
Abstract
The placenta is the primary link between the mother and the fetus. It regulates the exchange of nutrients, oxygen, and waste between the mother and the fetus. It also protects the fetus from maternal insults such as infections and exposure to toxicants, thereby minimizing any teratogenicity. Therefore, ensuring placental health is essential for a successful pregnancy. However, the development, differentiation, functions, and dysfunctions of the human placenta are difficult to study in vivo due to ethical constraints. On the other hand, the differences in the morphologies and the functions of the placenta between different species make the data obtained from animal models less reliable. Availability of various human placental cell lines as well as recent advances in microfluidics and bioprinting technologies has led to the evolution of the placenta-on-chip models. These models can effectively simulate the anatomical and physiological characteristics of the human placenta by successfully mimicking the in vivo environment inside a microfluidic chip. By incorporating relevant placental cell types into the device design and controlling the microenvironment, microfluidics-based in vitro models hold clues to the pathogenesis of placental dysfunction and facilitate drug testing across the maternal-fetal interface. In this chapter, we highlight some existing microfluidic platforms that have attempted to simulate various aspects of placental biology. In addition, we review the applications of these models and discuss the limitations in the context of placental physiology and pathophysiology.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

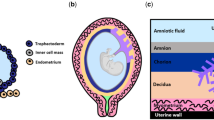
16.1 Introduction
Disorders of pregnancy due to placental defects are almost unique to the human species. The placenta-related pregnancy disorders include miscarriage, gestational diabetes, preterm births, and pre-eclampsia. Accounting for more than a third of human pregnancies, these disorders are a cause of significant morbidity and mortality both to the mother and the fetus. Thus, an understanding of the placental functions in normal conditions and in pathological pregnancies is crucial for developing management strategies for these disorders. Pregnant women and hence their fetuses are treated with a significant number of prescription and non-prescription medications. As the mother undergoes significant adaptations during pregnancy, the pharmacokinetics and pharmacodynamics of drugs can change, resulting in changes in effectiveness of the therapies. Unfortunately, pregnant women are excluded from most drug studies as there are concerns regarding fetal risk and hence the data on placental-mediated drug interactions are sparse. Thus, studying the placenta and investigating placental dysfunctions are of interest to clinicians, developmental and reproductive biologists, and toxicologists. Therefore, versatile in vitro models that simulate human placenta both anatomically and physiologically are highly desirable.
Advancements in the availability of human placental cell lines, microfluidics, and bioprinting have contributed to the evolution of placenta-on-chip models. Over the conventional two-dimensional (2D) systems, the three-dimensional (3D) cell culture systems, the placenta-on-chip models, are multicellular in nature recapitulating the spatial organization observed in vivo, reiterating some aspect of the tissue functions. The microfluidic systems provide mechanical cues like shear stress, tension and compression which has an important role in cell/tissue physiology. These bioengineered in vitro models of human placenta promise to offer precise control over the microenvironment allowing recapitulation of specific aspects of human placentation in health and disease.
Herein, we discuss (1) development and the structure of human placenta; (2) current placenta-on-chip models; (3) their applications and limitations in studying placental physiology and disorders of pregnancy; and (4) present limitations and critical aspects for future considerations.
16.2 Development and Structure of Human Placenta
Fertilization in humans is characterized by fusion of sperm and egg and formation of a zygote. This zygote then undergoes multiple rounds of cell division and forms a blastocyst. The blastocyst stage comprises Inner Cell Mass (ICM), Trophectoderm (TE), and the blastocoel. The inner cell mass develops into the embryo proper, while the trophectoderm cells give rise to the placenta. Before the formation of placenta, the blastocyst implants itself into the maternal uterus. At the time it enters the endometrial lumen, the blastocyst hatches itself from the zona coat and is closely apposed to the wall of the endometrium termed as the luminal epithelium. This is followed by the attachment stage where the contact between trophectoderm and the luminal epithelium of the uterus is established. The next step is embryo invasion wherein the trophectoderm invades into the endometrium to successfully embed the embryo into the uterus. The trophectoderm invasion marks the beginning of placenta development [1]. The sequelae of events involved in formation of human placenta are summarized in Fig. 16.1 and described below. The reader is requested to go through the following papers for a detailed description of placental development [2,3,4,5].
-
1.
Primary syncytium formation: The trophectoderm after initial invasion gets completely embedded inside the endometrium. By this stage, the embryo is bathed by endometrial secretions. The trophectoderm cells proliferate and expand in numbers and differentiate into cytotrophoblasts. The cytotrophoblasts, by a fusion event, lose their cell boundaries and form multinucleated syncytiotrophoblast surrounding the embryo forming the primary syncytium. This primary syncytium further invades and erodes the decidua.
-
2.
Formation of primary villi: While the outer syncytial layer is in direct contact with the maternal blood, the inner layer of the cytotrophoblast cells completely surrounds the conceptus to form the cytotrophoblast shell. At this stage, the cytotrophoblast proliferates and solid outgrowths of cytotrophoblast cells protrude into the syncytiotrophoblast layer giving rise to the primary villi. In a cross section, the primary villus has a central core surrounded by the layer of syncytiotrophoblasts.
-
3.
Development of the secondary and tertiary villi: Towards the embryonic side, the extraembryonic mesoderm is differentiated and layers above the cytotrophoblast cells. The cells of the extraembryonic mesoderm extend and penetrate into the primary villi, resulting in the inner stromal core, the cytotrophoblast layer, and the outer syncytiotrophoblast layer. The fetal blood vessels that originate from the extraembryonic mesoderm then penetrate into the primary villus making them vascular. By the process of constant proliferation and invasion, the primary villi branches into secondary villi and tertiary villi resulting in the placental tree. The endothelium in the villi never penetrates into the trophoblast lacunae or anastomose with the material blood vessels. Numerous “daughter” villi arise out of the tertiary villi. These remain either free and project into the intervillous space.
-
4.
The floating villi: The villi that bathe in maternal blood are referred to as the floating villi, and through the syncytiotrophoblast layer, there is exchange of nutrients and exclusion of waste products between the mother and fetus. These floating villi act as a selectively permeable structure and are primarily responsible for the barrier function of the placenta. The gases, nutrients, and waste products that diffuse through the maternal and fetal blood must pass through a total of four layers. From outside to inside, these are (1) Syncytiotrophoblast, (2) Cytotrophoblast, (3) Loose connective tissue that surrounds the endothelium, (4) Capillary endothelium of the villus. These four elements together form the placental barrier of the first trimester which continues to be so till 16 weeks (fourth month of human pregnancy). After 16 weeks, cytotrophoblast in the tertiary villi disappears and the villi branches further and becomes very thin. This results in reduction of the intervillous space and the fetal vessels become smaller and come in direct contact with the outermost syncytiotrophoblast layer. By the end of the second trimester, the barrier is now a bilayered structure with the outer syncytiotrophoblast layer and the fetal endothelium. After the mid second trimester, the gases, nutrients, and waste products that diffuse through the maternal and fetal blood must pass through these two layers to enter the fetal circulation.
