% among Taiwanese children, highlighting a significant portion of the child population in Taiwan as being allergic to mango [16]. Several mango allergens have been identified. In Paschke's study, two major allergens from mango, namely Man i 1 and Man i 2, have been identified. Man i 1 has a molecular weight of 40 kDa, and Man i 2 has a molecular weight of 30 kDa [17]. Additionally, our research group has identified the 40-kDa mango major allergen as a glyceraldehyde 3-phosphate dehydrogenase (GAPDH) [18]. It is worth noting that several other allergens, such as wheat allergen (Tri a 34), the indoor mold Aspergillus versicolor spore allergen (Asp v 1–3), American cockroach allergen (Per a 13), and striped catfish allergen (Pan h 13), have also been classified as GAPDH [19,20,21,22]. These findings provide valuable insights into the shared allergenic properties of GAPDH proteins across various species, including those found in food sources. Profilin in mango has been identified as the minor allergen Man i 3 (14 kDa), whose structure is similar to that of birch tree profilin, causing cross-reactions between mango and pollen [23]. Since no cure has been developed for mango allergy, which can induce anaphylactic shock in mango-sensitized individuals, a reliable and sensitive method for identifying mango in foods is urgently needed.
To prevent accidental exposure to food allergens, a precautionary allergen label (PAL) on packaging is commonly required to warn food-allergic people of a significant risk of allergic reactions to the product [24]. However, cross-contamination by food allergens can happen during food manufacture: the handling of materials, the sharing of equipment, storage and transportation, leading to risks of unexpected exposure to hidden food allergens. Thus, reliable and sensitive methods for detecting allergens in foods are necessary. Methods for identifying food allergens are typically divided into protein-based and DNA-based methods, based on the target molecule. The former directly target allergens; typical assays are the enzyme-linked immunosorbent assay (ELISA), the lateral flow assay (LFA), and the liquid chromatography–tandem mass spectrometry (LC–MS/MS) [25]. LC–MS/MS is a high-throughput method and often used for multiplex food allergen detection, but it involves a relatively costly, sophisticated device [26]. ELISA and LFA are user-friendly and suitable for routine analysis. To identify mango allergen residues in foods, a monoclonal antibody-based sandwich ELISA has been developed with a limit of detection (LOD) of 3.9 ng/mL of recombinant mango major allergen GAPDH, and an LFA that uses immunomagnetic nanoparticles has been established with an LOD of 6.2 ng/mL of this specific target protein [27, 28].
DNA is thermally stable and highly tolerant of many harsh conditions during food processing and sample extraction, making it a good target for the identification of allergenic materials in processed foods [29]. To detect food allergens, the targets of DNA-based methods can be either an allergen gene or a specific DNA region in ribosomal, mitochondrial or chloroplast DNA in the allergenic food, which are detected using the polymerase chain reaction (PCR), real-time quantitative PCR (qPCR), or loop-mediated isothermal amplification (LAMP). For example, the chloroplast trnL gene of wheat, barley or rye has been identified using PCR to detect gluten contamination in gluten-free foods, and 16S rDNA of a crustacean detected using PCR was used to identify crustacean in processed foods [30, 31]. qPCR methods that use SYBR Green have been used to detect the walnut jug r3 and hazelnut allergen genes (cor a9, cor a11, and cor a13) [32, 33]. TaqMan probe-based qPCR has been used to identify peanut ara h2, cashew ana o1, and chestnut cas s9 allergen genes [34,35,36]. The TaqMan probe-based qPCR is a fast method with high specificity and sensitivity. It exhibits a low LOD, at the parts per million (ppm) level, which depends on factors such as the copy number of the target gene, matrix effect, processing effect, and DNA extraction efficiency [37]. For instance, in Sanchiz’s study, the Ct values for detecting peanut targeting trnH-psbA showed significant variation among peanut samples processed with boiling, controlled instantaneous depressurization, and autoclave treatments [38]. Many commercial TaqMan probe-based qPCR kits have been launched for detecting allergenic foods, such as peanut, hazelnut, walnut, wheat, barley, rye, and buckwheat.
In this study, a TaqMan probe-based qPCR assay that targets the major mango allergen GAPDH gene was developed to identify mango in processed foods. An IAC was also developed to prevent false negative results due to the presence of PCR inhibitors in foods. Two food models (juice and cookie) were used to find the optimal DNA extraction method and to study the effects of the food matrix and processing on assay performance. Finally, the developed assay was used to analyze 40 processed foods to verify its effectiveness in detecting mango in foods. These samples were also analyzed using a TaqMan probe-based qPCR method that is officially recognized by the Taiwan FDA and specifically targets the internal transcribed spacer 2 (ITS2) region of the ribosomal DNA of mango, to verify the results of the developed assay.
Materials and methods
Materials
Mango (Mangifera indica), other fruits, vegetables, and other food products were purchased from local supermarkets in Taiwan. Cetyltrimethylammonium bromide (CTAB) was purchased from VWR International LLC. (Radnor, PA, USA). Ethylenediaminetetraacetic acid (EDTA), sodium dodecyl sulfate (SDS), beta-mercaptoethanol (β-ME), and Tris hydrochloride (Tris–HCl) were obtained from Sigma-Aldrich Corporation (St. Louis, MO, USA). ABI Fast SYBR Green Master Mix, TaqMan Universal Master Mix II, and RNAse A were purchased from Thermo Fisher Scientific Inc. (Waltham, MA, USA). Primer sets and probes were synthesized by Genomics (Taipei, Taiwan), and their sequences are provided in Table 1. BamHI, NdeI, and T4 DNA ligase were obtained from New England Biolabs (Ipswich, MA, USA). A T&A cloning kit and pUC19 plasmid were purchased from Yeastern Biotech Co., Ltd (Kaohsiung, Taiwan). DNA polymerase was purchased from GeneMark (Taipei, Taiwan).
DNA extraction
Solid samples of fruits, vegetables, crackers, and cookies, were ground into fine powders for DNA extraction. Liquid samples of juices and drinks underwent DNA extraction without pre-treatment. Samples that were analyzed using the developed qPCR assay were extracted by the CTAB-SDS method [41]. Samples were ground and incubated with 400 μL TES buffer (0.1 M Tris, 10 mM EDTA and 2% SDS, pH 8.0) at 60 °C for 1 h. Then 5 M NaCl was used to adjust the final salt concentration to 1.4 M, and 10% CTAB solution was added to yield a final concentration of 2%. The mixture was incubated at 65 ℃ for 30 min and centrifuged for 10 min. The supernatant was isolated, mixed with an equal volume of PCIA solution (phenol:chloroform:isoamyl alcohol = 25:24:1, v/v), and centrifuged. The supernatant was then transferred to a new tube, extracted with an equal volume of CIA solution (chloroform:isoamyl alcohol = 24:1, v/v) and centrifuged. It was then transferred to a fresh tube, mixed with 1.5 μL of RNase A (20 mg/mL) and incubated at 37 ° for 30 min. After two volumes of absolute alcohol were added to precipitate DNA overnight at − 20 °C, the sample was centrifuged. The supernatant was removed and pellets were washed twice using cold 70% alcohol, dried in a vacuum centrifuge concentrator, and dissolved in 50 μL of double distilled water (ddH2O).
