
Abstract
The quintessential example of evolutionary convergence is that between the shark, ichthyosaur, and dolphin. Although not closely related, the three exemplar taxa have independently evolved adaptations in morphology, physiology, and behavior that result in concomitant levels of performance that meet the requirements associated with operating in a dense, viscous, and thermally conductive marine environment. These apex marine predators display a remarkable amount of homoplasy. All three taxa have developed streamlined fusiform bodies to reduce drag when swimming. The position, type, and morphology of the control surfaces (i.e., fins, flippers, flukes) are similar for the convergent taxa. The control surfaces have different internal support structures, but function similarly to generate lift forces for stability and maneuverability. The main departure in control surface design among the three taxa is that dolphins lack pelvic fins. For dolphins, the loss of pelvic appendages is directly related to the possession of horizontally oriented caudal flukes, which perform double duty as a propulsive device and posterior stabilizer for trim control. The flukes of dolphins and caudal fins of ichthyosaurs and sharks have a lunate shape that function as an oscillating wing to generate high efficiency, lift-based thrust for high-speed swimming. The three convergent taxa are homeothermic, with a body temperature above that of the water in which they live. The advantages of an elevated body temperature are the attainment of higher maximum swimming speeds, longer and faster sustained swimming speeds, improved digestion, brain heating, and enhanced visual acuity. The convergence of the shark, ichthyosaur, and dolphin with respect to morphology, physiology, and locomotor performance reflects similar selective pressures imposed by the physical fluid environment that have dictated the independent evolutionary trajectories of these high-performance marine predators.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
15.1 Introduction
In terms of evolution, similarity is expressed in two different ways, resulting in homology (evolutionary similarity) and analogy (functional similarity). Homology results from similarity due to common descent and is expressed as derived traits within monophyletic groups that share common inheritance and phylogeny (Haas & Simpson, 1946; Chang & Kim, 1996; Wake et al., 2011). Homology is used to generate phylogenies to show the degree of relatedness among organisms or trace evolutionary pathways back through deep time. Analogy reflects similarity of function between different structures (Haas & Simpson, 1946). When analogy is combined with homoplasy, there is not only similarity in function but also similarity in appearance arrived at from multiple independent origins. Both analogy and homoplasy point to natural selection as the driver of the separate evolution of similar structures among lineages, but homology need not imply the operation of selective mechanisms (Gould, 2002).
Darwin (1859) considered analogy to be associated with “adaptive characters” that were beneficial to the individual that had been subjected to selection, but to be “valueless to the systematist.” Much of evolutionary theory was driven by Darwin’s idea of common descent traced through homologous characters, putting it at odds with the perceived importance and utility of analogous features, particularly where homoplasy was involved (Wake, 1991; Brooks, 1996). Homoplasy was viewed as false homology (Wake, 1991). Analogy and homoplasy were considered barriers to a full comprehension of evolutionary trajectories. However, homologous features and shared genetic attributes provide only an understanding of the historical connections among organisms and their adaptations but cannot be employed to address why particular adaptations have evolved. Analogy and homoplasy provide clues about factors leading to the origin of particular adaptations and their association with the ecology of the organisms under consideration.
The combination of similar functions (analogy) and designs (homoplasy) resulting from the independent evolution of unrelated lineages leads to convergence (Haas & Simpson, 1946; Wake, 1991; Wake et al., 2011; Stayton, 2015). In this case, convergence manifests as analogy and homoplasy without homology, and thus stems from different developmental-structural origins (Powell, 2007). The incorporation of shape and function differentiates convergence from mimicry (Haas & Simpson, 1946). Convergence differs from parallelism in that the latter entails analogy, homoplasy, and homology that depend upon the same developmental genetic mechanisms (Wake et al., 2011). Both convergence and parallelism yield similarity as a result of common selective pressures imposed by the interaction of organisms with their biotic and abiotic environments. Convergence and parallelism permit testing of adaptive hypotheses (Larson & Losos, 1996; Stayton, 2015). Comparative studies have used the concepts of convergence and parallelism to elucidate the mechanisms and constraints that underpin directed phylogenetic changes (Wake et al., 2011). The characters that evolve from unrelated or distantly related groups demonstrate similar adaptive solutions to common selective pressures imposed by similar relatively stable environmental constraints. Convergence is one of the most powerful lines of evidence for how and why natural selection drives form toward particular functional adaptations that are specific to similar ecological circumstances in varied localities in unrelated groups of organisms (Patterson, 1988; Wake, 1991).
It is not uncommon for multiple evolutionary pathways in unrelated groups to arrive independently at a common solution to an environmental challenge. Convergence not only demonstrates that there is more than one pathway to the same end point, but also demonstrates the power of natural selection to arrive at a favorable solution from a variety of starting points (Fish & Beneski, 2014). The organisms associated with any such evolutionary pathway are subject to the same physical laws of nature whether at the molecular, microscopic, or macroscopic level. The laws of motion and thermodynamics, in conjunction with the ecological niche and the available genetic variation within a lineage, constrain phylogenetic trajectories to a particular area of morphospace.
There are myriad ways of negotiating movement on or under land, in air, and through water. Locomotion in each of these environments imposes a specific set of selective pressures that influence the morphological and physiological evolution of animals to be able to operate with a positive energy budget. The majority of the surface of the earth is covered in expansive seas that harbor the majority of animal phyla. The physical properties of the aqueous medium place intense selective pressures on animals that attempt to move in it with speed based upon a high-energy economy. Design space is highly constrained in the aquatic realm.
To understand how and why convergence is driven to a similar endpoint, this chapter focuses on a textbook case associated with morphological and functional similarity in three highly derived types of aquatic vertebrates. Comparison of the shark, ichthyosaur, and dolphin body form represents a prominent example of the expression of convergent evolution (Fig. 15.1; Howell, 1930; Irving, 1966; Hildebrand, 1995; Liem et al., 2001; Thewissen & Nummela, 2008; Wicander & Monroe, 2012; Kardong, 2019). The three groups exhibit similarities in shape, physiology, and mechanics despite their phylogenetic separation (Fig. 15.2). This textbook example demonstrates how similar functional requirements have been met by different clades that have sharply focused on a nearly identical solution to the same environmental challenges. Such convergence, with its resulting analogy and homoplasy, is associated with similar environmental selection pressures imposed on these aquatic predators, enabling them to operate in the open ocean as fast, efficient swimmers that exploit similar trophic opportunities (Gans, 1974; Fish, 1996; Motani, 2000; Bernal et al., 2001a; Donley et al., 2004; Kelley & Motani, 2015; Fischer et al., 2016; Lingham-Soliar, 2016). If aquatic vertebrates are adapted to swim in a manner that minimizes energy expenditure, there should be distinct hydrodynamic advantages to morphological designs and propulsive modes employed by the most derived species in each group. Analysis of swimming mechanics and energetics may thus elucidate physical and biological constraints that may have influenced the convergence of these aquatic animals.

