
Abstract
The blood-brain barrier (BBB) is a dynamic barrier essential for maintaining the microenvironment of the brain. Although the special anatomical features of the BBB determine its protective role for the central nervous system (CNS) from blood-borne neurotoxins, however, the BBB extremely limits the therapeutic efficacy of drugs into the CNS, which greatly hinders the treatment of major brain diseases. This chapter summarized the unique structures of the BBB; described a variety of in vivo and in vitro experimental methods for determining the transport properties of the BBB and the permeability of the BBB to water, ions, and solutes including nutrients, therapeutic agents, and drug carriers; and presented recently developed mathematical models which quantitatively correlate the anatomical structures of the BBB with its barrier functions. Recent findings for modulation of the BBB permeability by chemical and physical stimuli were described. Finally, drug delivery strategies through specific trans-BBB routes were discussed.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


1 Introduction
The most complicated and amazing organ in our body is the brain. It contains ∼100 billion neurons and ∼1 trillion glial cells (supporting cells in the brain including astrocytes, oligodendrocytes, microglia, and ependymal cells). Along with a tremendous amount of blood vessels, these cells and surrounding extracellular matrix form a highly complex, though well-organized, 3-D interconnecting arrays. In order to perform its highly complicated tasks, the brain needs a substantial amount of energy to maintain electrical gradients across neuronal membranes and consequently requires a sufficient supply of oxygen and nutrients. Although it only accounts for ∼2% of the body weight, the brain uses ∼20% of the blood supply. The blood is delivered through a complex network of blood vessels that runs >650 km and passes a surface area of ∼20 m2. The mean distance between adjacent capillaries is ∼40 μm, which allows almost instantaneous equilibration in the brain tissue surrounding the microvessels for small solutes such as glucose, amino acids, vitamins, oxygen, etc. However, unlike peripheral microvessels in other organs where there is a relatively free small solute exchange between the blood and tissue, the microvessels in the brain (cerebral microvessels) constrain the movement of molecules between blood and the brain tissue (Abbott 1992; Pardridge 1998). This characteristic provides a natural defense against toxins circulating in the blood, which, on the other hand, prevents the delivery of therapeutic agents to the brain.
The vascular barrier system in the brain consists of the blood-brain barrier (BBB) and the blood-cerebrospinal fluid (CSF) barriers. The blood-brain barrier (BBB) is the name for the wall of the cerebral microvessels in the brain parenchyma. At the surface of the brain parenchyma, microvessels running in the pia mater are called pial microvessels, which are often used as in vivo models for studying the BBB permeability. Due to its unique structure that will be discussed in the next section, the BBB maintains very low permeability to water and solutes. In the middle of the brain parenchyma, there are ventricular cavities (ventricles) filled with cerebrospinal fluid (CSF) secreted by the epithelial cells of choroid plexus (Brown et al. 2004a, b; Engelhardt and Coisne 2011). The choroid plexus is a highly vascular tissue with leaky, fenestrated capillaries covered with ependymal epithelium, which has relatively tight junctions. The multicell layer between the blood and the CSF in the choroid plexuses is called the blood-CSF barrier. Since the area of the BBB is about 1000 times that of the blood-CSF barrier, it is more important to circumvent the impermeability of the BBB for delivering drugs to the brain (Pardridge 2007). The total surface area of the BBB constitutes by far the largest interface for blood-brain exchange, which is between 12 and 18 m2 for the average human adult (Nag and Begley 2005). Unlike these two tight blood barriers, the interface between the CSF and brain tissue along the ependymal surface of the ventricles and that between pia mater and brain tissue, so called the brain-CSF barriers, is rather leaky, implying a possible route for drug delivery to the brain. The CSF is formed by the choroid plexuses of the ventricles, passes the ventricles to the subarachnoid space over the pia mater, and is finally absorbed to the venules in the dura mater through arachnoid microvilli and arachnoid granulations (Redzic and Segal 2004). Recent studies (reviewed in Bakker Erik et al. 2016; Brinker et al. 2014) revealed that the process of CSF absorption is far more complicated than the previous view, due to the discovery of brain lymphatic system (Aspelund et al. 2015; Louveau et al. 2015; Greenwood 2017) and perivascular and paravascular clearance mechanisms (Carare et al. 2008; Iliff et al. 2013).
2 The Blood-Brain Barrier
The blood-brain barrier (BBB) is a unique dynamic regulatory interface between the cerebral circulation and the brain tissue, and it is essential for maintaining the microenvironment within the brain. No other body organ so absolutely depends on a constant internal microenvironment as does the brain. In the brain, the extracellular concentrations of amino acids and ions such as Na+, K+, and Ca2+ must be retained in very narrow ranges. If the brain is exposed to big chemical variations for these molecules, neurons would not function properly because some amino acids serve as neurotransmitters and certain ions modify the threshold for neuronal firing. The BBB also protects the central nervous system (CNS) from blood-borne neuroactive solutes, such as glutamate, glycine, norepinephrine, epinephrine, and peptide hormones (Smith 2000), which can increase with physiological changes (e.g., diet and stress) and pathological changes (e.g., injury and diseases). In addition, the BBB plays a key role in facilitating the brain uptake of essential nutrients like glucose, hormones, and vitamins and larger molecules like insulin, leptin, and iron transferring to sustain brain growth and metabolism (Zhang and Pardridge 2001).
The term “blood-brain barrier” was coined by Lewandowsky in 1900 while he demonstrated that neurotoxic agents affected brain function only when directly injected into the brain but not when injected into the systemic circulation (Lewandowsky 1900). Nevertheless, the first experimental observation of this vascular barrier between the cerebral circulation and the CNS should date back to the 1880s, when Paul Ehrlich discovered that certain water-soluble dyes, like trypan blue, after injected into the systemic circulation, were rapidly taken by all organs except the brain and spinal cord (Ehrlich 1885). Ehrlich interpreted these observations as a lack of the affinity of the CNS for the dyes. However, subsequent experiments performed by Edwin Goldmann, an associate of Ehrlich, demonstrated that the same dyes, when injected directly into the CNS, stained all types of cells in the brain tissue but not any other tissues in the rest of the body (Goldmann 1913). It took additional 70 years until this barrier was firstly localized to cerebral microvascular endothelial cells by electron microscopic studies performed by Reese and Karnovsky (1967). Although the concept of the BBB has continued to be refined over the past several decades, the recent understanding of the basic structure of the BBB is built on the general framework established by their studies in the late 1960s; more specifically, the BBB exists primarily as a selective transport barrier at the level of cerebral capillary endothelium.
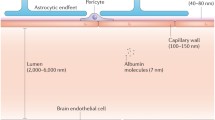
The anatomical structure of the blood-brain barrier (BBB) is sketched in Fig. 1. The circumference of the BBB lumen is surrounded by endothelial cells, the opposing plasmalemma membranes of which are connected by tight junctions. At the luminal surface of the endothelial cell, there is a rather uniform fluffy glycocalyx layer (Ueno et al. 2004; Haeren et al. 2016; Yoon et al. 2017). This mucopolysaccharide structure is highly hydrated in electrolytic solution and contains large numbers of solid-bound fixed negative charges due to the polyanionic nature of its constituents abundant in glycoproteins, acidic oligosaccharides, terminal sialic acids, proteoglycan, and glycosaminoglycans aggregates. Pericytes attach to the abluminal membrane of the endothelium at irregular intervals. In a peripheral microvessel, there is a loose and irregular basal lamina (or basement membrane) surrounding the pericytes. In contrast, in the BBB, pericytes and endothelial cells are ensheathed by a very uniform basement membrane of 20–100 nm thickness (Farkas and Luiten 2001), which is composed of collagen type IV, heparan sulfate proteoglycans, laminin, fibronectin, and other extracellular matrix proteins (Bakker Erik et al. 2016). The basal lamina is contiguous with the plasma membranes of astrocyte end feet that wrap almost the entire abluminal surface of the endothelium (Pardridge 1998).

Schematic of the three-dimensional view (a) and cross-sectional view (b) of the blood-brain barrier (BBB) or cerebral microvessel (the microvessel in the brain) wall. In addition to other structures as in a peripheral microvessel, the BBB is wrapped by astrocyte foot processes (AP). BM, basement membrane (or basal lamina); E, endothelial cell; EN, nucleus of endothelial cell; P, pericytes; G, surface glycocalyx layer; TJ, tight junction. Modified from (Li et al. 2010a, b)
In addition to the anatomical structures, the BBB differs from the peripheral microvessels in the following aspects. The mitochondrial content of the endothelial cells forming the BBB is greater than that of such cells in all nonneural tissues. It is suggested that this larger metabolic work capacity may be used to maintain the unique structural characteristics of the BBB and/or by metabolic pumps that may require energy to maintain the differences in composition of the cerebral circulation and the brain tissue (Oldendorf et al. 1977). The BBB has high electrical resistance, much less fenestration, and more intensive junctions, which are responsible for restricting paracellular passage of water and polar solutes from the peripheral circulation entering into the CNS (Butt et al. 1990; Hawkins and Davis 2005). Between adjacent endothelial membranes, there are junctional complexes which include adherens junctions (AJs), tight junctions (TJs), and possibly gap junctions (Simard et al. 2003). The structure of the junction complexes between endothelial cells is shown in Fig. 2 (Kim et al. 2006; Abbott et al. 2010). Both AJs and TJs act to restrict paracellular transport across the endothelium, while gap junctions mediate intercellular communication. AJs are ubiquitous in the vasculature, and their primary component is vascular endothelial (VE)-cadherin. They basically mediate the adhesion of endothelial cells to each other and contact inhibition during vascular growth and remodeling. Although disruption of AJs at the BBB can lead to increased permeability, TJ is the major junction that confers the low paracellular permeability and high electrical resistance (Romero etal. 2003). The tight junction complex includes two classes of transmembrane molecules: occludins and claudins. These transmembrane proteins from adjacent endothelia cells interact with each other and form seals in the spaces between adjacent endothelial cells. The cytoplasmic tails of the transmembrane proteins are linked to the actin cytoskeleton via a number of accessory proteins such as members of the zonula occludens family, ZO-1, ZO-2, and ZO-3.

Schematic of junctional complex in the paracellular pathway of the BBB. Modified from (Abbott et al. 2010)
A number of grafting and cell culture studies have suggested that the ability of cerebral endothelial cells to form the BBB is not intrinsic to these cells, but the cellular milieu of the brain somehow induces the barrier property into the blood vessels. It is believed that all components of the BBB are essential for maintaining functionality and stability of the BBB. Pericytes seem to play a key role in angiogenesis, structural integrity, and maturation of cerebral microvessels (Ballabh et al. 2004). The extracellular matrix of the basal lamina appears to serve as an anchor for the endothelial layer via interaction of laminin and other matrix proteins with endothelial integrin receptors (Moody 2006). It was suggested that astrocytes are critical in the development and/or maintenance of unique features of the BBB. Additionally, astrocytes may act as messengers to or in conjunction with neurons in the moment-to-moment regulation of the BBB permeability (Ballabh et al. 2004).