Two DNA extraction CTAB-PVP and CTAB-PVP-βME methods were tested. The CTAB-PVP method is adapted from that in Pafundo’s work [42]. Briefly, pellets of samples were first mixed with extraction buffer (2% CTAB, 0.1 M Tris, 20 mM EDTA, 1.4 M NaCl, 1% PVP) and then 5 M NaCl was added to yield a final concentration to 2 M. After it was incubated at 65 °C for 30 min, the sample was centrifuged. The supernatant was further processed by following the same steps as in the CTAB-SDS method. The third method, CTAB-PVP-βME, is based on the CTAB-PVP method with the addition of 2% of β-mercaptoethanol (β-ME) to the extraction buffer, which is pre-warmed before extraction. The subsequent steps were the same as those in the CTAB-SDS and CTAB-PVP methods. The quality of DNA was evaluated as the ratio of absorbance at 260 nm to that at 280 nm, and the DNA concentration (ng/μL) was estimated from the absorbance at 260 nm using a NanoDrop™ 1000 spectrophotometer (Thermo Fisher Scientific, MA, USA). All DNA samples were stored at −20 °C until use.
Primer and probe design
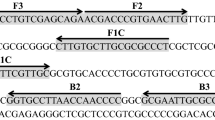
To design primers for the identification of the major mango allergen GAPDH gene, its genomic structure was obtained by an in silica analysis by the NCBI (National Center for Biotechnology Information), and the entire genomic sequence was obtained in a chromosome walking experiment [43]. For the in silica analysis, the total genomic sequence (Project ID: PRJNA771370) of 392 million bases of the tropical fruit Mangifera indica (cultivar Alphonso Mango; Taxonomy ID: 29780) was retrieved from the NCBI genome assembly CATAS_Mindica_2.1 (RefSeq GCF_011075055.1) [44]. The draft genomic sequence was assembled de novo from the reads into 20 chromosome contigs using the ABySS package v.2.1.5. From the assembled 14 M genomic contig (pseudomolecules Chromosome 14, NC_058150.1), a putative exon–intron structure of mango GAPDH gene (ANQ43386.1) was determined using GeneWise v.2.4.1. The genomic sequence of GAPDH gene was obtained by performing a chromosome walking experiment, with primer sets that were designed based on the cDNA sequence of mango GAPDH (accession number KU975060.1), published by our group [18]. Figure S1 and Table S1 present the strategy and primer sets that were used herein. Primer 3 was used for primer design and the optimal primer set for identifying the major mango allergen GAPDH gene was selected owing to its specificity in differentiating mango from nine commonly consumed fruits and vegetables—celery, guava, papaya, grape, orange, tomato, carrot, peach, and pineapple—using the SYBR Green qPCR method. The SYBR Green qPCR reaction was conducted in a final volume of 20 μL, containing 10 μL 2X SYBR Green Master Mix, 400 nM of each of the forward and the reverse primer, and 2 μL of 33 ng/μL sample DNA and added ddH2O up to a final volume of 20 μL. The qPCR was performed with the following program; initial denaturation at 95 °C for 10 min, followed by 40 cycles of denaturation at 95 °C for 5 s and primer annealing and elongation at 60 °C for 30 s. A melting curve analysis of each sample was performed from 60 to 95 °C with intervals of 0.3 °C to identify the melting temperature (Tm) of the qPCR amplicons. PCRs with primer sets that are specific to the ITS2 region of mango and the 5.8S rRNA gene of plants were used to verify DNA extraction results [39]. Primers SA442-F/R were designed based on the gene that encodes SA442 in Staphylococcus aureus and used to construct the IAC [45].
Construction of internal amplification control (IAC) and real-time qPCR
The addition of an IAC in assay development ensures result accuracy and reliability by addressing false negatives caused by qPCR reaction inhibitors in food samples. In this study, the IAC was incorporated with the primers and probes, allowing simultaneous amplification of the target gene and IAC plasmid. A negative target gene result with a positive IAC result indicates a true negative for tested samples, while a negative result for both indicates the presence of qPCR reaction inhibitors, leading to a false negative outcome. Utilizing the IAC in the developed qPCR method ensures the accuracy and reliability of obtained results. The IAC primer set IAC-F/R primers were designed by connecting partial sequences of SA442-F/R primers to the 3’ ends of i4e5-F/R primers that were used to identify the major mango allergen GAPDH gene; the scheme is presented in Fig. 1a. Briefly, the SA442-F/R primers were used to amplify the 94 bp amplicon of the SA442 gene from S. aureus (Fig. 1b), which was used as a template and amplified using IAC-F/R primers to produce a 99 bp product, which was further used as template and reacted with i4e5-F/R primers to generate a 115 bp IAC product [46]. The obtained IAC amplicon was ligated into a TA vector, and the IAC region was cut from the TA vector using BamHI and NdeI; it was then ligated into a pUC19 plasmid. The IAC-pUC19 plasmid was transformed into Escherichia coli DH5α, and the positive clones were identified using a PCR screening method with i4e5-F/R primers to generate a 115 bp IAC amplicon (Fig. 1b). The IAC-pUC19 plasmid (2839 bp) was purified using the method that was developed by Birnboim et al. [47], and its copy number was calculated as follows: \(\frac{[\mathrm{amount }\left(\mathrm{ng}\right) \times {6.02\times 10}^{23} (\mathrm{number}/\mathrm{mole})]}{[\mathrm{length }\left(\mathrm{bp}\right) \times 650 (\frac{\mathrm{g}}{\mathrm{mole}}\mathrm{x bp })\times 1\times {10}^{9}\mathrm{ng}/\mathrm{g})}\). One ng of IAC-pUC19 plasmids was calculated to contain 3.26 × 108 copies.