Early illustration from Howell (1930) showing convergence in morphology of a shark, ichthyosaur, and dolphin for a fully aquatic life as fast-swimming predators. Figure from Howell, A. B., Aquatic Mammals: Their Adaptations to Life in Water, 1930. Courtesy of Charles C. Thomas, Publisher, Ltd, Springfield, Illinois

Phylogenetic relationships of the morphologically convergent shark, ichthyosaur, and dolphin
15.2 Physical Characteristics of Water That Affect Swimming Performance
To understand the limitations on locomoting through an aquatic medium that would lead to in convergence, a brief examination of the physical characteristics of water is required. The morphology and locomotor performance of animals in water are dictated by its physical properties (Daniel & Webb, 1987; Webb, 1988). The most pertinent of these physical properties with respect to movement in the aquatic environment, are density and viscosity (Webb, 1975; Daniel & Webb, 1987). Density is the mass per unit volume, whereas viscosity is the resistance to deformation (flow) by the fluid, when there is relative motion between different points in the fluid (Webb, 1975). Density affects inertial and pressure forces within a fluid, whereas viscosity is produced by friction within the fluid due to its “stickiness” resulting from cohesive forces between the water molecules and adhesion between the water and any solid surface (Webb, 1988; Denny, 1993). Pure water is 770 to 890 times denser than air at the same temperature, and at least 50 times more viscous. Thus, the resistance against moving through water is greater than that for air. Density and the pressure that it exerts is directed perpendicular to the surface of a submerged body, whereas viscosity is directed tangential to the surface of a submerged body.
As opposed to a solid, water as a fluid yields when pushed against (Lindsey, 1978). The distortion of the fluid can occur as a swimmer’s body changes the direction and magnitude of the flow, known as vorticity, as it swims through stationary water, as moving water flows around a stationary body, or as the fluid is acted on due to the propulsive movement of the body and/or appendages. The increase in vorticity transfers the kinetic energy of the swimmer’s movement to the water. Energy is lost to eddy formation and frictional forces in the water. As a consequence of the density and viscosity of water, movement through water imposes severe limitations on speed and energetic performance for swimming animals.
Water covers roughly 70% of the Earth’s surface. Despite this enormous two-dimensional area, the oceans, seas, lakes and rivers have a third dimension of depth. As the density of water is related directly to its mass, water is affected by gravity so that the pressure experienced in a water column increases with depth. In seawater, the pressure increases by one atmosphere at sea level (760 mm of mercury, 14.7 pounds per square inch, 101 kPa) with every 10 meters of depth. Many of the tissues and total body composition of marine animals are close to the density of seawater, so they are not compressed with increased depth of submergence (Aleyev, 1977). However, air-filled spaces (e.g., swim bladder, lungs, sinuses, fur, feathers) are compressible (Brawn, 1962; Lovvorn et al., 1999; Fish et al., 2002; Moore et al., 2011). This compression with depth reduces the volume of the air-filled spaces and consequently increases the animal’s density and reduces its hydrostatic positive buoyancy (Kooyman, 1973; Ridgway & Howard, 1979; Moore et al., 2011).
Despite the limitations imposed on aquatic animals by the physical environment, there are advantages to moving in water. Aquatic locomotion can be the most economical form of transport (Tucker, 1970, 1975; Schmidt-Nielsen, 1972). As the density of the body tissues is close to that of water, a swimmer can be near neutrally buoyant, thereby negating the effect of gravity. In addition, both the near neutral buoyancy and viscosity of the water reduces sinking rates when air-filled spaces are compressed. Indeed, even with a loss of positive buoyancy, animals do not “sink like a stone” (Williams et al., 2000; Williams, 2001; Mitani et al., 2010). Unlike terrestrial and flying animals, swimmers do not have to expend energy to support the body against gravity during locomotion (Rayner, 1986; Withers, 1992). The power required for swimming is determined by speed and stress developed in muscles, but the effect of gravity is reduced (Pennycuick, 1992). This has allowed aquatic animals to attain huge sizes, ranging up to the 150-ton blue whale (Balaenoptera musculus) that are far beyond the size attainable by the largest terrestrial and flying animals (McClain et al., 2015; Goldbogen et al., 2019).
The near density match between marine animals and water that reduces the effect of gravity affects the energetics of swimming. Swimming fishes have a lower cost of transport (COT) compared to animals that fly and run (Tucker, 1975). COT is defined as the metabolic energy required to transport a unit mass a unit distance and is calculated by dividing the mass-specific metabolic rate by the swimming velocity (Fish, 1992). COT is inversely proportional to the efficiency of energy expenditure (Tucker, 1970). COT represents the energetic cost by which the metabolic power input is converted to thrust production (Tucker, 1970, 1975; Schmidt-Nielsen, 1972; Williams, 1987; Fish, 1992). The minimum COT is the most efficient and is considered to occur at the velocity which the animal can cover the greatest distance for the smallest energy cost.
Endothermic and homeothermic animals, which can include marine mammals and some species of tuna and lamnid (mackerel) shark, have elevated costs of transport compared to similarly sized ectothermic fishes (Williams, 1999; Fish, 2000; Watanabe et al., 2015). Williams (1999) asserted that the maintenance costs of endothermic tuna and aquatic mammals are higher than those of similarly-sized ectotherms due to the costs of the maintenance of an elevated body temperature and a high basal metabolism. When maintenance costs were omitted, yielding a net cost of transport, the endothermic swimmers were found to have similar locomotor costs, with the minimum COT, to ecothermic fishes. The maintenance costs of marine mammals are 22–77% of the gross COT (Williams, 1999; Fish, 2000). The similarity of locomotor cost of endothermic swimmers and ectothermic fishes, which have the lowest costs of transport among vertebrates, indicates that these endothermic swimmers have reached an optimum in terms of energetic performance (Williams, 1999). Swimming is also relatively economical because propulsive forces are easy to generate in water (Rayner, 1985, 1986).
As endothermy can affect swimming energetics, the thermal conductance and high heat capacity of water is an important physical property that challenges the retention of heat generated by the animal to maintain homeothermy. Water is about 23 times more thermally conductive than air (Denny, 1993). Thus, heat is transferred from an endotherm to the aquatic environment faster than in air, and this is exacerbated for endothermic aquatic animals that maintain an elevated homeothermic body temperature because the thermal gradient is increased. Furthermore, movement through water increases heat transfer by convective exchange. Heat transfer can be reduced with a body geometry that minimizes surface area, where exchange takes place, and the employment of specialized circulatory systems and insulative blubber.
