
Abstract
Treatment options for autosomal dominant cerebellar ataxias are still scarce. Transcranial magnetic stimulation (TMS), a neuromodulation technique currently used for the treatment of depression, pain, vascular motor deficit, and posttraumatic stress disorder, can be a symptomatic treatment for ataxic patients. In this chapter, we reviewed current medical literature for the use of cerebellar TMS in spinocerebellar ataxias. Ten articles, including 170 ataxic patients, reported ataxia improvement after cerebellar TMS, with variable, but overall small effect sizes. This procedure appears to be safe since no severe side effect was reported. Additionally, cerebellar TMS can increase cerebellar blood flow, decrease oxidative stress, and decrease inhibition of the cerebellum over the contralateral motor cortex. However optimistic, these results still need to be better investigated in larger, longer, and more homogeneous trials.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
Autosomal dominant spinocerebellar ataxias (SCA) are a genetically inherited group of diseases related to degeneration of the cerebellum and cerebellar pathways (Klockgether et al. 2019). They can be divided into repeat expansions and non-repeat mutations SCAs (Klockgether et al. 2019). The prevalence of SCA, although challenging to assess, is estimated to be 2.7 cases per 100,000 individuals (Ruano et al. 2014). All SCAs are invariably progressive, and are responsible for premature disability, impairment in quality of life, and death (Diallo et al. 2018; Schmitz-Hübsch et al. 2010). Currently, there is no treatment capable of slow or cease SCAs evolution, and treatment is limited to symptomatic options. Even so, there is little treatment choices to effectively improve ataxic symptoms.
Pharmacological treatments with positive evidence for SCA treatment include riluzole, valproic acid, lithium carbonate, and varenicline, but evidence of quality of life improvement is scarce (Klockgether et al. 2019). Currently, rehabilitation therapies (physiotherapy, speech, and occupational therapy) remain the best studied treatment options for SCA patients. Although improvement in ataxia scores and in daily function after rehabilitation is well documented, size effects are small, and rehabilitation alone is often not capable of reestablishing functionality (Zesiewicz et al. 2018).
In light of this gap in ataxia treatment, transcranial magnetic stimulation (TMS) has emerged as a possible option treatment for SCA patients (França et al. 2018; Cury et al. 2020). TMS is a non-invasive, safe, and well-tolerated neuromodulation technique that can reach different areas of the nervous system and is capable of long-lasting benefits (Lefaucheur et al. 2020). Currently, TMS is successfully used in the treatment of pain, depression, motor recovery after stroke, Parkinson’s disease, spasticity, and posttraumatic stress disorders (Lefaucheur et al. 2020). Since the cerebellum is a neural structure abundantly connected to almost all the central nervous system, involved in the pathogenesis of SCA, endowed with neuromodulation properties, and accessible through non-invasive neuromodulation techniques, it is a possible hub of interest for TMS (Cury et al. 2020).
In this chapter, we reviewed studies using TMS aimed at the cerebellum for the treatment of ataxia. We searched for articles published between January 1, 1996, and October 18, 2021, on Medline (PubMed) using terms “Transcranial Magnetic Stimulation” AND “ataxia.” Articles selected for this review should demonstrate clinical results in patients with spinocerebellar ataxia after TMS targeting the cerebellum. We included only articles written in English.
2 Principles of Transcranial Magnetic Stimulation
TMS was introduced by Barker et al. in 1985, following the success of transcranial electric stimulation in modulating the motor cortex, as a less painful way to deliver the electric current to the brain (Barker et al. 1985). Based on the electromagnetic induction principle described in 1831 by Faraday, it can generate up to 2T magnetic field that lasts for 100 μs, and that is able to go unattenuated through scalp structures and then generate an electric field in the brain (Farzan 2014).
The electric field, and consequently the neural structures affected, can be shaped through several variables, such as coil geometry, current orientation, and intensity. Circular coils were the first types of coils used and allow a large, albeit not deep, area of cortical stimulation (Deng et al. 2013). For a more focal stimulation, figure-of-eight and double-cone coils are preferred, and these are also responsible for deeper stimulation fields (the former more than the latter). However, there is a rapid attenuation of the electric field in depth, which implies that more superficial structures receive most of the electric field (Deng et al. 2013). The stimulation of deeper structures, however, can increase depending on the delivered stimulation intensity, since the intensity of the induced current reduces with the square distance to the stimulation site (Deng et al. 2013). Regarding current orientation, it is known TMS stimulates preferentially axons than cell bodies, and the former are best stimulated by a parallel current. However, additionally to depth, shape, and intensity of stimulation, the effects of TMS must be accounted also for structures distant from the stimulation site, since TMS acts by circuit activation (Lefaucheur 2016). After axonal excitation by TMS, the changes in neuronal membrane spread in both orthodromic and antidromic directions, activating postsynaptic and presynaptic structures, respectively (Lefaucheur 2019). Although the effects of TMS are not exclusively consequence of local effects, but also distant circuit effects, it is important to precisely determine the stimulation target, and for this purpose the use of neuronavigation systems seems to be preferred over skull landmarks (Lefaucheur 2019).