IAC construction. a IAC construction and b amplification results. Lane M: 100 bp DNA ladder. Lane B: negative control. Lane 1: 94 bp amplicon of SA442 gene of Staphylococcus aureus. Lane 2: 115 bp amplicon of the selected IAC-pUC19 plasmid amplified using i4e5-F/R primers
The real-time qPCR was carried out in a final volume of 20 μL, comprising 10 μL TaqMan™ Universal Master Mix II, 600 nM of each i4e5-F/R primers, 200 nM of IAC probe, 250 nM of target probe, 1 μL of IAC template (815 copies), 5 μL of sample DNA ,and added ddH2O up to a final volume of 20 μL. The program was as follows; activation at 50 °C for 2 min, initial denaturation at 95 °C for 10 min, 45 cycles of denaturation at 95 °C for 15 s, and primer annealing and elongation at 62 °C for 1 min.
Assay performance
The performance of the developed qPCR was evaluated in terms of sensitivity, specificity, broad-spectrum, accuracy, and precision. The sensitivity of the assay in detecting mango DNA was obtained using the assay to analyze various amounts of mango genomic DNA (0–104 pg/μL), and thus plot a dose–response curve. The LOD and amplification efficiency (E) values were, thus, estimated. Additionally, the incurred assay was used to detect the major mango allergen GAPDH gene in mango-spiked cookie and juice samples with thermal processing. Independent triplicate incurred juice and cookie samples were prepared by adding freeze-dried mango powder to cookie and juice. Mixing was conducted by vortexing for 1 min to ensure the homogenous distribution of the mango powder in the juice. After mixing, juice samples were heated in a water bath at 90 °C for 1 min before extraction, and cookie samples were baked at 180 °C for 15 min and cooled to room temperature before being ground. To obtain incurred matrices that contained various amounts of mango powder, non-contaminated matrices were mixed with incurred matrices. Pure pineapple juice was used as a juice model; it comprised pineapple and water. The cookie dough comprised flour (21.2%, w/w), sugar (30.3%, w/w), egg (30.3%, w/w), and low-fat milk (18.2%, w/w). To evaluate assay specificity, DNA extracts of 21 fruits and vegetables were prepared and analyzed using the developed assay. To examine the broad spectrum of the developed qPCR, genomic DNA from six mango cultivars—Irwin, Tu, Yuwen, Tainung No.1, Jinhuang, and Wuxiang—were tested. The accuracy and precision of the developed assay were evaluated using inter- and intra-assays using mango-spiked cookie and juice. The intra-assay was conducted with 20 replications in 1 day, and the inter-assay was performed in duplicate on 10 consecutive days.
Statistical analysis
The results in this study are provided as mean ± standard deviation (SD) from triplicate measurements (n = 3). Statistical analyses were conducted using the SAS statistical package (SAS Institute, NC, USA). Differences in the results were regarded as statistically significant at a p value of less than 0.05.
Results and discussion
Primer selection and IAC construction
The goal of this study is to develop a TaqMan probe-based qPCR method to detect the major mango allergen GAPDH gene. First, the cDNA sequence of mango GAPDH gene was compared to GAPDH coding sequences of other food plants [18]. However, its cDNA sequence shares high identities with those of carrot (84%), tomato (83%), papaya (82%), apple (84%), and orange (88%), possibly because GAPDH is an essential enzyme in glycolysis and the Calvin–Benson cycle for plant cells [48]. The putative genomic structure of the major mango allergen GAPDH gene was determined by retrieving a total genomic sequence of Mangifera indica [44] to comprise ten exons with nine introns, as shown in Fig. S2-a. A total of 2109 bp of the sequence of the target gene were identified by chromosome walking (Fig. S2-b). As shown in Fig. S2-a, four primer sets were selected to identify the target gene, based on their locations in the intron or the junction between the intron and the exon, where sequences for species identification may be potentially conserved [49]. These primer sets were tested for their specificity in differentiating mango from nine food plants using the SYBR Green qPCR method. As shown in Fig. S3, i4e5-F/R primers reacted most strongly with mango and with better specificity than the other three primer sets. Therefore, this set was selected for subsequent assay development.
Food samples normally contain numerous PCR inhibitors, such as polyphenols and pigments. To eliminate the possibility of false negative PCR results, an internal amplification control (IAC) plasmid was constructed by incorporating the SA442 gene—a species-specific DNA fragment to Staphylococcus aureus. Consequently, this DNA fragment of S. aureus should not be present in properly prepared food products, and thus it can be employed as an IAC to co-amplify with the target mango allergen gene in this study. As shown in Fig. 1a, the IAC was established by including an SA442 gene fragment (94 bp) from S. aureus using SA442-F/R primers, and this fragment was further applied as a template to produce a 99 bp IAC product using the IAC-F/R primers, a pair of hybridized primers which are a hybridization of SA442 and i4e5 primers. Furthermore, i4e5-F/R primers were used to produce a 115 bp IAC amplicon, which was used to construct a 2839 bp IAC-pUC19 plasmid. To confirm the success of IAC insertion, i4e5-F/R primers were used to amplify the selected IAC-pUC19 plasmid. As shown in Fig. 1b, a 115 bp IAC amplicon was successfully produced, as confirmed by DNA sequencing. The IAC-pUC19 plasmid was used as an IAC for further assay development.
Assay optimization
Several factors were optimized in the development of a TaqMan probe-based qPCR method; they were annealing temperature (Ta), primer concentration, and probe concentration. A properly designed qPCR assay amplifies the target DNA with an amplification efficiency of 90–110% and a correlation coefficient (R2) > 0.98 [50]. First, the Ta for i4e5-F/R primers was optimized by analyzing serial dilutions of mango genomic DNA with three values of Ta (60 °C, 62 °C, and 64 °C). As shown in Fig. 2, only the assay with a Ta of 64 °C had a significantly low amplification efficiency of 79.63%, whereas those with Tas of 60 °C and 62 °C had efficiencies of 92.30% and 91.40%, respectively. The Ct values of the assay with a Ta of 64 °C were higher than those of the assays with Tas of 60 °C and 62 °C, possibly owing to the decrease in fluorescence intensity that is caused by an increase in Ta. As shown in Fig. S4, in analyzing a low concentration of target DNA (0.01 ng/μL), the delta Rn (△Rn) value of the assay with a Ta of 64 °C was only approximately 6.5% and 13.3% of those with Tas of 60 °C and 62 °C, respectively. Additionally, in detecting 0.01 ng/μL mango DNA, the assay with a Ta of 60 °C yielded a positive ratio of 1/3 whereas that with a Ta of 62 °C yielded a positive ratio of 3/3. The positive ratio at 60 °C might have been lower because 60 °C is suitable for amplifying both target gene and the IAC. However, 0.01 ng/μL of mango DNA was too little to compete with IAC for the qPCR reagents, yielding negative results in the detection target gene. Thus, 62 °C was selected for further assay optimization.