3 Transport Pathways Across the Blood-Brain Barrier
The BBB endothelial cells differ from those in peripheral microvessels by more intensive tight junctions, sparse pinocytic vesicular transport, and much less fenestrations. The transport of substances from the capillary blood into the brain tissue depends on the molecular size, lipid solubility, binding to specific transporters, and electrical charge (Moody 2006). Figure 3 summarizes the transport routes across the BBB (Neuwelt 2004). Compared to the peripheral microvessel wall, the additional structure of the BBB and tighter endothelial junctions greatly restrict hydrophilic molecules transport through the gaps between the cells, i.e., the paracellular pathway of the BBB, route A in Fig. 3. In contrast, small hydrophobic molecules such as O2 and CO2 diffuse freely across plasma membranes following their concentration gradients, i.e., the transcellular lipophilic diffusion pathway, route C in Fig. 3. The BBB permeability to most molecules can be estimated on the basis of their octanol/water partition coefficients (Sawchuk and Elmquist 2000). For example, diphenhydramine (Benadryl), which has a high partition coefficient, can easily cross the BBB, whereas water-soluble loratadine (Claritin) is not able to penetrate the BBB and has little effect on the CNS (Kay 2000).

Transport pathways across the brain endothelial cell. Modified from (Neuwelt 2004)
However, the octanol/water partition coefficients do not completely reflect BBB permeability to solutes. Some solutes with low partition coefficients that easily enter into the CNS generally cross the BBB by active or facilitated transport mechanisms, which rely on ion channels, specific transporters, energy-dependent pumps, and a limited amount of receptor-mediated transcytosis. Glucose, amino acids, and small intermediate metabolites, for example, are ushered into brain tissue via facilitated transport mediated by specific transport proteins (route B in Fig. 3), whereas larger molecules, such as insulin, transferrin, low-density lipoprotein, and other plasma proteins, are carried across the BBB via receptor-mediated (route D) or adsorptive transcytosis (route E). Some small molecules with high octanol/water partition coefficients are observed to poorly penetrate the BBB. Recent studies suggested that these molecules are actively pumped back into blood by efflux systems (route F in Fig. 3). These efflux systems greatly limit drug delivery across the BBB. For instance, P-glycoprotein (P-gp), which is a member of the adenosine triphosphate-binding cassette family of exporters, has been demonstrated to be a potent energy-dependent transporter. P-gp contributes greatly to the efflux of xenobiotics from the brain to the blood and has increasingly been recognized as having a protective role and being responsible for impeding the delivery of therapeutic agents (Schuetz et al. 1996). The organic anion transporters and glutathione-dependent multidrug resistance-associated proteins (MRPs) also contribute to the efflux of organic anions from the CNS, and many drugs with the BBB permeabilities that are lower than predicted are the substrates for these efflux proteins (Pardridge 1998; Abbott et al. 2010; Neuwelt 2004; Begley 2007). While the brain endothelium is the major barrier interface, the transport activity of the surrounding pericytes (Shimizu 2008), basement membrane, and astrocyte foot processes (Fig. 1) (Wolburg-Buchholz et al. 2009) also contributes to the BBB barrier function under physiological conditions and may act as a substitute defense if the primary barrier at the endothelium is compromised (Li etal. 2010a, b).
4 Quantification of Transport Across the Blood-Brain Barrier
4.1 Permeability of the Blood-Brain Barrier
The same as a peripheral microvessel, the wall of the BBB can be viewed as a membrane. The membrane transport properties are often described by Kedem-Katchalsky equations derived from the theory of irreversible thermodynamics (Curry 1983):
where J s and J v are the solute and volumetric fluxes and ΔC and Δp are the concentration and pressure difference across the membrane. L p, the hydraulic conductivity, describes the membrane permeability to water. P, the diffusive permeability, describes the permeability to solutes. σ f is the solvent drag or ultrafiltration coefficient that describes the retardation of solutes due to membrane restriction, and σ d, the reflection coefficient, describes the selectivity of a membrane to solutes. In many transport processes, σ f is equal to σ d, and thus we often use σ, the reflection coefficient, to represent both of them. These three coefficients can be determined experimentally and theoretically. In addition to these quantitative coefficients, there are other less quantitative permeability indicators for the BBB, e.g., brain uptake index (BUI) and brain efflux index (BEI) (Pardridge 1998). In the following sections, in vivo and in vitro experiments for determining permeability of the BBB are introduced, as well as the mathematical models.
4.2 Determination of the Blood-Brain Barrier Permeability In Vivo and Ex Vivo
Several in vivo and ex vivo rat models have been used for the study of the transport across the BBB, including pharmacokinetic methods (Cornford et al. 1992; Zlokovic et al. 1986), intracerebral microdialysis (de Lange et al. 1999; Zhang et al. 2017), positron emission tomography (PET) (Elsinga et al. 2004), magnetic resonance imaging (MRI) (Wang et al. 2007), the intravital microscopy study (Gaber et al. 2004), occluding single microvessel measurement (Easton and Fraser 1994), and single microvessel fluorescence imaging method (Yuan et al. 2009; Shi et al. 2014a, b, c).
Pharmacokinetic methods are used to evaluate the delivery of a molecule from the systemic circulation into the brain, in which the amount of solute delivered to the brain can be expressed by percentage of injected dose delivered per gram of the brain. Generally, a small volume of buffered Ringer’s solution containing the radiolabeled compound of interest and a radiolabeled diffusible reference compound as an internal standard (such as 3H-water) is injected into the common carotid artery, or the internal carotid artery, or the venule depending on different techniques. Then, the animal is sacrificed 5–15 s after injection, and the brain tissue and the injection solution are analyzed to calculate the brain uptake index (BUI), which is the ratio of radiolabeled test compound/3H reference in the brain, divided by the ratio of radiolabeled test compound/3H reference in the injection mixture. Another permeability indicator, brain efflux index (BEI), can also be determined using this method: BEI = (amount of test compound injected into the brain – amount of test compound remaining in the brain)/amount of test compound injected into the brain. The assumptions of these models are (1) the reference compound is freely diffusible across the BBB; (2) the drug does not back-diffuse from the brain to the blood; and (3) no metabolism of the compounds occurs before decapitation. The advantage of these pharmacokinetic methods is fast, and many compounds can be assessed in a short time, which is ideal in the high-throughput setting. The major disadvantages are (1) brain extraction only occurs over a limited time, making it difficult to accurately determine the brain uptake index and (2) the driving force for the transport is unknown.
Intracerebral microdialysis involves direct sampling of brain interstitial fluid by a dialysis fiber implanted into the brain parenchyma. The concentration of compound that has permeated into the brain following oral, intravenous, or subcutaneous administration can be monitored over time within the same animal. Any drug that enters the brain interstitial fluid will permeate into the physiological solution within the probe, and the solution may be subsequently assayed by an appropriate technique. The major advantage of this technique is that it provides pharmacokinetic profiles of drugs in the brain without killing animals at different time points. One limitation of this technique is that it greatly relies on and limited by the sensitivity of the assay technique (de Lange et al. 1999), since the solute concentrations may be extremely low in the dialysate. Another major disadvantage is that insertion of the probe can result in chronic BBB disruption.
More recently, various imaging techniques, including positron emission tomography (PET) and magnetic resonance imaging (MRI), have been used to determine BBB permeability in humans. PET is a noninvasive tracer technique used to quantify the BBB extravasation. Magnetic resonance imaging (MRI) is also a noninvasive technique, but it is more qualitative than quantitative. The major disadvantages for these techniques include their inherent costs, labor intensity, relatively low resolution (100 μm to 1 mm per pixel), and inability to differentiate between parent compound and metabolites (Yuan et al. 2009; Nicolazzo et al. 2006).
All of the abovementioned methods only measure certain indexes of relative permeability for the drug uptake to the brain since they cannot determine the driving force for the efflux. Because it is hard to measure the BBB permeability in brain parenchyma, the microvessels in pia dura at the surface of the brain are often used in in vivo BBB permeability study. Although pial microvessels do not have the entire ensheathment of astrocytes as those cerebral microvessels in the parenchyma, the pial and cerebral microvessels appear to have many morphophysiological properties in common. These include ultrastructural characteristics, permeability of cell junctions to electron-dense tracers, trans-endothelium electrical resistance, and molecular properties of endothelium. For these reasons, pial microvessels are often used in the BBB permeability studies (Allt and Lawrenson 1997).
Gaber et al. (2004) suggested a method to measure clearance or leakage of drug out of the pial microvessels rather than “true” permeability of the microvessels to solute. Again, this method cannot determine the driving force, the concentration difference of the test solute across the BBB. The occluding single microvessel measurement is done directly on one single exposed pial microvessel after the frontal craniotomy removing a small section of the skull and the dura mater (Easton and Fraser 1994). This method has well-controlled conditions including known concentration difference across the microvessel wall. However, recent study suggests that the exposed rat pial microvessels become leaky to both small and large molecules within 20–60 min following the craniotomy and the permeability of the exposed microvessels rises sharply after 160 min (Easton et al. 1997).
To quantify the permeability of intact rat pial microvessels and overcome the abovementioned disadvantages, Yuan et al. (2009) developed a noninvasive method, without exposing the cortex, to measure the solute permeability (P) of postcapillary venules on rat pia mater to various-sized solutes. The pial microvessels were observed by a high numerical aperture objective lens through a section of frontoparietal bones thinned with a micro-grinder (revised surgical method from Easton et al. 1997). P was measured on individual pial venular microvessels with the perfused fluorescence tracer solution through the carotid artery by using highly sensitive quantitative fluorescence microscope imaging method. Their results indicate that the solute permeability of rat pial microvessels is about an order of magnitude lower than that of rat mesenteric microvessels, from 1/11 for a small solute, sodium fluorescein, to 1/6 for a large solute, albumin or dextran 70 k. The permeability of rat mesenteric microvessels to these solutes was measured by Fu and Shen (2004). Recently, by employing multiphoton microscopy with longer wavelength lasers for the deeper penetration, the solute permeability of cerebral microvessels in the rat parenchyma was quantified as well as the solute brain tissue diffusion coefficient (Shi et al. 2014a, b, c). Their method is shown in Fig. 4. They found that the BBB permeability to solutes of molecular weight ranging from 4 kDa to 160 kDa, in the cerebral microvessels of rat brain parenchyma 100–250μm below the pia mater, is not significantly different from that of rat pial microvessels for the same-sized solutes except for the small solute, sodium fluorescein (molecular weight 376 kDa) (Yuan et al. 2009). They also found that the ratios of the effective brain tissue diffusion coefficient to the free one are 0.46, 0.19, 0.12, 0.12, 0.11, and 0.11, correspondingly, for sodium fluorescein; dextran 4 k, 20 k, 40 k, and 70 k; and IgG (molecular weight ∼160 kDa).

