
Abstract
The first organ which is formed and began to function during the embryonic periods is the heart. In fact, when the embryo could no more support its nutritional requirements just by the simple diffusion from the placenta, the heart appears as a new organ.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others



Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
Establishing Cardiac Crescent
The first organ which is formed and began to function during the embryonic periods is the heart. In fact, when the embryo could no more support its nutritional requirements just by the simple diffusion from the placenta, the heart appears as a new organ.
From epiblast, the primitive streak comes out; however, the primitive streak, in turn, gives origin to mesoderm. Myocardial progenitor cells come out of anterior splanchnic mesoderm. As described by Abu-Issa and Kirby, these myocardial progenitor cells start their development process through four “sequential but at times, overlapping” stages (Abu-Issa and Kirby 2007; Lin et al. 2012):
-
1.
Cardiogenic mesoderm specification
-
2.
Bilateral establishment of heart fields
-
3.
Composition and configuration of heart field
-
4.
Differentiation of cardiomyocyte and formation of heart tube
The heart as noted is one of the organs originating from anterior splanchnic mesoderm. Also, parts of the endoderm are involved in the development of the heart; these endodermal parts are enveloped by mesoderm to form the endodermal portions of the heart (Lin et al. 2012).
The cardiac progenitor cells develop locally into the splanchnic layers of the lateral mesoderm on both left and right sides of the embryo to form bilateral blood islands in splanchnic mesoderm, leading to cardiogenic mesoderm specification. Also, blood islands appear bilaterally in this mesoderm, which will later form blood cells and two dorsal aortae (Gittenberger-de Groot et al. 2005).
In the next stage, after the development of the cardiac progenitor cells in the lateral mesoderm, two “lateral mesodermal islands” or “lateral cardiogenic plates” are created. These two lateral islands fuse together in the midline to form the cardiac crescent which has the following features (Yutzey and Kirby 2002; Lockhart et al. 2011):
-
It has an “arch-shaped” or “bow-shaped” or “horseshoe-shaped” feature in the caudal part of the embryo.
-
It constitutes the so-called primary heart field or first heart field (PHF or FHF), in days 16–18.
-
Then, PHF gives rise to some of the main structure of the mature heart: atria, left ventricle, atrioventricular (AV) canal, and most part of the right ventricle (Fig. 2.1).
Fig. 2.1 
Early development of the heart; (a–c) Dorsal view of an embryo. Hemangioblasts reside in the splanchnic mesoderm in front of the neural plate and on each side of the embryo

PHF has two poles with the following order in migration and looping:
-
A cephalad pole will later constitute the bulbus cordis and aortic roots (i.e., the outflow tract).
-
The caudal pole, which is also known as the sinus venosus, will later constitute the ventricles and the ends of those major veins that bring the venous blood to the heart; also, some segments of the atria are made from the caudal pole (Lin et al. 2012).
While the cardiovascular system forms in about the middle of the third week, this primary heart tube has rhythmic peristalsis activity by the end of the third week. After rightward looping of the primary heart tube, these two poles of the primary heart tube are changed accordingly:
-
Anterior specification: ventricular segments
-
Posterior specification: atrial segments
During later stages of the cardiac development, another important part named secondary heart field (SHF) is created with the following characteristics (Moorman et al. 2003; Verzi et al. 2005; Restivo et al. 2006; Watanabe and Buckingham 2010; Xin et al. 2013; Calkoen et al. 2016):
-
SHF is “a second cellular pool.”
-
It is derived from the ventral pharyngeal splanchnic mesoderm.
-
SHF is located medial and anterior to the cardiac crescent (i.e., medial and anterior to PHF and dorsal to the primary heart tube).
-
Its main function is to give cellular origin to “the arterial and venous side of the heart.”
-
SHF, during days 20 and 21, primarily gives origin to the conotruncal region of the heart, which includes mainly the following segments in the mature heart: other primordial parts of the right ventricle, the interventricular septum (IVS), and endothelial and myocardial components of the outflow tract (i.e., conotruncus).
-
Another major function of the SHF is laterality of cardiac constituents.
Clinical Note
Some of the most common congenital heart disorders due to conotruncal defects are:
-
Persistent truncus arteriosus
-
Tetralogy of Fallot
-
Double-outlet right ventricle
-
Pulmonary atresia
-
Pulmonary stenosis (Restivo et al. 2006)
Formation of the Heart Tube
The heart arises from a common mesodermal pool of progenitor cells, which is part of the cardiopharyngeal region. During the early phases of cardiac development, some parts of the primary cardiogenic area are anterior to the neural tubes; however, due to the very rapid growth of brain vesicles, there is a cephalad movement of the oropharyngeal membrane, which in turn pushes the heart into the more caudal parts; then, the heart comes to an inner position to be part of the future thorax. From another point of view, the oropharyngeal region becomes compressed in between the brain, the yolk, and the heart (Soukup et al. 2013; Diogo et al. 2015).
The process of body folding involves both cephalocaudal and lateral folding of the embryonic plate. As the embryonic plates fold laterally, the endocardial tubes fuse with each other to form a heart tube; by this process, the heart tube is gradually formed (Yutzey and Kirby 2002; Abu-Issa and Kirby 2007).
Usually, looping of the heart tube which is discussed in the next paragraphs is considered as the primary visible sign for asymmetry during embryo development. However, formation of the atrioventricular canal is structurally asymmetric associated with left-sided bulging (Moorman et al. 2003).
Therefore, the heart tube is composed of three distinct layers (Yutzey and Kirby 2002; Lockhart et al. 2011):
-
1.
The inner endothelial cover, i.e., endocardium (which lines the inner layer of the heart tube).
-
2.
The outer myocardial layer, i.e., myocardium; the endocardium and myocardium are separated by an acellular space which is rich in extracellular matrix (ECM); this thick ECM, also named cardiac jelly, is secreted by myocardium and is rich in a number of specific molecules including hyaluronic acid, hyaluronan, fibronectin, fibrillin, proteoglycans, and collagens which have important roles in heart development, especially for proper development of endocardial cushion of the atrioventricular junctions.
-
3.
The epicardium (visceral pericardium) is derived from mesodermal cells that arise from splanchnic mesoderm on the surface of the septum transversum and sinus venous which migrate onto the outer surface of the myocardium.
During later developmental phases, the heart tube become swelled and progressively invaginates into the pericardial cavity. At first, it is attached to the dorsal wall of the body through the dorsal mesocardium which is a fold of the mesodermal tissue (Moorman et al. 2003; Lin et al. 2012). However, the central portion of dorsal mesocardium degenerates and forms the transverse pericardial sinus, which is located between the left and the right sides of the pericardial cavity (Fig. 2.2).

Transverse sections and longitudinal section through embryo at different stages of heart tube formation; (a) 17 days, (b) 18 days, (c) 22 days
The heart is now attached only at its caudal and cranial parts in the pericardial cavity by blood vessels; i.e., the ventricular loop of the heart has gained both “inlet and outlet components.” The inflow to the heart is initially supplied by three pairs of drainage veins into the tubular heart through vitelline veins (return poorly oxygenated blood from the umbilical vesicle), umbilical veins (carry well-oxygenated blood from the chorion), and common cardinal veins (return poorly blood with low oxygen from the embryo body) (Moorman et al. 2003; Lin et al. 2012). On the other side, the outlet supports the cardiac outflow tract; it means that the left and right dorsal aortae make the outflow of the heart vessels, which in turn leads to arteries emerging from the aortic sac and then going to pharyngeal arcs. In the area of the arches, the dorsal aortae are arranged as paired vessels, but caudal parts of the dorsal aorta join together to form a single aorta (thoracic aorta and abdominal aorta).
However, with improvements in heart development, the heart loop is completed and expands; the dorsal mesocardium breaks up, except in the most caudal segment which plays the role of venous pole for the heart; this dorsal mesenchymal part has a central role in some major events of heart development like pulmonary vein development and atrioventricular mesenchymal complex formation.
The Looping of Cardiac Tissue
The primitive heart tube begins to elongate on day 23; however, signaling pathway guaranteeing right-sided looping is initiated earlier. It means that the looping process is signaled during gastrulation; as mentioned earlier, looping of the heart tube is the primary visible sign for asymmetry during embryo development. The process of cardiac loop is completed by day 28 (Yutzey and Kirby 2002; Moorman et al. 2003; Jacobs et al. 2007).
As the primitive heart tube elongates in both cranial and caudal parts, the dorsal mesocardium is detached from the developing left ventricle, leading to liberation of the heart tube. The heart tube bends rightward after its liberation (Moorman et al. 2003).
Then, in the process of looping, the cephalic part of the heart tube bends in three directions:
-
Ventral
-
Caudal
-
To the right
However, the caudal part itself bends again and extends in these three directions:
-
Dorsal
-
Cranial
-
To the left
As mentioned before, these processes of bending are completed by day 28 and lead finally to the cardiac loop.
Over the next 5 weeks, a series of “strictures” and “bulged areas” are created in the primitive heart tube. Through the bulged regions, the following structures come out:
-
Bulbus cordis (which includes truncus arteriosus, conus arteriosus, and conus cordis)
-
Ventricle
-
Atrium
-
Sinus venosus
The resultant “ventricular loop” has two main components:
-
The inlet gives origin to the sinus venosus, which starts at the caudal end and consists of left and right sinus horns; these right and left horns are partially confluent; also, the common cardinal veins drain into the sinus venosus (Yutzey and Kirby 2002).
-
The outlet consists of the conus and truncus, and the arterial system (including aorta and pharyngeal arcs) originates from the outflow tract SHF which is the origin of the outflow tract myocardium which comes out of the ventricular outlet (Restivo et al. 2006; Watanabe and Buckingham 2010).
There are two other chambers that are cranial to the sinus venosus; these are:
-
The primitive atrium which will form the common atrium (also known as primitive atrium); this primitive atrium will form the left and the right atria.
-
The primitive ventricle which will form the left ventricle; however, the primitive ventricle is separated from the next expansion (i.e., the bulbus cordis) by the bulboventricular sulcus; the latter segment will form much of the right ventricle.
Conotruncal segment
The distal outflow tract of the right and left ventricles has the same origin. The conotruncal segment is the cranial-most segment of the ventricles and forms the following parts:
-
Outflow tract: the distal outflow region each of the left and right ventricles.
-
Conus cordis (or conus arteriosus): after creating the outflow tract, the conotruncal segment is further subdivided into the conus cordis (or conus arteriosus) and the truncus arteriosus; conus cordis will be eventually incorporated into the corresponding right and left ventricles.
-
Truncus arteriosus: this is the third part of the conotruncal segment and splits into two parts to form the pulmonary artery and the ascending aorta; however, the most cranial end of the truncus arteriosus is connected to a dilated expansion called the aortic sac (Fig. 2.3).

Developing of the cardiac loop. (a) 18 days. (b) 22 days. (c) 23 days. (d) 24 days. (e) 28 days. (f) 35 days
The aortic sac is continuous with the first aortic arch and, eventually, with the other four aortic arches. The aortic arches form major arteries that transport blood to the head and also to the trunk. On the other hand, the sinus venosus receives blood from the common cardinal vein, the umbilical vein, and the vitelline vein which bring the blood back to the heart from the chorion, the umbilical vesicle, and the embryo, respectively.
Clinical Note
In the case of abnormal looping, there may be:
-
Random looping
-
Anterior looping
-
Leftward looping
In clinic, abnormal looping could be seen in the following clinical syndromes:
-
Abnormal atrial situs (situs inversus or isomerism)
-
Dextrocardia
-
Ventricular inversion
We never see heterotaxia patterns in ventricles; heterotaxia is exclusively seen in the atria of the heart (Gittenberger-de Groot et al. 2005; Jacobs et al. 2007).
The Formation of the Sinus Venosus
At the end of the fourth week, the heart contracts synchronously that led to moving of the blood in one direction. During this time the venous blood from right and left sinus horns enters to the sinus venosus. Usually three important veins enter to the right and left horns including (1) embryo via the common cardinal veins, (2) developing placenta via the umbilical veins, and (3) umbilical vesicle via the vitelline vein. The primordial atrium receives blood from the sinus venosus under the control of sinoatrial valves. The blood then enters into the primordial ventricle through the atrioventricular canal. Simultaneously, with the contraction of the ventricle, the blood is pumped via the bulbus cordis and truncus arteriosus and enters into the aortic sac. The blood then distributes to the embryo, umbilical vesicle, and placenta through the dorsal aortas. Initially, the opening between the atrium and the sinus is big. Soon, however, the sinus entrance shifts to the right side. This shift is affected primarily by left-to-right shunts of blood, which take place in the venous system during the fourth and fifth weeks of development (Fig. 2.4).

Different stages of development in “sinus venosus”; (a) 24 days, (b) 35 days
The Next Steps in Differentiation: Atrial Development
Blood flow from left to right causes enlargement of the right sinus and right-sided veins. The right horn is the main confluence between the original sinus venosus and the atrium; this right horn develops gradually into the right atrium and makes the smooth part of the right atrial wall. Now, the sinoatrial orifice is edged on each side by a valvular fold; these folds will create the right and the left venous valves. After a while, the dorsocranial parts of these valves fuse together; the result is formation of a ridge called septum spurium (Fig. 2.5a).