There are several available TMS protocols. Single-pulse TMS (pulses separated by intervals ≥4 s) is largely used to measure neurophysiological variables, such as motor-evoked potentials, which reflect cortical excitability and the integrity of corticospinal pathways (Farzan 2014; Rodríguez-Labrada et al. 2018). Paired-pulse TMS consists of a conditioning stimulus followed by a test stimulus, and both stimuli are separated by an interstimulus interval (Rodríguez-Labrada et al. 2018). In the cerebellum, paired-pulse TMS can be used to measure cerebellar-brain inhibition (CBI) and cerebellar-brain facilitation (CBF) (Ugawa et al. 1995). CBI most likely reflects activation of cerebellar cortex Purkinje cells, which inhibits cerebellar facilitatory output through dentate-thalamic-cortical pathway to the contralateral cerebral motor cortex (Ugawa et al. 1995). The use of TMS in repetitive pulses—repetitive TMS (rTMS)—has a modulatory effect over neural structures possibly through long-term depression and long-term potentiation, and can generate plastic synaptic changes (Chen et al. 1997; Pascual-Leone et al. 1998). High frequency rTMS (≥5 Hz) is considered to be excitatory, while low frequency rTMS (≤1 Hz) is inhibitory. A type of rTMS, theta burst stimulation (TBS), can also be used for neuromodulation, and consists of 50 Hz bursts at 5 Hz delivered continuously (cTBS, considered inhibitory) or intermittently (iTBS, considered excitatory) (Suppa et al. 2016). This notion of inhibitory or excitatory is not always straightforward, since it can vary depending on the stimulation target and the prior state of circuits activation (Lefaucheur 2006; Fitzgerald et al. 2006). As dictated by the Bienenstock–Cooper–Munro model, if postsynaptic activity is high, it is more likely to be depressed; if it is low, it is more likely to be potentiated (Bienenstock et al. 1982). Therefore, the effects of rTMS are more dependent of baseline excitability levels than stimulation frequency (Daskalakis et al. 2006). This is probably one of the reasons why a typical plastic responses and altered excitability modifications to cortical stimulation have been reported in various neuropsychiatric diseases (Ueki et al. 2006; Quartarone et al. 2003; Pascual-Leone et al. 1996). Additionally, most of the knowledge about rTMS effects is derived from motor cortex studies; the effects after cerebellar rTMS are more limited. The size effect after one rTMS session is usually small, and short-lasting, but its effectiveness can be enhanced if patient is submitted to repeated sessions, especially in consecutive days (Valero-Cabré et al. 2008).
3 The Cerebellum as a Window to the Whole Brain
The cerebellum has emerged as an attractive and promising target for neuromodulation in neurological disorders over the last few years. Because cerebellar areas present several connections with important cortical and subcortical structures, the modulation of these different neuronal networks could potentially treat pathologic neuronal oscillations and thus influence motor and sensory integration (Fig. 1).

Schematic representation of cerebellar cortical and subcortical connections. Network model showing cerebellar connections to distant regions. The dentate nucleus receives inhibitory input from Purkinje cells and modulates other brain areas, including contralateral primary motor cortex (facilitatory tonus). There is intracortical inhibition between both motor cortices, which is related to maintaining the integrity of axial, and limbs movements. (Adapted from França et al. (2020))
Since the cerebral cortex is connected to the cerebellum only by polysynaptic circuits, and hence there are no monosynaptic connections, traditional techniques of anterograde and retrograde tracing cannot explore the topographic relationship between these two structures (Evarts and Thach 1969; Schmahmann and Pandya 1997; Strick 1985). Instead, inferences from deficits after specific lesions, as well as physiological and transneuronal tracing techniques, and functional neuroimaging could be used to investigate correlated areas.
Coherence is a spectral measure of the neural synchrony that can suggest communication between brain areas and can be measured using intrinsic low-frequency functional correlations by functional magnetic resonance imaging (MRI). Buckner et al. used this technique to create a complete functional map of the human cerebellum, and found functional connections between the cerebellum and the entire cerebral cortex, except perhaps primary visual and auditory cortices (Buckner et al. 2011). The cerebellum holds hubs of major functional brain networks, including Somatomotor Network, Default Mode Network, Limbic Network, Frontal Control Network, Ventral Attention Network, and Dorsal Attention Network (Buckner et al. 2011). Despite the previous concept of the cerebellum as a structure purely related to motor control, somatomotor regions occupy only a small portion of the cerebellum; functional connections to cerebral association networks are by far larger (Buckner et al. 2011). Moreover, the cerebellum has at least two complete homotopic maps of all aforementioned cortical networks: one inverted representation in the anterior lobe, and one mirrored upright representation in the posterior lobe. The size of a cerebellar region dedicated to a network is in fact proportionate to its representation in the cerebral cortex, meaning the largest cerebral networks are associated with the greatest representations in the cerebellum (Buckner et al. 2011). This evidence points to a comprehensive cortical representation in the cerebellum.
In addition to cortical areas, several brainstems structures receive cerebellar outputs: pontine reticular nucleus of the tegmentum, basilar pontine nuclei, pontine and medullary reticular formation, inferior olive, red nucleus, periaqueductal gray area, prerubral area, accessory oculomotor nuclei, and superior colliculus (Teune et al. 2000). The pontine reticular nucleus of the tegmentum is associated with motor learning (Takeichi et al. 2005), while the inferior olive plays a role not only in motor learning, but also in motor timing (De Zeeuw et al. 1998). Since the red nucleus receives fibers from the dentate nucleus and is connected to both motor cortex and spinal cord, it is associated with motor control, especially postural control (Herter et al. 2015).
The cerebellum is an important source of excitatory input to M1 via the dentato-thalamo-cortical pathway (Fig. 1) and when this input is diminished, there is a reduction in cortical excitability (increase in intracortical inhibition and decrease in intracortical facilitation) (Liepert et al. 2004). Injury in the dentato-thalamo-cortical pathway reduces excitability in the contralateral cortex (Rispal-Padel et al. 1981), whereas stimulation of the dentate nucleus increases cortical excitability and consequently promotes motor facilitation (Iwata and Ugawa 2005). Therefore, cerebellar neuromodulation techniques can modulate cortical excitability, since the cerebellum is a subcortical structure deputed to plastic mechanisms of motor learning (Ito 2008). However, it is not yet known whether cerebellar stimulation affects the dentate nucleus or Purkinje cells, structures with different roles in the cerebellum-thalamus-cortical activation.
