
Abstract
Panicoid grasses and crops include finger millet, switch grass, foxtail millet, napier grass, and pearl millet. Foxtail millet is an important C4 panicoid crop with a small genome size (~490 Mb), a short life cycle, inbreeding characteristics, and great abiotic stress tolerance. It is an important food and fodder crop in arid and semiarid parts of Asia and Africa, including North China and India. Setaria italica (cultivated) and Setaria viridis (wild) are two well-known Setaria species that have served as ideal model systems for evolutionary, architectural, and physiological studies in related potential bioenergy. Foxtail millet has high genetic diversity, with numerous core and mini-core collections holding a wide range of genotypes. Significant phenotypic variations enable the discovery and mining of new abiotic stress tolerance variants for crop improvement. Several foxtail millet accessions may be drought and salinity tolerant, or rather exploiting their agronomic and stress-tolerant traits may be especially important for marker-assisted selection and genetic engineering. In addition, the release and availability of the foxtail millet genome sequence have revealed numerous of its unique properties, such as abiotic stress tolerance, which may aid in a better understanding of its evolution, stress physiology, and adaptation. The foxtail millet genome sequence thus aids not only in the identification and introgression of agronomically important traits but also in the deciphering of this exceptionally stress-tolerant crop’s abiotic stress tolerance mechanisms, in addition to the evolution of climate-resilient crops, which is critical in this epoch of global climate change.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
2.1 Introduction
Agriculture’s advancement in some crop species was the most pivotal point in human history, and plant breeding plays a key role in supporting agricultural production by altering the genetic constituents of crop species in relation to their economic use. Systematic and planned studies on plant breeding began in the nineteenth century, following the rediscovery of Mendel’s laws of inheritance. The most significant limitations of traditional breeding methods are that selection is based on phenotype, which produces misleading results because it is heavily influenced by the environment. However, environmental effects have been lessened through the discovery of molecular markers that aid in the selection of desirable traits in an indirect manner. After the 1990s, the emergence of recombinant DNA technology, DNA sequencing methodologies, and other advanced breeding techniques marked turning points in several omics fields (Fig. 2.1). The genomics toolbox contains linkage and physical maps, genome sequencing, bioinformatics software and databases, and new generations of omics tools like transcriptomics, proteomics, metabolomics, genomics, and high-throughput phenomics. Indeed, over the last few decades, it had made spectacular radiances in genomics research. For example, at the dawn of the genomics era, Arabidopsis was chosen as the first model for whole-genome sequencing, followed quickly by the sequencing of other model genomes (rice for monocots, Medicago and Lotus for legume crops, and poplar for tree species) and crop species (soybean, cassava, sorghum, maize, pigeon pea, wheat, finger millet, foxtail millet, etc.). Generated sequence data is being analyzed in parallel for characterizing genes and validating their functions using comparative and functional genomics approaches. This information can be applied to evolutionary significance and crop improvement programs. There are currently a plethora of high-throughput genome sequencing technologies and tools, omics platforms, and strategies available for using genomics in plant breeding. However, the high costs of omics research, both invested in and associated with it, currently limit the implementation of omics-based crop breeding, particularly for underutilized, potential, minor, and orphan crops.

Historical timeline of plant breeding with respect to Omics
Foxtail millet (Setaria italica) is a small diploid C4 crop species whose genome has been sequenced collaboratively by the Joint Genome Institute (JGI), Berkeley, California, USA, and the Beijing Genome Initiative (BGI), China (Singh et al. 2017). It serves as a model for other biofuel grasses such as napier grass, switch grass, and pearl millet. The foxtail millet’s final drought genome size is (490 Mb), fastened onto nine chromosomes, and functional 38,801 genes are annotated. Because of its small genome size, short life cycle, and inbreeding nature, foxtail millet is an excellent model crop for studies in plant nutritional biology, stress response studies, and C4 metabolism research.
2.2 Characterization of Abiotic Stress Tolerance Mechanism in Foxtail Millet
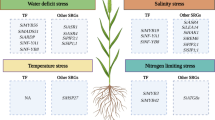
Drought, salt, water logging, high temperatures, and mineral toxicity all limit growth and development of plants, resulting in potential yield losses of more than half of major agricultural crop production. Positive feedback and tolerance to survive and reproduce under such stressful conditions is a complicated performance that is largely coordinated and maintained by correcting crops’ physiological, cellular, and molecular activities (Chinnusamy et al. 2004; Ahuja et al. 2010). As a result, addressing intricate regulatory pathways and the biological theories governing abiotic stress tolerance in plants necessitates a thorough understanding of genome-scale metabolic responses at critical growth stages of development regarding external triggers. Advanced technologies, particularly omics approaches, are of top priority and promising approaches for improving foxtail millet abiotic stress tolerance (Fig. 2.2).

Abiotic stress resistance responses to develop improved foxtail millet cultivars through different omics research
Omics refers to all a molecular biology applications that require a molecular understanding of stress response, viz., genomics, functional genomics, genetic engineering, gene expression, proteomics or metabolomics(s), and to understand their ultimate phenotypic consequences in response to environmental changes, which are usually followed by appropriate modifications in the crop transcriptome, proteome, or metabolome (Ahuja et al. 2010). Because of its dwarf genome size and diploidy, foxtail millet has been studied and characterized more thoroughly than other millet crops under abiotic stress conditions. Genome-wide studies and expression profiling studies on foxtail millet are underway. Furthermore, proteomic and metabolomic studies on millets in response to abiotic stress are still far behind. Gene modification, including regulatory genes, has proven to be more responsive than using mono or polygenes for improving abiotic stress tolerance in millets. As a result, recent advances in “omics” technology related to genome sequences have created a plethora of opportunities for increasing foxtail millet crop stress tolerance against abiotic stressors (Fig. 2.3) (Table 2.1).

Flow diagram showing different omics strategies
2.2.1 Genomics Studies in Foxtail Millets to Understand Abiotic Stress
The study of the structure and function of a living organism’s genome is known as genomics. The foxtail millet is an excellent model plant for a wide range of plant nutritional biology, stress-responsive investigations, and C4 metabolism research (Zhang et al. 2012). Because of its small genome size (490 Mbp), small plant size, and quick generation time, the foxtail millet genome sequencing provides a platform for organizing knowledge on various millets and comprehending the shared and independent features of C4 plant evolution. Characterization of plant genomes and the genes contained within them will aid geneticists and molecular biologists in their quest to understand C4 metabolism physiology and will help plant breeders in their goal of developing better products. It serves as a reference genome for molecular biological studies in the cereals, for the following reasons:
-
(i)
Availability of complete genome sequences of both genotypes, Yugu1 and Zhang gu (Bennetzen et al. 2012; Zhang et al. 2012), which enabled investigations of transcriptome activity in a range of tissues and developmental; stages.
-
(ii)
Large genetic resources for Millet Genome Database (https://db.cngb.org/datamart/plant/DATApla6/).
-
(iii)
Identification of candidate stress-responsive genes (Sood and Prasad 2017).
-
(iv)
Well-established genetic transformation techniques (Sood et al. 2020).
-
(v)
Strong genome databases facilitating access and depositing of information (Bonthala et al. 2013).
-
(vi)
The high-resolution mapping of epigenetic modifications for chromosomes (Ni et al. 2017; Wang et al. 2017).
-
(vii)
The genome-wide identification of genetic variation in gene expression between different Setaria species (Jaiswal et al. 2019).