Optimization of the annealing temperature (Ta) of the developed qPCR. Dose–response curves at three annealing temperatures (60 °C, 62 °C, and 64 °C) were plotted using the developed qPCR with i4e5-F/R primers to analyze samples that contained different concentrations of mango genomic DNA
The objective of this study is to establish a competitive TaqMan probe-based qPCR for identifying the target mango allergen gene. In a competitive IAC model, both the target and IAC are amplified using the same set of primers. However, in a non-competitive IAC model, the target and IAC are amplified using different primer sets. The use of different primer sets in the non-competitive IAC model may not accurately reflect the amplification results of the primary target. In a competitive IAC model, the competition between the IAC with target DNA for primers and other qPCR reagents affects the efficiency of the PCR and potentially reduces assay sensitivity [51]. Thus, various combinations of concentrations of i4e5 primers, the target probe, and the IAC probe were tested to optimize the developed qPCR. As shown in Table 2, when a 400 nM primer was used, the positive ratios of the IACs of these six reactions were all below 3/3, possibly because not enough primers were present for the simultaneous amplification of the IAC and the target gene. When a 600 nM primer was used, three assays with a 100 nM IAC probe still yielded the positive rations of the IACs below 3/3. Among the three assays with the 600 nM primer and the 200 nM IAC probe, the Ct values for the target gene did not vary significantly as their positive ratios for the target and the IAC were all 3/3. Based on these results, the 600 nM primer, the 250 nM target probe, and the 200 nM IAC probe were determined to be optimal for the developed qPCR.
DNA extraction
Food components, such as polyphenols, polysaccharides, lipids, and proteins, can affect DNA purity and interfere with qPCR amplification. An effective DNA extraction method has a high extraction efficiency, favoring the assay sensitivity of the qPCR. To determine the optimal DNA extraction method, pineapple juice and pancake powder were used as liquid and solid food models; these were spiked with mango and then extraction was carried out using the three methods of CTAB-SDS, CTAB-PVP, and CTAB-PVP-β-ME. β-ME is used to prevent the oxidation of polyphenols and PVP is used to remove polyphenols from DNA samples [41, 42]. In Fig. 3a, samples that were obtained using the CTAB-SDS method had the lowest Ct values for all spiking levels. The DNA pellets that were generated by CTAB-PVP method were yellowish, possibly retaining some of the polyphenols from the juice. Polyphenols from plants were easily oxidized and irreversibly bound to DNA, making their elimination difficult [52]. Additionally, PVP might have caused the hyperchromic effect by partially degrading dsDNA to ssDNA, increasing the absorbance at 260 nm, leading to an overestimation of the DNA concentration [53]; PVP might have co-precipitated with DNA, inhibiting the qPCR [54]. The high Ct values of samples that were obtained using CTAB-PVP-β-ME might have been caused by the contamination by β-ME, which is a PCR inhibitor and a DNA cutter [55]. In Fig. 3b, pancake samples that were spiked with the smallest amount of mango powder (0.033%) could only be detected in the extracts that were obtained by the CTAB-SDS method. Therefore, the CTAB-SDS method was subsequently used to isolate DNA from liquid and solid processed foods.

Selection of DNA extraction methods for a liquid food model: pineapple juice spiked with different amounts of mango puree (0.05–50%, w/v), and b solid food model: pancake powder spiked with various amounts of freeze-dried mango powder (0.033–33%, w/w). DNA were individually extracted from spiked samples by CTAB-SDS, CTAB-PVP, and CTAB-PVP-β-ME methods. The extracted DNA samples were analyzed by the developed qPCR assay to obtain their Ct values
Assay performance
To determine assay sensitivity, various concentrations of mango genomic DNA were analyzed using the assay to generate a dose–response curve. As shown in Fig. S5, the linearity was good (R2 = 0.9974), with a 3-log-unit dynamic range (10–10,000 pg/μL). The assay detected the target gene down to a concentration of 10 pg/μL of mango genomic DNA. To study the effects of the food matrix and processing on assay sensitivity, pre-spiked and incurred assays were performed using juice and cookie as liquid and solid food models, respectively. In the pre-spiked and incurred assays of juice samples, the LOD value was 10 μg/g of mango with a dynamic range of 10–10,000 μg/mL (Fig. 4a); in the pre-spiked and incurred assays of cookie samples, the LOD value was 0.033% of mango with a dynamic range as 330–100,000 μg/g (Fig. 4b). The values of R2 and amplification efficiency (E) from these four calibration curves satisfied the accepted criteria for a qPCR assay, with the exception of the E value for the incurred cookie samples [50]. The juice samples yielded higher SD values than the cookie samples, possibly owing to the presence of fibers, polysaccharides, and pigments in the juice, potentially affecting the DNA extraction and amplification [56]. These compounds may not be uniformly dispersed, especially in highly viscous juice samples. Despite homogenization, inherent variability introduced by these components may result in a larger standard deviation in juice samples compared to cookies. This hypothesis explains the observed high standard deviation values in the qPCR analysis of juice samples. Further investigation is required to better understand the potential causes behind these high standard deviation values. At each spiking amount, the Ct values of the pre-spiked samples were lower than those of the incurred samples, indicating that thermal processing might have caused irreversible binding of food compounds to DNA or DNA fragmentation, disfavoring DNA extraction and qPCR amplification [57]. The LOD of the developed assay in detecting the incurred juice (10 μg/g) was about 1/30th that in detecting the incurred cookie (0.033%; 330 μg/g), possibly because cookies were heated under harsher conditions (180 °C, 15 min) than the juice (90 °C, 1 min). Additionally, the constituents of the cookie (flour, egg, milk, and sugar) were more complicated than those of the juice (pineapple and water), leading to a strong matrix effect on DNA extraction and amplification [58]. Based on these results, we strongly recommend plotting a calibration curve for a qPCR method with matrix adaption and corresponding thermal processing to determine precisely the amount of the target in processed foods. To the best of our knowledge, the eliciting threshold of mango allergy has not been established. The lack of clinical research on this subject is likely due to the localized nature of mango allergy, primarily occurring in specific regions of Southeast Asia, thereby limiting its global significance as a public health concern. Consequently, determining a detection limit for an assay based on the eliciting threshold of mango allergy is currently challenging. In terms of detection limits, the official Taiwan FDA qPCR method can detect mango at a limit of 0.1% (w/w) in foods, while the developed qPCR assay in this study achieves a more sensitive detection limit of 0.033% (w/w) mango powder in cookies. Therefore, the developed qPCR assay demonstrates higher sensitivity compared to the official Taiwan FDA method, providing a potential alternative for the detection of mango residues in food samples.