4 Clinical Outcomes
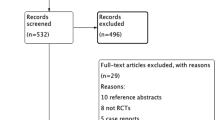
Presently, ten trials evaluated the effects of TMS in spinocerebellar ataxias (Table 1). Overall, 170 patients were evaluated, although additional causes of cerebellar ataxia other than spinocerebellar ataxia were included (multiple system atrophy (MSA-c), post-lesion ataxia, and idiopathic late-onset cerebellar atrophy). Five trials were double-blind and the remaining used open-label or single-blind designs. As for the TMS protocol, six studies used single-pulse TMS, one used low frequency rTMS, one used high frequency rTMS, one used cTBS, and one used iTBS.
All trials reported clinical improvement in ataxia after cerebellar TMS. Shimizu et al., in a preliminary open label study, demonstrated improvement in balance and gait after 21 sessions of single-pulse TMS in 4 SCA patients (Shimizu et al. 1999). Afterward, the same group used a similar protocol on 74 patients with cerebellar ataxia (etiologies not thoroughly described) using a double-blind design, also describing gait and balance improvement. Ihara et al. were the first to describe effects of single-pulse TMS on 20 ataxic patients (mixed etiologies) using a validated ataxia scale, the International Cerebellar Ataxia Rating Scale (ICARS). The most recent single-pulse TMS study included 20 spinocerebellar ataxia patients using a double-blind sham-controlled design and observed statistically significant improvement in the stance sub-score of the Scale for the Assessment and Rating of Ataxia (SARA) (Manor et al. 2019). Dang et al. were the first to use cerebellar rTMS in one SCA6 patient, reporting great improvement in SARA and ICARS (Dang et al. 2019). After 18 months of the last session, this improvement was not only sustained, but increased. The largest rTMS trial included 24 patients (9 SCA3 patients, 8 MSA-c patients, and 7 post-lesion ataxia patients) with a double-blind sham-controlled crossover design and applied five low-frequency rTMS sessions using a deep reaching (double-cone) coil (França et al. 2020). This trial was the first to use neuronavigation to better locate the stimulation target (dentate nucleus contralateral to the most ataxic hemibody) and reported a significant improvement in ataxia using SARA and ICARS comparing active and sham stimulations. Two other rTMS trials used TBS protocols and reported improvement in ataxia (iTBS) (Sanna et al. 2020), and more specifically in ataxic dysarthria (cTBS) (Lin et al. 2022).
It is important to highlight that, albeit encouraging, improvements reported in those studies demonstrated highly variable size effects. In the study conducted by Shiga et al., there was 31% decrease in time to walk 10 m, 18% decrease in the number of steps to walk 10 m, and 638% increase in number of Tandem steps achieved, comparing before TMS and after TMS in the active group (Shiga et al. 2002). Ihara et al. reported 5.1 points reduction (improvement) in ICARS score (ranging 0–100) comparing before and after TMS (Ihara et al. 2005). In another study evaluating 20 SCA patients, there was a 3.9 points reduction (improvement) in SARA score (ranging 0–40) after 20 TMS sessions comparing scores from baseline and 1 month follow up (Manor et al. 2019). In the only study using low-frequency rTMS, there was an improvement of 3.3 points in SARA score, and 5 points in ICARS score (França et al. 2020). Finally, in the last published trial, which included 6 SCA38 patients, there was an improvement of 4.4 points in the Modified International Cooperative Ataxia Rating Scale (MICARS, ranging 0–120) (Sanna et al. 2020).
The medical literature up to this point endorses the safety of cerebellar TMS. No clinical study so far reported severe side effects. Some mild side effects included headache or local discomfort, and were all self-limited (França et al. 2020).
5 Neurophysiological and Biochemical Outcomes
In addition to clinical outcomes, several studies also included neurophysiological outcomes, which could help us better understand the pathophysiology behind the clinical efficacy of cerebellar TMS for spinocerebellar ataxias. There is currently evidence pointing to changes of brain blood flow, oxidative stress markers and CBI after cerebellar TMS in ataxic patients.
After 21 sessions of single-pulse TMS over the cerebellum (four patients), single photon emission computed tomography revealed significantly increased blood flow in the cerebellar hemisphere, putamen, and pons, compared to the measures taken before TMS, which may be correlated with the clinical improvement (Shimizu et al. 1999). These findings were then corroborated by a two future clinical trials using single-pulse TMS (Shiga et al. 2002; Ihara et al. 2005). Previous reports showed increase in brain blood flow of normal subjects after TMS both in the stimulated area and associated regions (Siebner et al. 1998).
Ihara et al. evaluated several cerebrospinal fluidbiochemical oxidative stress parameters in 20 patients before and after 24 sessions of single-pulse TMS (Ihara et al. 2005). This is an interesting investigation, since there is evidence pointing to oxidative stress as a pathological mechanism of SCA (Torres-Ramos et al. 2018; Guevara-García et al. 2012; Araujo et al. 2011). Ataxic patients had higher oxidative stress compared to controls, and its levels were inversely correlated with clinical severity (Ihara et al. 2005). This finding suggests decrease of oxidative stress as a possible mechanism underlying the clinical improvement after TMS.