Coronal sections of the heart at the atrioventricular canal level; (a) 5 weeks, (b) fetal stage
In the next stage, the right sinus horn is merged into the atrial wall; however, the left venous valve and the septum spurium fuse with the atrial septum. Meanwhile, all of the superior parts of the right venous valve are drawn back, but the inferior parts are changed into two parts: the inferior vena cava valve and the coronary sinus valve (Fig. 2.5b).
As mentioned in the previous segments, during development of the heart, the heart segments are arranged from cephalad to caudal as the order (Fig. 2.3d):
-
Vitelline veins
-
Primitive atrium
-
Sinus venosus
-
Ventricle
-
Conotruncal segment (including outflow tract, bulbus cordis, and truncus arteriosus)
However, the sinus venosus is located between vitelline veins (cephalad) and primitive atrium chamber (caudad). In fact, the sinus venosus persists until adulthood, when the sinus venosus forms the smooth-walled parts of the right atrium; in the mature heart, this smooth part of the right atrium is called sinus venarum or venarum sinus, which composes the main parts of the posterior atrial wall and the majority of the interatrial septum and the lateral wall of right atrium; in the adult heart, sinus venarum surrounds the openings of the venae cavae and the coronary sinus (Taylor and Taylor 1997; Ho et al. 2002).
There is a junction between the sinus venosus and the primary atrium which is called crista terminalis (Fig. 2.5b); in the mature heart, crista terminalis is a fibromuscular ridge at the posterolateral region of the right atrium. Crista terminalis is the anatomical margin between the anterior wall of the right atrium (i.e., the trabeculated-walled right atrium which contains pectinate muscles and the right atrial appendage) and the posterior wall of the right atrium (i.e., the smooth-walled parts of the right atrium also named sinus venarum) (Freedom et al. 2005).
Clinical Note
Crista terminalis functions as the anterior pathway for typical atrial fibrillation or atrial flutter. Also, in some patients, crista terminalis mimics the right atrium masses in echocardiographic exams especially in patients with supraventricular arrhythmias in which a suspicious mass in the right atrium is of utmost importance and needs vigilance for differential diagnosis; sophisticated transesophageal echocardiography and/or cardiac computed tomography/magnetic resonance imaging are needed to rule out the differential diagnosis (Ellis et al. 2000; Gaudio et al. 2004; Akcay et al. 2007; Salustri et al. 2010; Na et al. 2011; Siddiqui et al. 2013; Nakanishi et al. 2015).
The primary atrium starts to be formed at about 2 weeks of gestation. As the primitive atrium enlarges, it partially envelops the bulbus cordis; meanwhile, the right atrium and the left atrium are created out of the primary atrium mainly due to the growth of the septum primum:
-
The primitive right atrium is created out of the primitive atrium by fusion of the right sinus horn; as mentioned above, the right atrium is consisted of two main parts, the trabeculated right atrial appendages which give origin to pectinate muscle and the smooth-walled sinus venarum originating from the right horn of the sinus venosus; the pectinate muscles cover the entire wall of the right atrial appendage (Ho et al. 2002).
-
The primitive left atrium expands according to the following order: first of all, an outgrowth of the posterior left atrial wall develops in the left side of the septum primum to the primary pulmonary vein (Fig. 2.5a); this primitive pulmonary vein connects with the veins of the developing lung buds; however, during the next expansion, the pulmonary vein and its branches are merged into the left atrium, making the large part of smooth wall of adult left atrium. In the left atrium, there is not such a structure like crista terminalis; so, the mouth of the appendage plays the differentiation role between rough and smooth parts of the left atrium. Only one vein opens initially into the left atrium; however, in the mature heart, four pulmonary veins drain into the left atrium. The left atrial wall is smooth in the majority of its segments; but, a few parts are rough; meanwhile, much less amount of pectinate muscle is seen in the left atrium compared to the right atrium. In complied heart, the left atrium originates from the appendage of the trabeculated atrial wall, and its roof is adjacent to the aorta and pulmonary artery with its trabeculated surface. On the other hand, the smooth-walled portion covers the majority of left atrial surface, initiates from the pulmonary vein component, and is continued up to the body and the vestibule of the left atrium; also, the smooth wall includes the superior and posterior walls of the left atrium (Fig. 2.5a, b) (Ho et al. 2002).
The Primordial Heart Septation
After cardiac looping, the heart, composed of inner endocardial lining and outer myocardial cells, is septated into four main independent regions, which eventually conform the typical anatomy of the future heart chambers and extracardiac arterial system (Lin et al. 2012):
-
The atrium
-
The atrioventricular canal (also contains endocardial cushions)
-
The ventricle
-
The outflow tract (also contains endocardial cushions)
Septation is a critical transitional stage event in heart development, since during septation, the heart is changed from a single chamber peristaltic tube to a four-chambered pump with unidirectional valves and specific cardiac routes for blood circulation; also, we should note that in normal developmental sequence of the heart, cardiac looping is an essential prerequisite for septation; so we will have the following three stages as the logical and subsequent stages which constitute cardiac development:
-
Cardiac looping
-
Septation
-
Chamber formation
Also, septation, though a single stage in cardiac development, takes place at three different anatomic levels that any defect in each of these levels results in specific lesions; these levels are (Gittenberger-de Groot et al. 2005):
-
The atrium
-
The ventricle
-
The arterial pole
However, myocytes participating in chamber formation are not materially involved in septation (Lamers and Moorman 2002; Lin et al. 2012).
Human heart septation starts at the fourth week and is accomplished by the end of the seventh week and involves two main phases which are composed of sequential events:
-
During the first phase, two actively growing tissue masses (known as ridges) advance toward each other, until they fuse and make a septum (Fig. 2.6a, b). This septum primarily divides the heart lumen into two main single canals (Fig. 2.6a). Similar septum may also be actively shaped by a single mass of growing tissue which will develop till it launches into the opposite side of the lumen (Fig. 2.6c). Synthesis of extracellular matrices and cell proliferation play a main role in the formation of such tissue masses.
Fig. 2.6 
(a) Ridge formation by two actively growing masses; (b) septation and septum formation by two actively growing masses that close to each other until they fuse and make a septum; (c) septation by a single actively growing mass
-
The endocardial cushions develop into the conotruncal and atrioventricular parts; endocardial cushions are local tissue swellings, made of accumulated “cardiac jelly”; in fact, cardiac jelly is abundant masses of extracellular matrix incorporated between the endocardium and the myocardium (Lin et al. 2012). Cardiac jelly is acellular at first; however, after formation of AV cushions, there is an epithelial-to-mesenchymal transition (EMT) of the endocardial cells which cover the cushions (Lockhart et al. 2011).
-
However, endocardial cushions when created participate in the formation of membranous parts of atrial and ventricular septa, the atrioventricular canals, the atrioventricular (AV) valves, and the aortic and pulmonary valves. But, we should remember that endocardium cushions usually do not participate in true septum formation. Instead, endocardial cushions take part in septation process in a different way. Their mechanism is that a thin strip of mass tissue in the walls of the atria and ventricles grows and spreads around to well expand the surrounding tissue, until it makes a thin ridge between two contralateral parts; each of these contralateral thin ridges grows on either side until they reach each other and eventually fuse to form a septum. However, this septation mechanism usually does not completely divide the original lumen; but, it leaves a thin link between the two contralateral parts; while later on, the thin canal is endorsed and secondarily supported by proliferation of neighboring tissues. This mechanism of septation separates the ventricles and atria partially (Fig. 2.6a–c). At times, due to this mechanism or other similar mechanisms, some of the cardiac structures are created that are known as classic septum while they are not real septum and are in fact folds or layers of the myocardial tissue that engulf some adipose tissue in between (Anderson et al. 2003a).

Whatever the mechanism of septation, the process of septum formation and cardiac chambering is not completed until three major subsequent events will happen which eventually will lead into the typical “four-chambered heart”:
-
Creation of the primitive atrial septum (PAS)
-
Creation of the atrioventricular (AV) cushions which also gives origin into tricuspid valve apparatus and mitral valve apparatus
-
Creation of the interventricular septum (IVS)
These three major events are discussed in the next paragraphs.
Septum development in the common atrium
Creation of the primitive atrial septum (PAS)
The septation process in the common atrium starts at the beginning of the fifth week and includes the following steps (McCarthy et al. 2003; Gittenberger-de Groot et al. 2005; Sukernik and Bennett-Guerrero 2007; Asrress et al. 2015; Calkoen et al. 2016):
-
A sickle-formed crest derived from the roof of the common atrium grows toward the middle of the heart lumen; this crest makes the first part of the structure called septum primum.
-
The caudal end of this septum primum develops toward the fused endocardial cushion which is located in the atrioventricular canal.
-
In this stage, there is a gap between two parts of the common atrium which is called the ostium primum and allows blood flow between the two parts of the common atrium (i.e., interatrial flow).
-
The septum primum is fenestrated spontaneously in its superior regions by apoptosis to create ostium secundum; these fenestrations appear in order to create right-to-left shunt in the fetal circulation which allows flow of oxygenated blood coming from the umbilical vein to the other organs of fetal body; in this way, the superior part of the septum primum will be obliterated, though it will be completed by septum secundum.
-
A crescent muscular mass of the ventrocranial common atrial wall originates from the right atrium and grows downward on the right side of the septum primum; this infolding will produce the superior segment of the future interatrial septum; this structure is called septum secundum and it will cover the main part of the ostium secundum; this muscular septum secundum develops during the fifth and sixth weeks of gestation.
-
So, we see that the future interatrial septum is the result of two merging septa: septum primum and septum secundum.
-
The left venous valve and the septum spurium merge with the right side of the septum secundum.
-
Meanwhile, the pulmonary veins will be relocated from the right atrium to the dorsal wall of the left atrium.
-
The defect in the septum secundum is called the fossa ovalis which is usually compensated by septum primum.
-
There is a defect in the borders of septum primum and septum secundum called the foramen ovale, an obliquely elongated cleft in the interatrial septum which is open as long as fetal circulation persists; after birth, transition of circulation from fetal circulation to normal circulation leads to increased pressure in the left cardiac chambers and closure of the foramen ovale, at first physiologically and after a while, anatomically.
-
The sinus venosus is the part of the tissue separating right pulmonary veins from the SVC from the posterior and inferior aspects of the free wall of the right atrium; coronary sinus septum is the part of the myocardial tissue separating the coronary sinus from the left atrium (Geva et al. 2014).
The communication between the endocardial cushions and the lower edge septum primum named the foramen primum or the ostium primum. The ostium primum, as a shunt, helps the oxygenated blood to cross from the left to the right atrium. In the next growth, expansions of the lower and higher endocardia cushions develop toward the rim of the septum primum and block the ostium primum.
Genetic factors related to the development of interatrial septum: a set of genetic studies have shown that atrial septal defects and defects in the conduction system are closely related to NKX2.5 mutations. The other genes that play a main role in the heart development are GATA4 gene. The product of this gene and its interaction with other gene products including TBX5 play an important role in cardiac development. Mutations in this gene alter the transcriptional activity of GATA4 and result in ASD, VSD, and pulmonary valvular stenosis (Gourdie et al. 1999; Gourdie et al. 2003; Christoffels et al. 2004; Moorman et al. 2005; Tomita-Mitchell et al. 2007; Remme et al. 2009; Moskowitz et al. 2011; Xin et al. 2013; Stefanovic and Christoffels 2015).
Clinical Notes
-
ASD is discussed in detail in Chap. 19; however, some of its developmental notes are discussed here in brief.
-
If the pulling-down process of septum primum and septum secundum is deficient, a type of defect occurs in the interatrial septum known as ASD secundum or ostium secundum ASD (ASD II) which is the most common type of ASD.
-
If the ostium primum is not closed by septum primum coming from the underlying AV cushions, primary atrial septal defect (ASD I) occurs which is often in combination with varying degrees of abnormalities in AV cushions, namely, AVSD which is discussed in Chap. 18—AV Septal Defects.
-
In a minority of newborns, fetal circulation persists, leading to a clinical status called persistent fetal circulation (PFC) which is discussed in detail in Chap. 3—Cardiac Physiology; however, PFC leads to increased pressure in the right side over the left side which is the main etiology for persistence of the foramen ovale, a disease known as persistent foramen ovale (PFO) which is discussed under the interatrial defects.
-
The last major type of interatrial defects is called sinus venosus atrial septal defect, which is due to abnormal attachment of venae cavae (often the superior vena cava) leading to atrial septal defect which is usually associated with anomalous attachment of pulmonary veins; this defect is often discussed under anomalous pulmonary venous drainage (Kerut et al. 2001; Sukernik et al. 2001; Oliver et al. 2002; McCarthy et al. 2003; Van Praagh et al. 2003; Sukernik and Bennett-Guerrero 2007; John et al. 2011; Briggs et al. 2012).
Septum development in the atrioventricular canal
Creation of the atrioventricular cushions associated with tricuspid and mitral valves
At the fifth week of gestation, the superior and inferior endocardial cushions are going to be created, so they gradually appear over the primitive left ventricle. Then, an endocardial mass is produced on the ventral and dorsal parts of atrioventricular (AV) canal, and at the same time, this mass is penetrated by mesenchymal cells. In this way, the AV endocardial cushions are made close to each other, while in the next stage, they merge together and are separated to form the left and the right AV canals; these AV canals could discriminate incompletely the primordial ventricle from the primordial atrium.
However, specialized parts of extracellular matrix or cardiac jelly play a crucial role in the development of endocardial cushion; this is why the effect of specific molecules including hyaluronic acid, hyaluronan, fibronectin, fibrillin, proteoglycans, and collagens in cardiac jelly is a leading role (Lockhart et al. 2011; Ray and Niswander 2012; Lalani and Belmont 2014).
The cells that constitute the endocardial cushion tissues are primarily endocardial in origin; however, these endothelial cells migrate into the inner layers of the heart tube to create the primitive mesodermal tissue of this tube which is located in the crux of the heart. This critical process in formation of cardiac cushions is called endothelial-to-mesenchymal transition of endothelial cells in cardiac cushions (Zhang et al. 2014; Davey and Rychik 2016).
There is a detailed list of cellular and molecular factors which play their role in the development of cardiac cushions, and any impairment in their role may lead to endocardial cushion defects; these factors include but are not limited to the following:
-
Transforming growth factors and proteins (like bone morphogenetic protein, BMP)
-
Intercellular signaling molecules and enzymes
-
Extracellular matrices
-
Transcription factors and mutations in their related genes, like GATA4 transcription factor, TGF beta, FOG factor, Smad4, Zic family member 3 (Zic3), NK2 homeobox 5 (Nkx2.5), and T-box protein 5 (Tbx5)
-
Mutations in genes such as CYSTEINE-RICH PROTEIN WITH EGF-LIKE DOMAINS (CRELD1, a cell adhesion molecule) (Yamagishi et al. 2009; Lockhart et al. 2011; Moskowitz et al. 2011; Ray and Niswander 2012; Garside et al. 2013; Liu et al. 2013; Paffett-Lugassy et al. 2013; Xin et al. 2013; Lalani and Belmont 2014; Kathiriya et al. 2015; Stefanovic and Christoffels 2015; Gordon and Gordon 2016)
Atrioventricular Valves
For the creation of the atrioventricular (AV) valves, the following steps happen:
-
The AV valves are produced in a process called endothelial-to-mesenchymal transition of endothelial cells in cardiac cushions through the following subsequent stages.
-
The endocardial cushions are the “progenitors” of the AV valves.
-
The embryologic cells that constitute the endocardial cushions are endothelial cells migrating into the inner layer of the heart tube; this migration creates the primary mesodermal tissue of the heart tube which is located in the heart crux.
-
Afterward, the AV endocardial cushions merge.
-
Then the mesenchymal tissue proliferates locally and surrounds the orifice of the AV canals.
-
In the next stage, the blood flow dips out the tissue on the ventricular surface of the mesenchymal proliferations, which leads to more final form of the valves.
-
However, there is still persistence of the valve links to the ventricular wall by muscular cords; in the last stage, the muscular cords are changed to a dense connective tissue, followed by obliteration of muscular part of the cords.
-
Now the valves include the connective tissue endorsed by endocardium, which are attached by chordae tendineae to the papillary muscles (Fig. 2.7).
Fig. 2.7 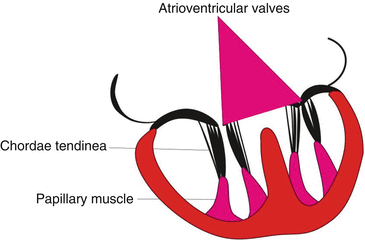
Development of the atrioventricular valves and chordae tendineae
-
And now we have two final valves: the two-leaflet valve in the left atrioventricular canal which is bicuspid, known as the mitral valve, and three-leaflet valve in the right atrioventricular canal known as the tricuspid valve (Gaussin et al. 2005; Zhang et al. 2014; Davey and Rychik 2016).