15.3 Convergent Design
15.3.1 Body Streamlining
Design is acknowledged to have a major impact on the ecological performance of organisms (Liem, 1990). The similarity of body design to that of a shark is displayed by ichthyosaurs and cetaceans (Fig. 15.2). Sharks are one of the oldest vertebrate lineages to have become apex predators in the aquatic realm (Sternes & Shimada, 2020). Ichthyosaurs and dolphins have converged upon analogous biological roles to those of highly derived sharks. Ichthyosaurs lived in the oceans for 245 million years, whereas modern dolphins and whales first evolved around 34 million years ago (Motani, 2000; Thewissen et al., 2009). The similarity of the two tetrapods to sharks developed after their terrestrial ancestors returned to the sea (Howell, 1930; Motani, 2000; Thewissen & Bajpai, 2001; Caldwell, 2002; Gingerich, 2015). Such convergence is associated with constraints imposed on these swimmers by the physical environment and the similar biological roles that they fulfill. As apex predators, selection favored morphological, physiological, and behavioral adaptations for swimming performance that maximized speed and efficiency (Daniel & Webb, 1987; Blake, 1991; Fish, 1992; Motani, 2000; Donley et al., 2004; Lingham-Soliar, 2016). The evolution of highly derived convergent morphologies and swimming modes represents the culmination of a sequence of transitional stages displayed by chondrichthyans, reptiles, and mammals (Howell, 1930; Massare, 1988; Gingerich et al., 1990; Fish, 1992; Lingham-Soliar & Reif, 1998; Motani, 2000; Buchholtz, 2001a; Thewissen & Bajpai, 2001; Donley et al., 2004; Sternes & Shimada, 2020).
The most strikingly analogous features of the convergent shark, ichthyosaur, and dolphin are the shapes of their bodies and appendages (Fig. 15.2). Specifically, the highly derived group of sharks that can be considered the template for the convergent design are those in the family Lamnidae. Lamnid sharks include the great white shark (Carcharodon carcharias), mako shark (Isurus oxyrinchus), porbeagle shark (Lamna nasus), and salmon shark (Lamna ditropis). The most derived ichthyosaurs are members of the clade Thunnosauria exemplified by the families Ichthyosauridae (e.g., Ichthyosaurus sp.), Ophthalmosauridae (e.g., Ophthalmosaurus icenicus), Stenopterygiidae (e.g., Stenopterygius sp.), and Temnodontosauridae (e.g., Temnodontosaurus sp.). The family of cetaceans that exemplifies convergence with lamnid sharks and ichthyosaurs is the Delphinidae (oceanic dolphins), which comprises about 32 species, including the ubiquitous bottlenose dolphin (Tursiops truncatus), common dolphin (Delphinus delphis), white-sided dolphins (Lagenorhynchus sp.), spotted dolphins (Stenella sp.), and the notorious killer whale (Orcinus orca). For the remainder of this discussion, unless otherwise specified, the highly derived exemplar species listed above are referred to broadly as sharks, ichthyosaurs, and dolphins.
Lamnid sharks, thunnosaurian ichthyosaurs and delphinids all exhibit similarities in body shape and appendage form that represent adaptations for optimizing swimming performance, maximizing energy efficiency, enhancing swimming speed, and maintaining stability in the open ocean. Overall body shape is the major determinant of resistance (i.e., drag) to movement through a fluid (Fish & Rohr, 1999: Gutarra et al., 2019). The body of the focal groups is streamlined and has smooth contours. Their streamlined bodies exhibit a fusiform shape similar to that of engineered high-performance hydrofoils (Hertel, 1966; Webb, 1975; Vogel, 1994; Fish, 1996, 2018). The fusiform shape resembles an elongate teardrop with a rounded leading edge that extends posteriorly to a maximum thickness and a gradually tapering tail (Fig. 15.3). This shape is accentuated by the anterior position of the bulk of the locomotor muscles (Pabst, 1990, 2000; Bernal et al., 2001a; Shadwick, 2005).

Fusiform body shape of a common dolphin (Delphinus delphis). The white line indicates the position of the maximum thickness, called the shoulder position
Streamlining minimizes the total drag on a body and reduces energy expenditure when swimming (Webb, 1975; Fish & Hui, 1991; Vogel, 1994; Fish et al., 2008; Gutarra et al., 2019). For a fully submerged, streamlined body, the total drag has two components: frictional or viscous drag and pressure or form drag (Webb, 1975; Fish, 1993a, b; Vogel, 1994). The frictional drag is due to the viscosity of the fluid, giving rise to tangential forces resulting from skin friction. Because the water immediately attached to the surface of a body does not move relative to the body (no slip condition; velocity = 0), a thin layer of water (boundary layer) encompasses a velocity gradient from the body surface to the free stream velocity outside of the boundary layer (i.e., outer flow). The velocity gradient generates shear (frictional) forces that consume kinetic energy from the movement of the body and transfers it to the water. The frictional drag is proportional to the wetted surface area of the body. The pressure drag component results from the distortion of the flow outside the boundary layer (i.e., deviation of the trajectory of the streamlines) and is dependent on pressure distribution as the body deflects the water. The pressure acts perpendicularly to the body surface.
The relationship between the pressure in a fluid and its velocity is expressed by the Bernoulli equation (Webb, 1975):
where ρ is the density of the fluid, U is the velocity of the fluid along a streamline (i.e., line tangent to the direction of flow at every point in a flow field; Fox et al., 2009), g is the acceleration due to gravity, h is the depth of the streamline, and P is the reference pressure. The first term is the dynamic pressure of the moving fluid, and the second term is the static pressure of the mass of fluid above the streamline. For streamlines of the same depth, the total pressure within the fluid is determined by the first term. The equation thus indicates that the pressure of the fluid is inversely proportional to the U2, so a high velocity gives a low pressure and vice versa.
The pressure is highest at the rostrum or leading edge, where the flow stagnates. As the flow moves around the rostrum it accelerates and reaches a high velocity at the maximum thickness of the body. By Bernoulli’s principle, the pressure at the rostrum is high because the velocity of the flow is zero, but as the velocity increases downstream, the pressure is reduced. This forebody region, therefore, has a favorable pressure gradient (i.e., water flow from high to low pressure). Downstream of the maximum thickness the flow starts to decelerate and pressure increases, but not to the extent displayed at the rostrum. This region has an adverse pressure gradient (i.e., water must move against an increasing pressure) and the boundary layer flow around the body must have enough energy to continue to remain alongside the body before separating into the wake. Separation of the boundary layer occurring at the trailing edge of the body will produce a narrow wake. If there is insufficient energy and momentum in the boundary layer flow, the increased pressure in the aft of the body will decelerate the flow. This reduction in flow velocity can lead to instabilities and flow reversal. Reversals in the form of eddies and vortices can interact within the boundary layer and cause it to prematurely detach from the body and interact with the outer flow. Such premature separation further increases the kinetic energy losses and associated drag on the body, which is manifested as a broad wake.