In a single-case study, Farzan et al. examined CBI in a patient with idiopathic late-onset cerebellar ataxia (precise diagnosis unknown). After 21 sessions of single-pulse cerebellar TMS, in addition to the clinical improvement, there was CBI decrease, and this reduction persisted for 6 months after TMS interruption (Farzan et al. 2013). This might suggest reduction of cerebellar tonic inhibition over the cerebral cortex as another mechanism responsible for clinical improvement. Moreover, decrease in the tonic Purkinje cell inhibition may increase vestibular nuclei activity, which could contribute to balance improvement seen in some ataxic patients after TMS (Rub 2002; Shin et al. 2011).
6 Targets and Coils
Location of coil placement, coils shapes, and sizes varied greatly among studies, regardless of the positive clinical outcomes.
The cerebellum is not a homogeneous structure, and is composed of several types of cells, fibers, and nuclei. Some of these components have opposite final effects. For instance, the dentate nucleus is responsible for the excitatory output to the thalamus, but Purkinje cells inhibit the dentate nucleus. Therefore, distinguishing modulation of Purkinje cells and dentate nucleus is paramount, considering these two structures have opposite roles in cerebellar effects over the motor cortex. However, the determination of the exact brain area being influenced by the induced electric current is a major inherent limitation of non-invasive modulation techniques (Lefaucheur et al. 2020). Most likely, more than one structure is being stimulated simultaneously, and that makes even more difficult to determine which stimulated structure is actually responsible for the final result. This issue is even more complex if we add to the equation the concomitant activation of distant parts of the network, away from the stimulated target (Al-Fatly et al. 2019; Horn et al. 2019).
To comprehend more about this issue, it is important to understand about different coils characteristics. Coils can vary in shapes (circular, figure-of-eight, double-cone, etc.), and sizes (coil diameter). Circular coils are considered superficial coils and can reach a large stimulation area—the larger the coil diameter, the larger the area stimulated. Figure-of-eight and double-cone coils, on the other hand, are deeper reaching coils, and stimulate smaller areas (Deng et al. 2013). The double-cone coil is considered to reach structures as deep as the foot motor cortex (Galhardoni et al. 2019). Since the dentate nucleus lies as deep from the skull surface as the foot motor cortex, it is safe to say double-cone coils are able to reach it (Cury et al. 2015; Hardwick et al. 2014). However, between the dentate nucleus and the skull surface lie Purkinje cells on the cerebellar cortex that could be also modulated by the magnetic field. The electric field diminishes as a function of coil distance; hence, it is possible that Purkinje and dentate nucleus, in addition to other cerebellar structures beneath the coil and its lateral wings, are concurrently modulated at different intensities (Hayward et al. 2007). The insula lies at a similar depth from the scalp as the dentate nucleus (4.5–5.0 cm). Interestingly, TMS insula studies found antinociceptive effects only when using double-cone coils (Ciampi de Andrade et al. 2012; Lenoir et al. 2018). More importantly, this analgesic effect was clinically equivalent to the effect obtained by direct stimulation of the posterior insula using electrodes during electroencephalography in patients with refractory epilepsy (Ciampi de Andrade et al. 2012). These data point to a relatively good specificity and target accuracy when performing TMS with a double-cone coil (Deng et al. 2013). Another study comparing TMS coils found no changes in cerebellar-brain inhibition after cerebellar 1 Hz repetitive transcranial magnetic stimulation with superficial figure-of-eight coil but only with deep-reaching coils (Hardwick et al. 2014). Cury et al. previously reported improvement in the SARA score after cerebellar rTMS using double-cone coil in one post-lesion ataxic patient, and after this same patient received a dentate nucleus Deep Brain Stimulation implant, the improvement in SARA was identical, which would argue in favor of the dentate nucleus as responsible for the clinical improvement (Cury et al. 2015; Teixeira et al. 2015). Currently, we do not have an answer for this conundrum. The most probable explanation might involve effects from multiple structures acting in resonance.
Another possible mechanism is derived from studies of neuromodulation for Parkinson’s disease (PD). In PD patients, it is well known that beta oscillations (13–30 Hz) are greatly enhanced, and its presence is correlated with parkinsonian symptoms (rigidity and bradykinesia) (Little and Brown 2014). Levodopa therapy and high frequency Deep Brain Stimulation can reduce beta oscillations, and improve parkinsonian symptoms. If there is a specific diseased cerebellar activity that correlates with ataxic symptoms, this activity could be disrupted by TMS, and this disruption could be responsible for the clinical benefit observed across studies. However, more studies are required to verify this hypothesis.
7 Little Brain, Big Expectations: A Glimpse into the Future
Albeit homogeneously reporting clinical improvement, studies investigating cerebellar TMS in SCA patients are wildly heterogeneous regarding coil type, frequency, intensity, location, number of sessions, follow-up, evaluation tools, and additional outcomes (França et al. 2018). It is therefore important to corroborate these finding with larger studies using the same stimulation parameters, and with longer follow-ups. Moreover, the fact that all studies reported positive clinical outcomes, but chose different types of TMS, makes us wonder what would be the best TMS setting. More likely, there is not a single answer for all SCA patients since there are different pathological mechanisms depending on the SCA type (Klockgether et al. 2019). In that line, studies should try to include homogeneous populations—a single type of SCA—or post-hoc analysis considering the molecular diagnosis. The main difficulty lies in the heterogeneous rarity of SCAs. SCA3, for instance, is one of the most common SCA, while SCA38 can only be observed in three family clusters (Klockgether et al. 2019; Sanna et al. 2020; Gazulla et al. 2020; Borroni et al. 2016). With this low prevalence in mind, and considering TMS effects are time-limited, multicentric crossover trials seem to be the best path.