Many cellular and molecular factors play their role in the normal development of AV cushions; any impairment in their process leads to impaired endocardial cushion development; some of these factors are presented here:
-
Transforming growth factors and proteins (like bone morphogenetic protein, BMP)
-
Intercellular signaling molecules and enzymes
-
Transcription factors and mutations in their related genes, like GATA4 transcription factor, TGF beta, FOG factor, Smad4, Zic family member 3 (Zic3), NK2 homeobox 5 (Nkx2.5), and T-box protein 5 (Tbx5)
-
Extracellular matrices (Yamagishi et al. 2009; Moskowitz et al. 2011; Ray and Niswander 2012; Garside et al. 2013; Liu et al. 2013; Kathiriya et al. 2015; Stefanovic and Christoffels 2015)
Clinical Note
If normal development of the endocardial cushions is impaired, the resulting defects would be seen as defects in the septum (at the crux of the heart) and, also, impairment in normal development of the AV valves, a clinical state known as atrioventricular septal defect “AVSD.” However, these abnormalities in the septum and the AV valves do not have a constant spectrum with varying degrees; so, there are different phenotypes seen in patients with AVSD; detailed discussion on this disease could be found in Chap. 18—AV Septal Defects.
Septum development in the ventricles
Creation of the interventricular septum (IVS)
At the beginning of the fifth week, the primeval ventricles start to expand and create the apical portions of the future ventricles from the primary heart tube. The interventricular septum (IVS) sulcus is the externally separating margin of right and left ventricles, while the internal separator of right and left ventricle is the bulboventricular flange which is part of the primitive ventricle and leads to the development of the muscular part of IVS. Development of the primeval ventricles is one of the main steps in the development of IVS. There are two main parts in IVS:
-
The muscular IVS developed from the bulboventricular flange.
-
The membranous IVS connects the upper margin of the bulboventricular flange to the anterior and posterior endocardial cushions.
However, there are a number of following sequential events leading to the development of IVS:
-
The first sign of division in primordial ventricle is the creation of muscular IVS which is a median ridge in the ventricle floor adjacent to its apex; the edge of muscular IVS is concave and free.
-
During early stage of development, IVS achieves its height by expansion of the ventricles on each side.
-
Afterward, IVS myoblasts start active proliferation, resulting in increased size.
-
The next step is conus septum completion, which happens as a result of tissue extension, starting from the inferior part of endocardial cushion alongside top of the muscular IVS; these parts of tissue finally merge with the neighboring portion of the conus septum.
-
And the final step is closure of the opening above the muscular IVS; when the interventricular foramen closes completely, the membranous part of the IVS is formed.
-
Three sources of tissue take part in the closure of the interventricular opening and formation of the membranous IVS: the left bulbar ridge, the right bulbar ridge, and the endocardial cushions.
The primary ventricular septum or primary ventricular fold is produced following the trabeculation of the ventral part of the muscular IVS. However, there is a smooth part on the dorsal wall of IVS, named the inlet septum; this nomenclature is used because it is located nearby the AV canals. The moderator band or septomarginal trabecula is located on the right wall of muscular IVS, between the primary trabeculated fold and the inlet septum. This structure is a firm connection between the muscular septum and the anterior papillary muscle. When the right ventricular chamber expands, the moderator band is formed nearby the AV canal and dorsal muscular IVS. Eventually, a large part of the mature right ventricular chamber is formed by this expansion. However, if this anatomic area expands incompletely, the developing tricuspid part of the atrioventricular canal remains attached to the interventricular foramen, leading to tricuspid atresia and/or other tricuspid valve anomalies (Lamers and Moorman 2002; Gittenberger-de Groot et al. 2005; Togi et al. 2006; Lin et al. 2012; Poelmann et al. 2014).
Failure or gaps in the development of IVS leads to different forms of VSD; a detailed discussion on VSD is found in Chap. 19.
Septum Development in the Truncus Arteriosus and Conus Cordis
There is a paired ridge, composed of two cushions, which appears in opposing sites of the truncus. These two ridges are located as follows:
-
On the right side of superior wall: the right swelling lies on the superior truncus.
-
On the left side of the inferior wall: the left swelling lies on the inferior truncus.
During the fourth week of gestation, the right-sided swelling located on the superior site of truncus progresses distally and toward the left. Meanwhile, the left-sided swelling located on inferior truncus develops distally to the right. Henceforward, these swellings grow toward the aortic arch, while at the same time, they turn around each other. During this turning movement, they foreshadow the spiral pathway of the upcoming septum (Webb et al. 2003; Anderson et al. 2010).
These swellings deal with a spiral twist of about 180°. Some of the neural crest cells are transferred from the embryonic pharynx and pharyngeal arches to arrive in these edges. The streaming of the blood from the ventricles is one of the main factors that may play a major role in the spiral orientation of the bulbar and truncal edges (Webb et al. 2003; Anderson et al. 2012).
Anyway, this model of development results in the formation of a spiral aorticopulmonary septum when the edges merge. However, after these edges are completely merged, they make the aorticopulmonary septum, separating the truncus into two parts: an aortic channel and a pulmonary channel (Fig. 2.8) (Steding and Seidl 1981; Webb et al. 2003; Okamoto et al. 2010).

Septation of the heart outflow tract and complete the ventricular separation. Formation of the conotruncal ridge; conotruncal ridge fuses with the other compartment to complete the interventricular septum
During the fifth week of gestation, ridges from the subendocardial tissue are formed in the common outflow tract. The spiral orientation of these ridges results in a spiral aorticopulmonary septum during fusion of these ridges. This is the septum that divides the outflow tract into two channels, the pulmonary trunk and the aorta. The second heart field (SHF) plays a crucial role in the development of the outflow tract. Any impairment in SHF or neural crest results in major defects in growth and development of the conotruncal region (Steding and Seidl 1981; Webb et al. 2003; Restivo et al. 2006).
Semilunar Valve Formation
When conotruncal septa are formed, two other cushions are developed which oppose each other in the outflow tract; they are called “the intercalated cushion in the distal conal fragment”; the new cushions, after being remodeled, make two main outflow tract cushions. These two cushions together with the lateral intercalated cushions are excavated; the final result of this process is creating cavities at the origin of the future pulmonary artery and ascending aorta. The primordial deviations from these cavities and the intervening tissues are valvular sinuses and semilunar valves. A set of studies in mice show that semilunar valve leaflets originate mainly from endocardial cushion tissue, associated somewhat with neural crest cells and epicardial cells. The development of semilunar valves in human is completed up to the ninth week (Fig. 2.9) (Anderson et al. 2003b; Hinton and Yutzey 2011; Goenezen et al. 2012; Lin et al. 2012; Sherif 2014; van Geemen et al. 2016).

Development of semilunar valves at weeks 6 (a), 7 (b), and 9 (c). The superior surface is hollowed to form the valves
Development of the Cardiac Conducting System in the Heart
The cardiac conducting system (CCS) is composed of several integral components in a delicate hierarchy which is the mainstay for effective mechanical contractions of the heart chambers (Desplantez et al. 2007; Dun and Boyden 2008; Atkinson et al. 2011).
-
The sinoatrial (SA) node which is the main excitatory and impulse generating location in the heart, generating the regular and rhythmic leading impulses of the heart; they have the most rapid intrinsic rate for impulse generation all over the cardiac cells.
-
Specialized conduction system known as conductive cells which is mainly composed of the atrioventricular conduction pathways, atrioventricular (AV) node, His bundle, and its right and left branches; finally, there are the Purkinje fiber cells or the Purkinje fiber network which is distributed over all parts of the ventricles and conducts the electrical impulse effectively and rapidly over the ventricles (Fig. 2.10).
Fig. 2.10 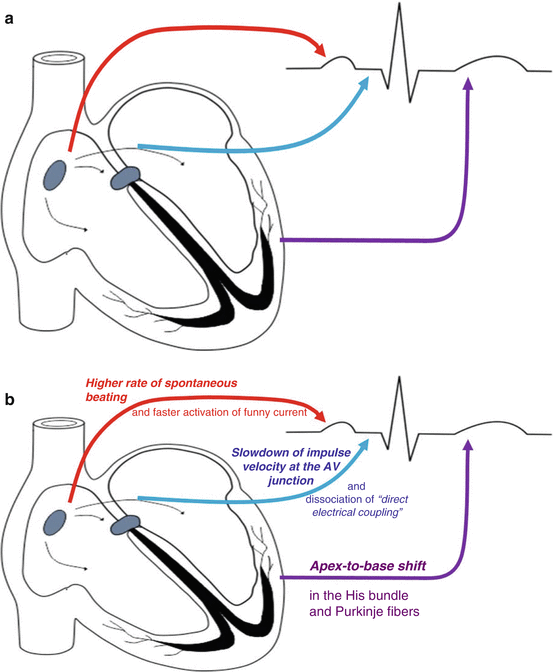
Cardiac conductive system and its elements; in the left, see the relationship of normal electrocardiography with the elements of the system. (a) Relationship of each anatomical segment with electrocardiograph; (b) the embryologic “role definition” for each of the electrical segments of the heart during its development (Modified from Dabbagh (2014). Published with kind permission from © Springer, 2014. All Rights Reserved)

Development of the sinoatrial (SA) node
During the very early phases of heart development, while there is no conduction system development, all of the epithelioid myocytes are electrically active; however, more sophisticated studies have shown that pacemaking area cells have been evolved as a primitive area in the primary sinus venosus and atrium well before the heart begins; these cells are developed and start signaling toward the outflow tract of the developing heart well before any other part of the CCS is developed; their impulses are spread over the heart through gap junctions and connexin proteins (Jalife et al. 1999; Mikawa and Hurtado 2007).
The pacemaker cells changing later to SA node are placed in the caudal portion of the left heart tube at the beginning of their development. At first, the primitive atrium plays the main role of pacemaker in the heart, but in the next phases of development, this role is transferred to sinus venosus. Later, during the fourth week, SA node develops. It is located in the right wall of the sinus venous during early phases of development; however, in the next couple of weeks, its gradual development leads to its merge with the right atrial wall. In the normal heart, SA node lies in the cephalic portion of the posterior wall of right atrium just near the orifice of SVC. During its normal function, SA node has a complex architecture leading to heterogeneous electrical activity. Also, pacemaking cells, especially in SA node, have a number of specific specifications:
-
Higher rate of spontaneous beating
-
Faster activation of the funny current (I f)
-
Greater density of the funny current (Mikawa and Hurtado 2007)
Development of AV node
During the next phases of CCS development, the cells derived from left wall of sinus venous form the atrioventricular node and bundle; these cells are usually placed in the inferior part of interatrial septum and anterior to the coronary sinus foramen. The AV node lies just superior to the endocardial cushions. Development of AV node is a multiple step mechanism. The first theory is that AV node cells are in fact a subpopulation of the primary myocardial cells which their differentiation into “normal” mature myocardial cells is restrained, leading to AV node cells with slow conducive properties; however, additional cellular and molecular theories (including signaling mechanism) are proposed for AV node development; these are beyond the scope of this chapter (Christoffels et al. 2004; Moorman et al. 2005; Mikawa and Hurtado 2007).
During the development of AV node, there are three basic changes in differentiation of the impulse propagation pattern of the CCS; these changes occur during looping of the heart tube and ensure productive and successful pumping of the blood in the four-chambered heart; they are mainly the result of the following three major developmental steps:
-
1.
Significant slowdown of impulse velocity at the AV junction: this impulse slowdown is synchronized with morphologic cleavage between atrial and ventricular chambers; also, this time delay in combination with development of AV cushions leads to production of an effective peristaltic wave in myocardial contraction; the final result will be augmented pumping efficiency of the developing heart.
-
2.
The second step is dissociation of direct electrical coupling between atrial syncytium and ventricular syncytium due to the creation of the interventricular septum.
-
3.
Shift in the electrical direction of the impulse: when septation occurs in the ventricular chambers, there is a very clear and specific change in the process of impulse generation and propagation and that is a dramatic shift in the electrical direction of the impulse from base-to-apex model to apex-to-base model (Mikawa and Hurtado 2007).
Development of bundle branches and His bundles
During the next developmental phases, the AV node bundle fibers pass through the atrium to the ventricles and divide into the right and left bundle branches, distributing over the ventricular myocardial tissue. The growth of the AV node is simultaneous with the formation of the bundle of His, a specialized conducting fiber, connecting the bundle branches to the Purkinje fibers in the peripheral ventricular conduction system. His bundles distribute into the left and the right ventricles by the moderator band.
Development of Purkinje fibers
Development of the Purkinje fibers is from the “working myocytes” as a result of interactions between contractile myocytes with endocardial cells and arterial cells, leading to the development of subendocardial and intramyocardial Purkinje cells, respectively; also, coronary arteries have a very essential role in differentiation of Purkinje cells; endothelin signaling is another main factor that promotes transdifferentiation process of the Purkinje fiber (Gourdie et al. 1999, 2003; Gassanov et al. 2004; Mikawa and Hurtado 2007; Hua et al. 2014).
The CCS has a pivotal role in the function of the heart; any developmental defects leading to abnormalities in the CCS lead to life-threatening cardiac arrhythmias (Wu et al. 2014). Also, the repair of congenital heart defects, especially in patients with ventricular septal defects, is associated with a great risk of damage during surgical correction in all of these conducting structures (Racker 2004).
Atrioventricular (AV) Valve Anomalies
Abnormalities in AV valve development significantly contribute to errors in the remodeling process that leads to formation of valve leaflets, chordae tendineae, and papillary muscles; they originate from endocardial cushion and ventricular myocardium. Partitioning of truncus arteriosus and conus arteriosus in the fifth week of embryologic development is associated with the spiral development and twisting migration of neural crest cells; the final result as mentioned before is the development of the structures known later as “outflow tracts” and “great arteries” (Anderson et al. 2003b; Restivo et al. 2006).
In the pulmonary tract system, the related conus known as pulmonary conus takes part in the formation of the infundibulum which is the anatomic structure between pulmonary valve and the AV valves on one side and the muscular infundibulum on the other side. In the outflow tract of the systemic circulation, the subaortic conus is obliterated leading to the development of the aortic valve surrounded by fibrous continuity of the AV valves (Hinton and Yutzey 2011).
Till now, the main pathogenesis mechanism of AV valve atresia (leading to complete obliteration of the orifice of each valve) is not well known. When the AV septum is not formed, the wedging and remodeling of valves are impaired, and consequently, alignment of the AV canals with their appropriate ventricles fails. It seems to be the underlying mechanism leading to single inlet for the ventricle: ventricular inflow from both atria.
Likewise, in double-outlet right ventricle (DORV), there may be double outlets for a ventricle, due to malalignment of the outflow tract; in other words, the affected ventricle has both the aorta and the pulmonary artery. In this disease, the aortic and pulmonary outflow tracts link to the right ventricle, associated in almost all cases with a ventricular septal defect, leading to arterial blood flow leaving the right ventricle while the blood is a mixture of high oxygenated and poor oxygenated blood. In these patients, the ductus arteriosus links the pulmonary trunk and aorta to each other. Impairment in biventricular circulation leading to a univentricular heart function leads to left ventricle-driven circulation and enlargement of the left ventricle and hypoplastic right ventricle; ductus arteriosus should remain open. Usually this condition leads to cardiac failure unless treated. This disease is discussed more in Chap. 30—Double Outlet Right Ventricle (Anderson et al. 2001, 2003b; Mahle et al. 2008).
Stenosis of Cardiac Semilunar Valves
Stenosis of the semilunar valve is defined as stenosis of either the aortic valve or the pulmonary valve. These conditions are discussed in Chap. 24, Right-Sided Obstructive Lesions, and Chap. 26, Congenital Aortic Valve Anomalies. Congenital valvular stenosis is due to abnormal cavitation and remodeling within the distal conal cushion tissue responsible for forming the aortic semilunar valves, leading to different congenital diseases of the aortic valve.
Cardiac Outflow Tract Septation Anomalies
Truncus arteriosus
Cardiac neural crest cells are multipotent migratory cells that contribute to the cardiac outflow tract formation and the pharyngeal arch arteries. Today we know that a great number of many of the outflow tract septation malformations are in association with abnormal development of neural crest cells; as a result, the conotruncal septa do not form at all, leading to persistent truncus arteriosus. This abnormality will inevitably embrace a defect in ventricular septation, leading to mixing of the blood during departure from two ventricles, i.e., in the common outflow tract; however, the result of this mixing is mainly a left-to-right shunt leading to pulmonary hypertension (Fig. 2.11). If not treated, the child usually dies within the first 2 years of life. This defect is usually corrected surgically, by fixing the VSD and implanting a valved prosthetic shunt connecting the right ventricle and the pulmonary arteries.