-
5.
Anchoring villi: These are villi that attach to the maternal decidua and are determinant of the depth of placentation. The major cell type of the anchoring villi are the extravillous trophoblasts. The cytotrophoblast shell at the proximal end of the growing villi facing the maternal decidua proliferates and forms a column of cells termed as the cytotrophoblast cell column. These column trophoblasts anchor on to the maternal decidua and initially plug the maternal arteries (Fig. 16.2). The cells at the tip of column trophoblast cells that anchor on the decidua leave the cytotrophoblast column and gain an invasive phenotype to form the extravillous trophoblast (EVTs) cells. These EVTs further differentiate into interstitial EVTs (iEVTs) and the endovascular EVTs (eEVTs). The iEVTs reside in the endometrial bed, while the eEVTs remodel the spiral arteries.
-
6.
Spiral artery remodelling: Spiral artery remodelling is the process where the endothelial cells of the artery are replaced by the eEVTs to give rise to a vessel that is capable of high conductance under physiological pressure. The eEVTs of the column trophoblasts invade the maternal arteries and displace the maternal endothelium and the surrounding smooth muscle cells. This occurs well under the upper two thirds of the decidua basalis almost until the myometrium.

Schematic process of formation of the Human Placenta

Anchoring villus, column trophoblast, extravillus trophoblasts and the process of spiral artery remodeling. At the tip of the anchoring villi, the syncytiotrophoblast anchor and form the cell columns. These cells are known as cytotrophoblast column cells. These cytotrophoblast cells initially plug the lumen of the spiral arteries and differentiate to form the Extravillous trophoblasts (EVTs). The EVTs are of two types, interstitial EVTS (iEVT) and endovascualr EVTs (eEVTs). The invasive EVTs invade both into the maternal decidua and the lumen of the spiral arteries in the first trimester of pregnancy. The eEVTs remodel the spiral arteries by replacing endothelial lining to accommodate a high flow volume of maternal blood without excessive velocity. Not all the arteries undergo the process of remodeling some remain in natural state as well. (Figure adapted with permission from Saghian et al. © (2019) The Royal Society (UK))
16.3 Structure Function Relationship of the Human Placenta in Physiology and Pathophysiology
In this section, we will describe the functions ascribed to the main cell types in the placenta and their roles in pregnancy.
-
Cytotrophoblasts and syncytiotrophoblasts: The placental villi are made of two cell populations, viz., the undifferentiated cytotrophoblasts and fully differentiated syncytiotrophoblasts.
-
Syncytiotrophoblast cells: This is the outermost layer of all villi and covers all placental villi as one single layer. Beneath the syncytiotrophoblasts are the cytotrophoblasts which are the precursors of syncytiotrophoblasts. Cytotrophoblasts continually differentiate into syncytiotrophoblasts during villous formation and development. The syncytiotrophoblast fetal cells are directly in contact with maternal blood and the cell layer acts as a primary barrier between the mother and fetus and is the site with the highest metabolic and endocrine activity. This layer plays a key function in the materno-fetal (and feto-maternal) transfer of biomolecules, drugs, and infectious agents. It is an active site for catabolism and synthesis of proteins and lipids. It is also a site of hormone synthesis of β-hCG (beta-subunit of human chorionic gonadotropin) and progesterone which are essential to maintain pregnancy [6]. The syncytiotrophoblast is the main site of exchange for drugs and a determinant of mother to child transmission of pathogens. In general, hydrophobic low molecular weight xenobiotics (<600 Da) can usually transfer across the placenta via passive diffusion, while hydrophilic and/or high molecular weight compounds (1000+ Da) cross very poorly [7]. The syncytiotrophoblast layer also protects the fetuses from maternal infections. However, some viruses (like Zika, SARS-CoV-2, CMV, EBV) can infect the syncytiotrophoblasts and act as reservoirs of the viruses [8,9,10,11]. In some instances, the viruses can also undergo lytic cycles in the placental cells and release the virions into the feto-maternal infection causing mother to child transmission. Beyond viruses, pathogens like malarial parasites have specific affinity to the placenta as they can reside into the syncytiotrophoblasts causing placental malaria which has life-threatening consequences [12, 13].
-
Extravillous trophoblasts: The extravillous trophoblasts are of two types, viz., the endovascular EVTs (eEVTs) and the interstitial EVTs (iEVTs). The extravillous trophoblast cells that infiltrate arterial walls of the spiral arteries make up the eEVTs. The EVTs that migrate into the maternal decidua are iEVTs. The migration of EVTs is a crucial step in human placentation. During pregnancy, the maternal blood vessels are extensively remodelled up to the upper third of the myometrium. To achieve this, the cells of the column trophoblasts migrate into the decidua. These iCTBs surround the maternal spiral arteries and initiate the remodelling process. Once the iCTBs align the maternal spiral arteries, they differentiate into eCTBs by adopting a vascular adhesion and interdigitate into the endothelial layer, and eventually replace the maternal endothelial cells completely. This allows the narrow spiral with relatively high resistance to be transformed into highly dilated, low-resistance channels (Fig. 16.2). These remodelled vessels alter the blood flow dynamics at an appropriate speed to ensure maximal perfusion and prevent damage to the villi.
Along with spiral remodelling, the eCTBs also form trophoblast plugs in early pregnancy to obstruct spiral arteries and prevent blood flow. As a consequence, the placenta exists in a low oxygen environment for the majority of the first trimester, and this is thought to be key to promote placental development, vasculogenesis, and angiogenesis. The failure of deep endovascular invasion and spiral artery remodelling is observed in a number of pregnancy-related disorders. Premature loosening of the trophoblast plugs is associated with miscarriages; inadequate spiral artery remodelling is observed in pregnancies complicated with pre-eclampsia and Intrauterine Growth Restriction (IUGR) [14].
16.4 Placenta-on-Chip
Since placental functions are crucial for several aspects of pregnancy, research in this direction has taken a centre stage. Attempts have been made to simulate various aspects of placental physiology using in vitro model systems. In the following sections, we have reviewed the existing models of human placenta-on-chip. Table 16.1 lists the cell lines derived from human placenta that are used for developing the placenta-on-chip models.