Among the most important genomic resources for genetic studies and molecular breeding approaches are molecular markers, genetic maps, and sequence information. As a result, genomic research is needed to develop molecular markers and identify quantitative trait loci (QTLs), as well as to use marker-assisted breeding to develop improved cultivars. Furthermore, these genetic resources aid crop development efforts by reducing the time and labor-intensive direct screening of germplasm cultivated in field and greenhouse conditions. Furthermore, genetic maps and molecular markers are used to help with comparative genomic research in agricultural plants. The foxtail millet genome sequences, as well as genetic and physical maps, are valuable tools for the discovery and characterization of genes and QTLs for a wide range of agronomic traits, which can help with marker-aided breeding (MAB) and crop improvement. Wang et al. (1998) identified 160 loci in the first foxtail millet genetic map based on restriction fragment length polymorphism (RFLP) markers in an inter-varietal cross. Since then, numerous researchers and scientific communities have been involved in the development of linkage maps, as well as the identification of numerous genes in foxtail millet using genomic or EST-based molecular markers for further use in genotyping, phylogenetic relationship, and transferability studies (Jia et al. 2007; Jia et al. 2009; Heng et al. 2011; Gupta et al. 2012; Gupta et al. 2013). Using referenced-based foxtail millet genome assembly and de novo assembly, Xu et al. (2013) created new 9576 and 7056 microsatellites, respectively. Several other researchers used Setaria genome sequence data to identify microsatellite repeat motifs (Pandey et al. 2013; Zhang et al. 2014), eSSRs (Kumari et al. 2013), and TE-based markers (Kumari et al. 2013; Yadav et al. 2014a). Gupta et al. (2011) and Muthamilarasan et al. (2014a) generated 98 and 5123 intron length polymorphic (ILP) markers in foxtail millet, respectively, and demonstrated the utility of ILP markers in cross-species transferability, germplasm characterization, comparative mapping, and genomic studies among millets or non-millet crop plants, respectively. Another noteworthy study produced 176 microRNA (miRNA)-based molecular markers from 335 mature foxtail millet miRNAs (Khan et al. 2014, Yadav et al. 2014b). Lata et al. (2011a) identified a synonymous SNP in a novel DREB2-like gene associated with dehydration tolerance and developed an allele-specific marker (ASM). The ASM was also validated in a core set of over 100 foxtail millet accessions, with regressions of lipid peroxidation (LP) and relative water content (RWC) on the ASM revealing that the SiDREB2-associated trait accounted for roughly 27% and 20% of total variation, respectively. As a result, the significance of this QTL for foxtail millet dehydration tolerance is demonstrated (Lata and Prasad 2011; Lata and Prasad 2013).
Furthermore, Jia et al. (2013) developed a haplotype map using 916 diverse foxtail millet accessions that yielded a total of 2,584,083 SNPs, of which 845,787 SNPs (minor allele frequency > 0.05) were responsible for haplotype map. In genomics-assisted crop breeding, these marker resources are used efficiently from time to time. Other than these markers, amplified fragment length polymorphism (AFLP) inter simple sequence repeat (ISSR), and orthologous microsatellite markers have been employed to analyze genetic relationships, investigate domestication-related geographical structure, and phylogenetic relationships between foxtail millet and wild relatives (Le Thierry d’Ennequin et al. 2000; Li and Brutnell 2011; Zhang et al. 2012). In addition to these, these are also utilized to evaluate genetic diversity, population structure, and linkage disequilibrium (LD) in various foxtail accessions (Li and Brutnell 2011; Vetriventhan et al. 2014) as well as for examining important agronomic traits (Mauro-Herrera et al. 2013; Gupta et al. 2014). These markers and markers associated studies if once validated, might be used to identify new genes/QTLs linked to traits of interest and, consequently, in the marker-assisted breeding in foxtail millet and related crops (Muthamilarasan and Prasad 2015). Qie et al. (2014) identified QTLs for a number of traits namely promptness index, radical root length, coleoptile length, lateral root number at germinating stage, and seedling survival rate in 190 F7 RILs population derived from foxtail millet cultivar (Yugu1) x wild Setaria viridis genotype. A total of eighteen QTLs were detected in nine linkage groups and among them, nine QTLs explained more than 10% of phenotypic variance for a given trait. Feng et al. (2016) and Li et al. (2017) identified six abscisic acid stress ripening genes SiASR1 to SiASR6 in tobacco and foxtail millet which enhanced drought and oxidative tolerance in plants through decreased H2O2 accumulation and improved antioxidant enzyme activities. Meng et al. (2021) conducted research to predict the cold responsiveness in different cold-tolerant grass species namely foxtail, proso, pearl millet, and switchgrass. In genomic assembly of four species identified 2605, 1872, 4888, and 3148 cold-responsive genes, among them 1154, 992, 1761, and 1609 cold-responsive genes showed comparative syntenic relationships. Non-syntenic genes, on the other hand, responded to cold stress in a more varied way across species than syntenic genes. Chen et al. (2021) identified ARF gene family which participated in stress tolerance in foxtail millet. Twenty-four SiARF genes were identified using GWAS and their expression not only revealed their potential functions in stress response but also indicates their functional redundancy (Chen et al. 2021). Stomatal density and canopy temperature are the most important traits that could be reduced to improve water use efficiency (WUE) in C4 crops. Therefore, quantitative trait loci (QTL) mapping was performed in RILs population derived from a Setaria italica × Setaria viridis cross under control and drought conditions and identified multiple QTLs that highlight the potential of Setaria as a model to understand the stress biology in C4 species (Prakash et al. 2021).
2.2.2 Functional Genomics
Extensive research using functional genomics techniques like overexpression, gene silencing, insertional mutagenesis, and targeted induced local lesion in genome (TILLING) has been critical in refining our understanding of complex regulatory networks involved in stress response, adaptability, and tolerance in plants throughout the post-genomic era. Various omics technologies are also generating massive data sets in millets, which could be used to identify important candidate genes for crop development projects utilizing MAS or transgenic technology. Hundreds of discovered genes or proteins still need to be functionally verified, despite the advent of genome sequencing programs and proteomics. Various reverse genetics methods will be essential in these instances. Overexpression of a gene in a homologous or heterologous system via a constitutive or stress-inducible promoter is a useful method for determining gene function. Knocking down or reducing the expression of a gene is another important method for determining its function. Transgenomics, often known as transgenic technology, is a gene-based functional genomics method that can provide crucial information on the regulatory mechanisms that support plant stress tolerance. Transgenomics in millets is still neglected, despite the economic and nutritional importance.
The majority of candidate genes functionally validated in Arabidopsis or tobacco in the case of millet crops are transcription factors (TFs), which can influence the expression of downstream stress-sensitive genes and thus play an important role in stress tolerance. Because of its close similarity to possibly bioenergy grasses, the foxtail millet is an ideal candidate for genetic change among millet crops. A novel gene from foxtail millet namely SiPf40 that is the member of ZIP gene family was characterized (Liu et al. 2009) and revealed that its overexpression causes vessel enlarging and xylary fibers enhancing with significantly decreased IAA content that could be enhanced cold tolerance in foxtail millet in lesser extent. Despite the fact that Agrobacterium-mediated transformation is becoming more common, there has only been one report of improved abiotic stress tolerance in foxtail millet. The SiARDP overexpression in foxtail millet improved drought tolerance, whereas constitutive expression of SiARDP in Arabidopsis thaliana improved drought tolerance throughout seed germination and seedling development. The expression levels of target genes of SiARDP were upregulated in transgenic Arabidopsis and foxtail millet. These finding revealed that SiARDP, one of the genes targeted by SiAREB, is involved in ABA-dependent signalling pathways, and plays a critical role in the abiotic stress response in plants (Li et al. 2014). Gene silencing