Important progress should also be directed to better understand cerebral activity in SCAs. In Parkinson’s disease, it is now known that the excess of beta oscillation is correlated with rigidity and bradykinesia (Kühn et al. 2009). Therefore, it is considered an oscillopathy. Both dopaminergic medications and Deep Brain Stimulation therapy can overwrite this pathological activity, and improve symptoms. In ataxia there could be a similar diseased-dominant frequency correlated with the symptoms, and this could potentially be overwritten by neuromodulation. Neurophysiological studies, and, in the future, studies using closed-loop DBS could aid in this matter (Arlotti et al. 2016).
Effect size reported in trials of cerebellar TMS studies for SCA are variable, and most are small. This is a constant in SCA clinical trials so far, regardless of the treatment approach. Romano et al. tested the efficacy of riluzole versus placebo in 55 patients (different types of SCAs and Friedreich ataxia) and found a decrease in SARA scores by 1.02 points in patients (Romano et al. 2015). Another group studied valproic acid in a smaller sample of 12 SCA3 patients and reported a 2.05-point decrease in SARA scores (Lei et al. 2016). However, although an one point decrease in SARA, a scale with a 40-point range, may seem small, it was considered to be clinically relevant in previous studies (Klockgether et al. 2019; Schmitz-Hübsch et al. 2010). Clinical trials combining treatment options (for instance, neuromodulation and physiotherapy) seem to be the natural next step, so we can best evaluate if the combination of treatments could enhance effect size.
An important shortcoming of most trials so far is the absence of quality-of-life measures. It is impossible to understand the degree of impact a certain scale improvement has in a patient’s life if quality of life is not evaluated. The only study so far in which quality of life was assessed did not report significant improvement after TMS (França et al. 2020). However, follow-up might have been insufficient to detect real improve in day-to-day activities. It is vital that future trials include quality of life in its outcomes (perhaps as main outcome) and have appropriate follow-ups.
To date, there are no studies comparing TMS to other non-invasive neuromodulation techniques in ataxic patients. In theory, TMS induces a more focal and deeper electric field when compared to transcranial direct current stimulation (tDCS), and can activate specific neural circuits (Di Lazzaro and Rothwell 2014). Comparative studies for pain and upper limb recovery after stroke showed superiority of TMS over tDCS in chosen protocols (Attal et al. 2016; Doris Miu et al. 2020). However, tDCS is a simpler technique, and can be used at bedside, which widens its use possibilities. Studies comparing TMS to tDCS in ataxic patients are needed.
Another gap that needs to be filled is the selection of good responders. Almost all trials up to this point included patients with different ataxia types. Shiga et al. divided patients into two groups—cerebellar type (hereditary and sporadic cerebellar atrophy, including SCA6), and olivopontocerebellar atrophy (OPCA; MSA, SCA1, SCA3)—and reported better outcomes in patients from the OPCA group (Shiga et al. 2002). França et al. included patients with MSA, SCA3, and post-lesion ataxia, and found best improvement in MSA patients (França et al. 2020). Despite these post-hoc analysis results, there is still a paucity of information regarding differences between good and bad responders. Does cerebellar connectivity influence clinical response? Or is it a matter or cerebellar atrophy? A previous study found no correlation between cerebellar volume and clinical outcome after low frequency rTMS (França et al. 2020). Is it possible that integrity of superior cerebellar peduncle (cerebellar efferent pathway) plays a role in clinical improvement after cerebellar TMS? Or are there biochemical differences responsible for the different outcomes? Manto et al. discussed the concept of cerebellar reserve—how much of the cerebellum cells and synapses are still intact—as a way a measure the potential of improvement after cerebellar non-invasive modulation (Manto et al. 2021). With that in mind, perhaps there are no good or bad responders, but good or bad treatment timings. Many questions still need to be answered before we can understand which patient profile could benefit the most.
8 Conclusions
There is evidence to suggest cerebellar TMS is safe and can reduce ataxic symptoms in SCA patients. Additional evidence suggests it can also increase cerebellar brain blood flow, decrease brain oxidative stress, and decrease CBI. Although encouraging, these results should be further explored in larger, more homogeneous trials, and trials with longer follow-ups. The pathophysiological mechanism of this improvement also should be better explored, as well as characterization of good and bad responders.
References
Al-Fatly B, Ewert S, Kübler D, Kroneberg D, Horn A, Kühn AA. Connectivity profile of thalamic deep brain stimulation to effectively treat essential tremor. Brain. 2019;142(10):3086–98.
Araujo J, Breuer P, Dieringer S, Krauss S, Dorn S, Zimmermann K, et al. FOXO4-dependent upregulation of superoxide dismutase-2 in response to oxidative stress is impaired in spinocerebellar ataxia type 3. Hum Mol Genet. 2011;20(15):2928–41.
Arlotti M, Rosa M, Marceglia S, Barbieri S, Priori A. The adaptive deep brain stimulation challenge. Parkinsonism Relat Disord. 2016;28:12–7.
Attal N, Ayache SS, Ciampi De Andrade D, Mhalla A, Baudic S, Jazat F, et al. Repetitive transcranial magnetic stimulation and transcranial direct-current stimulation in neuropathic pain due to radiculopathy: a randomized sham-controlled comparative study. Pain. 2016;157(6):1224–31.
Barker AT, Jalinous R, Freeston IL. Non-invasive magnetic stimulation of human motor cortex. Lancet. 1985;325(8437):1106–7.
Bienenstock EL, Cooper LN, Munro PW. Theory for the development of neuron selectivity: orientation specificity and binocular interaction in visual cortex. J Neurosci. 1982;2(1):32–48.