The outflow cardiac tract septation defects
Transposition of the great vessels
In about 5 per 10,000 live-born infants, the conotruncal septa develop but without the usual spiral pattern, leading to transposition of the great vessels, in which the left ventricle empties into the pulmonary circulation and the right ventricle empties into the systemic circulation. Inversion of the great vessels is often fatal unless the ductus arteriosus remains patent or is accompanied by intrinsic ASD or VSD or by surgically introduced atrial defects aiming to establish an interatrial communication and hence allowing the deoxygenated systemic and the newly oxygenated pulmonary blood to mix. Inversion can be surgically modified with a favorable prognosis. Nevertheless, it is the main cause of death in infants with cyanotic heart disease younger than 1 year old. A full discussion of the disease is presented in Chap. 21—Transposition of Great Vessels.
Tetralogy of Fallot (TOF)
TOF is a syndrome described by Danish Niels Stensen in 1671, then reported by Eduard Sandifort in 1777, and the exact anatomy of TOF illustrated by William Hunter from St George’s Hospital Medical School, London, in 1784. It was in 1888 that Etienne-Louis Arthur Fallot described L’anatomie pathologique de la maladie bleu; in 1924, the term “tetralogy of Fallot” was created for the first time by Canadian Maude Abbott (Berry 2006; Evans 2008; Van Praagh 2009). The term tetralogy refers to four classic malformations, demonstrated as a graphic in Fig. 2.12, and includes:

Tetralogy of Fallot: (1) pulmonary stenosis, (2) overriding of aorta, (3) interventricular septal defect, and (4) hypertrophy of the right ventricle
-
Pulmonary stenosis
-
Ventricular septal defect
-
Rightward displacement of the aorta (usually known as overriding of the aorta)
-
Right ventricular hypertrophy
The main etiologic mechanism is uneven separation of the outflow tract which results to overriding of the aorta and also malalignment of the muscular outlet septum regarding right and left ventricles; these defects lead to increased filling pressures in the right ventricle, with resultant right ventricular hypertrophy. TOF is among the common cyanotic congenital heart diseases and is generally corrected surgically, including release of the pulmonary trunk obstruction, repair of ventricular septal defect, and, at the same time, correction of overriding; a few years later a considerable number of patients refer for correction of the pulmonary insufficiency which is a sequel of pulmonary stenosis repair. TOF is presented in detail in Chap. 20 (Therrien et al. 2005; Apitz et al. 2009; Starr 2010).
Vascular Formation
Two main mechanisms are involved in the development of blood vessels:
-
1.
Vasculogenesis, a process producing vessels through the process of angioblast coalescence; this mechanism is responsible for the production of the two main vessels, the dorsal aorta and cardinal veins (Williams et al. 2010; Charpentier et al. 2015).
-
2.
Angiogenesis, new buds emerge from the preexisting vessels, and in this way, new vessels are created; this is the mechanism responsible for the development of all body vessels except for dorsal aorta and cardinal veins; however, vascular endothelial growth factor (VEGF) and other growth factors play a crucial role throughout this mechanism (Charpentier and Conlon 2014).
Arterial System
Development of Aortic Arches
The aortic arches are a series of paired developing vascular organs in the embryological development; in addition, they are also known as pharyngeal arch arteries or branchial arches, and they give origin to a number of important vascular structures.
The creation of aortic arches begins between 22nd and 24th days of development. At first a pair of arches is formed; then, following the folding of body, the endocardial tubes move toward the future thorax; meanwhile, the cranial ends of the attached aortae are drawn into a dorsoventral loop. This leads to the location of the first aortic arch pair into the condensed mesenchyme of the first pharyngeal arches in each side of the developing pharynx.
The arterial lumen of the aortic arch arises from the ventral part of the aortic sac, which is an enlargement at the cranial part of the truncus arteriosus. They are linked to the right and left dorsal aortae. Aortic arches are located ventral to the dorsal aorta. The dorsal aortae persist as discrete vessels in the aortic arches area; however, in the fourth week of gestation, they merge together throughout the fourth thoracic segment to the fourth lumbar segment to make a dorsal aorta in midline. Development of aorta occurs during the third week of development, in relation with development of the endocardial tube. During days 26–29, through the process of angiogenesis and vasculogenesis, the second, third, fourth, and sixth arches grow inside their related pharyngeal arches; then, they are incorporated with EPCs that are transferred from the inclosing mesoderm. Also, neural crest mesenchymal cells in the pharyngeal arches play a main role in the usual growth of the arch arteries; here the neural crest cells do not have a direct relationship with the endothelium of these vessels (Kau et al. 2007).
Development of Arterial Tree from Aortic Arches
During 28th to 32nd days of embryonic development, the blood departs the heart via the outflow tract and comes back to the heart through the extrapericardial aortic sac. There is a very important connection between the aortic sac and bilateral dorsal aortae; this connection is developed through third, fourth, and sixth pharyngeal arch arteries, which run in their corresponding pharyngeal arches and then separate in the dorsal region; finally, they merge with the paired dorsal aortae. Later, during the course of development, the aortic sac obliterates and is no more recognized (days 37–42). A summary of the organs created during the embryologic period from the aortic arches is discussed here for each aortic arch based on related extensive studies; also, a very brief summary is presented in Table 2.1 (Graham 2003; Kau et al. 2007; Strilic et al. 2009; Kellenberger 2010; Mirilas 2011; Lammert and Axnick 2012; Stojanovska et al. 2012; Bamforth et al. 2013; Neufeld et al. 2014; Rana et al. 2014; Gupta et al. 2015, 2016; Menshawi et al. 2015; Plein et al. 2015).
The first aortic arches
As the future arches are going to be formed, the first two arches go to remission and go to their earlier states. Later, as the second arch develops, the first arch degenerates absolutely (except a small remnant which makes a part of the maxillary arteries).
The second aortic arches
By day 26, the second aortic arch grows in the second pharyngeal arches and joins the dorsal aortae to the aortic sac; however, the second arch degenerates at the time that the sixth arch is developed; just a small part of the second arch remains which makes the stapedial artery, providing future blood supply to the stapes bone in the developing ear.
The third and fourth aortic arches
During regression of the first arch on day 25, the third and fourth aortic arches are formed; the aortic arch is made of the joint venture of left fourth aortic arch with the merged dorsal aorta and a small part of the aortic sac; this combination is converted to ascending aorta or to aortic arch and the most cranial part of the descending aorta. When the dorsal aortic segments are merged, they lead to caudal parts of the descending aorta at the fourth thoracic level.
During these developmental processes, the fourth arch is much more profuse and protuberant than the third and sixth arterial arches.
The third aortic arch is attached to the dorsal aortae at its cranial part; this attachment has an “end-to-side” fashion, leading to the development of internal carotid arteries as cranial extensions of this attachment.
There is another part of dorsal aorta located between the third and the fourth aortic arches, known as carotid duct; this arterial segment is narrower than caudal segments of the dorsal aorta.
In the next stage of the arterial system development, the dorsal aorta disappears on either side by day 35; these aortic parts join the third and the fourth arch arteries. Therefore, the third aortic arches drain totally into the cranial parts of the dorsal aortae which perfuse the head.
The arteries derived from the third arch include:
-
Right common carotid artery
-
Left common carotid artery
-
Proximal part of the right and left internal carotid arteries
The distal part of each internal carotid artery is developed from the cranial part of the ipsilateral dorsal aorta, and the right and left external carotid arteries develop from the common carotids.
The fifth aortic arches
The fifth pair is not involved in these developmental stages; as a matter of fact, some authors have even questioned its presence or at least claimed that no specific role is attributable to this arch in the development of congenital heart diseases; however, others have defined some role (Bamforth et al. 2013; Gupta et al. 2015, 2016).
The sixth aortic arches
It is formed on day 29 while the second arch degenerates simultaneously; the proximal end of the aortic sac gives rise to the right and left sixth arches which are asymmetrical in the next developmental phases. The right sixth arch loses its distal connection with the right dorsal aorta by the end of the eighth week; in comparison, the left sixth arch does not disappear and its distal part forms the ductus arteriosus. This duct lets the blood transfer from the pulmonary trunk into the descending aorta during gestation. After birth, this duct is closed and is transformed later to ligamentum arteriosum, a rudimentary joint between the pulmonary trunk and aorta.
The uneven growth of the right and left sixth arches has another consequence: asymmetry between left and right recurrent laryngeal nerves, which are branches of the vagus nerves. The laryngeal nerves rise initially under the level of the sixth arch; then, they pass under the right and left sixth arches. Recurrent laryngeal nerves innervate intrinsic muscles of the laryngeal system. Throughout development, the larynx moves cranially in relation to aortic arch; however, the left recurrent laryngeal nerve is trapped below the left sixth arch and remains circled under the developing ligamentum arteriosum. In contrast to the left recurrent nerve, the right recurrent laryngeal nerve is entrapped below the fourth arch which is changed later to the right subclavian artery due to the degeneration of the distal portion of the right sixth aortic arch and also no further development in the fifth arch.
Finally, during these phases, sprouts of pulmonary trunk emerge as the following order: sprouts of the pulmonary trunk could be visible at the caudal segment of the sixth aortic arch. The embryonic pulmonary arteries grow in the splanchnopleuric mesoderm; they initially join the fourth aortic arch; then, they create a new and secondary link with the sixth arch before missing their link with the fourth aortic arches. In fact, the pulmonary arteries join the sixth arch arteries and in the next stages, to the pulmonary trunk; however, pulmonary artery buds are not near each other in this stage; instead, the lumen of the aortic sac is insinuated between them; at this time, the right primary pulmonary artery is more laterally located compared to the left primary pulmonary artery. The distal part of the pulmonary arteries in the lung tissue anastomoses with the existing vasculature in the mesenchymal tissue surrounding the bronchial sprouts.
The seventh aortic arches
The seventh pair of dorsal intersegmental aortae meets and joins; left and right subclavian arteries originate at the caudal point of their convergence; then, they vegetate toward the sprouts of the corresponding upper limb. The right subclavian artery which will supply the upper limbs has a triple-based origin:
-
1.
The right seventh intersegmental artery
-
2.
A short segment of the right dorsal aorta
-
3.
The right fourth arch
The brachiocephalic arteries are derived by the modification of the joint area of the aortic sac and the right fourth arch. The left subclavian artery provides blood to left upper limb; it is developed progressively from the ascending aorta (Menshawi et al. 2015).
By the seventh week, the right dorsal aorta is disconnected from the merged “right sixth arch and midline dorsal aorta”; however, preserving its connection to the right fourth arch. In addition, the right dorsal aorta obtains a branch named the right seventh cervical intersegmental artery, which grows into the right upper limb sprout area.
Also, at the dorsal aspect of the dorsal aortae, a number of small intersegmental arteries sprout at regular distances; they run toward the developing spinal cord to support blood supply to the future spinal cord.
Fate of the Vitelline and Umbilical Arteries
The umbilical vesicle (yolk sac), allantois, and chorion are supplied by the unpaired ventral branches of the dorsal aorta. The three vitelline arteries perfuse the embryologic segments as they pass to the vesicle and later the primitive gut by the following order:
Celiac arterial trunk is the most superior of the three abdominal vitelline arteries and supplies the foregut. During its course, celiac trunk initially joins the dorsal aorta at the seventh cervical level; later, this connection continues down to the 12th thoracic level. The branches of celiac artery vascularize the abdominal part of the foregut, the abdominal esophagus up to the descending segment of the duodenum, and also the liver, pancreas, gallbladder, and spleen. The branch perfusing the spleen develops within the mesoderm of the dorsal mesogastrium. Dorsal mesogastrium is the portion of the dorsal mesentery that suspends the stomach (Cavdar et al. 1997; Yi et al. 2008; Stimec et al. 2011).
Superior mesenteric artery (SMA) is the second abdominal vitelline artery which supplies the midgut. At the beginning of its course, SMA joins the dorsal aorta at the second thoracic level. This connection moves later to the first lumbar level. This artery supplies the developing midgut, including the part of intestine extending from descending segment of the duodenum to transverse colon near the left colic flexure (Rouwet et al. 2000; Bhatnagar et al. 2013).
Inferior mesenteric artery (IMA) is the third and final segment of abdominal vitelline artery which supplies the hindgut. IMA initially joins the dorsal aorta at the 12th thoracic level and later ends at the third lumbar level supplying distal portion of transverse colon, descending colon, sigmoid colon, and the superior rectum (Moonen et al. 2012; Bhatnagar et al. 2013).
Blood supply to the inferior end of the anorectal canal comes from the branches of the iliac arteries. Two umbilical arteries pass from the connecting stalk (primordial umbilical cord) and locate along sides with vessels in the chorion, the embryonic part of the placenta. The umbilical arteries carry poorly oxygenated blood to the placenta. Proximal parts of the umbilical arteries form the internal iliac and superior vesical arteries. However, after birth, distal parts of umbilical artery obliterate and become the medial umbilical ligaments.
Dorsal Aorta Gives Rise to Lateral Branches
Lateral branches of the descending aorta supply the suprarenal glands, gonads, and kidneys. However, these three organs and their arteries have different developmental pathway.
The suprarenal glands originate in the posterior body wall between the sixth and 12th thoracic segments; they are supplied by a pair of lateral aortic branches that arise at an upper lumbar level. Also, some branches from the renal artery and inferior phrenic artery supply the suprarenal glands, but the suprarenal arteries developing from these aortic branches remain the major supply to the glands (Dutta 2010; Sato 2013).
The presumptive gonads become vascularized by gonadal arteries which arise initially at the tenth thoracic level. The gonads descend during development, but the origin of the gonadal arteries becomes stable at the third or fourth lumbar segments. The gonadal arteries elongate, as the gonads (especially the testes) descend further; there are many retroperitoneal anastomoses of the gonadal veins in fetal period, and afterward, left side anastomoses are much more common in both sexes (Raz 2004; Szpinda et al. 2005; Hen et al. 2014; Alfahad and Scott 2015).
In contrast, the definitive kidneys arise in the sacral region and move upward to a lumbar position just below the suprarenal glands. As they migrate, they are vascularized by a sequence of transient aortic side streams that originate at higher levels. These arteries do not elongate for following the ascending kidneys; in contrast, they degenerate. The definitive renal arteries develop from final pair of arteries in the upper lumbar region. Sometimes, a more inferior pair of renal arteries remains as accessory renal arteries (Alfahad and Scott 2015).
Intersegmental Branches
At the beginning of the fourth week, the vasculogenesis process leads to the rise of small sprouts in the posterolateral segments; these primitive sprouts are located alongside the cervical somites extending through to the sacral somites; they grow up and at the same time join to dorsal aorta. In lumbar and thoracic segments, the dorsal branch derived from each of these intersegmental vessels vascularizes toward the growing neural tube and also the epimeres (i.e., the dorsal portion of each somite which gives origin to dorsal muscles that are innervated from the related spinal somite). In addition, the dorsal skin is supplied by the cutaneous branches of these arteries (Gans and Northcutt 1983; Northcutt and Gans 1983; Technau and Scholz 2003).
Besides, these intersegmental vessels vascularize the hypomeres; i.e., hypomeric muscles and related skin are supplied by the ventral branches of intersegmental vessel (hypomeric muscles are those muscles derived from a hypomere and are also innervated by an anterior ramus of the related spinal nerve; also, hypomere is the lateral plate of the mesoderm which grows up to form the ventral body parts) (Finnegan 1961a, b; Technau and Scholz 2003).
The ventral intersegmental arteries in the thoracic segments are transformed to intercostal arteries and cutaneous branches, while in the sacral and lumbar regions, they are developed to lateral sacral and lumbar arteries.
A small branch, the median sacral artery, arises from the dorsal aorta in the area of common iliac artery bifurcation.
In cervical regions, the branches derived from intersegmental arteries link together and create a new complex form of vascularization. Some paired vertebral arteries rise from longitudinal branches that anastomose together in order to form a longitudinal vessel, while in back, they secondarily miss their intersegmental links to the aorta. The anastomoses of intersegmental arteries cause to the development of some arteries such as ascending cervical, deep cervical, internal thoracic, superior intercostal, and inferior and superior epigastric arteries (Adameyko and Fried 2016).
Formation of Limb Arteries
During the development of the limb buds, the arteries derived from seven cervical and five lumbar intersegmental arteries grow into the limb buds to provide blood to them; this primitive perfusion system works through an axial artery, which develops alongside the central axis of each limb. In the upper limb, the axial artery gives origin to the following arteries:
-
Brachial artery
-
Anterior interosseous artery of the forearm
-
Deep palmar arch of the hand
-
Radial, ulnar, and median arteries
In the lower limb, in contrast to the upper limb, the axial artery degenerates; so, the external arteries supply the blood flow of the lower limb, finally leading to these main arteries (Funke and Kuhn 1998):
-
The small sciatic artery which provides the blood for the sciatic nerve in the posterior thigh.
-
A branch of the popliteal artery.
-
A segment of the peroneal artery in the foreleg.
-
All additional arteries of the lower limb are derived from the external iliac artery.
The Formation of Coronary Arteries
Coronary arteries originate from two different segments:
-
1.
The proepicardial cells
-
2.
The epicardial cells
Vascular development in the myocardial tissue follows the initial development of cardiac loop. The primary coronary beds are formed in the trabeculations of the myocardial tissue, while the underling myocardial cells lead to some degrees of epicardial cell change in the form of “epithelial-to-mesenchymal transition,” in such a way that the newly formed mesenchymal cells penetrate the endothelial and smooth muscle cells located in the walls of coronary arteries. The endothelial plexus located in the subepicardial layer is connected to the endothelial sprouts which are located in the walls of the aortic sinuses.
On the other hand, the neural crest cells help smooth muscle cells alongside the proximal division of coronary arteries. The endothelial sprouts of coronary arteries develop some forms of vascular ring which is peritruncal; then, they penetrate and merge into the aortic wall. In fact, coronary arteries attach to the aorta following the development of endothelial cells in such a way that coronary arteries penetrate into the aorta. However, there are only two sprouts of coronary arteries which can produce their lumen and orifices: these are the left and the right coronary arteries. This is one of the main differences between coronary arteries and cardiac veins: coronary arteries are perfused from the systemic circulation through the root of aorta; however, coronary sinuses are the specific site for cardiac veins to connect general circulation. Coronary arteries are perfused in the third trimester. Chapter 34 deals in detail with coronary artery anomalies (Song et al. 2015; Perez-Pomares et al. 2016) (Fig. 2.13).