Calibration curves obtained by the developed qPCR, targeting the major mango allergen GAPDH gene using juice and cookie models that were mixed with different amounts of freeze-dried mango powder with and without thermal treatment. a Pre-spiked (◆) and incurred (▓) juice samples were mixed with 10–10,000 μg/g lyophilized mango powder without or with heating at 90 °C for 1 min, respectively. b Pre-spiked (◆) and incurred (▓) cookie samples were mixed with 330–100,000 μg/g lyophilized mango powder without or with heating at 180 °C for 10 min, respectively. DNA was first extracted from these pre-spiked or incurred samples using the CTAB-SDS method, and then analyzed using the developed qPCR assay
The specificity of the developed assay is critical and was evaluated by analyzing DNA samples that were extracted from 21 commonly consumed fruits and vegetables, As shown in the amplification plot in Fig. 5, only the mango sample gave a positive result and no positive reaction occurred in the analysis of DNA extracts of the other tested samples. In Fig. S3, cross-reactions were identified in analyses of celery, tomato, carrot, and pineapple, using an SYBR Green qPCR with i4e5-F/R primers. When the target probe was used, these non-specific results were eliminated, indicating that the developed TaqMan probe-based qPCR had good specificity in the identification of mango. Six mango cultivars (Irwin, Tu, Yuwen, Tainung No. 1, Jinhuang, and Wuxiang) were analyzed to assess the spectrum of the developed assay. Figure 6 shows that the method identified all of the mango cultivars except Jinhuang with similar sensitivities. The LOD value for Jinhuang was 0.1 μg/mL of genomic DNA, which was ten times those (0.01 μg/mL) for the other cultivars. In addition, the official Taiwan qPCR method was applied to analyze these six mango cultivars, using equal amounts of DNA (0.033 ng). The obtained Ct values for these cultivars did not show significant differences, with a range of Ct value of 27.21 ± 0.07 to 28.27 ± 1.85. However, the Tu mango cultivar displayed a slightly higher Ct value of 31.42 ± 0.70 compared to the other cultivars (data not shown). Based on the results obtained from the official Taiwan qPCR, the lower amplification observed specifically in the Jinhuang cultivar when using the developed qPCR assay could potentially be attributed to either a lower copy number of the target gene or a single nucleotide polymorphism (SNP) in the GAPDH gene among different mango cultivars, which could affect the annealing efficiency of the i4e5 primers to the target DNA [59]. Several factors, such as mismatch type and position, local sequence, KCl concentration, and type of polymerase, can influence primer extension in the presence of SNP. Notably, the presence of 3’ mismatches significantly decreases the optimal annealing temperature [60]. The hypothesis of gene copy number or SNP on qPCR amplification warrants further study of the full sequencing data of the mango allergen GAPDH gene of various mango cultivars, which are lacking in the open database. To further investigate the hypothesis regarding gene copy number or SNP influence on qPCR amplification, a thorough analysis of the full sequencing data of mango allergen GAPDH gene from various mango cultivars is required. Unfortunately, such data are currently lacking in the open database. To evaluate the precision and reproducibility of the developed qPCR, mango-spiked juice and cookie samples were tested using the inter-assay and intra-assay. In Table S2, the recovery rates of juice samples in the intra-assay and inter-assay were determined to be 94.84–98.93% and 91.05–107.41%, respectively, and the coefficients of variation (CV) were calculated to be 1.93–1.04% and 0.60–1.06%, respectively. The recovery rates of cookie samples for the intra-assay and inter-assay were determined to be 90.58–105.48% and 93.53–97.66%, respectively, and the CV values were 1.72–2.27% and 1.16–1.72%, respectively. The CVs for both the inter- and intra-assays of juice and cookie samples were all below 15%, revealing the high precision and reproducibility of the developed qPCR [61].

Specificity of the developed qPCR in differentiating mango (red) from 21 non-target food plants (orange; as shown in inset). The IAC result of testing mango is shown in green; those of testing non-targets are shown in blue

The broad spectrum of the developed assay, obtained by utilizing it to test six mango cultivars—Irwin, Tu, Yuwen, Tainung No. 1, Jinhuang, and Wuxiang. These mango cultivars were extracted using the CTAB-SDS method. Their DNA samples were diluted to five concentrations (0.01–10 μg/mL) and analyzed by the developed assay
Analysis of processed foods
To establish the feasibility of using the established qPCR to analyze processed foods, it was used to test 40 food products, including ice bars, juices, cakes, crackers, cookies, and jellies. The results of the qPCR were verified using an official qPCR method that is recognized by the Taiwan FDA, as shown in Table 3. In Taiwan, the packaging of foods that contain mango must display a food allergen alert. Twenty food products (No. 1–10 and No. 21–30) that contain mango all had food allergen alerts for mango, indicating thorough implementation of the mango allergen alert policy in Taiwan. The target gene was detected in 17 of those 20 food products, yielding a sensitivity of 85% (17/20). The three food products in which no mango was detected were a mango cookie (No. 26) and two mango jellies (No. 29 and No. 30), but no mango was detected in the two mango jelly products using the official qPCR method that specifically targets the ITS2 region of mango. These two items were manufactured using concentrated mango juice and the double processing procedure might have caused DNA damage, and thus the negative results. The consistent findings obtained from the analysis of the two mango jelly products further reinforce the reliability and accuracy of the developed qPCR method for identifying the mango allergen gene in processed foods. Moreover, processed foods may still contain DNA residues that affect the results of the qPCR, so 20 mango-free food products (No. 11–20 and No. 31–40) were tested. The developed method yielded negative results for all 20 of the non-mango-containing products, generating a specificity of 100% (20/20). In analyzing pineapple juice (No. 20), the official method yielded a false positive result with a 1/3 positive ratio, perhaps because of the sequence similarity in the ITS2 region between mango and pineapple. Based on the results of the food analyses, the sensitivity of the developed assay was calculated to be 85% (17/20) and its specificity was calculated to be 100% (20/20), indicating the effectiveness of the developed qPCR method in identifying mango in processed foods. In this study, the developed qPCR method specifically targets the mango allergen GAPDH gene, enabling a direct and relevant assessment of the potential allergenicity of tested samples. In contrast, the Taiwan FDA reference method primarily focuses on the ITS-2 region for species identification. Additionally, this method incorporates an IAC to ensure result accuracy and reliability by effectively identifying false negatives that may arise due to the presence of qPCR reaction inhibitors in food samples. These distinct features, including the targeted allergen gene and the inclusion of the IAC, make this developed method a valuable complement to existing techniques. It enhances allergen detection in processed foods and contributes to food safety and labeling regulations, thus addressing an important need in the field. Furthermore, when comparing the performance of the presented qPCR method with the previously developed LFA method by our research group for detecting mango allergens in processed foods, the qPCR method demonstrated superior specificity. Notably, the LFA method produced two false positive results in the analysis of two mango-free products: a vegetable fruit juice and pineapple juice (listed as food sample No. 13 and No. 20 in Table 3, respectively). These false positive outcomes can be attributed to cross-reactivity with the antibodies utilized in the LFA, as our previous research demonstrated that the crude protein extracts of carrot, celery, and pineapple exhibit cross-reactivity with the LFA antibodies [28]. In contrast, the developed qPCR method consistently generated negative results for these two samples, underscoring its remarkable specificity in detecting the mango allergen gene in processed foods.