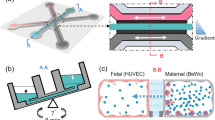
The existing placenta-on-a-chip models can be classified into those that simulate the placental barrier function and those that are used to study trophoblast migration. The placental barrier models can be sub-classified into two design categories: multi-layered vertical design and single-layered planar design (Fig. 16.3).

Classification of the placenta-on-chip (PoC) models
In the multi-layered vertical design, two microfluidic layers containing the microchannel features are aligned and bonded with an extracellular matrix membrane between them, mimicking the placental barrier that separates the maternal circulation in the intervillous space and fetal circulation in the fetal capillaries (Fig. 16.4). In these models, the cells are first introduced into the lower channel and the device is immediately inverted to allow cells to adhere to the bottom surface of the membrane. During this process, the inlets and outlets are blocked to prevent loss of cell suspension by outflow. After the cell adherence is confirmed, cells are introduced into the upper microchannel and allowed to adhere. Once cells are seeded and firmly attached to the membrane surfaces, they are allowed to form confluent monolayers, while media is flowed through the channels at physiologically relevant flow rates.

Vertical multi-layered placenta-on-chip model. (a) The engineered device is composed of two PDMS chambers separated by a vitrified collagen membrane. The upper surface is shown in blue and the lower surface is marked in red. (b) Trophoblasts are co-cultured with endothelial cells in close apposition on the either side of the membrane to form a microengineered placenta-on-a-chip device which mimics the placental barrier function. (The figure is adapted with permission from Lee et al. © (2016) Journal of maternal-fetal and neonatal medicine)
In the single-layered planar design, a single microfluidic layer is patterned to define both the maternal and fetal channels in the same layer and bonded to an unpatterned surface. The maternal and fetal cells are seeded on either side of a separating membrane or matrix and cultured under media perfusion at physiologically relevant flow rates.
16.4.1 Models of Placental Barrier
The first-ever placental barrier model was developed by Lee et al. [15], containing two patterned polydimethylsiloxane (PDMS) layers with a vitrified collagen membrane sandwiched between them (Fig. 16.5). The PDMS layers, with channel features of width 500 μm and height 200 μm, were fabricated using soft lithography techniques. The vitrified collagen membrane was developed using type 1 collagen mixed with distilled water and culture medium. This solution was dispensed over the central region of the lower PDMS microchannel and allowed to gelate, which was then dried overnight to develop into the membrane. The two PDMS layers, with the membrane attached to the lower layer, were plasma treated for bonding to each other. The membrane surface on the fetal and maternal sides were coated with fibronectin and gelatin, respectively, prior to cell seeding to enhance cell adhesion and growth. JEG-3 human trophoblast cells and human umbilical vein endothelial cells (HUVECs) were seeded on the maternal and fetal side of the vitrified collagen membrane, respectively. After cells were firmly attached to the membrane, appropriate growth media were perfused through the channels using a syringe pump at 30 μL/h for 3 days to allow the cells to form confluent monolayers. This model recapitulated the structural aspect of the placental villous tree of the feto-maternal interface (Fig. 16.4). This device was tested for functionality by comparing the efficiency of glucose transport across the barrier on-chip with ex-vivo models. The rate of transfer and the permeability coefficient of glucose were found similar to that of ex-vivo model systems. However, this design used Jeg-3 cells which resemble the EVTs and not the syncytiotrophoblasts. Thus, the results from this study need to be interpreted with caution.

Fabrication of the vertical multi-layered placenta-on-a-chip: (a) The microchannel master is produced using photolithography. (b) Replica molding of PDMS is carried out to define microchannels. The vitrified collagen membrane is incorporated to define the placental barrier. The upper and the lower PDMS slabs are permanently bonded, housing a microchannel covered with the vitrified membrane. (c) Sequential seeding of JEG-3 cells on the maternal side and HUVECs on the fetal side in the multilayered device. (The figure is adapted with permission from Lee et al. © (2016) Journal of maternal-fetal and neonatal medicine)
The above design from Lee et al. 2015 was extended by Blundell et al. who used a semipermeable polycarbonate membrane with 1 μm pores between the maternal and fetal channels [16, 17]. The two PDMS layers containing the microchannels (w = 1 mm, h = 135 μm, l = 1.5 cm) were bonded to the membrane using adhesive PDMS mortar. The channels were treated with fibronectin before seeding the cells. BeWo human trophoblasts and human primary placental villous endothelial cells (HPVECs) were cultured on either side of the membrane with culture media flowing through the channels at 100 μL/h. The device was developed to study the transport of substances from mother to fetus under continuous physiological flow conditions. This model was tested to study the transfer of glyburide (a drug used for treatment of gestational diabetes) across the placental barrier. They were able to show that glyburide transport across the barrier was very low and this finding was similar to that of other ex-vivo models (see below).
Another attempt was made to mimic the placental barrier structure in vitro. Mosavati et al. developed a model with BeWo trophoblast cells and HUVECs cultured on either side of a type 1 collagen-coated polycarbonate membrane with 0.4 μm sized pores [18]. The PDMS layers, with channel features of width 1 mm and height 200 μm, were bonded to the membrane using adhesive PDMS/toluene mortar. The cells were cultured in the microchannels under static conditions for 3 days with media change every day. Cell confluency was achieved at 72 h post cell seeding for HUVEC (86 ± 3%) and BeWo cell (90 ± 2%) layers. Intercellular junction formation for both the cell types was assessed by staining the cells using calcein-stain for HUVEC cells and Cell Tracker orange fluorescence-stain for BeWo cells growing on each surface of the porous membrane. Effects of membrane porosity and flow rate on glucose diffusion across the placental barrier were studied using numerical models. Glucose transfer rates of the device were determined in the absence of cells (93 ± 5.7%), in the presence of single cell type (70 ± 6.1% for HUVECs and 63 ± 4.2% for BeWo cells), and co-culture (35 ± 2.5%) which were similar to their glucose transfer rates determined by numerical models. Different flow rates (10–150 μL/h) were also assessed to see the effect of variable flow rate on transport of glucose across the chamber. Their results showed that at 50 μL/h, glucose concentration in the top channel is 6.81 mM and 5.76 mM in the bottom channel. At 150 μL/h, diffusion was negligible with which they concluded that the rate of glucose diffusion decreases with the flow rate which was concordant to their numerical model data. Effect of membrane porosity (0.01 and 0.1) was also checked on glucose transfer. The difference in the glucose concentration between the maternal and the fetal chamber was greater in the 0.01 porosity membrane as compared to the 0.1 porosity membrane. This occurred due to the lower resistance to mass transfer from the higher membrane porosity, suggesting that glucose diffusion increases with increasing membrane porosity.