Borroni B, Di Gregorio E, Orsi L, Vaula G, Costanzi C, Tempia F, et al. Clinical and neuroradiological features of spinocerebellar ataxia 38 (SCA38). Parkinsonism Relat Disord. 2016;28:80–6.
Buckner RL, Krienen FM, Castellanos A, Diaz JC, Yeo BTT. The organization of the human cerebellum estimated by intrinsic functional connectivity. J Neurophysiol. 2011;106(5):2322–45.
Chen R, Classen J, Gerloff C, Celnik P, Wassermann EM, Hallett M, et al. Depression of motor cortex excitability by low-frequency transcranial magnetic stimulation. Neurology. 1997;48(5):1398–403.
Ciampi de Andrade D, Galhardoni R, Pinto LF, Lancelotti R, Rosi J, Marcolin MA, et al. Into the Island: a new technique of non-invasive cortical stimulation of the insula. Neurophysiol Clin. 2012;42(6):363–8.
Cury RG, Teixeira MJ, Galhardoni R, Barboza VR, Alho E, Seixas CM, et al. Neuronavigation-guided transcranial magnetic stimulation of the dentate nucleus improves cerebellar ataxia: a sham-controlled, double-blind n = 1 study. Parkinsonism Relat Disord. 2015;21(8):999–1001.
Cury RG, França C, Reis Barbosa E, Jacobsen Teixeira M, Ciampi de Andrade D. Little brain, big expectations. Brain Sci. 2020;10(12):944.
Dang G, Su X, Zhou Z, Che S, Zeng S, Chen S, et al. Beneficial effects of cerebellar rTMS stimulation on a patient with spinocerebellar ataxia type 6. Brain Stimul. 2019;12(3):767–9.
Daskalakis ZJ, Möller B, Christensen BK, Fitzgerald PB, Gunraj C, Chen R. The effects of repetitive transcranial magnetic stimulation on cortical inhibition in healthy human subjects. Exp Brain Res. 2006;174(3):403–12.
De Zeeuw CI, Simpson JI, Hoogenraad CC, Galjart N, Koekkoek SK, Ruigrok TJ. Microcircuitry and function of the inferior olive. Trends Neurosci. 1998;21(9):391–400.
Deng Z-D, Lisanby SH, Peterchev AV. Electric field depth–focality tradeoff in transcranial magnetic stimulation: simulation comparison of 50 coil designs. Brain Stimul. 2013;6(1):1–13.
Di Lazzaro V, Rothwell JC. Corticospinal activity evoked and modulated by non-invasive stimulation of the intact human motor cortex. J Physiol. 2014;592(19):4115–28.
Diallo A, Jacobi H, Cook A, Labrum R, Durr A, Brice A, et al. Survival in patients with spinocerebellar ataxia types 1, 2, 3, and 6 (EUROSCA): a longitudinal cohort study. Lancet Neurol. 2018;17(4):327–34.
Doris Miu KY, Kok C, Leung SS, Chan EYL, Wong E. Comparison of repetitive transcranial magnetic stimulation and transcranial direct current stimulation on upper limb recovery among patients with recent stroke. Ann Rehabil Med. 2020;44(6):428–37.
Evarts EV, Thach WT. Motor mechanisms of the CNS: cerebrocerebellar interrelations. Annu Rev Physiol. 1969;31(1):451–98.
Farzan F. Single-pulse transcranial magnetic stimulation (TMS) protocols and outcome measures. In: Rotenberg A, Horvath JC, Pascual-Leone A, editors. Transcranial magnetic stimulation [Internet]. New York: Springer New York; 2014 [cited 2020 Jan 10]. p. 69–115. Available from: http://springerlink.bibliotecabuap.elogim.com/10.1007/978-1-4939-0879-0_5.
Farzan F, Wu Y, Manor B, Anastasio EM, Lough M, Novak V, et al. Cerebellar TMS in treatment of a patient with cerebellar ataxia: evidence from clinical, biomechanics and neurophysiological assessments. Cerebellum. 2013;12(5):707–12.
Fitzgerald P, Fountain S, Daskalakis Z. A comprehensive review of the effects of rTMS on motor cortical excitability and inhibition. Clin Neurophysiol. 2006;117(12):2584–96.
França C, de Andrade DC, Teixeira MJ, Galhardoni R, Silva V, Barbosa ER, et al. Effects of cerebellar neuromodulation in movement disorders: a systematic review. Brain Stimul. 2018;11(2):249–60.
França C, de Andrade DC, Silva V, Galhardoni R, Barbosa ER, Teixeira MJ, et al. Effects of cerebellar transcranial magnetic stimulation on ataxias: a randomized trial. Parkinsonism Relat Disord. 2020;80:1–6.
Galhardoni R, Aparecida da Silva V, García-Larrea L, Dale C, Baptista AF, Barbosa LM, et al. Insular and anterior cingulate cortex deep stimulation for central neuropathic pain: disassembling the percept of pain. Neurology. 2019. https://doi.org/10.1212/WNL.0000000000007396.
Gazulla J, Orduna-Hospital E, Benavente I, Rodríguez-Valle A, Osorio-Caicedo P, Alvarez-de Andrés S, et al. Contributions to the study of spinocerebellar ataxia type 38 (SCA38). J Neurol. 2020;267(8):2288–95.
Guevara-García M, Gil-del Valle L, Velásquez-Pérez L, García-Rodríguez JC. Oxidative stress as a cofactor in spinocerebellar ataxia type 2. Redox Rep. 2012;17(2):84–9.
Hardwick RM, Lesage E, Miall RC. Cerebellar transcranial magnetic stimulation: the role of coil geometry and tissue depth. Brain Stimul. 2014;7(5):643–9.