The flow within both the boundary layer and outer flow can be laminar, turbulent, or transitional (Webb, 1975). The type of flow is dependent on the Reynolds number (Re), which is the ratio of inertial to viscous forces in the fluid. Re is calculated as the product of the flow velocity, characteristic length, and density of the fluid divided by the viscosity. Typically, the flow will be laminar at Re < 5 × 105, turbulent at Re > 5 × 106, and transitional between these values (Webb, 1975; Fish, 1993a, b; Vogel, 1994). The shark, ichthyosaur, and dolphin swimming at cruising speeds will be in the turbulent regime with Re above 106. In a laminar flow, the virtual streamlines within the flow are parallel and orderly; whereas, in turbulent conditions, the flow is disordered and chaotic. There is increased momentum transfer and shear stresses in a turbulent flow. The ordered structure of laminar flow consumes less energy and has a lower drag than turbulent flow. However, laminar flow in the boundary layer is more susceptible to premature separation, with a concomitant increase in the pressure drag. A turbulent flow will energize and maintain attachment of the boundary layer and minimize the pressure drag at the expense of a slight increase in the frictional drag. Large animals swimming at high speeds, therefore, have a lower total drag with a turbulent boundary layer when compared to laminar conditions.
An indicator of the degree of streamlining is the fineness ratio (FR). FR is the ratio of the body length to the maximum diameter. An FR value of 1.0 would have a circular profile. The FR value of 4.5 is considered to induce the least drag and surface area for the maximum volume (Fig. 15.4; von Mises, 1945; Hertel, 1966; Webb, 1975; Ahlborn et al., 2009), although only a 10% increase in drag is realized in the range of FR of 3 to 7. The value of 4.5 was based on airship designs. Another study that examined the relationship of FR and drag on axisymmetrical torpedoes indicated that the optimal FR was 7 (Gertler, 1950). Depending on the contours of the body, animals should then have minimal drag in the FR range of 4.5 to 7.

Relationship of drag and Fineness ratio (black line) based on von Mises (1945). The dashed line shows the optimal Fineness ratio to be 4.5 for minimal drag based on airship design. The gray box illustrates the limits of Fineness ratio of 3–7 where drag increases by 10% from the optimal value
FR for lamnid sharks, ichthyosaurs, and dolphins overlaps substantially (Fig. 15.5). FR for lamnid sharks is concentrated around the value of 4.5. However, FR for ichthyosaurs and dolphins has a large range spanning values between 4.5 and 7 (Fig. 15.5). Data for ichthyosaurs from Massare (1988) were based on a length that was measured from the anterior edge of the orbit on the skull to the tip of the terminal caudal vertebra. Inclusion of the elongate rostrum in the total length of ichthyosaurs would have skewed the data to higher values of FR. The data for the sharks and dolphins used the fork or notch length, respectively, which was measured as the length from the tip of the rostrum to the notch in the caudal fin or flukes. Despite the differences in measurement, the distribution of FR values for the three convergent groups falls largely within the optimal range (4.5–7), indicating a low drag body form.

Fineness ratio distribution for lamnid sharks, ichthyosaurs, and delphinid dolphins. The solid line shows the optimal Fineness ratio of 4.5 for minimal drag based on airship design (from von Mises, 1945) and the dashed line represents the optimal value for minimal drag based on axisymmetrical torpedo bodies (from Gertler, 1950)
FR is a crude indicator of streamlining of the body because it does not provide information on changes in body contour. Body shapes can be compared to standardized two-dimensional airfoils that are classified by the United States National Advisory Committee for Aeronautics (NACA) (Abbott & von Doenhoff, 1959; Fish & Rohr, 1999). More recent data on engineered foil sections are available through Airfoil Tools (airfoiltools.com). The advantage of such comparisons is that the specific hydrodynamic characteristics have been measured for the foil sections. For example, a dolphin body has been compared to a NACA 66–018 foil (Hertel, 1966) and a killer whale (Orcinus orca) has a streamlined body shape similar to a NACA 66–026 (Pershin, 1983). Both these foil sections are designated as low drag laminar profiles (Fish & Rohr, 1999). However, the choice of foil design to represent body shape is subjective and overly simplistic in not accounting for body contours that do not match the idealized foil shape (Weber et al., 2009a; Fish, 2015). In addition, the animal’s body is three-dimensional with attached appendages that are not accounted for in the two-dimensional foil design.
Another important parameter related to streamlining is the position of maximum thickness or shoulder position (SP) relative to the body length (BL). SP influences the hydrodynamic performance associated with lift and drag for a fusiform body profile (von Mises, 1945; Hoerner, 1965). SP can be associated with the point of transition from laminar to turbulent flow and boundary layer separation (Fish et al., 2008). A rearward displacement of SP allows for the maintenance of laminar flow over a larger portion of the body. However, a placement of SP too far aft on the body will trade off low drag laminar flow for the premature separation of the boundary layer and development of turbulence when not closely oriented to the oncoming flow, such as when maneuvering (Walters, 1962; Lang, 1963; Webb, 1975; Vogel, 1981). Hertel (1966) assigned a generic shark with a FR of 5.56 an SP of 0.44 BL. Kabasakal and Kabasakal (2013) measured an SP of 0.42 BL on a shortfin mako (Isurus oxyrinchus). SP for dolphins is 0.34–0.45 BL.
SP data have not been collected for ichthyosaurs. Massare (1988) modeled the bodies of ichthyosaurs and other Mesozoic aquatic reptiles as streamlined prolate spheroids and measured BL as the distance from the anterior edge of the orbit to the tip of the tail, which would complicate any SP measurement because BL includes the caudal fin and excludes the elongate rostrum. From Fig. 4 (Massare, 1988), based on the ichthyosaur Ophthalmosaurus (after Andrews, 1910), an SP of 0.31 BL is calculated. However, if the rostrum is included into BL, then the SP is 0.26 BL. If BL is measured at the angle in the tail, where the caudal fin may have originated, then SP is 0.31 BL. If the rostrum and caudal fin are excluded, then SP is 0.38 BL. This latter value is within the range of SP found in dolphins and close to the value for lamnid sharks, indicating convergence on a hydrodynamically-optimized shape.