Quantitative multiphoton fluorescence microscopy for the measurement of the BBB solute permeability (a) and brain tissue diffusion coefficient (b) for a rat cerebral microvessel ∼200 μm below the pia mater. The images were collected during the in vivo experiments, and the fluorescence intensity was analyzed off-line. When the fluorescence-labeled test solute was injected into the carotid artery, the cerebral microvessel lumen is filled with fluorescent solute, producing I 0 in the dashed line enclosed area (ROI) in the left figure of (a). With continued perfusion, the measured fluorescence intensity increased indicating further transport of the solute out of the microvessel and into the surrounding tissue. The initial solute flux into the tissue was measured from the slope (dI/dt)0 of the intensity vs. time line, right figure in (a). The solute permeability P was calculated as P = 1/I 0 ×(dI/dt)0 × r/2. Here r is the microvessel radius. (b) The averaged intensity of the eight dashed lines on the right figure was plotted as a temporal function of the distance from the vessel edge (r t). Matching these lines (green) with the predictions (black) from a mathematical model for unsteady diffusion of a solute (right figure in (b)) can give the solute brain tissue diffusion coefficient D eff. D free shown here is the solute diffusion coefficient in a free solution. Redrawn from (Shi et al. 2014a, b, c)
4.3 In Vitro Blood-Brain Barrier Models
The development of in vitro models for the BBB has enabled the study of transport phenomena at the molecular and cellular levels. The aim of such in vitro BBB models is to functionally resemble as many as possible the unique characteristics of the BBB. Compared with in vivo animal models, the in vitro models are relatively accessible, flexible, reproducible, and abundantly available. Previous investigations showed that the permeability of the in vitro BBB models to various compounds such as sucrose, retinoic acid, retinol, haloperidol, caffeine, and mannitol was comparable to the permeability data obtained from in vivo models (Franke et al. 1999). Recent studies showed that the solute permeability of the in vitro BBB model generated from the cultured bEnd3 (mouse brain microvascular endothelial cell line) or cocultured together with primary astrocytes (Gaillard and de Boer 2000; Brown etal. 2007; Li et al. 2010a, b) is comparable to that of the cerebral microvessels from in vivo studies for the similar-sized solutes except for the small solutes of molecular weight less than 10 kDa (Li et al. 2010a, b; Yuan et al. 2009; Shi et al. 2014a, b, c; Fan and Fu 2016). The in vitro BBB model also showed good expression of essential junction proteins, claudin-5, occludin, ZO-1, and VE-cadherin, while the primary astrocytes used in the coculture expressed GFAP (glial fibrillary acidic protein, a specific protein of astrocytes) (Brown et al. 2007; Li et al. 2010a, b). But the hydraulic conductivity of this in vitro BBB model (Li et al. 2010a, b) is two orders of magnitude higher than that from the in vivo study (Fraser et al. 1990), while trans-endothelial electrical resistance (TEER) is two orders of magnitude lower (Crone and Olesen 1982).
To characterize the transport properties of in vitro BBB models, the solute permeability of the in vitro BBB was determined by measuring the flux of the selected tracer. The most commonly used cell culture substrate consists of a porous membrane support submerged in the culture medium (Transwell apparatus). The Transwell system is characterized by a horizontal side-by-side or vertical diffusion system. During the experiment, the flux of tracers into the abluminal compartment of the Transwell system is recorded as a function of the time, and the solute permeability is calculated from the slope of the flux. The tracers used in the transport experiments are labeled by a fluorescent dye or isotope whose intensity can be measured quantitatively (Li et al. 2010a, b). Another index, trans-endothelial electrical resistance (TEER), or the ionic conductance of the monolayer, is also a measurement of the “tightness” of the in vitro BBB models (Crone and Olesen 1982; Li et al. 2010a, b).
So far, two major types of in vitro BBB models have been developed: endothelial cell monolayer and coculture of endothelial cells with glial cells (the non-nerve cells in the brain). The cells for these models are basically obtained from primary/sub-passaged or immortalized cell cultures. The origins of the cells are also very diverse: human, primate, bovine, porcine, rodent, and murine species.
The brain capillary endothelial cells (BCEC) have been used to establish tissue culture systems ever since the technique of culturing highly purified populations of microvascular cells became available in the early 1980s. The first endothelial monolayers were established using BCEC grown on culture dishes, microcarriers (e.g., dextran beads), and various kinds of filters, including nylon mesh and polycarbonate. These cultured BCEC keep their endothelial phenotypes and provide a simple model for the study of the permeability of the BBB. For instance, they express angiotensin-converting enzyme, von Willebrand factor, and internalized acetylated low-density lipoprotein. However, they were reported to lose many BBB-specific features they possessed in vivo. For instance, they are lack of specific brain endothelial markers γ-glutamyl transpeptidase, marker enzyme alkaline phosphatase (Meyer etal. 1990), and glucose transporter system (Hemmila and Drewes 1993). Moreover, the permeability of the BCEC monolayer to sucrose was reported to be from 10−4 to 10−5 cm/s compared with 10−6 cm/s in vivo. The TEER for endothelial monolayer was also found to be pretty low, from 20 to 1400 Ω cm2, compared with more than 2000 Ω cm2 in vivo. So the BCEC monolayer alone is not a well-characterized model for the BBB. The major reason for this may be the lack of in situ environment and brain-derived signals.
In human body, the BBB are almost completely ensheathed by surrounding tissue, mostly astrocyte foot processes. Experimental results from electron microscopic techniques show that astrocytes do have significant effects on the formation of the unique BBB phenotype of brain endothelial cells (Abbot 2002; Haseloff et al. 2005). They induce formation of the tight junctions between endothelial cells and increase paracellular integrity of the BBB. To better mimic the in vivo BBB, a model with coculture of BCEC and astrocyte was developed. This coculture model was characterized on the basis of specific cell-type properties and specific BBB properties by electron microscopic evaluation and immunohistochemistry methods (Gaillard and de Boer 2000). The results showed that BCEC displayed (1) characteristic endothelial cell morphology, (2) expression of endothelial cell markers (i.e., CD51, CD62P, CD71, and cadherin 5), (3) tight junction formation between the cells, and (4) expression of typical barrier marker γ-glutamyl transpeptidase (γ-GTP) and P-glycoprotein (Pgp) and transferrin receptor. Astrocytes displayed characteristic astrocyte morphology and expressed glial fibrillary acidic protein (GFAP). Transmission electron microscopy showed evidence of tight junction formation between the endothelial cells and few pinocytic vesicles. A 15-fold increase in γ-glutamyl transpeptidase activity was measured in the endothelial cells cocultured with astrocytes (Demeuse et al. 2002). The permeability of the coculture system to several tracers was reported to be lower than the endothelial monolayer. These results indicate that the coculture system is a better model to study the transport across the BBB.
Primary brain capillary endothelial cells have the closest resemblance to the BBB phenotype in vivo and exhibit excellent characteristics of the BBB at early passages (Nicolazzo et al. 2006). They, however, have inherent disadvantages such as being extremely time-consuming and costly to generate, being easily contaminated by other neurovascular unit cells, losing their BBB characteristics over passages, and requiring high technical skills for extraction from brain tissue (Deli et al. 2005; Brown et al. 2007). An immortalized mouse brain endothelial cell line, bEnd3, has recently been under investigation for in vitro BBB models because of its numerous advantages over primary cell culture: the ability to maintain BBB characteristics over many passages, easy growth and low cost, formation of functional barriers, and amenability to numerous molecular interventions (Brown et al. 2007; Soga et al. 2001; Yoder 2002; Yuan et al. 2010a, b; Tyagi et al. 2009). Previous RT-PCR analysis showed that bEnd3 cells express the tight junction proteins ZO-1, ZO-2, occludin, and claudin-5 and junctional adhesion molecules (Brown et al. 2007; Li et al. 2010a, b; Omidi et al. 2003). They also maintained functionality of the sodium- and insulin-dependent stereospecific facilitative transporter GLUT-1 and the P-glycoprotein efflux mechanism (Omidi et al. 2003), formed fairly tight barriers to radiolabeled sucrose, and responded like primary cultures to disrupting stimuli (Brown etal. 2007).
To characterize the transport properties of in vitro BBB models, Malina et al. (2009) (Bowman et al. 1983; Thompson et al. 1994; Salvetti et al. 2002; Karyekar et al. 2003; Hamm et al. 2004; Kemper et al. 2004; Boveri et al. 2005; Kraus et al. 2008; Poller et al. 2008) measured the diffusive permeability of endothelial cell monolayer and coculture of endothelial cells with astrocytes to fluorescence- or isotope-labeled tracers, e.g., sucrose, inulin, and mannitol. Sahagun et al. (1990) reported the ratio between abluminal concentration and luminal concentration of different-sized dextrans (4 k, l0 k, 20 k, 40 k, 70 k, and 150 k) across mouse brain endothelial cells. Gaillard and de Boer (2000) measured the permeability of sodium fluorescein and FITC-labeled dextran 4 k across a coculture of calf brain capillary endothelial cells with rat astrocytes. Many investigators have measured the trans-endothelial electrical resistance (TEER) of brain endothelial monolayers and cocultures as an indicator of ion permeability (Santaguida et al. 2006; de Vries etal. 1996; Cucullo et al. 2002; Zhang et al. 2006).
To seek for in vitro BBB models that are more accessible than animals for investigating drug transport across the BBB, Li et al. (2010a, b) characterized the junction protein expression and quantified the TER and permeability to water (L p) and solutes (P) of four in vitro BBB models: bEnd3 monoculture, bEnd3 coculture with astrocytes, coculture with two BM substitutes (collagen type I and IV mixture), and Matrigel. Collagen type IV network is the basic framework of native BM (Engvall 1995; Miosge 2001), and Matrigel is a soluble and sterile extract of BM derived from the EHS tumor, which has been widely used as a reconstituted BM in studying cell morphogenesis, differentiation, and growth (Kleinman and Martin 2005). Their results show that L p and P of the endothelial monoculture and coculture models are not different from each other. Compared with in vivo permeability data from rat pial microvessels, P of the endothelial monoculture and coculture models are not significantly different from in vivo data for dextran 70 k, but they are 2–4 times higher for small solutes TAMRA and dextran 10 k. This suggests that the endothelial monoculture and all of the coculture models are fairly good models for studying the transport of relatively large solutes (drugs or drug carriers) across the BBB.
4.4 Transport Models for the Paracellular Pathway of the Blood-Brain Barrier
Transport across the BBB include both paracellular and transcellular pathways (Pardridge 2005). While large molecules cross the BBB through transcellular pathways, water and small hydrophilic solutes cross the BBB through the paracellular pathway (Hawkins and Davis 2005). The paracellular pathway of the BBB is formed by the endothelial surface glycocalyx, the tight junction openings, the BM filled with extracellular matrix, and the openings between adjacent astrocyte foot processes (Fig. 1b). In addition to the endothelial tight junctions, the BM and the astrocyte foot processes provide a significant resistance to water and solute transport across the BBB.
The breakdown of the BBB and increased permeability are widely observed in many brain diseases such as stroke, traumatic head injury, brain edema, Alzheimer’s disease, AIDS, brain cancer, meningitis, and hypertension (Beaumont et al. 2000; Dietrich et al. 1994; Fukuda et al. 1995; Baldwin et al. 1996; Cernak et al. 2004; Montagne et al. 2017; Setiadi et al. 2017). Although numerous biochemical factors are found to be responsible for the breakdown of the BBB in disease, the quantitative understanding of how these factors affect the structural components of the BBB to induce BBB leakage is poor. On the other hand, to design therapeutic drugs with better transport properties across the BBB relies greatly on this understanding. Therefore, it is important to investigate how the structural components in the paracellular pathway of the BBB affect its permeability to water and solutes through mathematical modeling.