Schematic drawings of the aortic arches and dorsal aorta before transformation into the definitive vascular pattern and after transformation
The Formation of Venous System
During the early 2nd month, three main veins can be found in each side of the body: vitelline vein, umbilical vein, and cardinal vein; these veins are discussed here (Fig. 2.14):

Illustrations of the primordial veins; initially, three systems of veins are present: the umbilical veins from the chorion, the vitelline veins from the yolk sac, and the cardinal veins from the body of the embryo
-
1.
Vitelline veins or omphalomesenteric veins transfer blood from yolk sac to the sinus venosus. The vitelline veins make a plexus around the duodenal part of the small intestine; afterward, they arrive in sinus venosus after crossing septum transversum. After trespassing the septum transversum, they are invaded by the liver cords growing into the septum; this will lead to interruption of venous courses. In the final stage, hepatic sinusoids are formed as a wide vascular network. Blood transfer from the left side to the right side of the liver leads to formation of an expanded right vitelline vein. In the next developmental phases, the right hepatocardiac channel gives origin to the hepatocardiac segment of IVC; but the proximal portion of the left vitelline vein gradually disappears.
The set of arteries around the duodenum anastomose and grow as a single vessel: portal vein. A small bud of artery grows from the right vitelline vein and creates the superior mesenteric vein. Later on, the distal part of left vitelline vein degenerates.
-
2.
Umbilical veins originate from the chorionic villi and transfer highly oxygenated blood from the placenta to the embryo body. During the early stages, a pair of veins, i.e., the umbilical veins, passes from both sides of the liver, to connect in later stages to sinusoids of the liver. At this time, the proximal portion of each umbilical vein and the remnants of the right umbilical vein begin to disappear. Therefore, the left vein is the only vein that transfers blood from the placenta to the liver. With increased blood flow through the placenta, a direct connection, named ductus venosus, is created between the right hepatocardiac channel and the left umbilical veins. Ductus venosus bypasses the sinusoidal plexus in the liver. However, obliteration of the left umbilical vein leads to ligamentum venosum and obliteration of ductus venosus leads to ligamentum teres.
-
3.
Cardinal veins drain the whole embryo body. In the embryonic period, cardinal veins are the early main veins draining the cephalic portion of the embryo. Anterior and posterior cardinal veins are derived from this early venous system; the anterior cardinal vein drains the cephalic segment of the embryo, while the posterior cardinal vein drains other segments of embryo. These anterior and posterior veins connect together before arriving into the sinus horn, making the short common cardinal vein. Throughout the 2nd embryologic month, a considerable number of veins are created including:
-
The sacrocardinal veins which drain the lower limbs
-
The subcardinal veins which drain the left and right kidneys
-
The supracardinal veins which drain the body wall
The anastomosis between the left and right veins leads to development of vena cava system, transferring the blood from the left side to the right side.
Inferior vena cava (IVC) development
The development of IVC occurs during the fourth to eighth gestation weeks. Creation of IVC has many different stages with many developmental complexities, involving all the anatomic venous segments originating from different venous parts (Spentzouris et al. 2014). It was in 1793 that IVC anomalies were first described by Abernethy in a 10-month-old baby with concomitant polysplenia and dextrocardia (Petik 2015).
IVC has four distinct segments (Spentzouris et al. 2014; Petik 2015; Smillie et al. 2015):
-
Hepatic IVC: vitelline vein is the origin of the hepatic IVC.
-
Suprarenal IVC is originated from right subcardinal vein.
-
Renal IVC is created after anastomosis between right subcardinal and right supracardinal veins.
-
Infrarenal IVC: embryologic development of infrahepatic IVC is the neatly ordered sequence of regression and formation between three paired embryonic veins:
-
Subcardinal veins
-
Supracardinal veins
-
Posterior cardinal veins
-
The anterior cardinal veins anastomose together and form the left brachiocephalic vein. This will lead to an important event in the future: the venous blood drains from the left side of the head and the left upper limbs to the right venous side. Also, the right common cardinal vein and the proximal part of the right anterior cardinal vein join to make the superior vena cava.
The end part of the left posterior cardinal vein enters the left brachiocephalic vein; this venous segment is preserved as a small vessel named the left superior intercostal vein; venous blood from the second and third intercostal spaces drains into these vessels.
The left renal vein is formed by the anastomosis between the subcardinal veins. After that, the left subcardinal vein gradually disappears; only remnants of its distal portion remain which are named the left gonadal vein. Therefore, the right subcardinal vein remains as a main drainage channel which forms the renal segment of the inferior vena cava.
The sacrocardinal veins merge together and make the left common iliac veins. The right sacrocardinal vein forms the sacrocardinal portion of the inferior vena cava.
When the renal and hepatic parts of inferior vena cava anastomose together, the formation of hepatic, renal, and sacrocardinal segments is completed.
After degeneration of the posterior cardinal veins, the supracardinal veins take the main role in body wall venous drainage. The azygos vein is formed after merging of these veins:
-
The fourth to 11th right intercostal veins
-
The right supracardinal vein
-
Part of the posterior cardinal vein
However, the left fourth to seventh intercostal veins merge and create the hemiazygos vein which is drained into the left supracardinal vein; later on, the hemiazygos vein drains into the azygos vein. One should always consider that during the fetal life and later on until adulthood, there are many anatomical variations of the azygos and hemiazygos veins (Krakowiak-Sarnowska et al. 2003; Keskin et al. 2013; Piciucchi et al. 2014).
The Formation of Lymphatic System
The development of the lymphatic system starts around the end of the sixth week, i.e., 2 weeks after recognition of the primordial cardiovascular system. The growth pattern of the lymphatic vessels is much similar to blood vessels. In the final fate of the lymphatic system, there are some connections with the venous system. The early lymphatic capillaries connect with each other to make a lymphatic network (Park et al. 2015).
Development of Lymph Sacs and Lymphatic Ducts
At the end of the embryonic period, there are six primary lymph sacs that include:
-
Two jugular lymph sacs which are located near the junction of the subclavian veins with the anterior cardinal veins which will be the future internal jugular veins
-
Two iliac lymph sacs close to the junction of the iliac veins with the posterior cardinal veins
-
One retroperitoneal lymph sac which is located in the root of the mesentery, located on the posterior abdominal wall
-
One cisterna chili located dorsal to the retroperitoneal lymph sac
In earlier phases, lymphatic vessels create some connections with the lymph sacs and pass along the main veins, in such a way that they create these connections:
-
From the jugular lymph sacs to the head, neck, and upper limbs.
-
From the iliac lymph sacs to the lower trunk and lower limbs.
-
From the retroperitoneal lymph sac and the chyle cistern to the primordial gut; the cistern connects two large channels (right and left thoracic ducts) which are connected to the jugular lymph sacs; later on, between these channels, large anastomosis develops.
The Development of Thoracic Duct
The thoracic duct develops from different parts including:
-
The caudal part of the right thoracic duct
-
The anastomosis between the left and right thoracic ducts
-
The cranial part of the left thoracic duct
Therefore, there are various variations in the origin, course, and insertion of the adult thoracic duct. The right lymphatic duct is created after merging of the right thoracic duct in its cranial part. Both the thoracic duct and the right lymphatic duct join with the venous system at the venous angle which is located between the internal jugular vein and the subclavian vein (Butler et al. 2009; Park et al. 2015).
Circulation Before and After Birth
Fetal Circulation
Prior to birth, the umbilical vein transfers the blood from placenta to the fetus while 80 % of its content is oxygen saturated; when the blood reaches the liver, it goes directly through the ductus venosus into the inferior vena cava (IVC) in a phenomenon called “short-circuiting the liver.” A small amount of this blood arrives to the liver sinusoids; then, it is combined with the blood coming from the portal circulation. The flow of the umbilical blood via the liver sinusoids is regulated by the sphincter of the ductus venosus, located near the entry of the umbilical vein. The sphincter of ductus venosus closes as soon as the first uterine contraction occurs in order to increase venous entry as much as possible and avoid unexpected overloading of the heart.
The blood coming from the placenta is well oxygenated; it comes from the maternal circulation, going through placenta to the umbilical veins and ductus venosus; there, the deoxygenated blood coming from the lower limbs is mixed with the oxygenated blood; from here, the mixed, though still oxygenated, blood is transferred through IVC, going finally to the right atrium.
From this point, part of oxygenated blood trespasses the foramen ovale to go from the right atrium to the left atrium, then the left ventricle and aorta to perfuse systemic organs, including brain and heart muscles, which are the first organs receiving well-oxygenated blood coming from the placenta.
In addition, the remaining oxygenated blood coming from IVC goes through the right atrium to the right ventricle and goes to the pulmonary trunk perfusing the lungs and then returning through pulmonary veins to the left atrium, left ventricle, and descending aorta to go to the to the umbilical artery. However, part of the blood goes through the pulmonary trunk to the ductus arteriosus and aorta to flow into the systemic circulation.
Superior vena cava (SVC) drains the desaturated blood coming from the arms and head; then, in the left atrium, this blood is combined with the desaturated blood coming from the lungs. Afterward, the blood goes into the left ventricle and ascending aorta.
In fetal life, because of high resistance of pulmonary vessels, the majority of blood directly goes from the ductus arteriosus to the descending aorta and does not pass through pulmonary vessels. In the descending aorta, the blood is combined with the blood coming from the proximal aorta. Finally, the desaturated blood goes through the descending aorta and then two umbilical arteries toward the placenta to leave fetal circulation.
As a brief, mixture of the saturated and desaturated blood takes places in the following locations:
-
1.
In the liver, a small amount of the desaturated blood coming from the portal system mixes the saturated blood.
-
2.
In IVC, the deoxygenated blood returned from the organs such as the lower limbs, kidneys, and pelvis is added to the saturated blood.
-
3.
In the right atrium, the desaturated blood returned from the upper and lower limbs and the head is added to the saturated blood.
-
4.
In the left atrium, the blood returned from the lungs is added to the saturated blood.
-
5.
At the junction of the ductus arteriosus with the descending aorta, the desaturated blood coming from the pulmonary trunk is added to “now relatively saturated blood” which finally enters the descending aorta.
Circulatory Alterations After Birth
The events in circulation after birth are discussed in detail in Chap. 3—Pediatric Cardiovascular Physiology. In summary, the fetal circulation shift to a neonatal circulation pattern is an adaptive change in order to tolerate the new “out of the uterus” environment; the final goal is achieving a biventricular parallel circulation pattern instead of the series circulation of the fetus; here, these steps are discussed in brief (Rabi et al. 2006; Gao and Raj 2010; Katheria and Leone 2012; Noori et al. 2012; Rabe et al. 2012; Duley and Batey 2013; Galinsky et al. 2013; McDonald et al. 2013; Azhibekov et al. 2014; van Vonderen et al. 2014a, b; Baik et al. 2015; van Vonderen and Te Pas 2015; Yigit et al. 2015):
-
1.
The establishment of the first inspiratory effort leads to inflation of the lungs and commencement of gas exchange; the result is increased oxygen pressure in the pulmonary vascular bed and the alveoli, which is among the earliest initiatives for transition of circulation; as a result, pulmonary vascular resistance (PVR) drops suddenly in the first 10 min after birth.
-
2.
Placental vessels are clamped; abrupt increase in systemic vascular resistance (SVR) happens.
-
3.
Foramen ovale is closed due to increased pressure in the left atrial chamber over the right atrial chamber; the mechanism of closure is anatomic relationship between septum primum and septum secundum; however, very shortly afterward, the ductus arteriosus is closed, preventing flow between pulmonary artery and the aorta; this is an essential “shift” in body circulation from a “series circulation” to a “parallel circulation.”
-
4.
Complete anatomical closure of the foramen ovale and ductus arteriosus occurs later; often, the foramen ovale is closed physiologically first and then anatomically during a few days, with a minority having only physiologic closure of the foramen ovale even in adulthood; however, the ductus arteriosus is closed during the first 48–72 h after birth.
-
5.
The umbilical vein stops its venous flow at birth.
-
6.
Portal venous system pressure increases, leading to redirection of flow through the hepatic veins.
-
7.
β-receptors are not as much frequent in the newborn heart; but, they increase in myocardium after birth.
-
8.
This transitional circulation needs a minimum of 5 min during the first stages of life, resulting in the “normal postnatal oxygen saturation” state.
-
9.
Blood pressure in the early postnatal period depends on a number of factors, mainly cardiac output (CO) and systemic vascular resistance (SVR); in addition, these factors affect blood pressure: neonatal asphyxia, drugs transferred from the mother like antihypertensive agents or some anesthetics, term infants vs preterm infants, vaginal delivery compared to cesarean section, and gender (female vs male neonates).
-
10.
Cardiac output considered as left and right heart output assumes the “normal” pattern after birth; in fact, in the fetal circulation, the main load of cardiac output is on the right heart; however, just after birth, i.e., in the postnatal period, while the transition from series circulation to parallel circulation ensues, left ventricle and right ventricle cardiac output equalizes and the dominancy of right ventricle output (RVO) disappears, and at the same time, left ventricle output (LVO) and left ventricle stroke volume (SV) increase; also, cardiac output increases in the early postnatal 2–5 h, while after that, due to closure of ductus arteriosus, cardiac output gradually drops to a bit lower level to become stable.
-
11.
If any pathology impairs this process, the transition from fetal to neonatal circulation does not occur correctly, leading to persistence of fetal circulation.
-
12.
Increased PVR due to hypoxia, acidosis, and hypercarbia, history of diabetes in mother during pregnancy, intrauterine inflammation due to infection or other reasons, increased pressure in the right atrium over the left atrium leading to impaired closure of foramen ovale, or patency of ductus arteriosus are among the main etiologies for persistence of fetal circulation.
Angiomas
Abnormal and unnecessary growth of small capillary networks is named a capillary hemangioma. On the other side, accumulating and germinating bundle of large venous sinuses is a cavernous hemangioma. Infantile hemangiomas are the most common vascular tumors in childhood. These tumors develop rapidly and cause an uncontrolled growing mass composed of endothelial cells, lumen obstruction, multilayered basement membranes, and fibrous tissue. Their only risk is when they are located on critical anatomical points like the skull or vertebral canal or in the airways (Marler and Mulliken 2005).
Genetic basis linked with developmental syndromes or chromosomal abnormalities has been proposed for some hemangiomas. For example:
-
5q31-33 of chromosomal region has close link with some hemangioma, containing genes such as FGF4, PDGFβ, and FMS-RELATED TYROSINE KINASE, coding the molecules essential in blood vessel growth.
-
Dysregulation of the TIE/ANG signaling pathways.
-
Mutations of VEGFR2.
-
Multiple hemangioblastomas are related with von Hippel-Lindau disease, with mutations in a gene at chromosome 3p25-26.
-
Excessive angiogenesis may be due to great levels of vascular endothelial growth factor (VEGF) and hypoxia inducible factor 1-alpha (HIF1α) secretion by stromal tumor cells.
Abnormalities of the Ductus Arteriosus
The ductus arteriosus which is located in the right side of the aortic arch grows toward the right side either in front or behind the esophagus and trachea, making ligamentum arteriosum after its postnatal closure.
If it passes from behind of the esophagus, it can lead to a constriction of the esophagus and trachea with clinical dysphagia and/or dyspnea.
When the right and left fourth aortic arches are disappeared and the distal right of dorsal aorta persists, a disease state known as interrupted aortic arch happens. After birth, the aorta supplies the upper body, upper limbs, and head, but the pulmonary artery (contains blood with poorly oxygenated) supplies the lower body and limbs through a patent ductus arteriosus. Detailed discussion about interrupted aortic arch could be found in Chap. 27.
Aortic Coarctation
Coarctation of the aorta (CoA) is one of the common congenital defects, more common in males than females. CoA may occur as a separated abnormality or in association with various other injuries, usually bicuspid aortic valve (BAV) and ventricular septal defect. The disease is discussed in Chap. 24.
CoA is often defined as a restricted aortic segment encompassing localized medial thickening, with some enfolding of the medial and overlaid neointimal tissue.
The underlying mechanism for aortic coarctation is still controversial, although the deformity may be initiated by genetic factors or by teratogens.
There are two main potential etiologies proposed as the causative mechanism for CoA, namely, “ductal theory or ductus tissue theory” and “hemodynamic theory or flow theory” described as follows:
-
Hemodynamic theory or flow theory: more widely accepted, this theory known as Rudolph theory states that during the fetal period, the development of the aortic arch (including the length and the diameter of the arch) depends on the amount of blood flow which passes through the arch; if this blood flow is impaired, it leads to a narrowed and/or hypoplastic aortic arch. This phenomenon explains three main features of CoA: (1) the posterior shelf, (2) the intracardiac defects which are concomitant embryologic lesions, (3) tubular hypoplasia which is often a concomitant lesion of CoA, and (4) the right-sided obstructive lesions which are very rarely associated with CoA.
-
Ductal theory or ductus tissue theory: this theory is also known as Skodaic hypothesis which assumes “abnormal distribution or aberrant migration” of smooth muscle cells (SMC) from the ductus arteriosus to the adjacent aortic tissue as the etiology of CoA; in other words, the ectopic ductal tissue in the aortic isthmus causes CoA and aortic constriction, usually in the isthmus of the aorta (i.e., the junction of aorta with ductus arteriosus) (Krediet 1965; Gillman and Burton 1966; Hutchins 1971; Heymann and Rudolph 1972; Rudolph et al. 1972; Shinebourne and Elseed 1974; Moore and Hutchins 1978; Ho and Anderson 1979; Momma et al. 1982; Van Meurs-Van Woezik and Krediet 1982; Russell et al. 1991; Jimenez et al. 1999; Liberman et al. 2004; Carroll et al. 2006; Kenny and Hijazi 2011).
CoA occurs in three forms:
-
Juxtaductal location which is close to the ductus arteriosus, which is more common than other forms.
-
Preductal CoA, i.e., proximal location which is upstream of the ductus; usually, collateral circulation does not develop, since the ductus arteriosus connects most of the oxygen- and nutrient-enriched blood from the placenta to the lower portion of the body (Fig. 2.15).
Fig. 2.15 
Coarctation of the aorta: (a) preductal type, (b) postductal type
-
Postductal CoA, i.e., distal location which is downstream of ductus; postductal CoA is asymptomatic in some newborn infants, since other arteries including subclavian, internal thoracic, transverse cervical, suprascapular, superior epigastric, intercostal, and lumbar arteries create collateral circulation throughout the embryonic and fetal period (Fig. 2.15).