Although the general body is fusiform for lamnid sharks, ichthyosaurs and delphinids, the rostrum is elongate in ichthyosaurs and to a lesser extent in some delphinids. The long, narrow rostrum is probably present to serve as a means of housing a battery of numerous teeth that enable feeding on small prey (e.g., fish, squid), despite potentially conflicting with the fusiform profile. However, the elongated rostrum of ichthyosaurs and some delphinids may act to reduce drag (Aleyev, 1977; Bandyopadhyay, 1989; Videler, 1993). The rostrum can potentially affect the pressure distribution over the forebody. The intersection of the rostrum and more posterior region of the head is marked by a concave region that facilitates transition from laminar to turbulent flow in the boundary layer (Aleyev, 1977). The turbulized boundary layer is less likely to separate and the pressure drag can be minimized over the aft of the body. In addition, the presence of an elongate rostrum reduces the pressure on the forebody, resulting in a reduction of the pressure differential over the body (Bandyopadhyay, 1989; Videler, 1993; Nesteruk, 2020).
15.3.2 Control Surfaces and Fin Shape
Aside from the analogous fusiform body shape exhibited by all three of the convergent taxa, it is the appendages, with their shape, position, and function that truly cement the idea of morphological convergence (Fish, 2004). The appendages are all considered to be highly streamlined (Fig. 15.6; Lang, 1966; Carey et al., 1971; Fish & Rohr, 1999; Motani, 2000). The fins, flippers, and flukes all function similarly as control surfaces. Control surfaces first evolved in aquatic organisms in association with their role in moderating stability and maneuvering (Nursall, 1962; Radinsky, 1987).

Streamlining of body and control surfaces of a dolphin. Images of cross-sections of flipper, dorsal fin, and flukes were obtained from CT scans
Control surfaces are structures that allow for adjustments to, and management of, the attitude of a body in a fluid (Fish & Lauder, 2017). When suspended within the water column, animals can move freely about three orthogonally arranged axes that intersect at the center of mass (CM) (Fig. 15.7). CM is the point where the weight of the animal is considered to be concentrated and acts as the ‘balance point’. Movement about CM permits translation and rotation that give six degrees of freedom. The degrees of freedom of the translational movements are surge (anterior-posterior), heave (vertical displacement), and slip (lateral displacement), whereas rotation about the axes is termed roll for the longitudinal axis, pitch for the lateral axis, and yaw for the vertical axis (Fig. 15.7; Webb, 2004, 2006). Control about the roll axis governs lateral stability, about the yaw axis governs directional stability, and about the pitch axis imparts longitudinal stability. Longitudinal stability (i.e., horizontal orientation) is associated with ‘trim’, which relates to fore and aft balance. Trim is determined by the alignment of the centers of mass and buoyancy in the vertical axis, and the longitudinal axis of the horizontally oriented body (Burcher & Rydill, 1994).

The six degrees of freedom that specify movements of a free body. For the three orthogonal orientation axes running through the center of mass (black circle), there are three translational movements (Slip, Heave, Surge) and corresponding rotational movements (Pitch, Yaw, Roll)
Governance of each degree of freedom by various combinations of control surfaces maintains stability that combats internal and external perturbations (Fish & Lauder, 2017). Perturbations are forces and torques that cause undesired changes in attitude and trajectory (Webb, 2006). Internal perturbations are related to self-generated motions of the animal and changes in body density (e.g., muscle contraction, gas distribution). External perturbations are a function of forces impinging on the animal from the environment (e.g., waves, currents, vortices shed from structures in flow, interactions with other animals). The highly streamlined bodies of the shark, ichthyosaur, and dolphin should be unstable and necessitate the addition of control surfaces for stability (Triantafyllou, 2017; Fish & Lauder, 2017). The position, size, and geometry of the control surfaces help to maintain stability and suppress instabilities by generating forces that counter perturbations (Harris, 1936; Aleyev, 1977; Weihs, 1993; Bandyopadhyay et al., 1997; Fish, 2002, 2004; Fish & Lauder, 2017). Alternatively, when deployed asymmetrically these same stabilizing structures can initiate instabilities to assist maneuvering. It is this duality of function that makes control surfaces so important in the locomotor performance and is expressed in the similar design and position of the various appendages in the three convergent taxa.
Like the rudder and dive planes of a submarine (Burcher & Rydill, 1994; Gabler, 2000), the appendages have evolved into hydrofoils (i.e., wing-like planing surfaces) that hydrodynamically generate lift from the animal’s movement dedicated to the control of stability and maneuverability (Harris, 1936, 1938; Lang, 1966; Fish & Shannahan, 2000; Fish, 2002, 2004; Webb, 2004, 2006; Cooper et al., 2008; Weber et al., 2009b, 2014; Fish & Lauder, 2017). The lift force can be vectored in a particular direction for stabilization or to generate instability for maneuvering. The lift force created by the control surface is a function of the surface area of its planform (i.e., shape) and Aspect Ratio (AR = fin span2/planform area), where span is defined as the linear distance from base to tip of the control surface. Well-performing hydrofoils maximize the lift to drag ratio (L/D) (von Mises, 1945; Webb, 1975; Vogel, 1994; Weber et al., 2009b, 2014). Increased lift can be fostered by cambering (i.e., asymmetry between dorsal and ventral surfaces of a hydrofoil).
The control surfaces are modifications of the paired appendages (including the pectoral and pelvic limbs), the median dorsal fin, and the caudal fin or flukes (Harris, 1936, 1938; Riess, 1986; Fish, 2004; Lingham-Soliar, 2004; Fish & Lauder, 2017). Each of these control surfaces has an elongate teardrop design in cross-section (Lang, 1966; Fish, 2002). As with the fusiform body, the cross-sectional profile maintains low drag and reduces the energy lost by moving through the dense aquatic medium (Lang, 1966; Fish et al., 2007).
Although the caudal fin or flukes have a propulsive function (see below), these caudal extensions can do double duty as control surfaces. The caudal fins of the shark and ichthyosaur can be used as a rudder to stabilize the body in the yaw axis or to generate torques to effect a turn (Harris, 1936; Webb, 1975; Fish & Lauder, 2017); whereas, the horizontal orientation of the dolphin flukes permits control of pitch and the compressed peduncle can be laterally flexed to induce yawing motions (Fish, 2002).
All three of the convergent taxa possess pectoral appendages and caudal and dorsal “fins”. Pelvic fins are present in both the shark and ichthyosaur, but not the dolphin. Another prominent difference is the orientation of the caudal fin. Both the shark and ichthyosaur have a vertically oriented caudal fin that can be displaced laterally, but the caudal flukes of the dolphin are oriented in the horizontal plane and move dorsoventrally. The dolphin’s dorsoventral propulsive movements are a legacy of the change in posture to an up-right stance of mammals from a sprawling stance and an associated reorientation of the axial muscles (Fish, 2001). Another difference is that the shark has additional median fins, with a second, smaller dorsal fin and a ventrally located anal fin.