Extended from a previous three-dimensional model for studying the transport across the peripheral microvessel wall with endothelium only (Fu et al. 1994; Fu and Chen 2003; Fu and Shen 2004), Li etal. (2010a, b) developed a new model for the transport across the BBB, which included the BM and wrapping astrocyte foot processes. The simplified model geometry is shown in Fig. 5. This is the enlarged view for the part near tight junction shown in Fig. 1b. At the luminal side, there is an endothelial surface glycocalyx layer (SGL) with a thickness of L f from 100 to 400 nm under normal physiological conditions (Squire etal. 2001; Arkill et al. 2011, 2012; Schulze and Firth 1992). Between adjacent endothelial cells, there is an inter-endothelial cleft with a length of L ∼500 nm and a width of 2B ∼20 nm (Schulze and Firth 1992; Adamson et al. 2004). In the inter-endothelial cleft, there is a L jun (∼10 nm) thick junction strand with a continuous slit-like opening of width 2B s, which varies depending on the location of the cerebral microvessels (from ∼1 to 10 nm). The distance between the junction strand and luminal front of the cleft is L 1. At the tissue side of the cleft, a BM separates the endothelium and the astrocyte foot processes. The thickness of the BM is 2L b (20–40 nm), and the length of the astrocyte foot processes is 2W a (∼5000 nm). Between adjacent astrocyte foot processes, there is a cleft with length L a (∼1000 nm) and width 2B a (20–2000 nm). The anatomic parameters for the BBB structural components were obtained from the electron microscopy studies in the literature.

Model geometry for the paracellular pathway of the BBB (not in scale) showing the dashed line enclosed region in Fig. 1b. The thickness of the endothelial surface glycocalyx layer is L f. The inter-endothelial cleft has a length of L and a width of 2B. The length of the tight junction strand in the inter-endothelial cleft is L jun. The width of the small continuous slit in the junction strand is 2B s. The distance between the junction strand and luminal front of the cleft is L 1. The width of the basement membrane is 2L b and the length of the astrocyte foot processes is 2W a. The cleft between astrocyte foot processes has a length of L a and a width of 2B a. The surface glycocalyx layer and the endothelial cells are defined as the endothelium only while the BBB is defined to include the endothelium, the basement membrane, and the astrocytes. Redrawn from Li et al. (2010b)
Unlike the peripheral microvessel wall, the endothelium of the BBB has negligible large discontinuous breaks in the junction strand of the inter-endothelial cleft, and the small slit in the junction strand is assumed continuous (Hawkins and Davis 2005). As a result, the cross-sectional BBB geometry is the same along the axial direction (y direction in Fig. 5), and thus the model could be simplified to 2-D (in x, z plane). It could be further simplified to a unidirectional flow in each region due to very narrow clefts and the BM. In addition, the curvatures of the BM and the endothelium can be neglected because their widths are much smaller than the diameter of the microvessel. The fluid flow in the cleft regions of the BBB was approximated by the Poiseuille flow while those in the endothelial SGL and BM by the Darcy and Brinkman flows, respectively. Diffusion equations in each region were solved for the solute transport. After solving for the pressure, water velocity, and solute concentration profiles, the hydraulic conductivity L p and solute permeability P can be calculated.
Figure 6a shows the model predictions for L p as a function of tight junction opening B s when the BM has different fiber densities. K b is Darcy permeability in the BM. When the fiber density in the BM is the same as that in the SGL, K b = 3.16 cm2. The green line in Fig. 6a shows the case of peripheral microvessels with only endothelium. When B s increases from 0.5 to 2 nm, L p will increase by ∼20-fold. In contrast, when the endothelium is wrapped by the BM and the astrocytes as for the BBB, increase in B s from 0.5 to 2 nm only induces fivefold increase in L p when the fiber density in the BM is the same as that in the SGL (dash-dot-dash line). If the fiber density in the BM is ten times of that in the SGL, the increase is only 1.6-fold in L p (solid line), while if the fiber density in the BM is 1/10 of that in the SGL, the increase is 12-fold in L p (dashed~line). Even at a large B s of 5 nm, when the BM is filled with the same density fibers as in the SGL, the BBB permeability is only 17% of that of endothelium only. This percentage can be as low as 2% if the fiber density in the BM is ten times of that in the SGL. Figure 6b shows the model predictions for L p as a function of the endothelial SGL thickness L f. The green line is for the case of endothelium only while the solid line for that of the BBB. We can see the decrease in L f from 400 to 0 nm increases L p by threefold in the case of endothelium only, while in the case of the BBB, the increase is only 25% in L p with the protection of the BM and the astrocytes. Similar results are predicted for the solute permeability (Li et al. 2010a, b). These results indicate that the BM and astrocytes of the BBB provide a great protection to the CNS under both physiological and pathological conditions. However, on the other hand, these unique structures also impede the drug delivery to the brain through the BBB. Most recently, an electro-diffusion model for the blood-brain barrier to the charged molecules was developed to take into consideration of the effect of charges in the SGL and BM on the solute transport across the BBB (Li and Fu 2011).

Model predictions for hydraulic conductivity L p (a) as a function of B s, the half width of the small slit in the junction strand under two cases: when considering transport across the endothelium only (endothelium only, green line) and when considering transport across the entire BBB (BBB). In the BBB case, three different fiber densities were considered for the basement membrane: the same as the fiber density in the surface glycocalyx layer (K b = 3.16 nm2, the dash-dot-dash line), ten times lower (K b = 31.6 nm2, the dashed line) and higher (K b = 0.316 nm2, the solid line); (b) as a function of the surface glycocalyx layer thickness L s. Redrawn from (Li et al. 2010a, b)
5 Modulation of the Blood-Brain Barrier Permeability by Chemical and Physical Stimulations
5.1 Modulation of the BBB Permeability by Chemical Stimuli
During cerebrovascular and neurological diseases, the BBB is compromised, and its permeability can be increased by various types of stimuli including cytokines, inflammatory mediators, metabolites of arachidonic acid, excitatory amino acids, nitric oxide, and bacterial infection. Among cytokines is vascular endothelial growth factor (VEGF), which is upregulated in many brain diseases (Ohlin et al. 2011; Suidan et al. 2010). By using submicron resolution multiphoton fluorescence microscopy with a longer penetration depth into brain parenchyma of rats, Shi et al. (2014b) quantified the temporal VEGF effects on the BBB permeability to various-sized molecules. They found that exposure to 1 nM VEGF transiently increased the BBB permeability to 2.2, 10.5, 9.8, and 12.8 times their control values, for sodium fluorescein, dextran 20 k and 70 k, and IgG, respectively, within 30 s, and all returned to control in 2 min.
Previous studies have shown that elevated intracellular 3,5-cyclic monophosphate (cAMP) levels enhance endothelial junction barriers and thus reduce permeability in peripheral microvessels (Adamson et al. 1998; Fu et al. 2006; Fukuhara et al. 2005; Mehta and Malik 2006; Moore et al. 1998; Sayner 2011) and in cultured BBB models (Balyasnikova et al. 2000; Rubin et al. 1991; Wolburg et al. 1994). Studies on intact microvessels also showed that elevation of cAMP levels abolished the increase in the hydraulic conductivity stimulated by adenosine triphosphate (ATP) in frog and hamster mesenteric microvessels (He and Curry 1993) as well as in the solute permeability induced by VEGF in the same type of microvessels of frogs and rats (Fu et al. 2006, 2015). Administration of a cAMP analogue abolished VEGF-induced rat mesenteric microvessel hyperpermeability (Fu etal. 2015). Employing multiphoton microscopy, Shi et al. (2014b) demonstrated that after 20 min pretreatment of 2 mM cAMP analogue, 8-bromo-cAMP, the initial increase by 1 nM VEGF was completely abolished in the BBB permeability to all solutes. The response pattern of the BBB permeability to VEGF and cAMP and the ratios of the peak to control values for rat cerebral microvessels are similar to those for rat mesenteric (peripheral) microvessels, except the ratios are higher in permeability of cerebral microvessels for the intermediate and large solutes. Their results imply a new approach for delivering large therapeutic agents to the brain.
5.2 Modulation of the BBB Permeability by Physical Stimuli
Among the numerous strategies to deliver therapeutic drugs into the central nervous system, the focused ultrasound (FUS) sonication combined with microbubbles (MBs) has been shown to be effective in transiently disrupting the BBB for noninvasive drug delivery (Hynynen et al. 2006; Chen et al. 2013; Yang et al. 2011; Curley etal. 2017). At appropriate acoustic power density (or pressure), burst repetition rate, duty cycle, and sonication duration and in the presence of MBs with proper materials, sizes, and concentrations, FUS can achieve noninvasive, selective and localized disruption of BBB without visible damage to the brain tissue (Konofagou 2012; Deng 2010). FUS-induced BBB disruption has been widely assessed by using magnetic resonance imaging (MRI) or post-FUS histological examination (Yang et al. 2011: Chu et al. 2013; Park et al. 2010). Indirect MRI collects the images of brain slices in the presence of a MRI contrast agent such as gadolinium to access the BBB disruption in different locations of the brain. Alternatively, post-FUS histological examination relates the BBB disruption to the amount of Evans blue extravasation in the brain tissue after the dye is intravenously injected into the blood circulation before or after sonication. However, the submillimeter spatial resolution of the MRI studies and the postmortem histological examinations only assess the relatively large leakage of the BBB, and these methods are restricted to the exploration of a region that is much larger than a single microvessel with its surrounding tissue. Therefore the BBB disruption assessed by the MRI and histological examination is more qualitative rather than quantitative. To overcome aforementioned limitations, two-photon microscopy was employed to quantitatively access the enhanced permeability of the cerebral vasculature after FUS (Nhan et al. 2013). In addition to micrometer spatial resolution, two-photon microscopy offers the advantage of deep tissue penetration, which is essential for the BBB permeability measurement. By employing the two-photon microscopy, Shi et al. (2014c) quantified the BBB permeability to dextran 155 k with similar molecular weight to an antibody by applying different doses of FUS in the presence of MBs with an optimal size and concentration. After ∼5 min by applying the FUS on the thinned skull in the presence of MBs for 1 min, TRITC-dextran 155 k in 1% BSA mammalian Ringer’s solution was injected into the cerebral circulation via the ipsilateral carotid artery by a syringe pump. Simultaneously, the temporal images were collected from the brain parenchyma ∼100–200 μm below the pia mater. At the optimal dose, permeability increased by ∼14-fold after 5 min post-FUS, and permeability returned to the control level after 25 min. FUS without MBs or MBs injected without FUS did not change the permeability. Their method provides an accurate in vivo assessment for the transient BBB permeability change under the treatment of FUS. The optimal FUS dose found for the reversible BBB permeability increase without BBB disruption is reliable and can be applied to future clinical trials.