is a well-known reverse genetics approach for discovering the function of a known gene in an organism that produces morphological changes. As a result, virus-induced gene silencing (VIGS) with dsRNA has emerged as a valuable and simple functional genomics method (Baulcombe 2004). VIGS works by utilizing viral vectors that contain a target region of the host gene to decrease the expression of target plant genes. The foxtail millet has been chosen as a high-throughput platform for virus-induced gene silencing (VIGS) to anticipate gene inhibition findings due to its proximity to a number of bioenergy grasses (Doust et al. 2009). The VIGS technique for functional assessment of cell wall associated genes in foxtail millet was developed by Ramanna et al. (2013) using the Brome mosaic virus as a vector. Late embryogenesis abundant (LEA) proteins played important roles in protecting higher plants from by environmental stresses. Wang et al. (2014) characterized a novel LEA gene (SiLEA14) from foxtail millet which was expressed in roots, stems, leaves, inflorescences, and seeds at different levels under normal growth conditions. Overexpression of this gene significantly enhances the salt and drought tolerances of foxtail millet due to responsiveness to ABA, PEG, and NaCl. Furthermore, SiLEA14 could be used in crops genetic engineering with the aim of improving stress tolerance. Nitrogen (N) is the macronutrient and is one of the most limiting factors in agricultural plant production which deficiency or excess tends to slow down plant growth or to produce toxicity that indirectly invites drought vulnerability in crop. Nitrogen is obtained in the form of nitrate and ammonium, which are regulated by nitrate redistribution transporters (NRT) and ammonium transporters (AMT) (AMT) (Loqué and von Wirén 2004; Nadeem et al. 2020). Recently research was conducted to identify the functional genes that respond to NRT and AMT in foxtail millet genotypes. Nadeem et al. (2018) characterized two genes (SiNRT1.11 and SiNRT1.12) for NRT and one gene for ammonium transporters (SiAMT1.1) and reported that N acquisition was accelerated by upregulating these genes expression. Muthamilarasan et al. (2020) performed expression profiling in the Setaria model species namely Setaria italica and Setaria viridis for distinct stress specific genes which were subjected to abiotic stresses showed upregulation of SiαCaH1, SiβCaH5, SiPEPC2, SiPPDK2, SiMDH8, and SiNADP-ME5 in the tolerant cultivar. Overexpression of SiNADP-ME5 in yeast in dehydration stress conditions highlights the putative role in conferring dehydration tolerance. Methylation on nitrogenous bases of DNA may also exhibit stress tolerance. Therefore, research had conducted by Hao et al. (2020) to analyze the differences of millet seedlings under osmotic stress using qRT-PCR, bisulfate sequencing PCR, DNA methyltransferase (DNMT). Findings revealed that osmotic stress induced the transcriptional expression and activity of key enzymes in H2S biosynthesis which may improve osmotic stress tolerance of foxtail millet by mediating DNA methylation. Phosphorus (P), absorbed from soil solutions, is a limiting nutrient for plant growth and yield. PHT1 family genes seem to play a key role for low Pi stress tolerance in foxtail millet. Therefore, twenty genotypes of foxtail millet with contrasting degree of growth and Pi uptake responses under low Pi (LP) and high Pi (HP) supply were screened and identified SiPHT1 genes in shoot tissues of the LP best-performing genotypes (LPBG) (Roch et al. 2020). Gene families namely LEA, DREB, PIP, HSP, and P5CS played crucial roles in salt tolerance and dehydration tolerance. Therefore, da Cunha Valença et al. (2020) exploited qPCR analysis to evaluate the expression of selected genes in roots of Setaria virdis genotypes on three key sampling days (3, 6, and 10) under the stress conditions and stated that all the genes were significantly upregulated on day 3 (da Cunha Valença et al. 2020). Gene family containing CCT motif played a key role in photoperiod-regulation flowering pathway that can enhance multiple stress resistance. Studied phylogenetic relationship among foxtail millet, sorghum, and maize for CCT motif genes. Real-time PCR analysis showed that the expression level of SiCCT gene was higher in leaf than in young panicle and leaf sheath that can exhibit stress tolerance in foxtail millet. PTI1 (Pto-interacting 1) protein kinase proteins participate in a variety of abiotic stress and defense responses against pest or pathogens in several plant species. Huangfu et al. (2021) identified 12 SiPTI1 genes from the foxtail genome using qPCR analysis after induction of multiple stresses. Heterologous expression of SiPTI1–5 in yeast and E. coli enhanced tolerance to salt stress (Huangfu et al. 2021). Plants regulate salt uptake via calcineurin B-like proteins (CBLs). Yan et al. (2021) had done expression analysis and revealed that overexpression of SiCBL5 increased its salt tolerance in foxtail millet through lower accumulations of Na+ and stronger Na+ efflux but its knockdown increased salt hypersensitivity.
2.2.3 Transcriptomics
The term “transcriptome” refers to an organism’s whole set of transcripts (mRNA), whereas “transcriptomics” refers to the study of these mRNAs. The transcriptome approach, in and of itself, provides for a comprehensive analysis and quantification of abiotic stress-induced alterations at the whole-organism level. The use of genome-wide expression profiling to discover candidate genes involved in a variety of biological processes and stress regulation networks has been proven to be beneficial. Transcriptome research is crucial for the development of molecular markers such as microsatellites and single nucleotide polymorphisms (Lata 2015). When whole-genome sequencing is not possible, transcriptome sequencing, or RNA-Seq, becomes more important. It provides cost-effective, functionally relevant information and the production of a large number of genetic resources in a reasonable amount of time as compared to traditional approaches. Differential gene expression, isoforms or allele-specific expression, regulatory element characterization, alternative splicing, RNA editing, population genetics, system biology, etc. are areas that may be benefited from the information generated using transcriptomics analysis (Arya et al. 2015). Sreenivasulu et al. (2004) used a microarray filter with 711 cDNA spots suggesting 620 unigenes from a barley EST collection to identify many differentially expressed transcripts in salt-treated tolerant and sensitive foxtail millet seedlings. Following that, Zhang et al. (2007) used suppression subtractive hybridization and cDNA microarray methods to identify many drought-induced differentially expressed genes in foxtail millet. The expression analysis of these genes revealed that in response to drought stress, different sets of genes are activated in the roots and shoots of foxtail millet, with most of them contributing to the protein degradation pathway. Jayaraman et al. (2008), on the other hand, conducted the first comprehensive study to identify salt-responsive differentially expressed transcripts using the cDNA-AFLP approach in two foxtail millet cultivars, tolerant and sensitive to salinity tolerance, and discovered 90 transcripts derived fragments. Lata et al. (2010) discovered 86 upregulated genes in the resistant foxtail millet cultivar “Prasad” under polyethylene-induced dehydration stress. Puranik et al. (2011) discovered 159 differentially expressed transcripts in a comparative transcriptome study of salinity tolerant and sensitive cultivars in order to better understand the salt tolerance pathway. Deep sequencing was used to analyze the whole transcriptome of foxtail millet by Qi et al. (2013), who discovered 2824 drought-responsive genes with variable expression patterns. Up to 48.23% and 51.77% of the genes were up- and downregulated, respectively. The most frequently upregulated proteins were LEA proteins, dehydrins, HSPs, aquaporins, and phosphatase 2C, indicating a possible role in foxtail millet drought tolerance. Yin et al. (2014) performed transcript profiling on drought-stressed foxtail millet as well. In this study, an F-box protein was discovered to be involved in drought and ABA response. Apart from the aforementioned, thorough genome-wide analyses and expression profiling examinations of numerous TF families such as AP2/ERFs, NAC, C2H2 zinc finger, and MYB (Lata et al. 2014; Puranik et al. 2013; Muthamilarasan et al. 2014b, c) and stress-responsive genes such as WD40, DCL, AGO, and RDR polymerases and ALDH (Mishra et al. 2014; Yadav et al. 2015; Zhu et al. 2014) have also been found in foxtail millet. Studies These were conducted to characterize gene regulatory networks and molecular cross linking for abiotic stress responses in foxtail millet, as well as to analyze gene families involved in stress tolerance, recognize relevant candidate genes, and identify relevant candidate genes.