16.4.2 Modifications of the Barrier Design
-
1.
Single-layered design: Miura et al. developed a similar placental barrier model with trophoblast cells [19]. However, in their design they used BeWo cells that mimic the syncytiotrophoblasts which are key cells involved in barrier function. No cells were seeded in the fetal channel making it a single-layered device. The device consisted of a 15 mm maternal and a 20 mm fetal channel, both of 2 mm width and 200 μm height - patterned on two PDMS layers by soft lithography. The device has a multi-layered vertical design in terms of the microfluidic layers, but has only one type of cell, making it single-layered in terms of the cellular layers in the device. A 4 mm circular chamber for cell seeding was made with a biopsy punch in the maternal channel and sealed using a thin PDMS membrane. BeWo cells were adhered to a 10 μm thick vitrified collagen membrane and they studied how fluid shear stress (FSS) affects the formation of microvilli under static and dynamic flow conditions. In the static condition, cells had sparse microvillar surfaces. Under flow conditions, the entire cell surface in the maternal chamber was covered with microvilli. The FSS was low at the centre of the chamber (∼0.001 dyn/cm2) as compared to the site near inlet and outlet of the microvillar (∼0.1 dyn/cm2). At the centre of the chamber, microvillar protrusions were long (>2 μm); however, near the site of inlet and outlet, the microvillar protrusions were shortened (<2 μm). The microvilli formation induced by FSS was quantified, by measuring total length of microvilli/field from the scanning electron microscopy (SEM) images for each FSS condition. The measured lengths of microvilli were significantly different under high- and low-FSS conditions. Also, the microvilli formation was increased 10.8-fold at the low-FSS area and 5.6-fold at the high-FSS area compared to the static culture conditions.
-
2.
Multicellular multi-layered design: Most of the on-chip platforms use filter membrane as scaffold to act as barrier, but these membranes affect cell behaviour as well as add a level of barrier which is not present in in vivo conditions. Therefore, a different multi-layered model was developed by Kreudar et al. called Membrick, which is a cell culture insert designed to be used in 24 well culture plates (Fig. 16.6) [20]. The model is a 3D bioprinted gelatin methacrylate (GelMA) biomembrane, which was used as an alternative to filter membranes. The model replicated early-stage placental barrier with human villous mesenchymal fibroblasts (resembling the stromal tissue of placental villi) embedded in a 3D bioprinted gelatin methacrylate (GelMA) biomembrane. The bioink was polymerised by light initiated polymerization using a 3D bioprinter to form the hydrogel membrane with a thickness of 330–400 μm within the Membrick body. After 3 days of culture with fibroblasts, the basolateral side of the membrane was seeded with HPVECs, while the apical side was seeded with BeWo trophoblast cells and cultured for 12 days with media change every day. Electrical resistance was measured to assess biomembrane integrity. The membrick showed lower resistance 18.22 ± 1.6 cm2 as compared to polyethylene terephthalate (PET) membrane 36.2 ± 2.8 cm2 devices. Membrick permeability was tested by using different weight molecules [457 Da Lucifer Yellow (LY), 3 kDa Dextran-Texas Red (DTR) and 70 kDa Dextran-FITC (DF)] in serum-free medium. After 24 h, permeation of smaller weight molecules (457 Da LY and 3 kDa DTR) was observed similar in both PET (10.37 μg/mL/cm2 LY and 10.1 μg/mL/cm2 DTR) and biomembrane (10.83 μg/mL/cm2 LY and 10.13 μg/mL/cm2 DTR). For 70 kDa DF PET membrane displayed a low hindrance (8.62 μg/mL/cm2) after 24 h, but the biomembrane showed much reduced biological membrane (2.41 μg/mL/cm2). This trilayer design was tested for successful growth of each type with cell-specific markers (HPVEC-CD31; BeWo-cytokeratin 7; HVMF-Vimentin) and showed a proof-of-concept study of culturing 3 cell types on a device.
-
3.
Design to simulate the barrier functions with villi-like projections: During the course of development, the placenta undergoes several structural changes which affect the surface area, cellular arrangement, and size of the organ. Thus, the barrier design described above was modified to resemble the in vivo like placental villous structure [21]. Using 2 photon polymerization which allows precision in structural patterns like external shape, pore size, and internal porosity of fabricated structures, Mandt et al. developed a planar placental barrier model wherein a separating GelMOD-AEMA hydrogel membrane of 100 μm thickness was fabricated within a microfluidic chip. The chip was fabricated in poly-(ethylene glycol)-dimethacrylate (PEGdma) and consisted of an X-shaped chamber with an intersection area of width 1 mm and length 1.4 mm. The GelMOD-AEMA membrane was fabricated at the intersection region, separating the chamber into a maternal and fetal compartment, and coated with fibronectin. The membrane consisted of five consecutive loops to mimic the villous structure of the placental barrier. BeWo trophoblast cells and HUVECs were seeded on the maternal and fetal sides of the membrane. After cells were attached, appropriate growth media were perfused through the channels at 50–70 μL/h. This design was made to assess the barrier function of the placenta. The barrier integrity was checked using riboflavin and fluorescently labelled dextran. Riboflavin was observed to pass through the barrier, while dextran did not. This design provides a much closer platform to study the barrier function as the hydrogel barrier and the villi-like projections would be similar to in vivo conditions compared to a membrane.

Schematic of placental cross-section (First trimester) and biological membrane system (Membrick) (a) placental villus contains fetal blood vessels (fb), endothelial cells (HPVEC), basal lamina (bl), mesenchymal stroma (st), human villous mesenchymal fibroblasts (HVMF), villous cytotrophoblast cells (vCTB), syncytiotrophoblast (STB) maternal blood (mb); (b) Scheme of the placental barrier model ofthe Membrick, the biological membrane is made with methacrylated gelatine (GelMA) with human villous mesenchymal fibroblasts (HVMF) embedded in it. BeWo and Human placental vascular endothelial cells (HPVEC) are cultured on the apical and basal side on the membrane respectively; (c) Depiction of cell culture insert scheme; biological membrane (1) two compartments for cell seeding (2, 3). The Membrick device can be kept hanging in 24 well by the device arms; the bottom of the cylindrical body includes a pipette rest for facilitated media exchange (4). (Figure is adapted from Kreudar et al. 2020)
16.4.3 Applications of the Placenta-on-Chip Devices that Simulate the Barrier Function
-
1.