Transcranial direct current stimulation (tDCS) is a noninvasive electrical stimulation technique investigated for a broad range of medical and performance indications. Most recently, Shin et al. (2016) found that after 1 mA tDCS treatment for 20 min, the permeability of rat cerebral microvessels 100–200μm below pia mater to sodium fluorescein (molecular weight 376 kDa) and dextran 20 k and 70k increased to about 8-fold, 35-fold, and 35-fold of their respective control values, 10 min post-tDCS treatment, correspondingly. All of the increased BBB permeability returns to the control in 20 min posttreatment. Their method provides another noninvasive approach for delivering therapeutic agents to the brain through the BBB.
6 Drug Delivery Through the Blood-Brain Barrier
A large number of people in the world are now suffering from CNS diseases. The total number of patients with CNS diseases is reported to be larger than that with cardiovascular diseases (Pardridge 2007). While the BBB serves as a natural defense that safeguards the brain against the invasion of various circulating toxins and infected cells, it also provides a significant impediment toward the delivery of diagnostic and therapeutic agents to the brain via the systemic route. Various methods such as intracerebral implantation, microdialysis, convection-enhanced distribution (CED), osmotic shock, and chemical modification of the BBB have been developed for delivering drugs into the brain. However, the applications of these methods are limited, and they can only partially keep with the demands of modern therapies. For instance, the efficiency of intracerebral implantation, microdialysis, and CED methods is low since their major transport mechanisms are diffusion and convection of interstitial fluid. The penetration distances of drugs delivered by the first two methods are reported to be less than 1 mm with simple diffusion (Mak et al. 1995). CED has been shown in laboratory experiments to deliver high-molecular-weight proteins 2 cm from the injection site in the brain parenchyma after 2 h of continuous infusion (Bobo et al. 1994). However, the success of CED relies on precise placement of the catheters and other infusion parameters for delivery into the correct location in the brain parenchyma. For effective treatment of the CNS diseases, therapeutic agents have to reach the specific regions of the brain at an adequate amount. As discussed earlier, due to the abundance and the largest contact area of the BBB for blood-brain exchange, it is more reasonable to develop strategies for drug delivery through the BBB.
The above session for the modulation of the BBB permeability by chemical and physical stimuli discusses potential noninvasive methods for drug delivery through the BBB. Novel nanomaterials-based nanocarriers have also been developed for drug delivery across the BBB (Tsou et al. 2017). More specifically, as shown in Fig. 3, we can directly deliver therapeutic agents through paracellular pathway (route A), through lipophilic diffusion pathway (route C), or through transporters at the BBB by closely mimicking their substrates (route B) or mounting the drugs on the ligands of the specific receptors expressed at the BBB (e.g., low-density lipoprotein receptor-related protein) for transcytosis (Trojan horse approach or receptor-mediated transcytosis, RMT, route D), as well as using cationized proteins, peptides, and nanoparticle carriers for adsorptive-mediated transcytosis (AMT, route E). The following summarizes the delivery strategies through these routes, respectively.
6.1 Delivery Through Paracellular Pathway (Route A)
To increase the hydrophilic drug delivery from the blood to the brain tissue, we can transiently open the barriers in the paracellular pathway of the BBB, e.g., the cleft opening (2B in Fig. 5), the tight junction opening (2B s), and the BM width (2L b), or degrade the fiber matrix in the endothelial surface glycocalyx and in the BM. Osmotic shock by intracarotid administration of a hyperosmotic mannitol causes endothelial cells to shrink and increase 2B, 2B s, and 2L b. Subsequent administration of drugs can increase their concentrations in the brain to a therapeutic level (Kroll and Neuwelt 1998; Doolittle et al. 2002). Physical means such as application of electric and magnetic fields can increase the drug brain uptake. Focused ultrasound, guided by MRI, combined with microbubbles injected into the blood stream has been shown to disrupt the BBB and increase the distribution of Herceptin in brain tissue by 50% in mice (Hynynen et al. 2001, 2006; Kinoshita 2006). Application of inflammatory agents such as bradykinin analogue can open the tight junction of the BBB and increase the drug transport to the brain (Dean et al. 1999; Borlongan and Emerich 2003). However, these approaches are relatively costly and non-patient friendly. They may also enhance tumor dissemination after BBB disruption and damage the neurons by allowing the passage of unwanted blood components into the brain (Gabathuler 2010).
6.2 Delivery Through Lipophilic Diffusion Pathway (Route C)
Some molecules, e.g., alcohol, nicotine, and benzodiazepine, can freely enter the brain through route C in Fig. 3. Their ability to passively (diffusion by concentration differences across the cell membrane) cross the BBB depends on the molecular weight, charge (low hydrogen-bonding capabilities), and lipophilicity (Lipinski 2000). Therefore, if we can modify the drugs through medicinal chemistry, e.g., reduce the relative number of polar groups, or incorporate them with a lipid carrier, we can enhance their brain uptake (Shashoua and Hesse 1996). Modification of antioxidants with pyrrolopyrimidines increases their ability to access target cells in the CNS (Sawada etal. 1999). Covalently attaching 1-methyl-1,4-dihydronicotinate to a hydroxymethyl group can enhance the delivery of ganciclovir (Cytovene, an antiviral medication) to the brain (Bodor etal. 1981; Brewster et al. 1994). However, the modification which helps for the drug delivery to the brain often results in loss of the therapeutic function of a drug. In addition, increase of lipophilicity of a drug can result in making it a substrate for the efflux pump P-glycoprotein (route F in Fig. 3) (Gabathuler 2010).
6.3 Delivery Through Transporter-Mediated Pathway (Route B)
The brain requires tremendous amount of essential substances for survival and function, e.g., glucose, insulin, hormones, low-density lipoprotein (LDL), etc. These nutrients and substances are transported into the brain, not by paracellular or lipophilic diffusion pathway as described earlier but by specific transporters or receptors at the BBB. Drugs can be modified to take advantages of the native BBB nutrient transporter systems or by being conjugated to ligands that recognize receptors expressed at the BBB for the receptor-mediated transcytosis. This physiological approach is by far recognized as the most likely successful drug delivery method to the brain.
Peptides and small molecules may use specific transporters expressed on the luminal and basolateral sides of the endothelial cells to cross into the brain. So far, at least eight different nutrient transporters have been identified to transport a group of nutrients with similar structures. Drugs can be modified to closely mimic the endogenous carrier substrates of these transporters and be transported through the specific transporter-mediated transcytosis. Dopamine can be used to treat Parkinson’s disease, but itself is non-brain penetrant. Instead, dopamine’s metabolic precursor, L-Dopa, if delivered by a neutral amino acid carrier through its transporter at the BBB, shows a clear clinical benefit on patients with Parkinson’s disease (Pardridge 2017). To use a BBB transporter for drug delivery, several important factors must be considered: the kinetics and structural binding requirements of the transporter, therapeutic compound manipulation so that the compound binds but also remains active in vivo, and actual transport of the compound into the brain instead of just binding to the transporter (Gabathuler 2010).
6.4 Delivery Through Receptor-Mediated Pathway (Route D)
Instead of by transporters, larger essential molecules are delivered into the brain by specific receptors highly expressed at the endothelial cells of the BBB. The receptor-mediated transcytosis (RMT) includes three steps: receptor-medicated endocytosis of the molecule at the luminal side of the endothelium (blood side), transport through the endothelial cytoplasm, and exocytosis of the molecule at the abluminal side of the endothelium (brain side). Although the exact mechanisms of RMT have not been well understood, drug delivery targeting three receptors (the insulin receptor, the transferrin receptor, and the LDL receptor) has been developed since the start of this century (Pardridge 2017). More and more receptors have been targeted for the drug delivery since then (Gabathuler 2010). This physiological approach is often called molecular Trojan horse since the therapeutic compounds are conjugated to the specific ligands or the antibodies, which can be recognized and delivered through transcytosis by the specific matching receptors at the endothelial cell membrane. In addition to molecular Trojan horses, drugs can be packaged to liposomes and other nanoparticles coated with targeting molecules such as antibodies to the specific receptors to improve the drug-loading capacity.
Although the Trojan horses for the BBB drug delivery are very promising in delivering large peptides and recombinant proteins such as neurotrophins, enzymes, and monoclonal antibodies (Pardridge 2007), the traffic is limited by the number and carrying capacity of the receptors, as well as by the number of drug molecules that can attach to each antibody (Miller 2002). In addition, Gosk etal. (2004) showed that using anti-transferrin mAb for drug delivery through the systemic administration, although the total amount of the drug in the brain is high, most of it stays associated with brain microvessel endothelial cells instead of in the brain parenchyma. Due to the high affinity of the antibodies, it is a challenge to dissociate from their specific receptors. Furthermore, widespread distribution of the receptors on peripheral organs would limit the specific delivery to the brain, and on the other hand, may induce additional toxicity.
6.5 Delivery Through Adsorptive-Mediated Pathway (Route E)
Adsorptive-mediated transcytosis (AMT) involves endocytosis and exocytosis of charged substances by the endothelial cells of the BBB. Its mechanism is different from that of the RMT, which needs specific matching receptors and ligands. Kumagai et al. (1987) observed that polycationic proteins such as protamine could not only bind to the endothelial cell surface but also penetrate the BBB. Mixing protamine, poly-L-lysine, or other cationic molecules with proteins (e.g., albumin) largely increased the BBB permeability to these proteins. These findings can be explained by AMT triggered by electrostatic interactions between the positively charged proteins and negatively charged membrane regions at the brain endothelium. At normal physiological pH, the luminal surface of the cerebral endothelium and the surrounding BM (see Fig. 1b) carry negative charge (Ueno et al. 2004; Lawrenson et al. 1997) and provide the necessary environment for delivering positively charged drugs and drug carriers. Recently, a quantitative in vivo animal study by Yuan etal. (2010a, b) found that the charge density of the endothelial surface glycocalyx and that of the BM in rat pial microvessels is ∼30 mEq/L. In another in vitro cell culture study, Yuan et al. (2010a, b) found the similar charge density on the surface of a cell monolayer of bEnd3, an immortalized mouse cerebral microvessel endothelial cell line.
To efficiently deliver a therapeutic protein or peptide across the BBB, the simplest way is to cationize the protein or peptide by amidation of its carboxylic acid groups, as well as glutamic and aspartic acid side chain groups with positively charged amines (Hervé et al. 2008). The degree of cationization of a protein or peptide may be critical for its pharmacokinetic fate. Cationization enhances the delivery while induces potential toxicity and immunogenicity of these proteins. PEGylation of cationized molecules can minimize the immunogenicity of these molecules. Positively charged cell-penetrating peptides (CPPs) are often used as the drug carriers for the brain delivery. Commonly used CPPs are penetratin, transportan, Syn-B, and Tat (Hervé et al. 2008). Brain uptake of enkephalin analogue dalargin was enhanced several hundred folds after carried by the CPPs (Rousselle et al. 2003). Decoration of CPPs on the surface of liposome- and biopolymer-based nanoparticles containing drugs has shown to promote their uptake by the brain and entrance to the cytoplasm of neurons (Liu et al. 2008). The drawbacks through AMT are lack of tissue selectivity although the BBB may contain higher concentrations of negative charges than other tissues and possible disruption of the BBB and binding of polycationic substances to the negatively charged plasma proteins and other anionic sites resulting in toxicity (Lockman et al. 2004).