When the ductus is closed after birth, the infants suffering from CoA are imposed with many abnormalities. In this disease the upper part of the body and head are well perfused but the lower part is somewhat ischemic. CoA repair may be done by surgery, stenting, or balloon dilation angioplasty with different outcomes and chance of recurring stenosis. More detailed discussion on this topic is found in Chap. 24 of this book.
Alagille syndrome refers to genetic problems that result in various symptoms in different parts of the body, including the liver, kidney, heart, and other systems of the body such as paucity of bile ducts and arterial stenosis. Abnormalities related to the disorder generally become apparent in early childhood or infancy.
Vena Cava Anomalies
The anomalies of venae cavae are categorized under two main classes: IVC anomalies and SVC anomalies.
IVC Anomalies
Inferior vena cava (IVC) anomalies were first described in 1793 by Abernethy in a 10-month-old baby who had polysplenia and dextrocardia (Petik 2015).
Embryologic development of IVC is the neatly ordered sequence of regression and formation between three paired embryonic veins (Fig. 2.14):
-
Subcardinal veins
-
Supracardinal veins
-
Postcardinal veins
As mentioned in the previous pages, under IVC development, these events occur between the fourth and eighth gestational weeks, in a multistage process; so, embryologic development of IVC is a potential process for many developmental malformations. Since the development of cross-sectional imaging, congenital anomalies of the IVC and its side streams have been encountered much more frequently, and in a large number of affected patients, these anomalies have been symptom-free.
Congenital anomalies of IVC are diverse, and at least 14 types of anomalies have been reported till now; among these 14, the most important ones are these four types (Srivastava et al. 2005; Dutta 2010):
-
Double IVC, also known as duplication of IVC
-
Left IVC or left-sided IVC
-
Retroaortic left renal vein
-
Circumaortic left renal vein
Among the other lesions are interrupted IVC, other left renal vein anomalies, gonadal vein anomalies, and preduodenal portal vein (Malaki et al. 2012; G et al. 2014; Hagans et al. 2014; Spentzouris et al. 2014; Petik 2015).
Persistent left IVC is usually the result of two events which happen together: regression of the right supracardinal veins plus persistence of the left supracardinal veins.
The typical form of persistent left IVC is that it will merge with the left renal vein then crosses to the other side, reaching the right renal vein, anterior to the aorta; this crossover from left to right is almost always at the level of renal veins. The final result is a normal right-sided IVC which is located in prerenal position (Bass et al. 2000; Malaki et al. 2012; Petik 2015).
There may be two forms of azygos continuation of IVC:
-
Left IVC and absent infrarenal IVC with azygos continuation of IVC
-
Left IVC and absent infrarenal IVC with azygos and hemiazygos continuation
Double IVC is a rare anomaly; its etiology is failure of regression in the caudal part of the left supracardinal veins, leading to persistence of both supracardinal veins and creation of an abnormal left IVC accompanied with right IVC; hence, there will be two IVCs. Significant asymmetry between the sizes of right and left IVCs is possible. The venous blood arriving left IVC will ultimately drain into one of the following veins:
-
Right IVC via the left renal vein
-
Hemiazygos vein rising from thoracic part of the supracardinal system (Bass et al. 2000; Malaki et al. 2012)
SVC Anomalies
Left Superior Vena Cava
In some patients, the left anterior cardinal vein remains, leading to preserved connection between the anterior cardinal vein and the left sinus venosus. Its incidence in general population is about 0.3–0.5 %. If left SVC persists, the blood coming from the left side of the head, neck, and the upper extremity is emptied through the abnormal left SVC into the coronary sinus. This anomaly is presented either as a steady left (double) SVC or a single left SVC. A single left SVC is seen if the left anterior cardinal vein persists while at the same time, the right one is eliminated. In this situation, the left anterior cardinal vein leads to a superior vena cava draining the blood from the entire head and neck, both upper extremities, and the azygos system, then directing all this venous blood into the coronary sinus and right atrium. However, when double and left SVC are present, left SVC drains directly into the left atrium; this condition is seen more commonly in heterotaxy cases (Fig. 2.16). In about 65 % of all left SVC cases, the left brachiocephalic vein is either very small or does not exist at all.