Maternal infection: E. coli is the predominant microorganism observed during infections and was chosen as a model to simulate these conditions. The presence of an immune response against the bacteria and whether it can cross the barrier was tested using the placenta-on-chip model. The design was principally similar to that described above with minor modifications. Zhu et al. cultured BeWo human trophoblast cells and HUVECs on the maternal and fetal channel sides of a semipermeable polycarbonate membrane [22]. The membrane was attached to the upper PDMS layer by electrostatic interaction and the bottom surface of the membrane was smeared with PDMS glue. Following this, the glue was cured and the two PDMS layers were plasma bonded together. The channels (w = 1.5 mm, h = 400 μm, l = 1.5 cm) were coated with type 1 collagen to promote cell adherence and growth before seeding the cells. Growth media were perfused through the channels at 10 μL/h using a syringe pump, resembling the maternal and fetal circulation in vivo. To simulate maternal infection, E. coli was applied to the maternal side, and the cells were tested for inflammatory responses. An increased secretion of inflammatory cytokines by trophoblasts and the adhesion of maternal macrophages was observed in response to E. coli. There was significant cell death in both the compartments, mimicking cell death during membrane rupture. Due to bacterial infection, there was a surge of inflammatory cytokines in both the chambers. The integrity of the fetal cell monolayer was also compromised, i.e. the endothelial barrier was disrupted. Overall, this model was shown to simulate not just the barrier functions, but also that of membrane rupture in women experiencing preterm delivery.
-
2.
Drug transport from mother to child: Caffeine is a widely consumed xenobiotic and studies have shown its transfer from the maternal to the fetal chamber which could cause long-term damage to the developing fetus as it lacks the enzymes which could inactivate caffeine. Therefore, it is essential to understand the rate of caffeine transfer across the placenta. Naltrexone is a medication prescribed for opioid use disorder with unknown potential harm to the fetus. To address if placenta-on-chip devices are useful in studying drug transport, Pemathilaka et al. utilized the placental barrier model [23, 24]. Essentially a design similar to that described above, two layers of PDMS, with channel features of width 400 μm and height 100 μm, were bonded by plasma treatment with the membrane sandwiched between them. The membrane was coated with an E-C-L (Entactin–collagen IV–laminin) solution and BeWo trophoblast cells and HUVECs were cultured on either side of a polyester track etched membrane with 0.4 μm sized pores under media perfusion at 50 μL/h. The integrity of the device and cells cultured was tested using cell type-specific antibodies. They observed a steady transfer of caffeine from the maternal to the fetal chamber. In this device, the transport of 6β-naltrexol across the placental barrier was also studied and the authors found that ~ 2.5% of the total maternal concentration of 6β-naltrexol could be passed on in the fetal chamber. The authors also demonstrated that the HUVECs in the fetal chamber had altered expression of cytokines, suggestive of the harmful effect of the compound on the fetal cells. Thus, this study provides proof of concept on application of this device in drug transport from mother to child.
-
3.
Nanoparticles: Nanomedicine is gaining momentum and there are several studies demonstrating its therapeutic usefulness. The risk of exposure to such particles in pregnant women in vivo is yet unknown. These exposures can lead to production of reactive oxygen species. Reactive oxygen species are unstable, free radical oxygen carrying molecules which are highly reactive. Increase in ROS can cause damage to DNA, RNA, and protein which could lead to cell death. Two studies have reported the effect of nanoparticle exposure on the placental function (mainly the placenta’s barrier function). Schuller et al. developed a multi-layered placental barrier model consisting of a porous PET membrane of pore size 3 μm and thickness 9 μm coated with type 1 collagen with an interdigitated electrode (impedance microsensor) array for nanoparticle risk assessment [25]. The channel features were patterned on microscope glass slides of height 1 mm and the layers were bonded together using double-sided adhesive tape. BeWo trophoblast cells were adhered to the membrane and the placental barrier integrity with exposure to nanoparticles was monitored. This model was tested using different concentrations of nanomaterials made of silicon dioxide (SiO2), titanium dioxide (TiO2), and zinc oxide (ZnO), which are present in various cosmetics and day to day consumables and further assessed with Reactive oxygen species (ROS) production. To show that a confluent layer of BeWo cell line was achieved on the device, tight junction immunofluorescence protein Zonula occludens protein 1 (ZO1), which was seen to be intact at all the cell junctions. Molecule transport across the microsensor membrane showed that their procedure did not affect the cellular function. A comparative study of the newly designed interdigitated membrane-bound impedance sensor array model vs standard nanotoxicological viability assays including Presto Blue and reactive oxygen species (ROS) assays was done. TiO2 and SiO2 devices showed no loss of barrier integrity after 24 h of exposure which was found in line with Presto blue viability assay. Reactive oxygen species were found to be produced equally in both models after exposure with both types of nanoparticles. ZnO nanoparticles in both standard and on-chip models showed similar levels of increase in ROS after 4 h and 24 h of exposure. The increasing ROS in turn created a similar level of cytotoxicities after 4 h and 24 h of exposure, respectively. Yin et al. developed a single-layered planar microfluidic device consisting of a PDMS layer with a central matrix channel and two parallel channels on either side of it [26]. This PDMS layer was bonded to an unpatterned PDMS slab after plasma treatment. Matrigel was perfused into the central channel of width 300 μm and height 50 μm and allowed to gelate. BeWo and HUVECs were introduced into the side channels and cultured on either side of the matrigel extracellular matrix by tilting the device after seeding. After 24 h of culture, appropriate growth media flowed through the channels at 20 μL/h. On the device, they were able to show that increasing concentration of nanoparticles would directly affect the viability of the cells in both maternal and fetal chambers which implied that it could cause placental barrier to rupture. Increase of inflammatory molecules in both the chambers indicates severe effects of these nanoparticles on the placental function. This model system has a high potential for assessment of various drug delivery systems and biomedical research purposes.
16.4.4 Placenta-on-Chip Models of Trophoblasts Migration
In models of trophoblast migration, trophoblast invasion or migration in response to chemical gradients is studied. Unlike the barrier function devices, there are very few designs available to study invasion. Most of the studies are limited to an early proof of concept and very few applications have been tested.