Bonthala et al. (2014) undertaken in silico study and identified 2297 putative TFs and categorized them in 55 families in foxtail millet genome. Lata et al. (2011b) identified a DREB2-like TF (SiDREB2) that is associated with drought tolerance in foxtail millet accessions that can also be useful for other millet species for the purposes of functional complementation of orthologous genes. Similarly, another abscisic acid (ABA)-responsive DREB-binding protein gene (SiARDP), cloned from foxtail millet that increases tolerance to drought and high salinity stress (Li et al. 2014). Furthermore, also found that two TFs namely SiAREB1 and SiAREB2 were able to bind SiARDP (Li et al. 2017). Tang et al. (2017) studied phenotypic and transcriptomic differences between two foxtail millet drought-sensitive varieties (An04 and Yugu1). Gene expression profiling and a cluster analysis suggested that transcriptomic regulatory mechanisms controlling stress responses in these cultivars were significantly affected by genotype x environment interactions. After combining the RNA sequencing data with previously mapped QTLs (Qie et al. 2014), 20 potential candidate genes were identified. A functional enrichment analysis indicated that genes involved in phytohormone metabolism and signalling, TFs, cellular detoxification, and osmotic adjustment may play important roles in modulating the genotype-specific drought stress responses of foxtail millet. Drought-resistant foxtail millet F1 hybrid (M79) and its parental lines E1 and H1 were subjected to transcriptomic analysis by Shi et al. (2018) and revealed 17 Gene Ontology (GO) enrichments and 14 Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways in hybrid, including photosystem II (PSII) complex, peroxidase activity, plant hormone signal transduction, and chlorophyll biosynthesis. RNA sequencing analysis identified 5258 DEGs between M79 and E1, and 3594 between M79 and H1, indicating that the drought-tolerant cultivar M79 and its parental lines had different transcriptional profiles. These results indicate that various molecular pathways, including photosynthesis, respond to drought stress in M79, and provide abundant molecular information for further analysis of the underlying mechanism responding to this stress. Similarly, recently Pan et al. (2020) employed transcriptomics in foxtail millet cultivars Yugu2 and An04, respectively, and results revealed that 8887 and 12,249 DEGs identified in both cultivars in response to salinity, respectively, and 3149 overlapped between two varieties. Shkolnik et al. (2019) reported that calmodulin-binding transcription activator 6 (CAMTA6) is a key negative regulator of most salt-responsive genes and ion homeostasis during early germination. Recently, in foxtail millet, Pan et al. (2020) identified one CAMTA gene (Si000282m.g), and its decrease also confirmed that Yugu2 is less sensitive to salt. Kumar and Trivedi (2018) explained that overexpression of glutathione-S-transferase (GST)/ glutathione peroxidase (GPX) from different species enhances tolerance toward abiotic stress through assisting peroxide scavenging and oxidative stress metabolism. Therefore, a study was conducted by Pan et al. (2020) to check the expression of GST/GPX in foxtail cultivar and reported higher expression levels of GST, GPX, and peroxidases (POD) possessing stronger antioxidant capacity. Flavonoids produced in foxtail germplasm act as the main anti-salinity effectors which are regulated by a series of transcription factors (TFs). Pan et al. (2020) identified five R2R3 MYB TFs which increased in differentially expressed genes (DEGs) YG2TvsAn04T and YG2TvsYG2C. The identified DEGs of TFs might be regulators for activating the transcription of the anthocyanin biosynthetic genes, and further contribute to flavonoids elevating and salt resistance improvement of foxtail millet. Salt stress inhibits root meristem development by stabilizing IAA17 and reducing PINFORMED (PIN) gene expression, lowering auxin levels and suppressing auxin signalling, respectively ((Liu et al. 2015). Pan et al. (2020) identified five PIN genes and six IAA genes were also evidently repressed in both foxtail millet cultivars An04 and Yugu2, which can result in reduction of auxin accumulation and growth inhibition in root. Stress-induced DELLA accumulation could hold back growth and enhance the tolerance by upregulation of ROS detoxification enzymes under stress. The higher expression level of DELLA genes (Si009972m.g and Si000959m.g) with higher expression level of ROS scavenging related genes conferred that Yugu2 possessing higher salt tolerance than An04. Qin et al. (2020) researched to evaluate different physiological parameters under drought conditions and expression of Δ1-pyrroline-5-carboxylate synthetase genes (SiP5CS1 and SiP5CS2) in foxtail millet cultivar, Jigu 16. RNA sequencing analysis results revealed upregulation of SiP5CS1 and SiP5CS2 under drought stress conditions and described that these genes involve in various physiological functions such as photosynthesis, signal transduction, phenylpropanoid biosynthesis, starch and sucrose metabolism, and osmotic adjustment. Micro RNAs (miRNAs) were playing crucial role in developmental gene expression regulation in plants and act as a important regulator in abiotic stress adaptation. Therefore, a small RNA sequencing approach was employed to profile the salt stress-responsive miRNA landscapes of the shoot and root tissues of two Setaria viridis accessions. RT-PCR analysis identified two regulatory modules (miR160 and miR167) that were accession- and tissue-specific responses in accessions shoot and root tissues and provide tolerance against salt stress (Pegler et al. 2020).
2.2.4 Proteomics
The term “proteome” refers to the entire set of proteins produced or modified by an organism, whereas “proteomics” refers to a large-scale analysis of proteins expressed in a cell, tissue, or organism at a specific stage or condition in order to better understand biology or causal regulatory mechanisms. Every organism’s current state is determined by the proteome, which is essentially a link between the transcriptome and the metabolome. Because mRNA levels are rarely associated with protein accumulation, and proteins have direct effects on biochemical processes, a proteomics approach to evaluating plant stress responses is becoming more important (Gygi et al. 1999). Despite advancements in proteomics research, very few reports on the application of proteomics to crops have been published in comparison to other organisms (Reddy et al. 2012). Furthermore, compared to transcriptome analysis, proteome analysis in plants in response to various stimuli is still relatively limited (Canovas et al. 2004), with only one study reported in foxtail millet among all millets. Veeranagamallaiah et al. (2008) performed proteomic study in 7-day old salt-treated tolerant foxtail millet cultivar “Prasad” using 2D electrophoresis that identified 29 distinct expressed proteins among them some of involved in stress response. Pan et al. (2018) identified a total of 2474 differentially expressed proteins (DEPs) by quantitative proteomic analysis in foxtail millet seedlings for drought tolerance. There were 321 proteins with significant expression changes, including 252 proteins that were upregulated and 69 proteins that were downregulated. Western blot analysis was used to confirm the expression patterns of five drought-responsive proteins. The transcription levels of 21 differentially expressed proteins were also investigated using qRT-PCR. The findings revealed significant inconsistency in the variation between proteins and their corresponding mRNAs, demonstrating once again that posttranscriptional modification plays a critical role in gene expression regulation.
Grain protein contents (GPCs) of seeds is a main indicator of nutritional values of foxtail millet also play roles in drought tolerance. The GPC were also played important roles in different metabolic pathways such as starch and sucrose metabolism, glycolysis/gluconeogenesis, amino acids biosynthesis, detoxification and defense, protein degradation, tricarboxylic acid (TCA) cycle, protein synthesis, energy metabolism, transporter, pentose phosphate pathway, and signal transduction pathway. Li et al. (2019) had done 2D electrophoresis coupled with matrix-assisted laser desorption/ionization-tandem time-of-flight (MALDI-TOF) analysis to characterize the response of foxtail millet under drought stress at a proteomic level. A total of 104 differentially abundant protein spots (DAPs) were identified; among them, 57 were upregulated and 47 were downregulated under drought treatment. This study provides new information on proteomic changes in foxtail millet. Xu et al. (2020) studied proteomic profiles of mature seeds under three drought stress levels, conducted on two varieties, Longgu6 and Huangjinmiao (HJM) of foxtail millet by 2D electrophoresis. Using proteomics study identified levels of DEPs moderate level of drought system. Protein biosynthesis was upregulated, and drought stress-related pathways were enhanced in HJM under moderate level of drought system, thereby resulting in the increase of GPC in HJM under this condition (Xu et al. 2020). Selenium response in foxtail could be useful for analyzing the Se response pathway and producing Se-enriched foxtail millet in the future. Liang et al. (2020) investigated Selenium (Se) effect on foxtail millet proteomic profiling, identified 123 differentially expressed proteins and results showed that foliar sodium selenite (Na2SeO3) applied at critical growth stages significantly improved the Se content in foxtail millet. C-terminally encoded peptides (CEPs) are small peptides, typically posttranslationally modified, and highly conserved in many species. CEPs are known to inhibit plant growth and development. Zhang et al. (2021) identified 14 CEPs in foxtail millet. Moreover, expression of all SiCEPs was induced by abiotic stresses and phytohormones. In the presence of abscisic acid (ABA), growth inhibition and ABA content in seedlings increased with the concentration of SiCEP3 gene.