The paired (pectoral, pelvic) fins and the dorsal and anal fins of the shark are generally thin in cross-section. Flattened cartilaginous basal elements are situated at the base of the fin and numerous smaller radial cartilages extend distally from the basals. The majority of fin area is supported by a fan-like array of collagenous ceratotrichia (Kemp, 1977), which cannot be actively bent. However, it was proposed that an increase in the hydrostatic pressure within the body could be transferred through the cross-helical collagen fibers surrounding the body and into the dorsal fin of the white shark (Carcharodon carcharias) to stiffen the fin (Wainwright et al., 1978; Lingham-Soliar, 2005a). Pectoral and pelvic fin movements are somewhat limited in sharks (Harris, 1936, 1938; Wilga & Lauder, 2000, 2001; Fish & Shannahan, 2000).
The fore and hind limbs of ancestral tetrapods have been adapted for use in water as flippers in the aquatic tetrapods. These limbs enclose a bony skeleton homologous with the bones of terrestrial tetrapods (Williston, 1914; Howell, 1930; Caldwell, 2002; Fish, 2004; Cooper et al., 2007; Kelley & Pyenson, 2015; Massare & Lomax, 2019). Flippers represent modifications of the pectoral and pelvic limbs. There is limited mobility of the elements within these appendages. In particular, the digits are not separated, resulting in a loss of independent movement within the flippers. Flippers are used to control stability and maneuverability. Movement of the flippers is confined to the shoulder and hip joints (Williston, 1914; Howell, 1930; Felts, 1966; Fish, 2004; Cooper et al., 2007, 2008). The number of phalanges in each digit of the flipper is variable among species. Hyperphalangy is the condition found in cetaceans and ichthyosaurs in which the maximum number of phalanges in the digits often greatly exceeds the number in the ancestral state (Williston, 1914; Howell, 1930; Riess, 1986; McGowan, 1991; Fish & Battle, 1995; Caldwell, 2002; Cooper et al., 2007; Maxwell, 2012; Massare & Lomax, 2019). Polydactyly (i.e., extra digits) also occurs in some aquatic tetrapods (Fedak & Hall, 2004).
The effectiveness of the various control surfaces is dependent on their shape and AR. High AR indicates a long narrow control surface associated with high lift generation, whereas low AR indicates a broad surface area with a short span. High AR hydrofoils are characteristic of relatively fast swimmers and have a high lift to drag ratio (L/D). Highly effective fins maximize L/D (Webb, 1975; Weihs, 1989). The AR for the pectoral fins of lamnid sharks is about 3.0 (Hoffman et al., 2020). Dolphins, such as Delphinus, Lagenorhynchus, Stenella, and Tursiops, have pectoral flippers with a pointed tip and a range of AR of 3.5–6.3 (Fig. 15.8a), whereas Orcinus, with rounded flippers, has a lower AR of 1.9–2.9 (Fig. 15.8b; Fish unpubl. Data; Fish et al., 1988). Ichthyosaur specimens of Ichthyosaurus and Stenopterygius had pectoral flippers with AR of 4.5–6.0, within the range of dolphins with flippers of a similar planform (Fig. 15.8c).

Foreflipper design of tetrapods. (a) Pectoral flipper of the striped dolphin, Stenella, (b) pectoral flipper of the killer whale, Orcinus, and (c) pectoral flipper of the ichthyosaur, Stenopterygius (courtesy of Judy Massare)
It is curious that, unlike the shark and ichthyosaur, dolphins lack pelvic or hind flippers. Indeed, all modern cetacean and sirenian (manatee and dugong) species lack external hind flippers despite their quadrupedal ancestors having had hind legs (Gingerich et al., 1990, 1994, 2001, 2015; Thewissen & Fish, 1997; Domning, 2000, 2001; Bejder & Hall, 2002; Thewissen et al., 2007, 2009; Uhen, 2010; Díaz-Berenguer et al., 2018, 2019; Lambert et al., 2019). Internally the pelvis and hind limbs of cetaceans have become vestigial structures (Adam, 2009). The loss of the external hind limbs in modern species is associated with the absence of the genetically controlled signaling cascade for formation of the hind limb (Thewissen et al., 2006; Thewissen, 2018). However, hind limb buds do develop for a brief period before reabsorption in embryonic cetaceans, and occasionally atavistic rudimentary hind limbs appear in modern whales and dolphins (Struthers, 1893; Andrews, 1921; Ohsumi, 1965; Hall, 1984; Bejder & Hall, 2002; Thewissen et al., 2006; Thewissen, 2018). This atavism indicates that the genes controlling hind limb formation have not been entirely lost. The loss of the hind limbs, along with other morphological changes, in the transition from terrestrial to fully aquatic habits, has been heralded as one of the best characterized examples of macroevolution (Thewissen & Bajpai, 2001; Gingerich, 2015; Huelsmann et al., 2019).
Why were the hind limbs lost in cetaceans and not sharks and ichthyosaurs? As control surfaces, the possession of hind flippers in cetaceans would be redundant. A submarine has control surfaces with dive planes anterior to the center of mass either near the bow or on the conning tower and posteriorly in combination with the rudder. Submarines adjust their trim hydrostatically through buoyancy control with the ballast tanks or hydrodynamically with lift generated by the dive planes (Burcher & Rydill, 1994; Fish & Lauder, 2017). The fore and aft position of the two sets of dive planes balances the vessel to keep it in trim. The shark and ichthyosaur both move their caudal fins laterally for propulsion; therefore, to maintain trim, rely upon their pelvic fins/flippers, located posteriorly, working in concert with the anterior pectoral fins (Harris, 1936, 1938; Standen, 2008). However, cetaceans and sirenians have horizontally oriented flukes that act as a control surface at the posterior end of the animal in concert with the anterior pectoral flippers to maintain trim, while the flukes also are used for propulsion (Fish, 2002; Kojeszewski & Fish, 2007). Like the feathers of an arrow, the flukes located far posterior to the CM can generate large directionally-correcting torques because of their long lever arms (Harris, 1936; Wegner, 1991; Webb et al., 1996; Fish, 2002, 2004; Fish & Lauder, 2017). Thus, the posteriorly located flukes, in concert with the anterior pectoral flippers, can perform trim control, so that extra sets of horizontally-oriented control surface such as pelvic flippers are redundant.