16.4.4.1 Devices to Study Trophoblast Migration in Response to Chemical Gradients
Abbas et al. developed a trophoblast invasion model using a PDMS device with a central channel and two side channels, each of length 2 cm, width 1.3 mm, and height 150 μm, separated by micropillars (Fig. 16.7) [27]. The PDMS layer was fabricated by soft lithography techniques and plasma bonded to a glass coverslip. Matrigel embedded with primary extravillous trophoblasts was loaded into the central microchannel and allowed to polymerise. One of the side channels was perfused with media containing granulocyte-macrophage colony-stimulating factor (GM-CSF), while the other side channel was perfused with media alone (control), both at 50 μL/h for 12 h. The model was used to study the migration of the trophoblasts under the influence of the cytokine GM-CSF. The migration of the trophoblast cells in the absence of GM-CSF was unidirectional, whereas in the presence of GM-CSF, cellular migration was increased with increasing directionality and velocity. This approach allows accurate estimation of migration of individual cell types’ response to chemo attractants by quantifying their migration speed, directionality, and pattern of migration (single/collective).

Placenta-on-chip model for studying trophoblast invasion. (a) The microfluidic device consists of a central channel and two side channels (channel A and B). EVTs isolated from first trimester placenta are stained with a cell tracker and are embedded in a Matrigel matrix in the central channel. Media with and without GM-CSF are perfused through channel A and channel B, respectively, creating a gradient of GM-GSF across the central channel. The migration of the cells in the central channel is tracked using time lapse microscopy. (b) The purity of the cells embedded in the central channel was confirmed by immunostaining of EVT for HLA-G (green) and staining the nucleus of each cell using DAPI dye. (Figure is adapted with permission from Abbas et al © (2017) the Royal Society)
While the design described above finds application in studying trophoblast migration, the application is restricted to chemical gradients and is a monocellular design. To circumvent this limitation, Yong Pu et al. developed a 3D microfluidic platform to mimic the trophoblast microenvironment for studying trophoblast invasion in vitro (Fig. 16.8) [28]. The platform, fabricated in PDMS, consisted of a central compartment and two outer channels (w = 200 μm, h = 100 μm) on either side of it, separated by a barrier of width 50 μm consisting of pillars. After coating the channels with fibronectin, HUVECs and extravillous trophoblasts (HTR8/SVneo cells) were seeded in the central compartment and the outer channels, respectively. Growth media were perfused through the channels at 0.6 μL/h using syringe pumps. The model was used to study the invasion properties of the trophoblast cells. To achieve a confluent monolayer of cells on the device, three matrices, Gelatin, Matrigel, and Fibronectin, were tested. Attachment of cells was found on all the three matrices, but fibronectin had the highest cell attachment maximum. Integrity of the barrier and barrier permeability was assessed using FITC-Dextran diffusion. The cell layers on the pillar and device restricted the diffusion of FITC-dextran into the central compartment and resulted in a gradual build-up gradient reaching the centre of the central compartment by 72 h. They assessed the invasion of HTR8/SVneo cells under the influence of folic acid which aids in the invasion of the extravillous trophoblast cells in vivo. They observed enhanced trophoblast migration from the outer chamber (HTR8/SVneo) to the central compartment (HUVECs) in the device.

3D planar multilayer chip (a) 3D microfluidic chip scheme depicting the following components: (1) a central compartment (red) with a central feeder line supplied by two inlet ports (a and b) connected to two outlet ports (c and d); (2) two outer channels (blue) with two outer feeder lines supplied by two inlet ports (e and f) connected to two outlet ports (g and h), and a (3) pillar barrier in between. Arrow corresponds to the directionality of medium flow; (b) Close up scheme of the center of the 3D microfluidic chip with the central compartment (red), outer channels (blue, width (CW): 200 μm), and pillar barrier (width (BW): 50 μm), filled with pillars (pillar spacing (PS): 3 μm). (Figure adapted permission from Pu et al. © (2021) Royal society of chemistry)
16.4.4.2 Design to Simulate Spiral Artery
Kuo et al. developed a 3D printed bioengineered placenta model using gelatin methacrylate (GelMA) to mimic the features of maternal decidua for studying trophoblast migration/invasion (Fig. 16.9) [29]. The cylindrical model was loaded with different components such as cells, ECM, and growth factors at different radial positions. BeWo trophoblast cells were embedded in the GelMA hydrogel at the periphery of the construct, while epithelial growth factor (EGF) was loaded at the centre. EGF gradients were created to understand migration patterns. This bioengineered model is designed to recapitulate spiral artery remodelling under dynamic flow conditions. The use of this model is to study the migration of cells under the influence of a stimulus. Using this model, they showed a positive correlation between migration pattern of trophoblast cells and epidermal growth factor (EGF). This design can be used to understand pharmacological and molecular interactions of therapeutic agents for pre-eclampsia.

Characterization of the GelMA printing platform (a and b) Lattice structures with 1 × 1 mm spacing. (c) Solid disc structure (diameter = 10 mm). (d) CAD model for spiral (height, 10 mm; channel diameter, 1 mm; pitch, 3 mm; taper helix angle, 20°; 3 revolutions). (e) Overall CAD model used for 3D printing: spiral embedded in a cube. (f) 3D printed spiral (blue) in a cube (transparent): side view. (g) 3D printed spiral (blue) in a cube (transparent): cross-sectional view. (Figure adapted with permission from Kuo et al. © (2016) American Chemical Society)
16.5 Limitation of the Placenta-on-Chip Devices
There are multiple reported designs of placenta-on-chip which have attempted to simulate the barrier functions or trophoblast migration. However, most of these are still at the proof of concept stage and very little advancement is made in terms of design improvements or applications. Listed below are some major limitations that the placenta-on-chip devices pose. It is important that one is aware of these limitations while interpreting the data from various studies. Most of the placenta-on-chip designs have a single layer of trophoblast cells or they are coupled with HUVECs. Such bilayer devices, mimic the floating villus, are similar to those of term placenta villi. The first and second trimester placenta villi are three to four layered structures (see above) and the molecules need to pass through these layers to reach the fetus. This aspect is not captured in the existing devices. The floating villus consist of cytotrophoblast cells and syncytiotrophoblast cells, each having different gene expression patterns as well as variable affinities towards biomolecules for their transport from maternal to fetal chamber. BeWo cells have cytotrophoblast-like properties and they need to be synctialized to become syncytiotrophoblast cells, making this device less homologous to the in vivo conditions. However, most models have used cytotrophoblast cells as the representative of the trophoblast layer, which is not physiologically relevant. The physiological shear stress levels at the feto-maternal interface are less explored, but studies have tried to calculate levels of shear stress in vivo [30,31,32]. The shear stress levels experienced in vivo are difficult to recapitulate in these devices. Also, the placental tissue produces a lot of hormones like βhCG, progesterone, oestrogen, etc. All the models have failed to test this important feature of the placental tissue.