2.2.5 Metabolomics
The term “metabolome” refers to the totality of low molecular weight biochemicals found in a sample that are either substrates or byproducts of enzymatic processes and have a direct influence on the phenotype of a cell, tissue, or organism. Metabolomics, on the other hand, attempts to assess a sample’s metabolite profile at a specific time, stage, or environment, and thus provides a functional account of an organism’s physiological state. Metabolomics concepts such as target analysis, metabolite profiling, and fingerprinting are examples of metabolomics concepts that can be applied to large-scale applications such as phenotyping transgenics, gene function analysis, significant equivalence testing, and stress response tracking. As a result, metabolomics may be regarded as bridging the genotype–phenotype divide. A complete transcriptome, proteome, and metabolome study is necessary to have a detailed knowledge of both gene function and molecular mechanisms mediating complicated biological processes. Only one research used gas chromatography-time-of-flight mass spectrometry (GC-TOFMS) to evaluate the variety of primary metabolites and phenolic acids in three proso millet types (Kim et al. 2013). A variety of metabolites, including suitable solutes, nitrogen-containing chemicals, antioxidants, and signal macromolecules, have been discovered to be involved in salt tolerance (Kim et al. 2007). Li et al. (2018) recognized and quantified more than 300 secondary metabolites using LC-MS, and inheritance patterns of metabolome in foxtail hybrids. Among them secondary metabolites, flavonoids and its derivatives are also identified that play a key role in drought resistance. The integrative omics analysis suggested that phenyl propanoid, flavonoid, and lignin biosynthesis pathways, as well as lysophospholipids, were important in determining foxtail millet salinity tolerance. Importantly, Yugu2 tolerance was linked to improved ion channel and antioxidant system efficiency. The results of an integrated analysis of metabolomic and transcriptomic data from foxtail millet roots revealed that 17 flavonoids biosynthesis-related genes were significantly upregulated in the Yugu2 cultivar when exposed to salinity. The overabundance of 27 flavonoids metabolites was obtained in accordance with gene expression (Pan et al. 2020).
2.2.6 Phenomics
Phenomics is the high-throughput study of phenotype through the evaluation of an organism’s morphological, physiological, and biological features, as well as the linkage of genetic, epigenetic, and environmental factors (Furbank and Tester 2011; Deshmukh et al. 2014). Phenotyping technology advances have been widely used to analyze germplasm collections in order to predict phenotype based on genetic markers. This method is referred to as “genetic symptoms” (Deshmukh et al. 2014). When combined with other omics tools, phenomics can reveal a great deal about how cell biochemical or biophysical processes result in the final phenotype. Plant phenotypes are inherently complex because they result from genotypes interacting with a wide range of environmental factors (Bilder et al. 2009). Automatic and high-throughput phenotyping platforms have been developed to investigate background variation, trait relationships, plant growth behaviors, and multi-parametric phenotypic data. Automated imaging tools are used to generate nondestructive phenotypic data for quantitative studies such as growth, tolerance, resistance, physiology, and yield that are required to link to molecular studies such as germplasm collections, population mapping, breeding materials, and the design of “omics” and quantitative trait locus (QTLs) studies (Rahaman et al. 2015). The field of plant-based phenotyping is being transformed by automated high-throughput hardware platforms and associated software packages (Rahaman et al. 2015). Fahlgren et al. (2015), for example, studied drought responses in Setaria using a conveyer belt-based platform and scan 3D images. The authors discovered that Setaria virdis grows faster than foxtail millet using image analysis. Specialized imaging systems, such as near infrared (NIR) imaging, spectroscopy imaging, and fluorescence imaging, can also be used to measure physiological traits. For example, Fahlgren et al. (2015) discovered significant differences in water content in Setaria species using NIR imaging. Furthermore, fluorescence imaging accurately measures photosynthesis rate in 2D leaves (Cruz et al. 2016). Spectroscopy imaging can also be used to investigate stress responses (Rahaman et al. 2015), but this technology has yet to be used in millet research. It is critical to have a method for obtaining root images. Rhizotrons are root visualization systems that consist of a thin layer of soil or nutrient substrates sandwiched between two plastic sheets. This system has been used to assess the rate of root growth in pearl millet (Passot et al. 2016). Growth and Luminescence Observatory for Roots (GLO-Roots), an imaging system with a cleaner background, aids in the detection of luciferase reporter expression in S. viridis transgenic lines (Sebastian et al. 2016). As phenotyping systems rapidly develop, it is important to have software packages that can efficiently extract biologically meaningful information from images. Software such as ImageJ, a new generation of high-throughput, customizable and open-source software is available (Singh et al. 2016). PlantCV is another Image Analysis package that has been developed at the Danforth Plant Science Center, Missouri, USA, which detected different timely related changes to water availability between the wild and domesticated Setaria species (Fahlgren et al. 2015) combined with the Bellwether Phenotyping Platform helps to collect data. Setaria dataset of 79,000 images available on the iPlant Data Store and through the Bio-Image Semantic Query User Environment (Bis Que) platform (Kvilekval et al. 2010; http://plantcv.danforthcenter.org/pages/data.html; publicly; Goff et al. 2011). Proximal sensing techniques, which can be used to obtain more accurate plant-level measurements, can be a useful tool in studying plant stress biology. As a result, a study was conducted to assess water use efficiency (WUE) in foxtail millet using thermal imaging and visual-near infrared spectroscopy. The findings indicated that visible-near-infrared reflectance data and thermal images could be used as a potential rapid technique for assessing plant physiological responses such as WUE (Wang et al. 2016).
2.3 Conclusion
In conclusion, foxtail millet can be used as a genetic model to study the domestication process, population structure, comparative mapping, evolution, and physiology of C4 photosynthesis, and biomass production in panicoid grasses. By comparing the genome sequences of foxtail millet and panicoid grasses, potential genes and molecular markers linked with useful agronomic traits can be identified for utilization in crop improvement programs. Foxtail millet can be used to study the regulation of C4 photosynthesis. So, extensive research must be conducted to collect and identify potential germplasm resources of foxtail millet for useful agronomic traits and abiotic stress tolerance. Further studies have to be done on the nutritional and medicinal properties of foxtail millet in order to popularize and increase production globally.
References
Ahuja I, de Vos RCH, Bones AM, Hall RD (2010) Plant molecular stress responses face climate change. Trends Plant Sci 15:1360–1385