In addition to the caudal flukes performing the task of trim control, loss of the pelvic flippers would be of benefit in reducing drag on cetaceans (Bejder & Hall, 2002). Possession of hind flippers with their additional surface area and concomitant increased drag would effectively limit swimming speed and increase energetic costs for locomotion. Possession of hind flippers would, therefore, be hydrodynamically disadvantageous. The lack of pelvic limbs in cetaceans is related to the orientation of the propulsive oscillatory movement of the spine. Dorsoventral movement of the spine is a legacy of the ancestral terrestrial mammals. These mammals stood on erect limbs and reorganized the axial muscles for dorsoventral bending of the spine from the reptilian lateral bending condition (Howell, 1930; Fish, 2001). The ichthyosaurs’ reptilian ancestors retained the ancestral muscle architecture associated with the sprawling posture and employed lateral bending (undulation). Primitive ichthyosaurs had a more anguilliform swimming mode that was fostered by lateral undulations (Motani et al., 1996; Buchholtz, 2001a). This committed the ichthyosaurs to lateral oscillations of the tail, as performed by sharks. Because the tail beat laterally, there was need for posteriorly located pelvic fin control surfaces for trim control. Buchholtz (2001a), however, contended that Stenopterygius quadriscissus and Ophthalmosaurus icenicus, with small hind limbs, were in the process of reduction or loss of these trim stabilizers.
Irrespective of the paired appendages with their skeletal braces, other control surfaces lack internal skeletal support, such as the dorsal fin, flukes, and peduncle keels. In the shark, ichthyosaur, and dolphin, these features are reinforced by dense arrays of collagen fibers (Felts, 1966; Lingham-Soliar, 2001; Lingham-Soliar & Plodowski, 2007). The keels of the peduncle of dolphins are composed of blubber that contains structural collagenous and elastin fibers (Hamilton et al., 2004), which act as tensile stays. Collagen fibers with high tensile strength are found in the flukes of cetaceans (Sun et al., 2010a, b; Gough et al., 2018), and are arranged in a dense array of chordwise-oriented thin crossing fibers composing the inner core layer sandwiched between two layers of spanwise-oriented thick fibers composing the outer ligamentous layers (Felts, 1966; Sun et al., 2010a, b, 2011; Gough et al., 2018). This sandwich composite beam provides rigidity while allowing some bending (Sun et al., 2010b).
The presence of a relatively stiff, non-mobile triangular dorsal fin is a strong indicator of convergence upon a similar lifestyle of the three taxa (Lingham-Soliar & Plodowski, 2007). AR for the dorsal fins is generally less than 2. In all cases, the dorsal fin has an anterior insertion that is near the maximum diameter of the animals and coincides with the longitudinal position of CM (Fig. 15.7; Fish, 2002). The dorsal fin functions to resist roll and yaw (Fish, 2002; Lingham-Soliar, 2005a; Lingham-Soliar & Plodowski, 2007). Its position near CM allows the dorsal fin to perform as an anti-slip device to maintain the trajectory of rapid, small radius turns (Fish, 2002). The wing-like profile of the dorsal fin could also aid in generating lift that is oriented toward the center of rotation to supply the centripetal force for maneuvering.
Based on Fig. 1 of Lingham-Soliar (2016), comparison of the profiles of a great white shark (Carcharodon carcharias) and ichthyosaur (Stenopterygius quadriscissus SMF 457; Senckenberg Museum, Germany) permits the sweep of the dorsal fin to be measured. The sweep is measured as the angle between the line perpendicular to the longitudinal axis of the body and the one-quarter chord position on the fin (Fish & Rohr, 1999). The sweep was 30.0° and 31.3° for the ichthyosaur and shark, respectively. These values are within the range of sweep angles (29.7–57.5°) of the dorsal fins of dolphins (Fish unpubl. data).
The caudal fins/flukes of the three convergent taxa, the shark, ichthyosaur and dolphin, are extensions from the tail. The caudal fins/flukes are similar in their general planform shape (Fig. 15.9). These caudal extremities are relatively stiff, with a high AR lunate planform shape with tapering tips (Williston, 1914; Lighthill, 1969, 1970; Riess, 1986; Webb, 1975; Fish et al., 1988; Massare, 1988; Fish, 1998a, b; Lingham-Soliar, 2005b, 2016; Lingham-Soliar & Plodowski, 2007). The cross-sectional profiles of these caudal fins/flukes have a streamlined fusiform shape similar to that of engineered hydrofoils, although the fossil remains of ichthyosaurs do not allow for an exact determination of the three-dimensional geometry of the caudal fin (Lang, 1966; Fish et al., 2007; Crofts et al., 2019).

Images of a shark (top), ichthyosaur (middle), and dolphin (bottom) showing the high aspect ratio, wing-like caudal fin and narrow peduncle. Shark image courtesy of Rodney Fox
The caudal fins and flukes are used primarily for propulsion (Fig. 15.9). They act as an oscillating wing or hydrofoil to generate a lift-based thrust (see below). The caudal fins and flukes are connected to the body by a narrow peduncle that can be flexed in the direction of oscillation. This ‘narrow necking’, along with keels oriented in the plane of oscillation, reduces the resistance of the peduncle to reciprocating propulsive oscillations (Lighthill, 1969, 1970; Zhang et al., 2020). In addition, the narrow peduncle, in concert with the large inertial mass in the anterior region of the body of the swimmer, will minimize kinetic energy losses due to recoil in the anterior body and reduce drag (Lighthill, 1969; Webb, 1975; Sfakiotakis et al., 1999; Ben-Zvi & Shadwick, 2013).
The caudal fins and flukes largely differ in their composition. The vertebral column extends into the dorsal lobe of the caudal fin of sharks, into the ventral lobe of the caudal fin of ichthyosaurs, and between the two caudal flukes of dolphins (Howell, 1930; Fish, 1998b; Fish et al., 2006; Crofts et al., 2019). For the shark, the blades of the caudal fin are supported by collagenous ceratotrichia (Kemp, 1977; Crofts et al., 2019). The caudal fin and flukes of the ichthyosaur and dolphin, respectively, are supported by compact arrays of collagen fibers (Felts, 1966; Lingham-Soliar, 2005b, 2016; Lingham-Soliar & Plodowski, 2007; Gough et al., 2018).
The flukes of dolphins are symmetrical and have a range of AR of 3.4–5.5. The sweep on the caudal flukes ranges from 25.0° to 41.3° (Fish unpubl. data; Fish et al., 1988). Based on the representative species mentioned above (Lingham-Soliar, 2016), AR for the caudal fins is 3.8 for the shark and 4.2 for the ichthyosaur. The dorsal (hypercaudal) lobe of the caudal fin and ventral (hypocaudal) lobe are asymmetrical for the shark and ichthyosaur. Comparatively, the planar area of the hypercaudal lobe is 34% greater than the hypocaudal lobe of the shark, whereas the planar area of the hypocaudal lobe is only 5% greater than the hypercaudal lobe for the ichthyosaur. The increased size of the respective lobes appears to be associated with the bending direction of the vertebral column.