The process of migration in vivo is assisted by multiple factors produced by various cell types. All the model devices used to study the migration pattern have used only one such factor to show mere tendency of highly migratory cells to react towards stimulus. Migration of trophoblast cells is influenced by other cell types in vivo. For example, decidual cells stimulate the migration and allow passage of these trophoblast cells by secreting various pro-invasive factors [33, 34]. None of these devices have taken the true physiology into consideration for performing these studies.
16.6 Future Direction
In the field of placenta-on-chip (PoC), the recent research progress gives hope of using these devices for toxicity testing, drug testing, and in understanding the basic physiology of the placenta in the near future. However, like any other organ-on-chip models, PoCs are also facing some serious challenges as described below. The community needs to address these issues together.
First, every PoC is to be validated for the application for which it is fabricated. For example, if a PoC is developed as the model for transplacental barrier, they should be validated for the same. However, there is sufficient ambiguity in terms of how a PoC should be validated. Due to the lack of any broadly accepted standard, it is presently not possible to comment if a PoC model is mimicking the in vivo conditions reliably. We also need to keep in mind that a PoC model validated for one molecule may not give reliable results for a different molecule of very different size, charge, and diffusivity. However, developing separate models for separate systems and validating them individually every time against some standard is next to impossible. Hence, the community should come together to set standards against which a newly developed PoC model can be validated.
To resolve this issue, engineering knowledge merged with an understanding of placental biology can be very useful. In any transport process such as transport of biochemicals across the placental barrier, surface to volume ratio is important. It becomes further critical when we reach micron scale level as in microfluidic devices and tissue capillaries. Other important factors are flow velocity, thickness of the boundary layers, transport parameters such as diffusivity and viscosity, porosity of the membrane and the extracellular matrix, etc. The researchers need to estimate the range of these parameters in the human placenta and then to formulate one or more relevant dimensionless numbers that can be matched between in vivo and in vitro when designing a new device. Further, unlike many other organs, the placenta is a continuously developing dynamic structure. The development of the placenta happens over a specific timescale. While designing PoC models which are generally static, one needs to also consider the dynamic behaviour of the placenta.
Like most of the organ-on-chip devices, PoCs are also mostly made of PDMS, which is known to absorb hydrophobic molecules. Additionally, while PDMS is an excellent polymer for lab scale demonstration of a concept, it is not suitable for large scale production. Hence, to make PoCs popular among researchers, the researchers should search for alternative materials such as polymethyl methacrylate (PMMA) or polystyrene (PS). Another major issue with PoCs is availability of relevant cells. The cell type to be used in PoC models should be physiologically relevant. Quite often, the cell type used in PoC devices are carcinoma cells which do not truly represent a healthy placenta. The other alternative is to use primary cells isolated from aborted placenta. However, the availability of such primary cells is limited, and they often have serious ethical and sociocultural concerns. A plausible alternative is the creation of placenta-specific immortalized cell lines made from the tissues of healthy and consenting adult donors.
Last but not the least, microfluidic experiments are often not very easy to set up by the end user. To maintain the flow precisely in a microfluidic device, many accessories such as pumps, valves, and tubings are needed. Such additional accessories make the device bulky and difficult to handle. A slightly faulty operation may lead to situations such as backflow and leakage. Hence, if such devices are not designed to be of “Plug and Play” type, the researchers trained in biology may find a barrier in adopting the technology for regular use. Hence, it is important to keep the design and operation of the microfluidic platform simple. Ideally, the device should be easy to use, modular, and robust. Collection of samples and imaging of the cells should also be hassle-free in the device. However, doing so should not compromise the critical features of an in vivo placenta that the device intends to mimic. A very simple model may be excellent for analysis and imaging, but it may miss many important structural and functional features of placenta. On the other hand, a complex model may be able to mimic various aspects of the placenta, but may offer a significant challenge in operation. Striking the balance is a challenge for a PoC designer.
16.7 Conclusion
This chapter has presented the recent developments in designing and application of PoC devices. We have discussed various available models, their advantages, and limitations. With the development of placenta-specific cell lines, modular designs, and integration of the knowledge of biology and engineering, PoC devices can be invaluable models for drug and toxicity testing in near future.
References
Ashary N, Tiwari A, Modi D (2018) Embryo implantation: war in times of love. Endocrinology 159:1188–1198. https://doi.org/10.1210/EN.2017-03082
Turco MY, Moffett A (2019) Development of the human placenta. Development 146. https://doi.org/10.1242/DEV.163428/223131
Aplin JD, Ruane PT (2017) Embryo–epithelium interactions during implantation at a glance. J Cell Sci 130:15–22. https://doi.org/10.1242/JCS.175943
Burton GJ, Jauniaux E (2018) Development of the human placenta and fetal heart: Synergic or independent? Front Physiol 9:373. https://doi.org/10.3389/FPHYS.2018.00373/BIBTEX
Burton GJ, Redman CW, Roberts JM, Moffett A (2019) Pre-eclampsia: pathophysiology and clinical implications. BMJ 366. https://doi.org/10.1136/BMJ.L2381
Yabe S, Alexenko AP, Amita M et al (2016) Comparison of syncytiotrophoblast generated from human embryonic stem cells and from term placentas. Proc Natl Acad Sci U S A 113:E2598–E2607. https://doi.org/10.1073/PNAS.1601630113/-/DCSUPPLEMENTAL