Arya L, Verma M, Ram C (2015) Small millets transcriptomics. Transcriptomics 3:e114. https://doi.org/10.4172/2329-8936.1000e114
Baulcombe D (2004) RNA silencing in plants. Nature 431:356–363. https://doi.org/10.1038/nature02874
Bennetzen JL, Schmutz J, Wang H, Percifield R, Hawkins J, Pontaroli AC, Estep M, Feng L, Vaughn JN, Grimwood J, Jenkins J (2012) Reference genome sequence of the model plant Setaria. Nat Biotechnol 30(6):555–561
Bilder RM, Sabb FW, Cannon TD, London ED, Jentsch JD, Parker DS, Poldrack RA, Evans C, Freimer NB (2009) Phenomics: the systematic study of phenotypes on a genome-wide scale. Neuroscience 164(1):30–42
Bonthala VS, Muthamilarasan M, Misra G, Prasad M (2013) FmMDb: a versatile database of foxtail millet markers for millets and bioenergy grasses research. PLoS One 8(8):1–7. https://doi.org/10.1371/journal.pone.0071418
Bonthala VS, Muthamilarasan M, Roy R, Prasad M (2014) FmTFDb: a foxtail millet transcription factors database for expediting functional genomics in millets. Mol Biol Rep 41(10):6343–6348
Canovas F, Dumas-Gaudot E, Recorbet G, Jorrin J, Mock H-P, Rossignol M (2004) Plant proteome analysis. Proteomics 4:285–298
Chen Y, Liu B, Zhao Y, Yu W, Si W (2021) Whole-genome duplication and purifying selection contributes to the functional redundancy of auxin response factor (ARF) genes in foxtail millet (Setaria italica L.). Int J Genomics 2021
Chinnusamy V, Schumaker K, Zhu JK (2004) Molecular genetics perspectives on cross-talk and specificity in abiotic stress signalling in plants. J Exp Bot 55:225–236
Cruz JA, Savage LJ, Zegarac R, Hall CC, Satoh-Cruz M, Davis GA, Kovac WK, Chen J, Kramer DM (2016) Dynamic environmental photosynthetic imaging reveals emergent phenotypes article dynamic environmental photosynthetic imaging reveals emergent phenotypes. Cell Systems 2(6):365–377. https://doi.org/10.1016/j.cels.2016.06.001
da Cunha Valença D, de Moura SM, Travassos-Lins J, Alves-Ferreira M, Medici LO, Ortiz-Silva B, Macrae A, Reinert F (2020) Physiological and molecular responses of Setaria viridis to osmotic stress. Plant Physiol Biochem 155:114–125
Deshmukh R, Sonah H, Patil G, Chen W, Prince S, Mutava R, Vuong T, Valliyodan B, Nguyen HT (2014) Integrating omic approaches for abiotic stress tolerance in soybean. Front Plant Sci 5:1–12
Doust AN, Kellogg EA, Devos KM, Bennetzen JL (2009) Foxtail millet: a sequence-driven grass model system. Plant Physiol 149(1):137–141. https://doi.org/10.1104/pp.108.129627
Fahlgren N, Feldman M, Gehan MA, Wilson MS, Shyu C, Bryant DW, Hill ST, McEntee CJ, Warnasooriya SN, Kumar I, Ficor T (2015) A versatile phenotyping system and analytics platform reveals diverse temporal responses to water availability in Setaria. Mol Plant 8(10):1520–1535
Feng ZJ, Xu ZS, Sun J, Li LC, Chen M, Yang GX, He GY, Ma YZ (2016) Investigation of the asr family in foxtail millet and the role of ASR1 in drought/oxidative stress tolerance. Plant Cell Rep 35(1):115–128. https://doi.org/10.1007/s00299-015-1873-y
Furbank RT, Tester M (2011) Phenomics—technologies to relieve the phenotyping bottleneck. Trends Plant Sci 16:635–644
Goff SA, Vaughn M, McKay S, Lyons E, Stapleton AE, Gessler D, Matasci N, Wang L, Hanlon M, Lenards A, Muir A (2011) The iPlant collaborative : cyber infrastructure for plant biology. Front Plant Sci 2:1–16
Gupta S, Kumari K, Das J, Lata C, Puranik S et al (2011) Development and utilization of novel intron length polymorphic markers in foxtail millet (Setaria italica (L.) P. Beauv.). Genome 54:586–602
Gupta S, Kumari K, Muthamilarasan M, Parida SK, Prasad M (2014) Population structure and association mapping of yield contributing agronomic traits in foxtail millet. Plant Cell Rep 33(6):881–893
Gupta S, Kumari K, Sahu PP, Vidapu S, Prasad M (2012) Sequence-based novel genomic microsatellite markers for robust genotyping purposes in foxtail millet [Setaria italica (L.) P. Beauv]. Plant Cell Rep 31:323–337
Gupta S, Kumari K, Sahu PP, Vidapu S, Prasad M (2013) Sequence-based novel genomic microsatellite markers for robust genotyping purposes in foxtail millet [Setaria italica (L.) P. Beauv]. Plant Cell Rep 31:323–337
Gygi SP, Rochon Y, Franza BR, Aebersold R (1999) Correlation between protein and mRNA abundance in yeast. Mol Cell Biol 19:1720–1730
Hao X, Jin Z, Wang Z, Qin W, Pei Y (2020) Hydrogen sulfide mediates DNA methylation to enhance osmotic stress tolerance in Setaria italica L. Plant Soil 453(1):355–370
Heng L, Chih C, Song C, Chang K (2011) Development of simple sequence repeats (SSR) markers in Setaria italica (Poaceae) and cross-amplification in related species. Int J Mol Sci 12:7835–7845
Huangfu Y, Pan J, Li Z, Wang Q, Mastouri F, Li Y, Yang S, Liu M, Dai S, Liu W (2021) Genome-wide identification of PTI1 family in Setaria Italica and salinity-responsive functional analysis of SiPTI1–5. BMC Plant Biol 21(1):1–17
Jaiswal V, Gupta S, Gahlaut V, Muthamilarasan M, Bandyopadhyay T, Ramchiary N, Prasad M (2019) Genome-wide association study of major agronomic traits in foxtail millet (Setaria italica L.) using DdRAD sequencing. Sci Rep 9(1):1–11
Jayaraman A, Puranik S, Rai NK, Vidapu S, Sahu PP, Lata C, Prasad M (2008) cDNA-AFLP analysis reveals differential gene expression in response to salt stress in foxtail millet (Setariaitalica L.). Mol Biotechnol 40:241–251
Jia G, Huang X, Zhi H, Zhao Y, Zhao Q, Li W, Chai Y, Yang L, Liu K, Lu H, Zhu C (2013) A haplotype map of genomic variations and genome-wide association studies of agronomic traits in foxtail millet (Setaria italica). Nat Genet 45(8):957–961
Jia X, Zhang Z, Liu Y, Zhang C, Shi Y, Song Y, Wang T, Li Y (2009) Development and genetic mapping of SSR markers in foxtail millet [Setaria italica (L.) P. Beauv.]. Theor Appl Genet 118:821–829
Jia XP, Shi YS, Song YC, Wang GY, Wang TY, Li Y (2007) Development of EST-SSR in foxtail millet (Setaria italica). Genet Resour Crop Evol 54:233–236. https://doi.org/10.1007/s10722-006-9139-8
Khan Y, Yadav A, Suresh VB, Muthamilarasan M, Yadav CB, Prasad M (2014) Comprehensive genome-wide identification and expression profiling of foxtail millet [Setaria italica (L.)] miRNAs in response to abiotic stress and development of miRNA database. Plant Cell Tiss Organ. https://doi.org/10.1007/s11240-014-0480-x
Kim JK, Bamba T, Harada K, Fukusaki E, Kobayashi A (2007) Time-course metabolic profiling in Arabidopsis thaliana cell cultures after salt stress treatment. J Exp Bot 58:415–424
Kim JK, Park S-Y, Yeo Y, Cho HS, Kim YB, Bae H, Park CH, LeeJ-H and Park SU (2013) Metabolic profiling of millet(Panicum miliaceum) using gas chromatography–time-offlightmass spectrometry (GC-TOFMS) for qualityassessment Plant Omics J 6: 73–78
Kumar S, Trivedi PK (2018) Glutathione S-transferases: role in combating abiotic stresses including arsenic detoxification in plants. Front Plant Sci 9:751
Kumari K, Muthamilarasan M, Misra G, Gupta S, Subramanian A et al (2013) Development of eSSR-markers in Setaria italica and their applicability in studying genetic diversity, cross-transferability and comparative mapping in millet and non-millet species. PLoS One 8:e67742
Kvilekval K, Fedorov D, Obara B, Singh A, Manjunath BS (2010) Bisque: a platform for bioimage analysis and management. Bioinformatics 26:544–552
Lata C (2015) Advances in omics for enhancing abiotic stress tolerance in millets. Proc Indian Nat Sci Acad 81(2):397–417
Lata C, Bhutty S, Bahadur RP, Majee M, Prasad M (2011a) Association of an SNP in a novel DREB2-like gene SiDREB2 with stress tolerance in foxtail millet [Setaria italica (L.)]. J Exp Bot 62(10):3387–3401