Conclusion
In this study, a TaqMan probe-based qPCR with the IAC was developed for detecting mango in processed foods. CTAB-SDS was the best tested extraction method for isolating DNA from juice and cookie samples. Under optimal assay conditions, the LOD of this assay was determined to be 10 pg/μL with a 3-log-unit dynamic range (10–10,000 pg/μL) for mango genomic DNA. The results of incurred assays revealed the LOD of this assay was estimated to be 10 μg/g mango powder in juice and 330 μg/g (w/w) of mango powder in cookie. Finally, the developed assay was used to analyze 40 processed foods that were simultaneously analyzed using an official qPCR assay that targets the ITS-2 region of mango. The developed assay yielded negative results in analyzing three mango-containing food (No. 26, mango cookie; No.29 and No.30, mango jelly) and the official method also yielded negative results for the two mango jelly products. The assay yielded all negative results with 20 mango-free products. Therefore, its sensitivity and specificity were determined to be 85% and 100%, respectively. To the best of the authors’ knowledge, this work is the first to have established a qPCR for detecting the mango allergen GAPDH gene in food products. This assay provides an alternative method for screening food products for mango DNA residues.
Data availability
Data will be made available on request.
References
Sicherer SH, Sampson HA (2014) Food allergy: epidemiology, pathogenesis, diagnosis, and treatment. J Allergy Clin Immunol 133(2):291–307. https://doi.org/10.1016/j.jaci.2013.11.020
Loh W, Tang ML (2018) The epidemiology of food allergy in the global context. Int J Environ Res Public Health 15(9):2043. https://doi.org/10.3390/ijerph15092043
FDA. Food allergies. https://www.fda.gov/food/food-labeling-nutrition/food-allergies. Accessed 5 Jun 2023
World Health Organization (2022) Risk assessment of food allergens. Part 1: review and validation of Codex Alimentarius priority allergen list through risk assessment: Meeting report. https://www.fao.org/3/cb9070en/cb9070en.pdf Accessed 5 Jun 2023
Fernández-Rivas M (2015) Fruit and vegetable allergy. In Food Allergy: Molecular Basis and Clinical Practice 101:162–170. https://doi.org/10.1159/000375469
Hassan AK, Venkatesh YP (2015) An overview of fruit allergy and the causative allergens. Eur Ann Allergy Clin Immunol 47(6):180–187. DOI: https://doi.org/10.12688/f1000research.109080.2
Gabrielli S, Clarke AE, Morris J, Gravel J, Lim R, Chan ES, Goldman RD, O’Keefe A, Gerdts J, Chu DK, Upton J, Hochstadter E, Moisan J, Bretholz A, McCusker C, Zhang X, Protudjer JL, Abrams EM, Simons E, Ben-Shoshan M (2021) Fruit-induced anaphylaxis: clinical presentation and management. Allergy Clin Immunol Pract 9(7):2825–2830. https://doi.org/10.1016/j.jaip.2021.02.055
Costa J, Mafra I (2022) Rosaceae food allergy: a review. Crit Rev Food Sci Nutr. https://doi.org/10.1080/10408398.2022.2045897
Sareen R, Shah A (2011) Hypersensitivity manifestations to the fruit mango. Asia Pac Allergy 1(1):43–49. https://doi.org/10.5415/apallergy.2011.1.1.43
Hegde VL, Venkatesh YP (2007) Anaphylaxis following ingestion of mango fruit. J Investig Allergol Clin Immunol 17(5):341–344
Ukleja-Sokołowska N, Gawrońska-Ukleja E, Lis K, Żbikowska-Gotz M, Sokołowski Ł, Bartuzi Z (2018) Anaphylactic reaction in patient allergic to mango. Allergy Asthma Clin Immunol 14:78. https://doi.org/10.1186/s13223-018-0294-1
Mehta D, Dagar A, Kishan J, Singh P, Nehra T, Sharma H (2018) Common allergens prevalent in and around ambala, haryana: an intradermal study among patients with asthma and allergic rhinitis and atopic dermatitis. Indian J Dermatol 63(4):311–316. https://doi.org/10.4103/ijd.IJD_438_17
Sun X, Zhao J, Wang Q, Shi G, Yang J, Ming L (2019) Prevalence of allergen sensitization among 15,534 patients with suspected allergic diseases in Henan Province, China. Asian Pac J Allergy Immunol 37(2):57–64. https://doi.org/10.12932/AP-160817-0137
Kiguchi T, Yamamoto-Hanada K, Saito-Abe M, Sato M, Irahara M, Ogita H, Miyagi Y, Inuzuka Y, Toyokuni K, Nishimura K, Ishikawa F, Miyaji Y, Kabashima S, Fukuie T, Narita M, Ohya Y (2021) Pollen-food allergy syndrome and component sensitization in adolescents: a Japanese population-based study. PLoS ONE 16(4):e0249649. https://doi.org/10.1371/journal.pone.0249649
Wu TC, Tsai TC, Huang CF, Chang FY, Lin CC, Huang IF, Chu CH, Lau BH, Wu L, Peng HJ, Tang RB (2012) Prevalence of food allergy in Taiwan: a questionnaire-based survey. Intern Med J 42(12):1310–1315. https://doi.org/10.1111/j.1445-5994.2012.02820.x
Li SK, Liu Z, Huang CK, Wu TC, Huang CF (2022) Prevalence, clinical presentation, and associated atopic diseases of pediatric fruit and vegetable allergy: a population-based study. Pediatr neonatol 63(5):520–526. https://doi.org/10.1016/j.pedneo.2022.03.019
Paschke A, Kinder H, Zunker K, Wigotzki M, Weßbecher R, Vieluf D, Steinhart H (2001) Characterization of allergens in mango fruit and ripening dependence of the allergenic potency. Food Agric Immunol 13(1):51–61. https://doi.org/10.1080/09540100051074220
Tsai WC, Wu TC, Chiang BL, Wen HW (2017) Cloning, expression, and purification of recombinant major mango allergen Man i 1 in Escherichia coli. Protein Expr Purif 130:35–43. https://doi.org/10.1016/j.pep.2016.06.009