References
Abbott NJ (1992) Comparative physiology of the blood-brain barrier. In: Physiology and pharmacology of the blood-brain barrier. Springer, Heidelberg
Abbott NJ (2002) Astrocyte-endothelial interactions and blood-brain barrier permeability. J Anat 200(6):629
Abbott NJ, Patabendige AK, Dolman DE, Yusof SR, Begley DJ (2010) Structure and function of the blood-brain barrier. Neuorbiol Dis 37:13
Adamson RH, Liu B, Fry GN, Rubin LL, Curry FE (1998) Microvascular permeability and number of tight junctions are modulated by cAMP. Am J Phys 274(6 Pt 2):H1885–H1894
Adamson RH, Lenz JE, Zhang X, Adamson GN, Weinbaum S, Curry FE (2004) Oncotic pressures opposing filtration across non-fenestrated rat microvessels. J Physiol Lond 557:889
Allt G, Lawrenson JG (1997) Is the pial microvessel a good model for blood-brain barrier studies? Brain Res Rev 24:67
Arkill KP, Knupp C, Michel CC, Neal CR, Qvortrup K, Rostgaard J, Squire JM (2011) Similar endothelial glycocalyx structures in microvessels from a range of mammalian tissues: evidence for a common filtering mechanism? Biophys J 101(5):1046–1056
Arkill KP, Neal CR, Mantell JM, Michel CC, Qvortrup K, Rostgaard J, Bates DO, Knupp C, Squire JM (2012) 3D reconstruction of the glycocalyx structure in mammalian capillaries using electron tomography. Microcirculation 19(4):343–351
Aspelund A, Antila S, Proulx ST, Karlsen TV, Karaman S, Detmar M, Wiig H, Alitalo K (2015) A dural lymphatic vascular system that drains brain interstitial fluid and macromolecules. J Exp Med 212(7):991. https://doi.org/10.1084/jem.20142290
Bakker Erik NTP, Bacskai BJ, Arbel-Ornath M, Aldea R, Bedussi B, Morris AWJ, Weller RO, Carare RO (2016) Lymphatic clearance of the brain: perivascular, paravascular and significance for neurodegenerative diseases. Cell Mol Neurobiol 36:181–194
Baldwin SA, Fugaccia I, Brown DR, Brown LV, Scheff SW (1996) Blood-brain barrier breach following cortical contusion in the rat. J Neurosurg 85(3):476
Ballabh P, Braun A, Nedergaard M (2004) The blood-brain barrier: an overview: structure, regulation, and clinical implications. Neurobiol Dis 16:1
Balyasnikova IV, Pelligrino DA, Greenwood J, Adamson P, Dragon S, Raza H, Galea E (2000) Cyclic adenosine monophosphate regulates the expression of the intercellular adhesion molecule and the inducible nitric oxide synthase in brain endothelial cells. J Cereb Blood Flow Metab 20(4):688–699
Beaumont A, Marmarou A, Hayasaki K, Barzo P, Fatouros P, Corwin F, Marmarou C, Dunbar J (2000) The permissive nature of blood brain barrier (BBB) opening in edema formation following traumatic brain injury. Acta Neurochir Suppl 76:125
Begley DJ (2007) Structure and function of the blood-brain barrier. In: Enhancement in drug delivery. CRC Press, Boca Raton, p 575
Bobo RH, Laske DW, Akbasak A, Morrison PF, Dedrick RL, Oldfield EH (1994) Convection-enhanced delivery of macromolecules in the brain. Proc Natl Acad Sci U S A 91(6):2076
Bodor N, Farag HH, Brewster ME (1981) Site-specific, sustained release of drugs to the brain. Science 214(4527):1370
Borlongan CV, Emerich DF (2003) Facilitation of drug entry into the CNS via transient permeation of blood brain barrier: laboratory and preliminary clinical evidence from bradykinin receptor agonist, Cereport. Brain Res Bull 60(3):297
Boveri M, Berezowski V, Price A, Slupek S, Lenfant AM, Benaud C, Hartung T, Cecchelli R, Prieto P, Dehouck MP (2005) Induction of blood-brain barrier properties in cultured brain capillary endothelial cells: comparison between primary glial cells and C6 cell line. Glia 51(3):187
Bowman PD, Ennis SR, Rarey KE, Betz AL, Goldstein GW (1983) Brain microvessel endothelial cells in tissue culture: a model for study of blood-brain barrier permeability. Ann Neurol 14(4):396
Brewster ME, Raghavan K, Pop E, Bodor N (1994) Enhanced delivery of ganciclovir to the brain through the use of redox targeting. Antimicrob Agents Chemother 38(4):817
Brinker T, Stopa E, Morrison J, Klinge P (2014) A new look at cerebrospinal fluid circulation. Fluids Barriers CNS 11:10
Brown PD, Davies SL, Speake T, Millar ID (2004a) Molecular mechanisms of cerebrospinal fluid production. Neuroscience 129:957
Brown RC, Egleton RD, Davis TP (2004b) Mannitol opening of the blood–brain barrier: regional variation in the permeability of sucrose, but not 86Rb+ or albumin. Brain Res 1014(1–2):221–227
Brown RC, Morris AP, O'Neil RG (2007) Tight junction protein expression and barrier properties of immortalized mouse brain microvessel endothelial cells. Brain Res 1130(1):17
Butt AM, Jones HC, Abbott NJ (1990) Electrical resistance across the blood-brain barrier in anaesthetized rats: a developmental study. J Physiol 429:47
Carare R, Bernardes-Silva M, Newman T, Page A, Nicoll J, Perry V, Weller R (2008) Solutes, but not cells, drain from the brain parenchyma along basement membranes of capillaries and arteries: significance for cerebral amyloid angiopathy and neuroimmunology. Neuropathol Appl Neurobiol 34(2):131–144
Cernak I, Vink R, Zapple DN, Cruz MI, Ahmed F, Chang T, Fricke ST, Faden AI (2004) The pathobiology of moderate diffuse traumatic brain injury as identified using a new experimental model of injury in rats. Neurobiol Dis 17(1):29
Chen CC, Sheeran PS, Wu SY, Olumolade OO, Dayton PA, Konofagou EE (2013) Targeted drug delivery with focused ultrasound-induced blood–brain barrier opening using acoustically-activated nanodroplets. J Control Release 172(3):795–804
Chu PC, Chai WY, Hsieh HY et al (2013) Pharmacodynamic analysis of magnetic resonance imaging-monitored focused ultrasound-induced blood–brain barrier opening for drug delivery to brain tumors. Biomed Res Int 2013:627496
Cornford EM, Young D, Paxton JW, Sofia RD (1992) Blood-brain barrier penetration of felbamate. Epilepsia 33:944
Crone C, Olesen SP (1982) Electrical resistance of brain microvascular endothelium. Brain Res 241(1):49–55
Cucullo L, McAllister MS, Kight K, Krizanac-Bengez L, Marroni M, Mayberg MR, Stanness KA, Janigro D (2002) A new dynamic in vitro model for the multidimensional study of astrocyte-endothelial cell interactions at the blood-brain barrier. Brain Res 951(2): 243
Curley CT, Sheybani ND, Bullock TN, Price RJ (2017) Focused ultrasound immunotherapy for central nervous system pathologies: challenges and opportunities. Theranostics 7(15):3608–3623
Curry FE (1983) Mechanics and thermodynamics of transcapillary exchange. In: Handbook of physiology. The cardiovascular system. The American Physiology Society, Bethesda
Dean RL, Emerich DF, Hasler BP, Bartus RT (1999) Cereport (RMP-7) increases carboplatin levels in brain tumors after pretreatment with dexamethasone. Neuro-Oncology 1(4):268
Deli MA, Abraham CS, Kataoka Y, Niwa M (2005) Permeability studies on in vitro blood-brain barrier models: physiology, pathology, and pharmacology. Cell Mol Neurobiol 25(1):59
Demeuse P, Kerkhofs A, Struys-Ponsar C, Knoops B, Remacle C, de Aguilar PV (2002) Compartmentalized coculture of rat brain endothelial cells and astrocytes: a syngenic model to study the blood-brain barrier. J Neurosci Methods 121(1):21
Deng CX (2010) Targeted drug delivery across the blood–brainbarrierusing ultrasound technique. Ther Deliv 1(6):819–848
Dietrich WD, Alonso O, Halley M (1994) Early microvascular and neuronal consequences of traumatic brain injury: a light and electron microscopic study in rats. J Neurotrauma 11(3):289
Doolittle ND, Abrey LE, Ferrari N, Hall WA, Laws ER, McLendon RE, Muldoon LL, Peereboom D, Peterson DR, Reynolds CP, Senter P, Neuwelt EA (2002) Targeted delivery in primary and metastatic brain tumors: summary report of the seventh annual meeting of the blood-brain barrier disruption consortium. Clin Cancer Res 8(6):1702
Easton AS, Fraser PA (1994) Variable restriction of albumin diffusion across inflamed cerebral microvessels of the anaesthetized rat. J Physiol 475:147
Easton AS, Sarker MH, Fraser PA (1997) Two components of blood-brain barrier disruption in the rat. J Physiol 503(3):613
Ehrlich P (1885) Das sauerstufbudurfnis des organismus. Hireschwald, Berlin
Elsinga PH, Hendrikse NH, Bart J, Vaalburg W, van Waarde A (2004) PET studies on P-glycoprotein function in the blood-brain barrier: how it affects uptake and binding of drugs within the CNS. Curr Pharm Des 10:1493
Engelhardt B, Coisne C (2011) Fluids and barriers of the CNS establish immune privilege by confining immune surveillance to a two-walled castle moat surrounding the CNS castle. Fluids Barriers CNS 8(4):10–1186
Engvall E (1995) Structure and function of basement membranes. Int J Dev Biol 39(5):781
Fan J, Fu BM (2016) Quantification of malignant breast cancer cell MDA-MB-231 transmigration across brain and lung microvascular endothelium. Ann Biomed Eng 44(7):2189–2201
Farkas E, Luiten PG (2001) Cerebral microvascular pathology in aging and Alzheimer's disease. Prog Neurobiol 64(6):575
Franke H, Galla HJ, Beuckmann CT (1999) An improved low-permeability in vitro-model of the blood-brain barrier: transport studies on retinoids, sucrose, haloperidol, caffeine and mannitol. Brain Res 818(1):65
Fraser PA, Dallas AD, Davies S (1990) Measurement of filtration coefficient in single cerebral microvessels of the frog. J Physiol 423:343–361
Fu BM, Chen B (2003) A model for the structural mechanisms in the regulation of microvessel permeability by junction strands. ASME J Biomech Eng 125:620
Fu BM, Shen S (2004) Acute VEGF effect on solute permeability of mammalian microvessels in vivo. Microvasc Res 68:51
Fu BM, Tsay R, Curry FE, Weinbaum S (1994) A junction-orifice-entrance layer model for capillary permeability: application to frog mesenteric capillaries. ASME J Biomech Eng 116:502
Fu BM, Shen S, Chen B (2006) Structural mechanisms in the abolishment of VEGF-induced microvascular hyperpermeability by cAMP. J Biomech Eng 128(3):317–328