Lata C, Bhutty S, Bahadur RP, Majee M, Prasad M (2011b) Association of an SNP in a novel DREB2-like gene SiDREB2 with stress tolerance in foxtail millet [Setaria italica (L.)]. J Exp Bot 62:3387–3401
Lata C, Mishra AK, Muthamilarasan M, Bonthala VS, Khan Y, Prasad M (2014) Genome-wide investigation and expression profiling of AP2/ERF transcription factor superfamily in foxtail millet (Setaria italica L.). PLoS One 9:e113092
Lata C, Prasad M (2011) Role of DREBs in regulation of abiotic stress responses in plants. J Exp Bot 62:4731–4748
Lata C, Prasad M (2013) Setaria genome sequencing: an overview. J Plant Biochem Biotechnol 22:257–260. https://doi.org/10.1007/s13562-013-0216-8
Lata C, Sahu PP, Prasad M (2010) Comparative transcriptome analysis of differentially expressed genes in foxtail millet (Setaria italica L.) during dehydration stress. Biochem Biophys Res Commun 393:720–727
Le Thierry d’Ennequin M, Panaud O, Toupance B, Sarr A (2000) Assessment of genetic relationships between Setaria italica and its wild relative S. viridis using AFLP markers. Theor Appl Genet 100:1061–1066
Li C, Yue J, Wu X, Xu C, Yu J (2014) An ABA-responsive DRE-binding protein gene from Setaria italica, SiARDP, the target gene of SiAREB, plays a critical role under drought stress. J Exp Bot 65:5415–5427
Li J, Dong Y, Li C, Pan Y, Yu J (2017) SiASR4, the target gene of SiARDP from Setaria italica, improves abiotic stress adaption in plants. Front Plant Sci 7:2053
Li J, Li X, Yang Q, Luo Y, Gong X, Zhang W, Hu Y, Yang T, Dong K, Feng B (2019) Proteomic changes in the grains of foxtail millet (Setaria italica (L.) Beau) under drought stress. Span J Agric Res 17(2):802
Li P, Brutnell TP (2011) Setaria viridis and Setaria italica, model genetic systems for panicoid grasses. J Exp Bot 62:3031–3037. https://doi.org/10.1093/jxb/err096. PubMed: 21459768
Li S, Dong X, Fan G, Yang Q, Shi J, Wei W, Zhao F, Li N, Wang X, Wang F, Feng X (2018) Comprehensive profiling and inheritance patterns of metabolites in foxtail millet. Front Plant Sci 9:1716
Liang K, Liang S, Zhu H (2020) Comparative proteomics analysis of the effect of selenium treatment on the quality of foxtail millet. Lwt 131:109691
Liu W, Li RJ, Han TT, Cai W, Fu ZW, Lu YT (2015) Salt stress reduces root meristem size by nitric oxide mediated modulation of auxin accumulation and signaling in Arabidopsis. Plant Physiol 168(1):343–356
Liu Y, Feng X, Xu Y, Yu J, Ao G, Peng Z et al (2009) Overexpression of millet ZIP-like gene (SiPf40) affects lateral bud outgrowth in tobacco and millet. Plant Physiol Biochem 47:1051–1060
Loqué D, von Wirén N (2004) Regulatory levels for the transport of ammonium in plant roots. J Exp Bot 55:1293–1305
Mauro-Herrera M, Wang X, Barbier H, Brutnell TP, Devos KM, Doust AN (2013) Genetic control and comparative genomic analysis of flowering time in Setaria (Poaceae). G3 (3):283–295
Meng X, Liang Z, Dai X, Zhang Y, Mahboub S, Ngu DW, Roston RL, Schnable JC (2021) Predicting transcriptional responses to cold stress across plant species. Proc Natl Acad Sci U S A 118(10):1–9
Mishra AK, Muthamilarasan M, Khan Y, Parida SK, Prasad M (2014) Genome-wide investigation and expression analyses of WD40 protein family in the model plant foxtail millet (Setaria italica L.). PLoS One. https://doi.org/10.1371/journal.pone.0086852
Muthamilarasan M, Bonthala VS, Mishra AK, Khandelwal R, Khan Y, Roy R et al (2014b) C2H2type of zinc finger transcription factors in foxtail millet define response to abiotic stresses. Funct Integr Genomics 14:531–543
Muthamilarasan M, Khandelwal R, Yadav CB, Bonthala VS, Khan Y, Prasad M (2014c) Identification and molecular characterization of MYB transcription factor superfamily in C4 model plant foxtail millet (Setaria italica L.). PLoS One 9:e109920
Muthamilarasan M, Prasad M (2015) Advances in Setaria genomics for genetic improvement of cereals and bioenergy grasses. Theor Appl Genet 128(1):1–14
Muthamilarasan M, Singh RK, Suresh BV, Rana S, Dulani P, Prasad M (2020) Genomic dissection and expression analysis of stress-responsive genes in C4 panicoid models, Setaria italica and Setaria viridis. J Biotechnol 318:57–67
Muthamilarasan ME, Venkata Suresh B, Pandey GA, Kumari KA, Parida SK, Prasad MA (2014a) Development of 5123 intron-length polymorphic markers for large-scale genotyping applications in foxtail millet. DNA Res 21(1):41–52
Nadeem F, Ahmad Z, Ul Hassan M, Wang R, Diao X, Li X (2020) Adaptation of foxtail millet (Setaria italica L.) to abiotic stresses: a special perspective of responses to nitrogen and phosphate limitations. Front Plant Sci 11:187
Nadeem F, Ahmad Z, Wang R, Han J, Shen Q, Chang F, Diao X, Zhang F, Li X (2018) Foxtail millet [Setaria italica (L.) Beauv.] grown under low nitrogen shows a smaller root system, enhanced biomass accumulation, and nitrate transporter expression. Front Plant Sci 9(February):1–12
Ni X, Xia Q, Zhang H, Cheng S, Li H, Fan G, Guo T, Huang P, Xiang H, Chen Q, Li N (2017) Updated foxtail millet genome assembly and gene mapping of nine key agronomic traits by resequencing a RIL population. Giga Sci 6(2):1–8
Pan J, Li Z, Dai S, Ding H, Wang Q, Li X, Ding G, Wang P, Guan Y, Liu W (2020) Integrative analyses of transcriptomics and metabolomics upon seed germination of foxtail millet in response to salinity. Sci Rep 10(1):1–16
Pan J, Li Z, Wang Q, Garrell AK, Liu M, Guan Y, Zhou W, Liu W (2018) Comparative proteomic investigation of drought responses in foxtail millet. BMC Plant Biol 18(1):1–19
Pandey G, Misra G, Kumari K, Gupta S, Parida SK et al (2013) Genome-wide development and use of microsatellite markers for large-scale genotyping applications in foxtail millet [Setaria italica (L.)]. DNA Res 20:197–207
Passot S, Gnacko F, Moukouanga D, Lucas M, Guyomarc’h S, Ortega BM, Atkinson JA, Belko MN, Bennett MJ, Gantet P, Wells DM (2016) Characterization of pearl millet root architecture and anatomy reveals three types of lateral roots. Front Plant Sci 7:1–11
Pegler JL, Nguyen DQ, Grof CP, Eamens AL (2020) Profiling of the salt stress responsive microRNA landscape of C4 genetic model species Setaria viridis (L.) Beauv. Agronomy 10(6):837
Prakash PT, Banan D, Paul RE, Feldman MJ, Xie D, Freyfogle L, Baxter I, Leakey AD (2021) Correlation and co-localization of QTL for stomatal density, canopy temperature, and productivity with and without drought stress in Setaria. J Exp Bot 72(13):5024–5037
Puranik S, Jha S, Srivastava PS, Sreenivasulu N, Prasad M (2011) Comparative transcriptome analysis of contrasting foxtail millet cultivars in response to short-term salinity stress. J Plant Physiol 168:280–287
Puranik S, Sahu PP, Mandal SN, Parida SK, Prasad M (2013) Comprehensive genome-wide survey, genomic constitution, and expression profiling of the NAC transcription factor family in foxtail millet (Setaria italica L.). PLoS One 8:e64594
Qi X, Xie S, Liu Y, Yi F, Yu J (2013) Genome-wide annotation of genes and noncoding RNAs of foxtail millet in response to simulated drought stress by deep sequencing. Plant Mol Biol 83:459–473
Qie L, Jia G, Zhang W, Schnable J, Shang Z, Li W, Liu B, Li M, Chai Y, Zhi H, Diao X (2014) Mapping of quantitative trait locus (QTLs) that contribute to germination and early seedling drought tolerance in the interspecific cross Setaria italica x Setaria viridis. PLoS One 9(7):3–10. https://doi.org/10.1371/journal.pone.0101868
Qin L, Chen E, Li F, Yu X, Liu Z, Yang Y, Wang R, Zhang H, Wang H, Liu B, Guan YA (2020) Genome-wide gene expression profiles analysis reveal novel insights into drought stress in foxtail millet (Setaria italica L.). Int J Mol Sci 21(22):1–21