Sander I, Rozynek P, Rihs HP, Van Kampen V, Chew FT, Lee WS, Kotschy-Lang N, Merget R, Brüning T, Raulf-Heimsoth M (2011) Multiple wheat flour allergens and cross-reactive carbohydrate determinants bind IgE in baker’s asthma. Allergy 66(9):1208–1215. https://doi.org/10.1111/j.1398-9995.2011.02636.x
Benndorf D, Müller A, Bock K, Manuwald O, Herbarth O, Von Bergen M (2008) Identification of spore allergens from the indoor mould Aspergillus versicolor. Allergy 63(4):454–460. https://doi.org/10.1111/j.1398-9995.2007.01603.x
Asero R, Pravettoni V, Scala E, Villalta D (2020) House dust mite-shrimp allergen interrelationships. Curr Allergy Asthma Rep 20(4):9. https://doi.org/10.1007/s11882-020-0902-2
Ruethers T, Taki AC, Karnaneedi S, Nie S, Kalic T, Dai D, Daduang S, Leeming M, Williamson NA, Breiteneder H, Mehr SS, Kamath SD, Campbell DE, Lopata AL (2021) Expanding the allergen repertoire of salmon and catfish. Allergy 76(5):1443–1453. https://doi.org/10.1111/all.14574
Song J, Zhang H, Liu Z, Ran P (2008) Mango profilin: cloning, expression and cross-reactivity with birch pollen profilin Bet v 2. Mol Biol Rep 35(2):231–237. https://doi.org/10.1007/s11033-007-9075-5
DunnGalvin A, Chan CH, Crevel R, Grimshaw K, Poms R, Schnadt S, Taylor SL, Turner P, Allen KJ, Austin M, Baka A, Baumert JL, Baumgartner S, Beyer K, Bucchini L, Fernández-Rivas M, Grinter K, Houben GF, Hourihane J, Kenna F, Kruizinga AG, Lack G, Madsen CB, Mills EN, Papadopoulos NG, Alldrick A, Regent L, Sherlock R, Wal JM, Roberts G (2015) Precautionary allergen labelling: perspectives from key stakeholder groups. Allergy 70(9):1039–1051. https://doi.org/10.1111/all.12614
Senyuva HZ, Jones IB, Sykes M, Baumgartner S (2019) A critical review of the specifications and performance of antibody and DNA-based methods for detection and quantification of allergens in foods. Food Addit Contam Part A 36(4):507–547. https://doi.org/10.1080/19440049.2019.1579927
López-Pedrouso M, Lorenzo JM, Gagaoua M, Franco D (2020) Current trends in proteomic advances for food allergen analysis. Biology 9(9):247. https://doi.org/10.3390/biology9090247
Tsai WC, Yin HY, Chen SN, Chang HC, Wen HW (2021) Development of monoclonal antibody-based sandwich ELISA for detecting major mango allergen Man i1 in processed foods. J Food Saf 41(2):e12884. https://doi.org/10.1111/jfs.12884
Tsai WC, Yin HY, Lin YT, Liu HJ, Tseng CY, Wen HW (2021) A rapid lateral flow assay using immunomagnetic nanoparticles for detecting mango allergen residues in processed foods. J Food Saf 41(6):e12929. https://doi.org/10.1111/jfs.12929
Holzhauser T (2018) Protein or no protein? Opportunities for DNA-based detection of allergenic foods. J Agric Food Chem 66(38):9889–9894. https://doi.org/10.1021/acs.jafc.8b03657
Melinte G, Hosu O, Cristea C, Marrazza G (2022) DNA sensing technology a useful food scanning tool. TrAC. https://doi.org/10.1016/j.trac.2022.116679
Herrero B, Vieites JM, Espiñeira M (2012) Fast real-time PCR for the detection of crustacean allergen in foods. J Agric Food Chem 60(8):1893–1897. https://doi.org/10.1021/jf2043532
Linacero R, Ballesteros I, Sanchiz A, Prieto N, Iniesto E, Martinez Y, Pedrosa MM, Muzquiz M, Cabanillas B, Rovira M, Burbano C, Cuadrado C (2016) Detection by real time PCR of walnut allergen coding sequences in processed foods. Food Chem 202:334–340. https://doi.org/10.1016/j.foodchem.2016.01.132
Iniesto E, Jiménez A, Prieto N, Cabanillas B, Burbano C, Pedrosa MM, Rodríguez J, Muzquiz M, Crespo JF, Cuadrado C, Linacero R (2013) Real Time PCR to detect hazelnut allergen coding sequences in processed foods. Food Chem 138(2–3):1976–1981. https://doi.org/10.1016/j.foodchem.2012.11.036
López-Calleja IM, de la Cruz S, Pegels N, González I, García T, Martín R (2013) Development of a real time PCR assay for detection of allergenic trace amounts of peanut (Arachis hypogaea) in processed foods. Food Control 30(2):480–490. https://doi.org/10.1016/j.foodcont.2012.09.017
Sanchiz Á, Ballesteros I, Marqués E, Dieguez M, Rueda J, Cuadrado C, Linacero R (2018) Evaluation of locked nucleic acid and TaqMan probes for specific detection of cashew nut in processed food by real time PCR. Food Control 89:227–234. https://doi.org/10.1016/j.foodcont.2018.02.021
Sanchiz Á, Ballesteros I, López-García A, Ramírez A, Rueda J, Cuadrado C, Linacero R (2020) Chestnut allergen detection in complex food products: development and validation of a real-time PCR method. LWT 123:109067. https://doi.org/10.1016/j.lwt.2020.109067
Linacero R, Sanchiz A, Ballesteros I, Cuadrado C (2020) Application of real-time PCR for tree nut allergen detection in processed foods. Crit Rev Food Sci Nutr 60(7):1077–1093. https://doi.org/10.1080/10408398.2018.1557103
Sanchiz A, Sánchez-Enciso P, Cuadrado C, Linacero R (2021) Detection of peanut allergen by Real-Time PCR: looking for a suitable detection marker as affected by processing. Foods (Basel, Switzerland) 10(6):1421. https://doi.org/10.3390/foods10061421
Method of Test for Plant-Derived Ingredients in Foods Qualitative Test of Mango Ingredients (2013) Taiwan Food and Drug Administration (TFDA), Taipei. https://www.fda.gov.tw/tc/includes/GetFile.ashx?id=f636694248527946380&type=2&cid=1737. Accessed 7 Jun 2023
Peters RP, van Agtmael MA, Gierveld S, Danner SA, Groeneveld AB, Vandenbroucke-Grauls CM, Savelkoul PH (2007) Quantitative detection of Staphylococcus aureus and Enterococcus faecalis DNA in blood to diagnose bacteremia in patients in the intensive care unit. J Clin Microbiol 45(11): 3641–3646. https://doi.org/10.1128/JCM.01056-07