Left SVC: (a) double SVC, (b) single left SVC
Lymphatic System Anomalies
Lymphedema may result from lymphatic hypoplasia. Primary lymphedemas also known as lymphatic obstruction are swellings of the lymphatic vasculature and a major hereditary congenital disorder of the lymphatic system which is induced by hypoplasia of the lymphatic system. This condition usually is accompanied with other abnormalities. There are a number of genes involved in development of lymphedema (Dellinger et al. 2008; Stoll et al. 2013; Sabine and Petrova 2014; Yang and Oliver 2014; Park et al. 2015):
-
FOXC2 gene
-
The transcription factors related to the Forkhead family
-
SOX18 gene mutations and SRY-related transcription factor
-
Ang2, Nrp2, Net, Podoplanin, and Syk
The most severe swellings usually take place in the legs, but, for lymphedema related to Turner syndrome, the lymphatic ducts are obstructed in the neck region and upper trunk; however, this clinical feature may also be the result of lymph-filled cyst development (known as cystic hygromas). These cysts may withdraw if drainage of lymphatic recovers during subsequent stages of development.
Milroy disease is associated with impaired function of the lymphatic system; so, it is a primary lymphedema syndrome due to mutations in the VEGFR3 gene.
References
Abu-Issa R, Kirby ML. Heart field: from mesoderm to heart tube. Annu Rev Cell Dev Biol. 2007;23:45–68.
Adameyko I, Fried K. The nervous system orchestrates and integrates craniofacial development: a review. Front Physiol. 2016;7:49.
Akcay M, Bilen ES, Bilge M, Durmaz T, Kurt M. Prominent crista terminalis: as an anatomic structure leading to atrial arrhythmias and mimicking right atrial mass. J Am Soc Echocardiogr Off Publ Am Soc Echocardiogr. 2007;20:197.e199–110.
Alfahad A, Scott P. Testicular arteries originating from accessory renal arteries. J Vasc Interv Radiol. 2015;26:205.
Anderson RH, Chaudhry B, Mohun TJ, Bamforth SD, Hoyland D, Phillips HM, Webb S, Moorman AF, Brown NA, Henderson DJ. Normal and abnormal development of the intrapericardial arterial trunks in humans and mice. Cardiovasc Res. 2012;95:108–15.
Anderson RH, Cook A, Brown NA, Henderson DJ, Chaudhry B, Mohun T. Development of the outflow tracts with reference to aortopulmonary windows and aortoventricular tunnels. Cardiol Young. 2010;20 Suppl 3:92–9.
Anderson RH, McCarthy K, Cook AC. Continuing medical education. Double outlet right ventricle. Cardiol Young. 2001;11:329–44.
Anderson RH, Webb S, Brown NA, Lamers W, Moorman A. Development of the heart: (2) Septation of the atriums and ventricles. Heart. 2003a;89:949–58.
Anderson RH, Webb S, Brown NA, Lamers W, Moorman A. Development of the heart: (3) formation of the ventricular outflow tracts, arterial valves, and intrapericardial arterial trunks. Heart. 2003b;89:1110–8.
Apitz C, Webb GD, Redington AN. Tetralogy of Fallot. Lancet. 2009;374:1462–71.
Asrress KN, Marciniak M, Marciniak A, Rajani R, Clapp B. Patent foramen ovale: the current state of play. Heart. 2015;101:1916–25.
Atkinson A, Inada S, Li J, Tellez JO, Yanni J, Sleiman R, Allah EA, Anderson RH, Zhang H, Boyett MR, Dobrzynski H. Anatomical and molecular mapping of the left and right ventricular His-Purkinje conduction networks. J Mol Cell Cardiol. 2011;51:689–701.
Azhibekov T, Noori S, Soleymani S, Seri I. Transitional cardiovascular physiology and comprehensive hemodynamic monitoring in the neonate: relevance to research and clinical care. Semin Fetal Neonatal Med. 2014;19:45–53.
Baik N, Urlesberger B, Schwaberger B, Freidl T, Schmolzer GM, Pichler G. Cardiocirculatory monitoring during immediate fetal-to-neonatal transition: a systematic qualitative review of the literature. Neonatology. 2015;107:100–7.
Bamforth SD, Chaudhry B, Bennett M, Wilson R, Mohun TJ, Van Mierop LH, Henderson DJ, Anderson RH. Clarification of the identity of the mammalian fifth pharyngeal arch artery. Clin Anat (New York, NY). 2013;26:173–82.
Bass JE, Redwine MD, Kramer LA, Huynh PT, Harris Jr JH. Spectrum of congenital anomalies of the inferior vena cava: cross-sectional imaging findings. Radiographics. 2000;20:639–52.
Berry D. History of cardiology: Etienne-Louis Fallot, MD. Circulation. 2006;114:f152.
Bhatnagar S, Rajesh S, Jain VK, Patidar Y, Mukund A, Arora A. Celiacomesenteric trunk: a short report. Surg Radiol Anat. 2013;35:979–81.
Briggs LE, Kakarla J, Wessels A. The pathogenesis of atrial and atrioventricular septal defects with special emphasis on the role of the dorsal mesenchymal protrusion. Differentiation Res Biol Divers. 2012;84:117–30.
Butler MG, Isogai S, Weinstein BM. Lymphatic development. Birth Defects Res C Embryo Today. 2009;87:222–31.
Calkoen EE, Hazekamp MG, Blom NA, Elders BB, Gittenberger-de Groot AC, Haak MC, Bartelings MM, Roest AA, Jongbloed MR. Atrioventricular septal defect: from embryonic development to long-term follow-up. Int J Cardiol. 2016;202:784–95.
Carroll SJ, Ferris A, Chen J, Liberman L. Efficacy of prostaglandin E1 in relieving obstruction in coarctation of a persistent fifth aortic arch without opening the ductus arteriosus. Pediatr Cardiol. 2006;27:766–8.
Cavdar S, Sehirli U, Pekin B. Celiacomesenteric trunk. Clin Anat (New York, NY). 1997;10:231–4.
Charpentier MS, Conlon FL. Cellular and molecular mechanisms underlying blood vessel lumen formation. BioEssays News Rev Mol Cell Dev Biol. 2014;36:251–9.
Charpentier MS, Tandon P, Trincot CE, Koutleva EK, Conlon FL. A distinct mechanism of vascular lumen formation in Xenopus requires EGFL7. PLoS One. 2015;10:e0116086.
Christoffels VM, Hoogaars WM, Tessari A, Clout DE, Moorman AF, Campione M. T-box transcription factor Tbx2 represses differentiation and formation of the cardiac chambers. Dev Dyn Off Publ Am Assoc Anatomists. 2004;229:763–70.
Dabbagh A. Cardiac physiology. In: Dabbagh A, Esmailian F, Aranki SF, editors. Postoperative critical care for cardiac surgical patients. Berlin: Springer; 2014. p. 1–39.
Davey BT, Rychik J. The natural history of atrioventricular valve regurgitation throughout fetal life in patients with atrioventricular canal defects. Pediatr Cardiol. 2016;37:50–4.
Dellinger MT, Thome K, Bernas MJ, Erickson RP, Witte MH. Novel FOXC2 missense mutation identified in patient with lymphedema-distichiasis syndrome and review. Lymphology. 2008;41:98–102.
Desplantez T, Dupont E, Severs NJ, Weingart R. Gap junction channels and cardiac impulse propagation. J Membr Biol. 2007;218:13–28.
Diogo R, Kelly RG, Christiaen L, Levine M, Ziermann JM, Molnar JL, Noden DM, Tzahor E. A new heart for a new head in vertebrate cardiopharyngeal evolution. Nature. 2015;520:466–73.
Duley L, Batey N. Optimal timing of umbilical cord clamping for term and preterm babies. Early Hum Dev. 2013;89:4.
Dun W, Boyden PA. The Purkinje cell; 2008 style. J Mol Cell Cardiol. 2008;45:617–24.
Dutta S. Suprarenal gland-arterial supply: an embryological basis and applied importance. Rom J Morphol Embryol Revue roumaine de morphologie et embryologie. 2010;51:137–40.
Ellis WS, SippensGroenewegen A, Auslander DM, Lesh MD. The role of the crista terminalis in atrial flutter and fibrillation: a computer modeling study. Ann Biomed Eng. 2000;28:742–54.
Evans WN. “Tetralogy of Fallot” and Etienne-Louis Arthur Fallot. Pediatr Cardiol. 2008;29:637–40.
Finnegan CV. An analysis of the postgastrula differentiation of the hypomere. I. The influence of tissue mass and of endoderm in Ambystoma punctatum. J Embryol Exp Morphol. 1961a;9:294–309.
Finnegan CV. An analysis of the postgastrula differentiation of the hypomere. II. The influence of endoderm and tissue mass in Taricha torosa. J Embryol Exp Morphol. 1961b;9:609–17.
Freedom RM, Jaeggi ET, Lim JS, Anderson RH. Hearts with isomerism of the right atrial appendages – one of the worst forms of disease in 2005. Cardiol Young. 2005;15:554–67.
Funke C, Kuhn HJ. The morphogenesis of the arteries of the pelvic extremity. A comparative study of mammals with special reference to the tree shrew Tupaia belangeri (Tupaiidae, Scandentia, Mammalia). Adv Anat Embryol Cell Biol. 1998;144:1–97.
Galinsky R, Hooper SB, Wallace MJ, Westover AJ, Black MJ, Moss TJ, Polglase GR. Intrauterine inflammation alters cardiopulmonary and cerebral haemodynamics at birth in preterm lambs. J Physiol. 2013;591:2127–37.
Gans C, Northcutt RG. Neural crest and the origin of vertebrates: a new head. Science (New York, NY). 1983;220:268–73.
Gao Y, Raj JU. Regulation of the pulmonary circulation in the fetus and newborn. Physiol Rev. 2010;90:1291–335.
Garside VC, Chang AC, Karsan A, Hoodless PA. Co-ordinating Notch, BMP, and TGF-beta signaling during heart valve development. Cell Mol Life Sci. 2013;70:2899–917.
Gassanov N, Er F, Zagidullin N, Hoppe UC. Endothelin induces differentiation of ANP-EGFP expressing embryonic stem cells towards a pacemaker phenotype. FASEB J Off Publ Fed Am Soc Exp Biol. 2004;18:1710–2.
Gaudio C, Di Michele S, Cera M, Nguyen BL, Pannarale G, Alessandri N. Prominent crista terminalis mimicking a right atrial mixoma: cardiac magnetic resonance aspects. Eur Rev Med Pharmacol Sci. 2004;8:165–8.
Gaussin V, Morley GE, Cox L, Zwijsen A, Vance KM, Emile L, Tian Y, Liu J, Hong C, Myers D, Conway SJ, Depre C, Mishina Y, Behringer RR, Hanks MC, Schneider MD, Huylebroeck D, Fishman GI, Burch JB, Vatner SF. Alk3/Bmpr1a receptor is required for development of the atrioventricular canal into valves and annulus fibrosus. Circ Res. 2005;97:219–26.
Geva T, Martins JD, Wald RM. Atrial septal defects. Lancet. 2014;383:1921–32.
Gillman RG, Burton AC. Constriction of the neonatal aorta by raised oxygen tension. Circ Res. 1966;19:755–65.
Gittenberger-de Groot AC, Bartelings MM, Deruiter MC, Poelmann RE. Basics of cardiac development for the understanding of congenital heart malformations. Pediatr Res. 2005;57:169–76.
Goenezen S, Rennie MY, Rugonyi S. Biomechanics of early cardiac development. Biomech Model Mechanobiol. 2012;11:1187–204.
Gordon NK, Gordon R. The organelle of differentiation in embryos: the cell state splitter. Theor Biol Med Model. 2016;13:11.
Gourdie RG, Kubalak S, Mikawa T. Conducting the embryonic heart: orchestrating development of specialized cardiac tissues. Trends Cardiovasc Med. 1999;9:18–26.
Gourdie RG, Harris BS, Bond J, Justus C, Hewett KW, O’Brien TX, Thompson RP, Sedmera D. Development of the cardiac pacemaking and conduction system. Birth Defects Res C Embryo Today. 2003;69:46–57.
Graham A. Development of the pharyngeal arches. Am J Med Genet A. 2003;119a:251–6.
Gupta SK, Bamforth SD, Anderson RH. How frequent is the fifth arch artery? Cardiol Young. 2015;25:628–46.
Gupta SK, Gulati GS, Anderson RH. Clarifying the anatomy of the fifth arch artery. Ann Pediatr Cardiol. 2016;9:62–7.
Hagans I, Markelov A, Makadia M. Unique venocaval anomalies: case of duplicate superior vena cava and interrupted inferior vena cava. J Radiol Case Rep. 2014;8:20–6.
Hen G, Friedman-Einat M, Sela-Donenfeld D. Primordial germ cells in the dorsal mesentery of the chicken embryo demonstrate left-right asymmetry and polarized distribution of the EMA1 epitope. J Anat. 2014;224:556–63.
Heymann MA, Rudolph AM. Effects of congenital heart disease on fetal and neonatal circulations. Prog Cardiovasc Dis. 1972;15:115–43.
Hinton RB, Yutzey KE. Heart valve structure and function in development and disease. Annu Rev Physiol. 2011;73:29–46.
Ho SY, Anderson RH. Coarctation, tubular hypoplasia, and the ductus arteriosus. Histological study of 35 specimens. Br Heart J. 1979;41:268–74.
Ho SY, Anderson RH, Sanchez-Quintana D. Atrial structure and fibres: morphologic bases of atrial conduction. Cardiovasc Res. 2002;54:325–36.
Hua LL, Vedantham V, Barnes RM, Hu J, Robinson AS, Bressan M, Srivastava D, Black BL. Specification of the mouse cardiac conduction system in the absence of Endothelin signaling. Dev Biol. 2014;393:245–54.
Hutchins GM. Coarctation of the aorta explained as a branch-point of the ductus arteriosus. Am J Pathol. 1971;63:203–14.
Jacobs JP, Anderson RH, Weinberg PM, Walters 3rd HL, Tchervenkov CI, Del Duca D, Franklin RC, Aiello VD, Beland MJ, Colan SD, Gaynor JW, Krogmann ON, Kurosawa H, Maruszewski B, Stellin G, Elliott MJ. The nomenclature, definition and classification of cardiac structures in the setting of heterotaxy. Cardiol Young. 2007;17 Suppl 2:1–28.
Jalife J, Morley GE, Vaidya D. Connexins and impulse propagation in the mouse heart. J Cardiovasc Electrophysiol. 1999;10:1649–63.
Jimenez M, Daret D, Choussat A, Bonnet J. Immunohistological and ultrastructural analysis of the intimal thickening in coarctation of human aorta. Cardiovasc Res. 1999;41:737–45.
John J, Abrol S, Sadiq A, Shani J. Mixed atrial septal defect coexisting ostium secundum and sinus venosus atrial septal defect. J Am Coll Cardiol. 2011;58:e9.
Katheria A, Leone T. Altered transitional circulation in infants of diabetic mothers with strict antenatal obstetric management: a functional echocardiography study. J Perinatol. 2012;32:508–13.
Kathiriya IS, Nora EP, Bruneau BG. Investigating the transcriptional control of cardiovascular development. Circ Res. 2015;116:700–14.
Kau T, Sinzig M, Gasser J, Lesnik G, Rabitsch E, Celedin S, Eicher W, Illiasch H, Hausegger KA. Aortic development and anomalies. Semin Intervent Radiol. 2007;24:141–52.
Kellenberger CJ. Aortic arch malformations. Pediatr Radiol. 2010;40:876–84.
Kenny D, Hijazi ZM. Coarctation of the aorta: from fetal life to adulthood. Cardiol J. 2011;18:487–95.
Kerut EK, Norfleet WT, Plotnick GD, Giles TD. Patent foramen ovale: a review of associated conditions and the impact of physiological size. J Am Coll Cardiol. 2001;38:613–23.
Keskin S, Keskin Z, Sekmenli N. The independent right and left azygos veins with hemiazygos absence: a rare case presentation. Case Rep Vasc Med. 2013;2013:282416.
Krakowiak-Sarnowska E, Wisniewski M, Szpinda M, Krakowiak H. Variability of the azygos vein system in human foetuses. Folia Morphol. 2003;62:427–30.
Krediet P. An hypothesis of the development of coarctation in man. Acta Morphol Neerl Scand. 1965;6:207–12.
Lalani SR, Belmont JW. Genetic basis of congenital cardiovascular malformations. Eur J Med Genet. 2014;57:402–13.
Lamers WH, Moorman AF. Cardiac septation: a late contribution of the embryonic primary myocardium to heart morphogenesis. Circ Res. 2002;91:93–103.
Lammert E, Axnick J. Vascular lumen formation. Cold Spring Harb Perspect Med. 2012;2:a006619.
Latha GA, Kagali NA, Kagali NA, M S. Preduodenal portal vein in adult with polysplenia syndrome revisited with a case report. Indian J Surg. 2014;76:137–42.
Liberman L, Gersony WM, Flynn PA, Lamberti JJ, Cooper RS, Stare TJ. Effectiveness of prostaglandin E1 in relieving obstruction in coarctation of the aorta without opening the ductus arteriosus. Pediatr Cardiol. 2004;25:49–52.
Lin CJ, Lin CY, Chen CH, Zhou B, Chang CP. Partitioning the heart: mechanisms of cardiac septation and valve development. Development. 2012;139:3277–99.
Liu Y, Lu X, Xiang FL, Lu M, Feng Q. Nitric oxide synthase-3 promotes embryonic development of atrioventricular valves. PLoS One. 2013;8:e77611.
Lockhart M, Wirrig E, Phelps A, Wessels A. Extracellular matrix and heart development. Birth Defects Res A Clin Mol Teratol. 2011;91:535–50.
Mahle WT, Martinez R, Silverman N, Cohen MS, Anderson RH. Anatomy, echocardiography, and surgical approach to double outlet right ventricle. Cardiol Young. 2008;18 Suppl 3:39–51.