Fu BM, Yang J, Cai B, Fan J, Zhang L, Zeng M (2015) Reinforcing endothelial junctions prevents microvessel permeability increase and tumor cell adhesion in microvessels in vivo. Sci Rep 5:15697
Fukuda K, Tanno H, Okimura Y, Nakamura M, Yamaura A (1995) The blood-brain barrier disruption to circulating proteins in the early period after fluid percussion brain injury in rats. J Neurotrauma 12(3):315
Fukuhara S, Sakurai A, Sano H, Yamagishi A, Somekawa S, Takakura N, Saito Y, Kangawa K, Mochizuki N (2005) Cyclic AMP potentiates vascular endothelial cadherin-mediated cell-cell contact to enhance endothelial barrier function through an Epac-Rap1 signaling pathway. Mol Cell Biol 25(1):136–146
Gabathuler R (2010) Approaches to transport therapeutic drugs across the blood-brain barrier to treat brain diseases. Neurobiol Dis 37(1):48
Gaber MW, Yuan H, Killmar JT, Naimark MD, Kiani MF, Merchant TE (2004) An intravital microscopy study of radiation-induced changes in permeability and leukocyte-endothelial cell interactions in the microvessels of the rat pia mater and cremaster muscle. Brain Res Protocol 13:1
Gaillard PJ, de Boer AG (2000) Relationship between permeability status of the blood-brain barrier and in vitro permeability coefficient of a drug. Eur J Pharm Sci 12(2):95
Goldmann E (1913) Vitalfarbung am zentralnervensystem. Abhandl Kongil preuss Akad Wiss, vol 1, pp. 1
Gosk S, Vermehren C, Storm G, Moos T (2004) Targeting anti-transferrin receptor antibody (OX26) and OX26-conjugated liposomes to brain capillary endothelial cells using in situ perfusion. J Cereb Blood Flow Metab 24(11):1193
Greenwood J, Hammarlund-Udenaes M, Jones HC, Stitt AW, Vandenbrouke RE, Romero IA, Campbell M, Fricker G, Brodin B, Manninga H, Gaillard PJ, Schwaninger M, Webster C, Wicher KB, Khrestchatisky M (2017) Current research into brain barriers and the delivery of therapeutics for neurological diseases: a report on CNS barrier congress London, UK, 2017. Fluids Barriers CNS 14(1):31
Haeren RH, van de Ven SE, van Zandvoort MA, Vink H, van Overbeeke JJ, Hoogland G, Rijkers K (2016) Assessment and imaging of the cerebrovascular glycocalyx. Curr Neurovasc Res 13(3):249–260
Hamm S, Dehouck B, Kraus J, Wolburg-Buchholz K, Wolburg H, Risau W, Cecchelli R, Engelhardt B, Dehouck MP (2004) Astrocyte mediated modulation of blood-brain barrier permeability does not correlate with a loss of tight junction proteins from the cellular contacts. Cell Tissue Res 315(2):157
Haseloff RF, Blasig IE, Bauer HC, Bauer H (2005) In search of the astrocytic factor(s) modulating blood-brain barrier functions in brain capillary endothelial cells in vitro. Cell Mol Neurobiol 25(1):25
Hawkins BT, Davis TP (2005) The blood-brain barrier/neurovascular unit in health and disease. Pharmacol Rev 57:173
He P, Curry FE (1993) Differential actions of cAMP on endothelial [Ca2+]i and permeability in microvessels exposed to ATP. Am J Phys 265(3 Pt 2): H1019–H1023
Hemmila JM, Drewes LR (1993) Glucose transporter (GLUT1) expression by canine brain microvessel endothelial cells in culture: an immunocytochemical study. Adv Exp Med Biol 331:13
Hervé F, Ghinea N, Scherrmann JM (2008) CNS delivery via adsorptive transcytosis. AAPS J 10(3):455
Hynynen K, McDannold N, Vykhodtseva N, Jolesz FA (2001) Noninvasive MR imaging-guided focal opening of the blood-brain barrier in rabbits. Radiology 220(3):640
Hynynen K, McDannold N, Vykhodtseva N, Raymond S, Weissleder R, Jolesz FA, Sheikov N (2006) Focal disruption of the blood-brain barrier due to 260-kHz ultrasound bursts: a method for molecular imaging and targeted drug delivery. J Neurosurg 105(3):445
Iliff JJ, Wang M, Zeppenfeld DM, Venkataraman A, Plog BA, Liao Y, Deane R, Nedergaard M (2013) Cerebral arterial pulsation drives paravascular CSF-interstitial fluid exchange in the murine brain. J Neurosci 33(46):18190–18199
Karyekar CS, Fasano A, Raje S, Lu RL, Dowling TC, Eddington ND (2003) Zonula occludens toxin increases the permeability of molecular weight markers and chemotherapeutic agents across the bovine brain microvessel endothelial cells. J Pharm Sci 92(2):414
Kay GG (2000) The effects of antihistamines on cognition and performance. J Allergy Clin Immunol 105:S622
Kemper EM, Boogerd W, Thuis I, Beijnen JH, van Tellingen O (2004) Modulation of the blood-brain barrier in oncology: therapeutic opportunities for the treatment of brain tumours? Cancer Treat Rev 30(5):415
Kim JH, Park JA, Lee SW, Kim WJ, Yu YS, Kim KW (2006) Blood-neural barrier: intercellular communication at glio-vascular interface. J Biochem Mol Biol 39(4):339
Kinoshita M (2006) Targeted drug delivery to the brain using focused ultrasound. Top Magn Reson Imaging 17(3):209
Kleinman HK, Martin GR (2005) Matrigel: basement membrane matrix with biological activity. Semin Cancer Biol 15(5):378
Konofagou EE (2012) Optimization of the ultrasound-induced blood–brain barrier opening. Theranostics 2(12):1223–1237
Kraus J, Voigt K, Schuller AM, Scholz M, Kim KS, Schilling M, Schabitz WR, Oschmann P, Engelhardt B (2008) Interferon-beta stabilizes barrier characteristics of the blood-brain barrier in four different species in vitro. Mult Scler 14(6):843
Kroll RA, Neuwelt EA (1998) Outwitting the blood-brain barrier for therapeutic purposes: osmotic opening and other means. Neurosurgery 42(5):1083
Kumagai AK, Eisenberg JB, Pardridge WM (1987) Absorptive-mediated endocytosis of cationized albumin and a beta-endorphin-cationized albumin chimeric peptide by isolated brain capillaries.Model system of blood-brain barrier transport. J Biol Chem 262(31):15214
de Lange EC, de Boer BA, Breimer DD (1999) Microdialysis for pharmacokinetic analysis of drug transport to the brain. Adv Drug Deliv Rev 36:211
Lawrenson JG, Reid AR, Allt G (1997) Molecular characteristics of pial microvessels of the rat optic nerve. Can pial microvessels be used as a model for the blood-brain barrier? Cell Tissue Res 288:259–265
Lewandowsky M (1900) Zur lehre von der cerebrospinalflussigkeit. Z Klin Med 40:480
Li G, Fu BM (2011) An electro-diffusion model for the blood-brain barrier permeability to charged molecule. ASME J Biomech Eng 133(2):0210
Li G, Simon M, Shi Z, Cancel L, Tarbell JM, Morrison B, Fu BM (2010a) Permeability of endothelial and astrocyte cocultures: in vitro blood-brain barrier models for drug delivery. Ann Biomed Eng 38(8):2499
Li G, Yuan W, Fu BM (2010b) A model for water and solute transport across the blood-brain barrier. J Biomech 43(11):2133
Lipinski CA (2000) Drug-like properties and the causes of poor solubility and poor permeability. J Pharmacol Toxicol Methods 44(1):235
Liu L, Guo K, Lu J, Venkatraman SS, Luo D, Ng KC, Ling EA, Moochhala S, Yang YY (2008) Biologically active core/shell nanoparticles self-assembled from cholesterol-terminated PEG-TAT for drug delivery across the blood-brain barrier. Biomaterials 29(10):1509
Lockman PR, Koziara JM, Mumper RJ, Allen DD (2004) Nanoparticle surface charges alter blood-brain barrier integrity and permeability. J Drug Target 12(9–10):635
Louveau A, Smirnov I, Keyes TJ, Eccles JD, Rouhani SJ, Peske JD, Derecki NC, Castle D, Mandell JW, Lee KS (2015) Structural and functional features of central nervous system lymphatic vessels. Nature 523:337–341. https://doi.org/10.1038/nature14432
Mak M, Fung L, Strasser JF, Saltzman WM (1995) Distribution of drugs following controlled delivery to the brain interstitium. J Neuro-Oncol 26(2):91
Malina KC, Cooper I, Teichberg VI (2009) Closing the gap between the in-vivo and in-vitro blood-brain barrier tightness. Brain Res 1284:12
Mehta D, Malik AB (2006) Signaling mechanisms regulating endothelial permeability. Physiol Rev 86(1):279–367
Meyer J, Mischeck U, Veyhl M, Henzel K, Galla HJ (1990) Blood-brain barrier characteristic enzymatic properties in cultured brain capillary endothelial cells. Brain Res 514(2):305
Miller G (2002) Drug targeting. Breaking down barriers. Science 297(5584):1116
Miosge N (2001) The ultrastructural composition of basement membranes in vivo. Histol Histopathol 16(4):1239
Montagne A, Zhao Z, Zlokovic BV (2017) Alzheimer's disease: a matter of blood-brain barrier dysfunction? J Exp Med 214(11):3151–3169
Moody DM (2006) The blood-brain barrier and blood-cerebral spinal fluid barrier. Semin Cardiothorac Vasc Anesth 10(2):128
Moore TM, Chetham PM, Kelly JJ, Stevens T (1998) Signal transduction and regulation of lung endothelial cell permeability. Interaction between calcium and cAMP. Am J Phys 275(2 Pt 1):L203–L222
Nag S, Begley DJ (2005) Blood-brain barrier, exchange of metabolites and gases. In: Pathology and genetics. Cerebrovascular diseases. ISN Neuropath. Press, Basel
Neuwelt EA (2004) Mechanisms of disease: the blood-brain barrier. Neurosurgery 54(1):131
Nhan T, Burgess A, Cho EE, Stefanovic B, Lilge L, Hynynen K (2013) Drug delivery to the brain by focused ultrasound induced blood–brain barrier disruption: quantitative evaluation of enhanced permeability of cerebral vasculature using two-photon microscopy. J Control Release 172(1):274–280
Nicolazzo JA, Charman SA, Charman WN (2006) Methods to assess drug permeability across the blood-brain barrier. J Pharm Pharmacol 58:281
Ohlin KE, Francardo V, Lindgren HS, Sillivan SE, O'Sullivan SS, Luksik AS, Vassoler FM, Lees AJ, Konradi C, Cenci MA (2011) Vascular endothelial growth factor is upregulated by L-dopa in the parkinsonian brain: implications for the development of dyskinesia. Brain 134(Pt 8):2339–2357
Oldendorf WH, Cornford ME, Brown WJ (1977) The large apparent work capability of the blood-brain barrier: a study of the mitochondrial content of capillary endothelial cells in brain and other tissues of the rat. Ann Neurol 1:409