Möller EM, Bahnweg G, Sandermann H, Geiger HH (1992) A simple and efficient protocol for isolation of high molecular weight DNA from filamentous fungi, fruit bodies, and infected plant tissues. Nucleic Acids Res 20(22):6115–6116. https://doi.org/10.1093/nar/20.22.6115
Pafundo S, Gullì M, Marmiroli N (2011) Comparison of DNA extraction methods and development of duplex PCR and real-time PCR to detect tomato, carrot, and celery in food. J Agric Food Chem 59(19):10414–10424. https://doi.org/10.1021/jf202382s
Parker JD, Rabinovitch PS, Burmer GC (1991) Targeted gene walking polymerase chain reaction. Nucleic Acids Res 19(11):3055–3060. https://doi.org/10.1093/nar/19.11.3055
Wang P, Luo Y, Huang J, Gao S, Zhu G, Dang Z, Gai J, Yang M, Zhu M, Zhang H, Ye X, Gao A, Tan X, Wang S, Wu S, Cahoon EB, Bai B, Zhao Z, Li Q, Wei J, Chen H, Luo R, Gong D, Tang K, Chen Y (2020) The genome evolution and domestication of tropical fruit mango. Genome Biol 21(1):60. https://doi.org/10.1186/s13059-020-01959-8
Martineau F, Picard FJ, Roy PH, Ouellette M, Bergeron MG (1998) Species-specific and ubiquitous-DNA-based assays for rapid identification of Staphylococcus aureus. J Clin Microbiol 36(3):618–623. https://doi.org/10.1128/jcm.36.3.618-623.1998
Siebert PD, Larrick JW (1993) PCR MIMICS: competitive DNA fragments for use as internal standards in quantitative PCR. Biotechniques 14(2):244–249
Birnboim HC, Doly J (1979) A rapid alkaline extraction procedure for screening recombinant plasmid DNA. Nucleic Acids Res 7:1513–1523. https://doi.org/10.1093/nar/7.6.1513
Zaffagnini M, Fermani S, Costa A, Lemaire SD, Trost P (2013) Plant cytoplasmic GAPDH: redox post-translational modifications and moonlighting properties. Front Plant Sci 4:450. https://doi.org/10.3389/fpls.2013.00450
Martins RP, Leach RE, Krawetz SA (2001) Whole-body gene expression by data mining. Genomics 72(1):34–42. https://doi.org/10.1006/geno.2000.6437
Bustin S, Huggett J (2017) qPCR primer design revisited. Biomol Detect Quantif 14:19–28. https://doi.org/10.1016/j.bdq.2017.11.001
Hoorfar J, Malorny B, Abdulmawjood A, Cook N, Wagner M, Fach P (2004) Practical considerations in design of internal amplification controls for diagnostic PCR assays. J Clin Microbiol 42(5):1863–1868. https://doi.org/10.1128/jcm.42.5.1863-1868.2004
John ME (1992) An efficient method for isolation of RNA and DNA from plants containing polyphenolics. Nucleic Acids Res 20(9):2381. https://doi.org/10.1093/nar/20.9.2381
Abramo MD, Castellazzi CL, Orozco M, Amadei A (2013) On the nature of DNA hyperchromic effect. J Phys Chem B 117:8697–8704. https://doi.org/10.1021/jp403369k
Puchooa D (2004) A simple, rapid and efficient method for the extraction of genomic DNA from lychee (Litchi chinensis Sonn.). Afr J Biotechnol 3(4):253–255. https://doi.org/10.5897/AJB2004.000-2046
Yee W, Abdul-Kadir R, Lee LM, Koh B, Lee YS, Chan HY (2018) A simple and inexpensive physical lysis method for DNA and RNA extraction from freshwater microalgae. 3 Biotech 8:354. https://doi.org/10.1007/s13205-018-1381-1
Schrader C, Schielke A, Ellerbroek L, Johne R (2012) PCR inhibitors–occurrence, properties and removal. J Appl Microbiol 113(5):1014–1026. https://doi.org/10.1111/j.1365-2672.2012.05384.x
López-Andreo M, Aldeguer M, Guillén I, Gabaldón JA, Puyet A (2012) Detection and quantification of meat species by qPCR in heat-processed food containing highly fragmented DNA. Food Chem 134(1):518–523. https://doi.org/10.1016/j.foodchem.2012.02.111
Sidstedt M, Rådström P, Hedman J (2020) PCR inhibition in qPCR, dPCR and MPS—mechanisms and solutions. Anal Bioanal Chem 412(9):2009–2023. https://doi.org/10.1007/s00216-020-02490-2
Lefever S, Pattyn F, Hellemans J, Vandesompele J (2013) Single-nucleotide polymorphisms and other mismatches reduce performance of quantitative PCR assays. Clin Chem 59(10):1470–1480. https://doi.org/10.1373/clinchem.2013.203653
Rejali NA, Moric E, Wittwer CT (2018) The effect of single mismatches on primer extension. Clin Chem 64(5):801–809. https://doi.org/10.1373/clinchem.2017.282285
Jelliffe RW, Schumitzky A, Bayard D, Fu X, Neely M (2015) Describing assay precision—reciprocal of variance is correct, not CV percent: its use should significantly improve laboratory performance. Ther Drug Monit 37(3):389–394. https://doi.org/10.1097/FTD.0000000000000168
Funding
The authors would like to thank the Ministry of Science and Technology of the Republic of China, Taiwan, for financially supporting this research under the contract MOST 107-2320-B-005-011-MY3.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that there are no conflicts of interest among them.
Compliance with ethics requirements
This article does not contain any studies with human participants or animals performed by any of the authors.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Yin, HY., Liu, YF., Lin, YY. et al. Development of a TaqMan real-time PCR assay with an internal amplification control for detecting trace amounts of allergenic mango (Mangifera indica) in commercial food products. Eur Food Res Technol 249, 2647–2660 (2023). https://doi.org/10.1007/s00217-023-04310-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00217-023-04310-0