Hayward G, Mehta MA, Harmer C, Spinks TJ, Grasby PM, Goodwin GM. Exploring the physiological effects of double-cone coil TMS over the medial frontal cortex on the anterior cingulate cortex: an H2(15)O PET study. Eur J Neurosci. 2007;25(7):2224–33.
Herter TM, Takei T, Munoz DP, Scott SH. Neurons in red nucleus and primary motor cortex exhibit similar responses to mechanical perturbations applied to the upper-limb during posture. Front Integr Neurosci [Internet]. 2015 [cited 2017 Oct 27];9. Available from: https://www.frontiersin.org/articles/10.3389/fnint.2015.00029/full.
Horn A, Wenzel G, Irmen F, Huebl J, Li N, Neumann W-J, et al. Deep brain stimulation induced normalization of the human functional connectome in Parkinson’s disease. Brain. 2019;142(10):3129–43.
Ihara Y, Takata H, Tanabe Y, Nobukuni K, Hayabara T. Influence of repetitive transcranial magnetic stimulation on disease severity and oxidative stress markers in the cerebrospinal fluid of patients with spinocerebellar degeneration. Neurol Res. 2005;27(3):310–3.
Ito M. Control of mental activities by internal models in the cerebellum. Nat Rev Neurosci. 2008;9(4):304–13.
Iwata NK, Ugawa Y. The effects of cerebellar stimulation on the motor cortical excitability in neurological disorders: a review. Cerebellum. 2005;4(4):218–23.
Kawamura K, Etoh S, Shimodozono M. Transcranial magnetic stimulation for diplopia in a patient with spinocerebellar ataxia type 6: a case report. 2018;20(5):15. https://doi.org/10.1186/s40673-018-0094-x.eCollection.
Klockgether T, Mariotti C, Paulson HL. Spinocerebellar ataxia. Nat Rev Dis Primers. 2019;5(1):24.
Kühn AA, Tsui A, Aziz T, Ray N, Brücke C, Kupsch A, et al. Pathological synchronisation in the subthalamic nucleus of patients with Parkinson’s disease relates to both bradykinesia and rigidity. Exp Neurol. 2009;215(2):380–7.
Lefaucheur JP. The use of repetitive transcranial magnetic stimulation (rTMS) in chronic neuropathic pain. Neurophysiol Clin. 2006;36(3):117–24.
Lefaucheur J-P. Cortical neurostimulation for neuropathic pain: state of the art and perspectives. Pain. 2016;157(Suppl 1):S81–9.
Lefaucheur J-P. Transcranial magnetic stimulation. In: Handbook of clinical neurology [Internet]. Elsevier; 2019 [cited 2020 Feb 23]. p. 559–80. Available from: https://linkinghub.elsevier.com/retrieve/pii/B9780444640321000370.
Lefaucheur J-P, Aleman A, Baeken C, Benninger DH, Brunelin J, Di Lazzaro V, et al. Evidence-based guidelines on the therapeutic use of repetitive transcranial magnetic stimulation (rTMS): an update (2014–2018). Clin Neurophysiol. 2020;131(2):474–528.
Lei L-F, Yang G-P, Wang J-L, Chuang D-M, Song W-H, Tang B-S, et al. Safety and efficacy of valproic acid treatment in SCA3/MJD patients. Parkinsonism Relat Disord. 2016;26:55–61.
Lenoir C, Algoet M, Mouraux A. Deep continuous theta burst stimulation of the operculo-insular cortex selectively affects Aδ-fibre heat pain. J Physiol Lond. 2018;596(19):4767–87.
Liepert J, Kucinski T, Tüscher O, Pawlas F, Bäumer T, Weiller C. Motor cortex excitability after cerebellar infarction. Stroke. 2004;35(11):2484–8.
Lin Q, Chang Y, Liu P, Jones JA, Chen X, Peng D, et al. Cerebellar continuous theta burst stimulation facilitates auditory–vocal integration in spinocerebellar ataxia. Cereb Cortex. 2022;32(3):455–66.
Little S, Brown P. The functional role of beta oscillations in Parkinson’s disease. Parkinsonism Relat Disord. 2014;20:S44–8.
Manor B, Greenstein PE, Davila-Perez P, Wakefield S, Zhou J, Pascual-Leone A. Repetitive transcranial magnetic stimulation in spinocerebellar ataxia: a pilot randomized controlled trial. Front Neurol [Internet]. 2019 [cited 2020 May 14];10. Available from: https://www.frontiersin.org/article/10.3389/fneur.2019.00073/full.
Manto M, Kakei S, Mitoma H. The critical need to develop tools assessing cerebellar reserve for the delivery and assessment of non-invasive cerebellar stimulation. Cerebellum Ataxias. 2021;8(1):2.
Pascual-Leone A, Catala MD, Pascual AP-L. Lateralized effect of rapid-rate transcranial magnetic stimulation of the prefrontal cortex on mood. Neurology. 1996;46(2):499–502.
Pascual-Leone A, Tormos JM, Keenan J, Tarazona F, Cañete C, Catalá MD. Study and modulation of human cortical excitability with transcranial magnetic stimulation. J Clin Neurophysiol. 1998;15(4):333–43.
Quartarone A, Bagnato S, Rizzo V, Siebner HR, Dattola V, Scalfari A, et al. Abnormal associative plasticity of the human motor cortex in writer’s cramp. Brain. 2003;126(Pt 12):2586–96.
Rispal-Padel L, Cicirata F, Pons C. Contribution of the dentato-thalamo-cortical system to control of motor synergy. Neurosci Lett. 1981;22(2):137–44.