Malaki M, Willis AP, Jones RG. Congenital anomalies of the inferior vena cava. Clin Radiol. 2012;67:165–71.
Marler JJ, Mulliken JB. Current management of hemangiomas and vascular malformations. Clin Plast Surg. 2005;32:99–116, ix.
McCarthy K, Ho S, Anderson R. Defining the morphologic phenotypes of atrial septal defects and interatrial communications. Images Paediatr Cardiol. 2003;5:1–24.
McDonald SJ, Middleton P, Dowswell T, Morris PS. Effect of timing of umbilical cord clamping of term infants on maternal and neonatal outcomes. Cochrane Database Syst Rev. 2013;7:CD004074.
Menshawi K, Mohr JP, Gutierrez J. A functional perspective on the embryology and anatomy of the cerebral blood supply. J Stroke. 2015;17:144–58.
Mikawa T, Hurtado R. Development of the cardiac conduction system. Semin Cell Dev Biol. 2007;18:90–100.
Mirilas P. Lateral congenital anomalies of the pharyngeal apparatus: part I. Normal developmental anatomy (embryogenesis) for the surgeon. Am Surg. 2011;77:1230–42.
Momma K, Takao A, Ando M. Angiocardiographic study of coarctation of the aorta–morphology and morphogenesis. Jpn Circ J. 1982;46:174–83.
Moonen RM, Kessels CG, Zimmermann LJ, Villamor E. Mesenteric artery reactivity and small intestine morphology in a chicken model of hypoxia-induced fetal growth restriction. J Physiol Pharmacol Off J Polish Physiol Soc. 2012;63:601–12.
Moore GW, Hutchins GM. Association of interrupted aortic arch with malformations producing reduced blood flow to the fourth aortic arches. Am J Cardiol. 1978;42:467–72.
Moorman A, Webb S, Brown NA, Lamers W, Anderson RH. Development of the heart: (1) formation of the cardiac chambers and arterial trunks. Heart. 2003;89:806–14.
Moorman AF, Christoffels VM, Anderson RH. Anatomic substrates for cardiac conduction. Heart Rhythm. 2005;2:875–86.
Moskowitz IP, Wang J, Peterson MA, Pu WT, Mackinnon AC, Oxburgh L, Chu GC, Sarkar M, Berul C, Smoot L, Robertson EJ, Schwartz R, Seidman JG, Seidman CE. Transcription factor genes Smad4 and Gata4 cooperatively regulate cardiac valve development. [corrected]. Proc Natl Acad Sci U S A. 2011;108:4006–11.
Na JO, Kim EJ, Mun SJ, Choi EH, Mun JH, Lee HR, Kim YK, Yong HS. Prominent crista terminalis in patients with embolic events. J Cardiovasc Ultrasound. 2011;19:156–8.
Nakanishi T, Fukuzawa K, Yoshida A, Itoh M, Imamura K, Fujiwara R, Suzuki A, Yamashita S, Matsumoto A, Konishi H, Ichibori H, Hirata K. Crista terminalis as the anterior pathway of typical atrial flutter: insights from entrainment map with 3D intracardiac ultrasound. Pacing Clin Electrophysiol. 2015;38:608–16.
Neufeld S, Planas-Paz L, Lammert E. Blood and lymphatic vascular tube formation in mouse. Semin Cell Dev Biol. 2014;31:115–23.
Noori S, Wlodaver A, Gottipati V, McCoy M, Schultz D, Escobedo M. Transitional changes in cardiac and cerebral hemodynamics in term neonates at birth. J Pediatr. 2012;160:943–8.
Northcutt RG, Gans C. The genesis of neural crest and epidermal placodes: a reinterpretation of vertebrate origins. Q Rev Biol. 1983;58:1–28.
Okamoto N, Akimoto N, Hidaka N, Shoji S, Sumida H. Formal genesis of the outflow tracts of the heart revisited: previous works in the light of recent observations. Congenit Anom. 2010;50:141–58.
Oliver JM, Gallego P, Gonzalez A, Dominguez FJ, Aroca A, Mesa JM. Sinus venosus syndrome: atrial septal defect or anomalous venous connection? A multiplane transoesophageal approach. Heart. 2002;88:634–8.
Paffett-Lugassy N, Singh R, Nevis KR, Guner-Ataman B, O’Loughlin E, Jahangiri L, Harvey RP, Burns CG, Burns CE. Heart field origin of great vessel precursors relies on nkx2.5-mediated vasculogenesis. Nat Cell Biol. 2013;15:1362–9.
Park SJ, Park SY, Choi H. Aberrent thoracic duct cyst in postrior mediastinum. Korean J Thorac Cardiovasc Surg. 2015;48:225–7.
Perez-Pomares JM, de la Pompa JL, Franco D, Henderson D, Ho SY, Houyel L, Kelly RG, Sedmera D, Sheppard M, Sperling S, Thiene G, van den Hoff M, Basso C. Congenital coronary artery anomalies: a bridge from embryology to anatomy and pathophysiology-a position statement of the development, anatomy, and pathology ESC Working Group. Cardiovasc Res. 2016;109:204–16.
Petik B. Inferior vena cava anomalies and variations: imaging and rare clinical findings. Insights Imaging. 2015;6:631–9.
Piciucchi S, Barone D, Sanna S, Dubini A, Goodman LR, Oboldi D, Bertocco M, Ciccotosto C, Gavelli G, Carloni A, Poletti V. The azygos vein pathway: an overview from anatomical variations to pathological changes. Insights Imaging. 2014;5:619–28.
Plein A, Fantin A, Ruhrberg C. Neural crest cells in cardiovascular development. Curr Top Dev Biol. 2015;111:183–200.
Poelmann RE, Gittenberger-de Groot AC, Vicente-Steijn R, Wisse LJ, Bartelings MM, Everts S, Hoppenbrouwers T, Kruithof BP, Jensen B, de Bruin PW, Hirasawa T, Kuratani S, Vonk F, van de Put JM, de Bakker MA, Richardson MK. Evolution and development of ventricular septation in the amniote heart. PLoS One. 2014;9:e106569.
Rabe H, Diaz-Rossello JL, Duley L, Dowswell T. Effect of timing of umbilical cord clamping and other strategies to influence placental transfusion at preterm birth on maternal and infant outcomes. Cochrane Database Syst Rev. 2012;8:CD003248.
Rabi Y, Yee W, Chen SY, Singhal N. Oxygen saturation trends immediately after birth. J Pediatr. 2006;148:590–4.
Racker DK. The AV junction region of the heart: a comprehensive study correlating gross anatomy and direct three-dimensional analysis. Part II. Morphology and cytoarchitecture. Am J Physiol Heart Circ Physiol. 2004;286:H1853–71.
Rana MS, Sizarov A, Christoffels VM, Moorman AF. Development of the human aortic arch system captured in an interactive three-dimensional reference model. Am J Med Genet A. 2014;164a:1372–83.
Ray HJ, Niswander L. Mechanisms of tissue fusion during development. Development. 2012;139:1701–11.
Raz E. Guidance of primordial germ cell migration. Curr Opin Cell Biol. 2004;16:169–73.
Remme CA, Verkerk AO, Hoogaars WM, Aanhaanen WT, Scicluna BP, Annink C, van den Hoff MJ, Wilde AA, van Veen TA, Veldkamp MW, de Bakker JM, Christoffels VM, Bezzina CR. The cardiac sodium channel displays differential distribution in the conduction system and transmural heterogeneity in the murine ventricular myocardium. Basic Res Cardiol. 2009;104:511–22.
Restivo A, Piacentini G, Placidi S, Saffirio C, Marino B. Cardiac outflow tract: a review of some embryogenetic aspects of the conotruncal region of the heart. Anat Rec A Discov Mol Cell Evol Biol. 2006;288:936–43.
Rouwet EV, De Mey JG, Slaaf DW, Heineman E, Ramsay G, Le Noble FA. Development of vasomotor responses in fetal mesenteric arteries. Am J Physiol Heart Circ Physiol. 2000;279:H1097–105.
Rudolph AM, Heymann MA, Spitznas U. Hemodynamic considerations in the development of narrowing of the aorta. Am J Cardiol. 1972;30:514–25.
Russell GA, Berry PJ, Watterson K, Dhasmana JP, Wisheart JD. Patterns of ductal tissue in coarctation of the aorta in the first three months of life. J Thorac Cardiovasc Surg. 1991;102:596–601.
Sabine A, Petrova TV. Interplay of mechanotransduction, FOXC2, connexins, and calcineurin signaling in lymphatic valve formation. Adv Anat Embryol Cell Biol. 2014;214:67–80.
Salustri A, Bakir S, Sana A, Lange P, Al Mahmeed WA. Prominent crista terminalis mimicking a right atrial mass: case report. Cardiovasc Ultrasound. 2010;8:47.
Sato Y. Dorsal aorta formation: separate origins, lateral-to-medial migration, and remodeling. Dev Growth Differ. 2013;55:113–29.
Sherif HM. Heterogeneity in the segmental development of the aortic tree: impact on management of genetically triggered aortic aneurysms. Aorta (Stamford). 2014;2:186–95.
Shinebourne EA, Elseed AM. Relation between fetal flow patterns, coarctation of the aorta, and pulmonary blood flow. Br Heart J. 1974;36:492–8.
Siddiqui AU, Daimi SR, Gandhi KR, Siddiqui AT, Trivedi S, Sinha MB, Rathore M. Crista terminalis, musculi pectinati, and taenia sagittalis: anatomical observations and applied significance. ISRN Anat. 2013;2013:803853.
Smillie RP, Shetty M, Boyer AC, Madrazo B, Jafri SZ. Imaging evaluation of the inferior vena cava. Radiographics. 2015;35:578–92.
Song G, Ren W, Tang L, Hou Y, Zhou K. Coronary artery fistula from the left circumflex to coronary sinus in infant: case report with literature review. Int J Cardiol. 2015;188:37–9.
Soukup V, Horacek I, Cerny R. Development and evolution of the vertebrate primary mouth. J Anat. 2013;222:79–99.
Spentzouris G, Zandian A, Cesmebasi A, Kinsella CR, Muhleman M, Mirzayan N, Shirak M, Tubbs RS, Shaffer K, Loukas M. The clinical anatomy of the inferior vena cava: a review of common congenital anomalies and considerations for clinicians. Clin Anat (New York, NY). 2014;27:1234–43.
Srivastava A, Singh KJ, Suri A, Vijjan V, Dubey D. Inferior vena cava in urology: importance of developmental abnormalities in clinical practice. Scientific World Journal. 2005;5:558–63.
Starr JP. Tetralogy of fallot: yesterday and today. World J Surg. 2010;34:658–68.
Steding G, Seidl W. Contribution to the development of the heart, Part II: morphogenesis of congenital heart diseases. Thorac Cardiovasc Surg. 1981;29:1–16.
Stefanovic S, Christoffels VM. GATA-dependent transcriptional and epigenetic control of cardiac lineage specification and differentiation. Cell Mol Life Sci. 2015;72:3871–81.
Stimec BV, Terraz S, Fasel JH. The third time is the charm–anastomosis between the celiac trunk and the left colic artery. Clin Anat (New York, NY). 2011;24:258–61.
Stojanovska J, Cascade PN, Chong S, Quint LE, Sundaram B. Embryology and imaging review of aortic arch anomalies. J Thorac Imaging. 2012;27:73–84.
Stoll SJ, Bartsch S, Kroll J. HOXC9 regulates formation of parachordal lymphangioplasts and the thoracic duct in zebrafish via stabilin 2. PLoS One. 2013;8:e58311.
Strilic B, Kucera T, Eglinger J, Hughes MR, McNagny KM, Tsukita S, Dejana E, Ferrara N, Lammert E. The molecular basis of vascular lumen formation in the developing mouse aorta. Dev Cell. 2009;17:505–15.
Sukernik MR, Bennett-Guerrero E. The incidental finding of a patent foramen ovale during cardiac surgery: should it always be repaired? A core review. Anesth Analg. 2007;105:602–10.
Sukernik MR, Mets B, Bennett-Guerrero E. Patent foramen ovale and its significance in the perioperative period. Anesth Analg. 2001;93:1137–46.
Szpinda M, Frackiewicz P, Flisinski P, Wisniewski M, Krakowiak-Sarnowska E. The retroperitoneal anastomoses of the gonadal veins in human foetuses. Folia Morphol (Warsz). 2005;64:72–7.
Taylor JR, Taylor AJ. The relationship between the sinus node and the right atrial appendage. Can J Cardiol. 1997;13:85–92.
Technau U, Scholz CB. Origin and evolution of endoderm and mesoderm. Int J Dev Biol. 2003;47:531–9.
Therrien J, Provost Y, Merchant N, Williams W, Colman J, Webb G. Optimal timing for pulmonary valve replacement in adults after tetralogy of Fallot repair. Am J Cardiol. 2005;95:779–82.
Togi K, Yoshida Y, Matsumae H, Nakashima Y, Kita T, Tanaka M. Essential role of Hand2 in interventricular septum formation and trabeculation during cardiac development. Biochem Biophys Res Commun. 2006;343:144–51.
Tomita-Mitchell A, Maslen CL, Morris CD, Garg V, Goldmuntz E. GATA4 sequence variants in patients with congenital heart disease. J Med Genet. 2007;44:779–83.
van Geemen D, Soares AL, Oomen PJ, Driessen-Mol A, Janssen-van den Broek MW, van den Bogaerdt AJ, Bogers AJ, Goumans MJ, Baaijens FP, Bouten CV. Age-dependent changes in geometry, tissue composition and mechanical properties of fetal to adult cryopreserved human heart valves. PLoS One. 2016;11:e0149020.
Van Meurs-Van Woezik H, Krediet P. Changes after birth in the tunica media and in the internal diameter of the aortic isthmus in normal newborns. J Anat. 1982;134:573–81.
Van Praagh R. The first Stella van Praagh memorial lecture: the history and anatomy of tetralogy of Fallot. Semin Thorac Cardiovasc Surg Pediatr Card Surg Annu. 2009;19–38.
Van Praagh S, Geva T, Lock JE, Nido PJ, Vance MS, Van Praagh R. Biatrial or left atrial drainage of the right superior vena cava: anatomic, morphogenetic, and surgical considerations–report of three new cases and literature review. Pediatr Cardiol. 2003;24:350–63.
van Vonderen JJ, Roest AA, Siew ML, Blom NA, van Lith JM, Walther FJ, Hooper SB, te Pas AB. Noninvasive measurements of hemodynamic transition directly after birth. Pediatr Res. 2014a;75:448–52.
van Vonderen JJ, Roest AA, Siew ML, Walther FJ, Hooper SB, te Pas AB. Measuring physiological changes during the transition to life after birth. Neonatology. 2014b;105:230–42.
van Vonderen JJ, Te Pas AB. The first breaths of life: imaging studies of the human infant during neonatal transition. Paediatr Respir Rev. 2015;16:143–6.
Verzi MP, McCulley DJ, De Val S, Dodou E, Black BL. The right ventricle, outflow tract, and ventricular septum comprise a restricted expression domain within the secondary/anterior heart field. Dev Biol. 2005;287:134–45.
Watanabe Y, Buckingham M. The formation of the embryonic mouse heart: heart fields and myocardial cell lineages. Ann N Y Acad Sci. 2010;1188:15–24.
Webb S, Qayyum SR, Anderson RH, Lamers WH, Richardson MK. Septation and separation within the outflow tract of the developing heart. J Anat. 2003;202:327–42.
Williams C, Kim SH, Ni TT, Mitchell L, Ro H, Penn JS, Baldwin SH, Solnica-Krezel L, Zhong TP. Hedgehog signaling induces arterial endothelial cell formation by repressing venous cell fate. Dev Biol. 2010;341:196–204.
Wu M, Peng S, Zhao Y. Inducible gene deletion in the entire cardiac conduction system using Hcn4-CreERT2 BAC transgenic mice. Genesis (New York, NY : 2000). 2014;52:134–40.
Xin M, Olson EN, Bassel-Duby R. Mending broken hearts: cardiac development as a basis for adult heart regeneration and repair. Nat Rev Mol Cell Biol. 2013;14:529–41.
Yamagishi T, Ando K, Nakamura H. Roles of TGFbeta and BMP during valvulo-septal endocardial cushion formation. Anat Sci Int. 2009;84:77–87.
Yang Y, Oliver G. Transcriptional control of lymphatic endothelial cell type specification. Adv Anat Embryol Cell Biol. 2014;214:5–22.
Yi SQ, Terayama H, Naito M, Hirai S, Alimujang S, Yi N, Tanaka S, Itoh M. Absence of the celiac trunk: case report and review of the literature. Clin Anat (New York, NY). 2008;21:283–6.
Yigit M, Kowalski W, Hutchon D, Pekkan K. Transition from fetal to neonatal circulation: modeling the effect of umbilical cord clamping. J Biomech. 2015;48(9):1662–70.
Yutzey KE, Kirby ML. Wherefore heart thou? Embryonic origins of cardiogenic mesoderm. Dev Dyn Off Publ Am Assoc Anatomists. 2002;223:307–20.
Zhang H, von Gise A, Liu Q, Hu T, Tian X, He L, Pu W, Huang X, He L, Cai CL, Camargo FD, Pu WT, Zhou B. Yap1 is required for endothelial to mesenchymal transition of the atrioventricular cushion. J Biol Chem. 2014;289:18681–92.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Dabbagh, A., Amini, A., Abdollahifar, MA., Saghafi, M.A. (2017). Cardiovascular System Embryology and Development. In: Dabbagh, A., Conte, A., Lubin, L. (eds) Congenital Heart Disease in Pediatric and Adult Patients. Springer, Cham. https://doi.org/10.1007/978-3-319-44691-2_2
Download citation
DOI: https://doi.org/10.1007/978-3-319-44691-2_2
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-44689-9
Online ISBN: 978-3-319-44691-2
eBook Packages: MedicineMedicine (R0)