Rahaman M, Chen D, Gillani Z, Klukas C, Chen M (2015) Advanced phenotyping and phenotype data analysis for the study of plant growth and development. Front Plant Sci 6:1–15. https://doi.org/10.3389/fpls.2015.00619
Ramanna H, Ding XS, Nelson RS (2013) Rationale for developing new virus vectors to analyze gene function in grasses through virus-induced gene silencing. Methods Mol Biol 975:15–32
Reddy DS, Bhatnagar-Mathur P, Vadez V, Sharma KK (2012) Grain legumes (soybean, chickpea, and Peanut): omics approaches to enhance abiotic stress tolerance. In: Tuteja N, Gill SS, Tiburcio AF, Tuteja R (eds) Improving crop resistance to abiotic stress. Wiley-VCH Verlag GmbH & Co.KGaA. https://doi.org/10.1002/9783527632930.ch39
Roch GV, Maharajan T, Krishna TA, Ignacimuthu S, Ceasar SA (2020) Expression of PHT1 family transporter genes contributes for low phosphate stress tolerance in foxtail millet (Setaria italica) genotypes. Planta 252(6):1–9
Sebastian J, Yee MC, Viana WG, Rellán-Álvarez R, Feldman M, Priest HD, Trontin C, Lee T, Jiang H, Baxter I, Mockler TC (2016) Grasses suppress shoot-borne roots to conserve water during drought. PNAS USA 113(31):8861–8866. https://doi.org/10.1073/pnas.1604021113
Shi W, Cheng J, Wen X, Wang J, Shi G, Yao J, Hou L, Sun Q, Xiang P, Yuan X, Dong S (2018) Transcriptomic studies reveal a key metabolic pathway contributing to a well-maintained photosynthetic system under drought stress in foxtail millet (Setaria italica L.). PeerJ 6:e4752
Shkolnik D, Finkler A, Pasmanik-Chor M, Fromm H (2019) Calmodulin-binding transcription activator 6: a key regulator of Na+ homeostasis during germination. Plant Physiol 180:1101–1118
Singh A, Ganapathysubramanian B, Singh AK, Sarkar S (2016) Machine learning for high-throughput stress phenotyping in plants. Trends Plant Sci 21:110–124
Singh RK, Muthamilarasan M, Prasad M (2017) Foxtail millet: An introduction. In: Prasad M (ed) The foxtail millet genome. Compendium of plant genomes. Springer, Cham, pp 1–9. https://doi.org/10.1007/978-3-319-65617-5_1
Sood P, Prasad M (2017) Genetic transformation of Setaria: a new perspective. In: Prasad M (ed) The foxtail millet genome, compendium of plant genomes. Springer, Berlin, pp 105–121
Sood P, Singh RK, Prasad M (2020) An efficient agrobacterium-mediated genetic transformation method for foxtail millet (Setaria italica L.). Plant Cell Rep 39(4):511–525
Sreenivasulu N, Miranda M, Prakash HS, Wobus U, Weschke W (2004) Transcriptome changes in foxtail millet genotypes at high salinity: identification and characterization of aPHGPX gene specifically upregulated by NaCl in a salt tolerant line. J Plant Physiol 161:467–477
Tang S, Li L, Wang Y, Chen Q, Zhang W, Jia G, Zhi H, Zhao B, Diao X (2017) Genotype-specific physiological and transcriptomic responses to drought stress in Setaria italica (an emerging model for panicoideae grasses). Sci Rep 7(1):1–15. https://doi.org/10.1038/s41598-017-08854-6
Veeranagamallaiah G, Jyothsnakumari G, Thippeswamy M, Reddy PC, Surabhi GK, Sriranganayakulu G, Mahesh Y, Rajasekhar B, Madhurarekha C, Sudhakar C (2008) Proteomic analysis of salt stress responses in foxtail millet (Setaria italica L. cv. Prasad) seedlings. Plant Sci 175(5):631–641
Vetriventhan M, Upadhyaya HD, Anandakumar CR, Senthilvel S, Varshney RK, Parzies HK (2014) Population structure and linkage disequilibrium of ICRISAT foxtail millet (Setaria italica (L.) P. Beauv.) core collection. Euphytica 196:423–435. https://doi.org/10.1007/s10681-013-1044-6
Wang J, Wang Z, Du X, Yang H, Han F, Han Y, Yuan F, Zhang L, Peng S, Guo E (2017) A high-density genetic map and qtl analysis of agronomic traits in foxtail millet [Setaria italica (L.) P. Beauv.] using RAD-Seq. PLoS One 12(6):1–15
Wang M, Ellsworth PZ, Zhou J, Cousins AB, Sankaran S (2016) Evaluation of water-use efficiency in foxtail millet (Setaria italica) using visible-near infrared and thermal spectral sensing techniques. Talanta 152:531–539
Wang M, Li P, Li C, Pan Y, Jiang X, Zhu D, Zhao Q, Yu J (2014) SiLEA14, a novel atypical LEA protein, confers abiotic stress resistance in foxtail millet. BMC Plant Biol 14(1):1–16
Wang ZM, Devos KM, Liu CJ, Wang RQ, Gale MD (1998) Construction of RFLP-based maps of foxtail millet, Setaria italica (L.) P. Beauv. Theor Appl Genet 96:31–36
Xu B, Gao X, Dong K, Li X, Yang P, Yang T, Feng B (2020) Grain protein content comparison and proteomic analysis of foxtail millet (Setaria italica L.) seed response to different drought stress levels. Acta Physiol Plant 42(2):1–1
Xu J, Li Y, Ma X, Ding J, Wang K, Wang S, Tian Y, Zhang H, Zhu XG (2013) Whole transcriptome analysis using next-generation sequencing of model species Setaria viridis to support C4 photosynthesis research. Plant Mol Biol 83:77–87
Yadav CB, Bonthala VS, Muthamilarasan M, Pandey G, Khan Y, Prasad M (2014a) Genome-wide development of transposable elements-based markers in foxtail millet and construction of an integrated database. DNA Res. https://doi.org/10.1093/dnares/dsu039
Yadav CB, Muthamilarasan M, Pandey G, Khan Y, Prasad M (2014b) Development of novel microRNA-based genetic markers in foxtail millet for genotyping applications in related grass species. Mol Breed 34:2219–2224
Yadav CB, Muthamilarasan M, Pandey G, Prasad M (2015) Identification, characterization and expression profiling of dicer-like, argonaute and RNA-dependent RNA polymerase gene families in foxtail millet. Plant Mol Biol Rep 33:43–55
Yan J, Yang L, Liu Y, Zhao Y, Han T, Miao X, Zhang A (2021) Calcineurin B-like protein 5 (SiCBL5) in Setaria italica enhances salt tolerance by regulating Na+ homeostasis. Crop J 15. https://doi.org/10.1016/j.cj.2021.06.006
Yin H, Yu Q, Dai Y, An L and Li W (2014). Transcript profiling of foxtail millet seedlings under drought condition and discovery of a new drought-tolerant gene. In: 1stInternational Setaria Genetics Conference, pp 24
Zhang G, Liu X, Quan Z, Cheng S, Xu X et al (2012) Genome sequence of foxtail millet (Setaria italica) provides insights into grass evolution and biofuel potential. Nat Biotechnol 30:549–554
Zhang J, Liu T, Fu J, Zhu Y, Jia J, Zheng J, Zhao Y, Zhang Y, Wang G (2007) Construction and application of EST library from Setaria italica in response to dehydration stress. Genomics 90:121–131
Zhang L, Ren Y, Wan Y, Xu Q, Yang G, Zhang S, Huang J, Yan K, Zheng C, Wu C (2021) SiCEP3, a C-terminally encoded peptide from Setaria italica, promotes ABA import and signaling. J Exp Bot 72(18):6260–6273
Zhang S, Tang C, Zhao Q, Li J, Yang L, Qie L, Fan X, Li L, Zhang N, Zhao M, Liu X (2014) Development of highly polymorphic simple sequence repeat markers using genome wide microsatellite variant analysis in foxtail millet [Setaria italica (L.) P. Beauv]. BMC Genomics 15:78
Zhu C, Ming C, Zhao-shi X, Li L-C, Xue-ping C, Ma Y-z (2014) Characteristics and expression patterns of the aldehyde dehydrogenase (ALDH) gene superfamily of foxtail millet (Setaria italica L.). PLoS One 9:e101136
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2022 The Author(s), under exclusive license to Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Bhinda, M.S., Sanadya, S.K., Kumari, A., Kant, L., Debnath, A. (2022). Omics for Abiotic Stress Tolerance in Foxtail Millet. In: Pudake, R.N., Solanke, A.U., Sevanthi, A.M., Rajendrakumar, P. (eds) Omics of Climate Resilient Small Millets. Springer, Singapore. https://doi.org/10.1007/978-981-19-3907-5_2
Download citation
DOI: https://doi.org/10.1007/978-981-19-3907-5_2
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-19-3906-8
Online ISBN: 978-981-19-3907-5
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)