Rubinchik-Stern M, Eyal S (2012) Drug interactions at the human placenta: What is the evidence? Front Pharmacol 3:126. https://doi.org/10.3389/FPHAR.2012.00126/BIBTEX
Ashary N, Bhide A, Chakraborty P et al (2020) Single-cell RNA-seq identifies cell subsets in human placenta that highly expresses factors driving pathogenesis of SARS-CoV-2. Front Cell Dev Biol 8:783. https://doi.org/10.3389/FCELL.2020.00783/BIBTEX
Shende P, Gaikwad P, Gandhewar M et al (2021) Persistence of SARS-CoV-2 in the first trimester placenta leading to transplacental transmission and fetal demise from an asymptomatic mother. Hum Reprod 36:899–906. https://doi.org/10.1093/HUMREP/DEAA367
Adibi JJ, Marques ETA, Cartus A, Beigi RH (2016) Teratogenic effects of the Zika virus and the role of the placenta. Lancet 387:1587–1590. https://doi.org/10.1016/S0140-6736(16)00650-4
Ostrander B, Bale JF (2019) Congenital and perinatal infections. Handb Clin Neurol 162:133–153. https://doi.org/10.1016/B978-0-444-64029-1.00006-0
Mbachu II, Ejikunle SD, Anolue F et al (2020) Relationship between placenta malaria and mother to child transmission of HIV infection in pregnant women in South East Nigeria. Malar J 19:1–8. https://doi.org/10.1186/S12936-020-03171-2/TABLES/6
Ayisi JG, Van Eijk AM, Ter Kuile FO et al (2003) The effect of dual infection with HIV and malaria on pregnancy outcome in western Kenya. AIDS 17:585–594. https://doi.org/10.1097/01.aids.0000042977.95433.37
McNally R, Alqudah A, Obradovic D, McClements L (2017) Elucidating the pathogenesis of pre-eclampsia using in vitro models of spiral uterine artery remodelling. Curr Hypertens Rep 19:1–13. https://doi.org/10.1007/S11906-017-0786-2/TABLES/2
Lee JS, Romero R, Han YM et al (2016) Placenta-on-A-chip: A novel platform to study the biology of the human placenta. J Matern Neonatal Med 29:1046–1054. https://doi.org/10.3109/14767058.2015.1038518
Blundell C, Tess ER, Schanzer ASR et al (2016) A microphysiological model of the human placental barrier. Lab Chip 16:3065–3073. https://doi.org/10.1039/c6lc00259e
Blundell C, Yi YS, Ma L et al (2018) Placental drug transport-on-a-chip: a microengineered in vitro model of transporter-mediated drug efflux in the human placental barrier. Adv Healthc Mater 7:1–17. https://doi.org/10.1002/adhm.201700786
Mosavati B, Oleinikov AV, Du E (2020) Development of an organ-on-a-chip-device for study of placental pathologies. Int J Mol Sci 21:1–12. https://doi.org/10.3390/ijms21228755
Miura S, Sato K, Kato-Negishi M et al (2015) Fluid shear triggers microvilli formation via mechanosensitive activation of TRPV6. Nat Commun 6:1–11. https://doi.org/10.1038/ncomms9871
Kreuder AE, Bolaños-Rosales A, Palmer C et al (2020) Inspired by the human placenta: a novel 3D bioprinted membrane system to create barrier models. Sci Rep 10:1–14. https://doi.org/10.1038/s41598-020-72559-6
Mandt D, Gruber P, Markovic M et al (2018) Fabrication of biomimetic placental barrier structures within a microfluidic device utilizing two-photon polymerization. Int J Bioprinting 4:1–12. https://doi.org/10.18063/IJB.v4i2.144
Zhu Y, Yin F, Wang H et al (2018) Placental barrier-on-a-chip: modeling placental inflammatory responses to bacterial infection. ACS Biomater Sci Eng 4:3356–3363. https://doi.org/10.1021/acsbiomaterials.8b00653
Pemathilaka RL, Caplin JD, Aykar SS et al (2019) Placenta-on-a-chip: in vitro study of caffeine transport across placental barrier using liquid chromatography mass spectrometry. Glob Challenges 3:1800112. https://doi.org/10.1002/gch2.201800112
Pemathilaka RL, Reynolds DE, Hashemi NN (2020) Maternally administered naltrexone and its major active metabolite 6β-naltrexol transport across the placental barrier in vitro. bioRxiv 1–31. https://doi.org/10.1101/2020.04.16.045229
Schuller P, Rothbauer M, Kratz SRA et al (2020) A lab-on-a-chip system with an embedded porous membrane-based impedance biosensor array for nanoparticle risk assessment on placental Bewo trophoblast cells. Sens Actuators B Chem 312. https://doi.org/10.1016/j.snb.2020.127946
Yin F, Zhu Y, Zhang M et al (2019) A 3D human placenta-on-a-chip model to probe nanoparticle exposure at the placental barrier. Toxicol In Vitro 54:105–113. https://doi.org/10.1016/j.tiv.2018.08.014
Abbas Y, Oefner CM, Polacheck WJ et al (2017) A microfluidics assay to study invasion of human placental trophoblast cells. J R Soc Interface 14. https://doi.org/10.1098/rsif.2017.0131
Pu Y, Gingrich J, Veiga-Lopez A (2021) A 3-dimensional microfluidic platform for modeling human extravillous trophoblast invasion and toxicological screening. Lab Chip 21:546–557. https://doi.org/10.1039/d0lc01013h
Kuo CY, Eranki A, Placone JK et al (2016) Development of a 3D printed, bioengineered placenta model to evaluate the role of trophoblast migration in preeclampsia. ACS Biomater Sci Eng 2:1817–1826. https://doi.org/10.1021/acsbiomaterials.6b00031
Tun WM, Yap CH, Saw SN et al (2019) Differences in placental capillary shear stress in fetal growth restriction may affect endothelial cell function and vascular network formation. Sci Rep 9:1–10. https://doi.org/10.1038/s41598-019-46151-6
Lecarpentier E, Bhatt M, Bertin GI et al (2016) Computational fluid dynamic simulations of maternal circulation: wall shear stress in the human placenta and its biological implications. PLoS One 11:1–18. https://doi.org/10.1371/journal.pone.0147262
Sprague B, Chesler NC, Magness RR (2010) Shear stress regulation of nitric oxide production in uterine and placental artery endothelial cells: experimental studies and hemodynamic models of shear stresses on endothelial cells. Int J Dev Biol 54:331–339. https://doi.org/10.1387/ijdb.082832bs
Sharma S, Godbole G, Modi D (2016) Decidual control of trophoblast invasion. Am J Reprod Immunol 75:341–350. https://doi.org/10.1111/AJI.12466
Godbole G, Suman P, Malik A et al (2017) Decrease in expression of HOXA10 in the decidua after embryo implantation promotes trophoblast invasion. Endocrinology 158:2618–2633. https://doi.org/10.1210/EN.2017-00032
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2022 The Author(s), under exclusive license to Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Bhide, A., Aboo, A., Sawant, M., Majumder, A., Paul, D., Modi, D. (2022). Placenta on Chip: A Modern Approach to Probe Feto-Maternal Interface. In: Mohanan, P.V. (eds) Microfluidics and Multi Organs on Chip . Springer, Singapore. https://doi.org/10.1007/978-981-19-1379-2_16
Download citation
DOI: https://doi.org/10.1007/978-981-19-1379-2_16
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-19-1378-5
Online ISBN: 978-981-19-1379-2
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)