Omidi Y, Campbell L, Barar J, Connell D, Akhtar S, Gumbleton M (2003) Evaluation of the immortalised mouse brain capillary endothelial cell line, bEnd3, as an in vitro blood-brain barrier model for drug uptake and transport studies. Brain Res 990(1–2):95
Pardridge WM (1998) CNS drug design based on principles of blood-brain barrier transport. J Neurochem 70:1781
Pardridge WM (2005) Molecular biology of the blood-brain barrier. Mol Biotechnol 30(1):57
Pardridge WM (2007) Drug targeting to the brain. Pharm Res 24:1733
Pardridge WM (2017) Delivery of biologics across the blood-brain barrier with molecular trojan horse technology. BioDrugs 31(6):503–519
Park J, Fan Z, Kumon RE, El-Sayed ME, Deng CX (2010) Modulation of intracellular Ca2+ concentration in brain microvascular endothelial cells in vitro by acoustic cavitation. Ultrasound Med Biol 36(7):1176–1187
Poller B, Gutmann H, Krahenbuhl S, Weksler B, Romero I, Couraud PO, Tuffin G, Drewe J, Huwyler J (2008) The human brain endothelial cell line hCMEC/D3 as a human blood-brain barrier model for drug transport studies. J Neurochem 107(5):1358
Redzic ZB, Segal MB (2004) The structure of the choroid plexus and the physiology of the choroid plexus epithelium. Adv Drug Deliv Rev 56:1695
Reese TS, Karnovsky MJ (1967) Fine structural localization of a blood-brain barrier to exogenous peroxidase. J Cell Biol 34(1):207
Romero IA, Radewicz K, Jubin E, Michel CC, Greenwood J, Couraud PO, Adamson P (2003) Changes in cytoskeletal and tight junctional proteins correlate with decreased permeability induced by dexamethasone in cultured rat brain endothelial cells. Neurosci Lett 344(2):112
Rousselle C, Clair P, Smirnova M, Kolesnikov Y, Pasternak GW, Gac-Breton S, Rees AR, Scherrmann JM, Temsamani J (2003) Improved brain uptake and pharmacological activity of dalargin using a peptide-vector-mediated strategy. J Pharmacol Exp Ther 306(1): 371
Rubin LL, Hall DE, Porter S, Barbu K, Cannon C, Horner HC, Janatpour M, Liaw CW, Manning K, Morales J, Tanner LI, Tomaselli KJ, Bard F (1991) A cell-culture model of the blood-brain-barrier. J Cell Biol 115(6):1725–1735
Sahagun G, Moore SA, Hart MN (1990) Permeability of neutral vs. anionic dextrans in cultured brain microvascular endothelium. Am J Phys 259(1 Pt 2):H162
Salvetti F, Cecchetti P, Janigro D, Lucacchini A, Benzi L, Martini C (2002) Insulin permeability across an in vitro dynamic model of endothelium. Pharm Res 19(4):445
Santaguida S, Janigro D, Hossain M, Oby E, Rapp E, Cucullo L (2006) Side by side comparison between dynamic versus static models of blood-brain barrier in vitro: a permeability study. Brain Res 1109:1
Sawada GA, Williams LR, Lutzke BS, Raub TJ (1999) Novel, highly lipophilic antioxidants readily diffuse across the blood-brain barrier and access intracellular sites. J Pharmacol Exp Ther 288(3):1327
Sawchuk RJ, Elmquist WF (2000) Microdialysis in the study of drug transporters in the CNS. Adv Drug Deliv Rev 45:295
Sayner SL (2011) Emerging themes of cAMP regulation of the pulmonary endothelial barrier. Am J Physiol Lung Cell Mol Physiol 300(5):L667–L678
Schuetz EG, Schinkel AH, Relling MV, Schuetz JD (1996) P-glycoprotein: a major determinant of rifampicin-inducible expression of cytochrome P4503A in mice and humans. Proc Natl Acad Sci U S A 93:4001
Schulze C, Firth JA (1992) Interendothelial junctions during blood-brain-barrier development in the rat - morphological-changes at the level of individual tight junctional contacts. Dev Brain Res 69(1):85
Setiadi A, Korim WS, Elsaafien K, Yao ST (2017) The role of the blood-brain barrier in hypertension. Exp Physiol 103(3):337–342
Shashoua VE, Hesse GW (1996) N-docosahexaenoyl, 3 hydroxytyramine: a dopaminergic compound that penetrates the blood-brain barrier and suppresses appetite. Life Sci 58(16):1347
Shi L, Zeng M, Sun Y, Fu BM (2014a) Quantification of blood-brain barrier solute permeability and brain transport by multiphoton microscopy. J Biomech Eng 136(3):031005
Shi L, Zeng M, Fu BM (2014b) Temporal effects of vascular endothelial growth factor and 3,5-cyclic monophosphate on blood-brain barrier solute permeability in vivo. J Neurosci Res 92(12):1678–1689
Shi L, Palacio-Mancheno P, Badami J, Shin DW, Zeng M, Cardoso L, Tu R, Fu BM (2014c) Quantification of transient increase of the blood-brain barrier permeability to macromolecules by optimized focused ultrasound combined with microbubbles. Int J Nanomedicine 18(9):4437–4448
Shimizu S (2008) A novel approach to the diagnosis and management of meralgia paresthetica. Neurosurgery 63(4):E820
DW Shin, N Khadka, J Fan, M Bikson, BM Fu (2016) Transcranial direct current stimulation transiently increases the blood-brain barrier solute permeability in vivo SPIE Medical Imaging Conference, Feb. 27-March 3, 2016, San Diego, CA, USA
Simard M, Arcuino G, Takano T, Liu QS, Nedergaard M (2003) Signaling at the gliovascular interface. J Neurosci 23(27):9254
Smith QR (2000) Transport of glutamate and other amino acids at the blood-brain barrier. J Nutr 130:1016
Soga N, Connolly JO, Chellaiah M, Kawamura J, Hruska KA (2001) Rac regulates vascular endothelial growth factor stimulated motility. Cell Commun Adhes 8(1):1
Squire JM, Chew M, Nneji G, Neal C, Barry J, Michel CC (2001) Quasi-periodic substructure in the microvessel endothelial glycocalyx: a possible explanation for molecular filtering? J Struct Biol 136(3):239
Suidan GL, Dickerson JW, Chen Y, McDole JR, Tripathi P, Pirko I, Seroogy KB, Johnson AJ (2010 Jan 15) CD8 T cell-initiated vascular endothelial growth factor expression promotes central nervous system vascular permeability under neuroinflammatory conditions. J Immunol 184(2):1031–1040
Thompson SE, Cavitt J, Audus KL (1994) Leucine-enkephalin effects on paracellular and transcellular permeation pathways across brain microvessel endothelial-cell monolayers. J Cardiovasc Pharmacol 24(5):818
Tsou YH, Zhang XQ, Zhu H, Syed S, Xu X (2017) Drug delivery to the brain across the blood-brain barrier using nanomaterials. Small
Tyagi N, Moshal KS, Sen U, Vacek TP, Kumar M, Hughes WM Jr, Kundu S, Tyagi SC (2009) H2S protects against methionine-induced oxidative stress in brain endothelial cells. Antioxid Redox Signal 11(1):25
Ueno M, Sakamoto H, Liao YJ, Onodera M, Huang CL, Miyanaka H, Nakagawa T (2004) Blood-brain barrier disruption in the hypothalamus of young adult spontaneously hypertensive rats. Histochem Cell Biol 122(2):131
de Vries HE, BlomRoosemalen MCM, van Oosten M, de Boer AG, van Berkel TJ, Breimer DD, Kuiper J (1996) The influence of cytokines on the integrity of the blood-brain barrier in vitro. J Neuroimmunol 64(1):37
Wang R, Ashwal S, Tone B, Tian HR, Badaut J, Rasmussen A, Obenaus A (2007) Albumin reduces blood-brain barrier permeability but does not alter infarct size in a rat model of neonatal stroke. Pediatr Res 62:261
Wolburg H, Neuhaus J, Kniesel U, Krauss B, Schmid EM, Ocalan M, Farrell C, Risau W (1994) Modulation of tight junction structure in blood-brain barrier endothelial cells. Effects of tissue culture, second messengers and cocultured astrocytes. J Cell Sci 107(Pt 5):1347–1357
Wolburg-Buchholz K, Mack AF, Steiner E, Pfeiffer F, Engelhardt B, Wolburg H (2009) Loss of astrocyte polarity marks blood-brain barrier impairment during experimental autoimmune encephalomyelitis. Acta Neuropathol 18(2):219
Yang FY, Lin YS, Kang KH, Chao TK (2011) Reversible blood–brain barrier disruption by repeated transcranial focused ultrasound allows enhanced extravasation. J Control Release 150(1):111–116
Yoder EJ (2002) Modifications in astrocyte morphology and calcium signaling induced by a brain capillary endothelial cell line. Glia 38(2):137
Yoon JH, Lee ES, Jeong Y (2017) In vivo imaging of the cerebral endothelial glycocalyx in mice. J Vasc Res 54(2):59–67. https://doi.org/10.1159/000457799 Epub 2017 Apr 1
Yuan W, Lv Y, Zeng M, Fu BM (2009) Non-invasive measurement of solute permeability in cerebral microvessels of the rat. Microvasc Res 77:166
Yuan W, Li G, Fu BM (2010a) Effect of surface charge of immortalized mouse cerebral endothelial cell monolayer on transport of charged solutes. Ann Biomed Eng 38(4):1463
Yuan W, Li G, Zeng M, Fu BM (2010b) Modulation of the blood-brain barrier permeability by plasma glycoprotein orosomucoid. Microvasc Res 80(1):148–157
Zhang Y, Pardridge WM (2001) Rapid transferrin efflux from brain to blood across the blood-brain barrier. J Neurochem 76:1597
Zhang Y, Li CS, Ye YY, Johnson K, Poe J, Johnson S, Bobrowski W, Garrido R, Madhu C (2006) Porcine brain microvessel endothelial cells as an in vitro model to predict in vivo blood-brain barrier permeability. Drug Metab Dispos 34(11):1935
Zhang Q, Fu BM, Zhang ZJ (2017) Borneol, a novel agent that improves central nervous system drug delivery by enhancing blood-brain barrier permeability. Drug Deliv 24(1):1037–1044
Zlokovic BV, Begley DJ, Djuricic BM, Mitrovic DM (1986) Measurement of solute transport across the blood-brain barrier in the perfused Guinea pig brain: method and application to N-methyl-alpha-aminoisobutyric acid. J Neurochem 46:1444
Acknowledgment
The author would like to thank the funding support from the National Institutes of Health RO1NS101362-01 and U54CA132378-09.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Springer International Publishing AG, part of Springer Nature
About this chapter

Cite this chapter
Fu, B.M. (2018). Transport Across the Blood-Brain Barrier. In: Fu, B., Wright, N. (eds) Molecular, Cellular, and Tissue Engineering of the Vascular System. Advances in Experimental Medicine and Biology, vol 1097. Springer, Cham. https://doi.org/10.1007/978-3-319-96445-4_13
Download citation
DOI: https://doi.org/10.1007/978-3-319-96445-4_13
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-96444-7
Online ISBN: 978-3-319-96445-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)