Rodríguez-Labrada R, Velázquez-Pérez L, Ziemann U. Transcranial magnetic stimulation in hereditary ataxias: diagnostic utility, pathophysiological insight and treatment. Clin Neurophysiol. 2018;129(8):1688–98.
Romano S, Coarelli G, Marcotulli C, Leonardi L, Piccolo F, Spadaro M, et al. Riluzole in patients with hereditary cerebellar ataxia: a randomised, double-blind, placebo-controlled trial. Lancet Neurol. 2015;14(10):985–91.
Ruano L, Melo C, Silva MC, Coutinho P. The global epidemiology of hereditary ataxia and spastic paraplegia: a systematic review of prevalence studies. Neuroepidemiology. 2014;42(3):174–83.
Rub U. Spinocerebellar ataxia type 3 (Machado-Joseph disease): severe destruction of the lateral reticular nucleus. Brain. 2002;125(9):2115–24.
Sanna A, Follesa P, Puligheddu M, Cannas A, Serra M, Pisu MG, et al. Cerebellar continuous theta burst stimulation reduces levodopa-induced dyskinesias and decreases serum BDNF levels. Neurosci Lett. 2020;716:134653.
Schmahmann JD, Pandya DN. The cerebrocerebellar system. Int Rev Neurobiol [Internet]. Elsevier. 1997 [cited 2021 Feb 11]:31–60. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0074774208603463.
Schmitz-Hübsch T, Fimmers R, Rakowicz M, Rola R, Zdzienicka E, Fancellu R, et al. Responsiveness of different rating instruments in spinocerebellar ataxia patients. Neurology. 2010;74(8):678–84.
Shiga Y, Tsuda T, Itoyama Y, Shimizu H, Miyazawa K-I, Jin K, et al. Transcranial magnetic stimulation alleviates truncal ataxia in spinocerebellar degeneration. J Neurol Neurosurg Psychiatry. 2002;72(1):124–6.
Shimizu H, Tsuda T, Shiga Y, Miyazawa K, Onodera Y, Matsuzaki M, et al. Therapeutic efficacy of transcranial magnetic stimulation for hereditary spinocerebellar degeneration. Tohoku J Exp Med. 1999;189(3):203–11.
Shin M, Moghadam SH, Sekirnjak C, Bagnall MW, Kolkman KE, Jacobs R, et al. Multiple types of cerebellar target neurons and their circuitry in the vestibulo-ocular reflex. J Neurosci. 2011;31(30):10776–86.
Siebner HR, Willoch F, Peller M, Auer C, Boecker H, Conrad B, et al. Imaging brain activation induced by long trains of repetitive transcranial magnetic stimulation. Neuroreport. 1998;9(5):943–8.
Strick PL. How do the basal ganglia and cerebellum gain access to the cortical motor areas? Behav Brain Res. 1985;18(2):107–23.
Suppa A, Huang Y-Z, Funke K, Ridding MC, Cheeran B, Di Lazzaro V, et al. Ten years of theta burst stimulation in humans: established knowledge, unknowns and prospects. Brain Stimul. 2016;9(3):323–35.
Takeichi N, Kaneko CRS, Fuchs AF. Discharge of monkey nucleus reticularis tegmenti pontis neurons changes during saccade adaptation. J Neurophysiol. 2005;94(3):1938–51.
Teixeira MJ, Cury RG, Galhardoni R, Barboza VR, Brunoni AR, Alho E, et al. Deep brain stimulation of the dentate nucleus improves cerebellar ataxia after cerebellar stroke. Neurology. 2015;85(23):2075–6.
Teune TM, van der Burg J, van der Moer J, Voogd J, Ruigrok TJ. Topography of cerebellar nuclear projections to the brain stem in the rat. Prog Brain Res. 2000;124:141–72.
Torres-Ramos Y, Montoya-Estrada A, Cisneros B, Tercero-Pérez K, León-Reyes G, Leyva-García N, et al. Oxidative stress in spinocerebellar ataxia type 7 is associated with disease severity. Cerebellum. 2018;17(5):601–9.
Ueki Y, Mima T, Ali Kotb M, Sawada H, Saiki H, Ikeda A, et al. Altered plasticity of the human motor cortex in Parkinson’s disease. Ann Neurol. 2006;59(1):60–71.
Ugawa Y, Uesaka Y, Terao Y, Hanajima R, Kanazawa I. Magnetic stimulation over the cerebellum in humans. Ann Neurol. 1995;37(6):703–13.
Valero-Cabré A, Pascual-Leone A, Rushmore RJ. Cumulative sessions of repetitive transcranial magnetic stimulation (rTMS) build up facilitation to subsequent TMS-mediated behavioural disruptions. Eur J Neurosci. 2008;27(3):765–74.
Zesiewicz TA, Wilmot G, Kuo S-H, Perlman S, Greenstein PE, Ying SH, et al. Comprehensive systematic review summary: treatment of cerebellar motor dysfunction and ataxia: report of the Guideline Development, Dissemination, and Implementation Subcommittee of the American Academy of Neurology. Neurology. 2018;90(10):464–71.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2023 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this chapter

Cite this chapter
França, C., Cury, R.G. (2023). Cerebellar Transcranial Magnetic Stimulation in Cerebellar Ataxias. In: Soong, Bw., Manto, M., Brice, A., Pulst, S.M. (eds) Trials for Cerebellar Ataxias. Contemporary Clinical Neuroscience. Springer, Cham. https://doi.org/10.1007/978-3-031-24345-5_21
Download citation
DOI: https://doi.org/10.1007/978-3-031-24345-5_21
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-24344-8
Online ISBN: 978-3-031-24345-5
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)