The asymmetries of the caudal fins of the shark and ichthyosaur will affect the pitching moments of the animals. The heterocercal tail of the shark will induce an upward pitch at the tail and a corresponding downward pitch of the head (Harris, 1936; Alexander, 1965). The pectoral fins situated anterior of CM would then be used to generate a restoring lift force to maintain trim of the body (Harris, 1936; Fish & Shannahan, 2000). The caudal fin of the ichthyosaur is a reversed heterocercal or hypocercal tail, which would induce an opposite rotation of the body compared to that of the shark. Taylor (1987) considered that because of a near neutral buoyancy of the ichthyosaur, the hypocercal fin would induce an upward oriented thrust vector through the center of balance and negate any pitching, as indicated by the model by Thomson and Simanek (1977). However, Wilga and Lauder (2004), and Flammang et al. (2011) used digital particle image velocimetry of a swimming shark and found a strong posteroventrally directed jet flow from the motion of the heterocercal tail. This direct evidence validated the assertions of Harris (1936) and Alexander (1965), while falsifying the hypothesis of Thomson and Simanek (1977). This result would indicate an upward pitch of the heterocercal tail of the shark and conversely a downward pitch of the hypocercal tail for the ichthyosaur. As an obligate air-breather, downward pitch of the tail and upward pitching of the head would be advantageous for the ichthyosaur to facilitate surfacing for breathing, in contrast to the gill-breathing shark (Taylor, 1987; McGowan, 1992; Crofts et al., 2019).
15.3.3 Integument
The integument, or skin, is the one part of the body that is intimate contact with the environment and can function to enhance locomotion (Garten & Fish, 2020). The magnitude of the frictional drag component is a function of the texture of the skin. The interaction of the surface of the body and appendages with the flow of water determines the development of the shear forces in the boundary layer.
Sharks have a rough skin, which appears counter-intuitive to the maintenance of an orderly, low-drag flow. Fast swimming sharks have scales that have flat crowns and sharp ridges oriented longitudinally, with rounded valleys between them (Pershin et al., 1976; Reif, 1978, 1985; Reif & Dinkelacker, 1982; Lang et al., 2008; Oeffner & Lauder, 2012). This scale morphology can potentially provide a 7–8% drag reduction by acting as riblets (Reidy, 1987; Walsh, 1990). Riblets are streamwise microgrooves that reduce drag by acting as small fences to break up spanwise vortices and reduce the surface shear stress and associated loss of momentum.
Small ridges, like riblets, on the epidermis of dolphins have been hypothesized to stabilize longitudinal vortices (Yurchenko & Babenko, 1980), but the geometry of the ridges, with rounded edges, does not suggest an effective analogy with riblets (Fish & Hui, 1991). The skin of cetaceans is generally described as being smooth (Shoemaker & Ridgway, 1991). Wainwright et al. (2019) validated that the skin is smooth, particularly when compared to other pelagic swimmers. It was shown that the size of the ridges on the body of cetaceans is small, or even absent, on skin of the control surfaces of most species.
Ridges were found on the fossilized skin of an ichthyosaur, although these ridges were interpreted to be due to post-mortem wrinkling (Delair, 1966; Lingham-Soliar, 1999). The skin exhibited orthogonally-oriented fibers arranged in dense pre-stressed layers (Lingham-Soliar, 1999). This arrangement was inferred to be for the prevention of wrinkling of the skin and bulging of the muscles in the living animal. Fibrous skin would maintain a smooth hydrodynamic surface (Lingham-Soliar, 1999, 2001). A layer of subdermal collagen fibers occurs in the body and tail of sharks, ichthyosaurs, and dolphins, which could be used for the transfer of muscle forces for swimming (Wainwright et al., 1978; Lingham-Soliar, 1999; Lingham-Soliar & Wesley-Smith, 2008; Flammang, 2010; Crofts et al., 2019).
15.4 Swimming Performance
15.4.1 Swimming Speed
The three convergent taxa are all marine predators and are considered to be capable of rapid and sustained swimming (Massare, 1988; Fish & Rohr, 1999; Buchholtz, 2001b). Lamnid sharks and oceanic dolphins undergo seasonal migrations maintaining a steady swimming speed over days (Miyazaki et al., 1974; Bonfil et al., 2005; Weng et al., 2007, 2008; Taylor et al., 2016). Satellite tag data for white sharks (Carcharodon carcharias) and salmon sharks (Lamna ditropis) indicate migratory speeds of 0.8 to 1.5 m/s (Weng et al., 2007, 2008; Bonfil et al., 2010). Dolphins routinely swim for prolonged periods at speeds of 0.4 to 3.7 m/s (Fish & Rohr, 1999). Based on the metabolically measured cost of transport, Williams et al. (1993) and Yazdi et al. (1999) each estimated that the optimal swimming speed for bottlenose dolphins (Tursiops truncatus) was 2.1 and 2.5 m/s, respectively. The optimal speed would coincide with the minimum cost of transport, which would have the highest efficiency.
Estimates of routine swimming speeds for ichthyosaurs were calculated by Massare (1988) and Motani (2002a, b). Both investigators used models based on metabolically and hydrodynamically derived energetics. Massare (1988) calculated speeds for ichthyosaurs ranging from 1.82 to 3.06 m/s. Initially, Motani (2002a) used a set of hydrodynamic equations to calculate the optimal swimming speed (the speed at which the energy consumption required to move a unit length is minimal) for Stenopterygius. Optimal swimming speed would correlate with the minimum cost of transport. The estimated optimal speeds ranged from 1.2 to 1.7 m/s. Motani (2002b) modified his model and calculated optimal swimming speeds based on the basal metabolic rates for each of three conditions (reptiles, tuna-leatherback turtle, and cetaceans-pinnipeds). His results produced speed ranges of 0.55 to 0.78 m/s (reptiles), 1.2 to 1.6 m/s (tuna-leatherback turtle), and 2.2 to 2.5 m/s (cetaceans-pinnipeds). Motani (2002b) considered the reptilian condition was probably not feasible, and he argued ichthyosaurs would have had a higher metabolism and an elevated body temperature due to their large body size. A cruising speed of at least 1 m/s would have been similar to the swimming performance of the blue marlin (Makaira nigricans) and yellowfin tuna (Thunnus albacares) that have a similar diet to Stenopterygius (Motani, 2002b).
While optimal swimming speed can be estimated to provide an indication of long-duration routine swimming speeds for activities such as cruising, searching, and migrating, burst swimming speeds cannot be predicted. Burst swimming represents an unsteady behavior of short-duration. High-speed swimming is most commonly associated with pursuit of prey and escape from predators (Webb, 1975). It is unknown what the maximum burst swimming speed was for ichthyosaurs, although given the similar morphology and swimming mode (see below), it can be assumed to be comparable to that attained by lamnid sharks and delphinid dolphins. Shortfin mako shark (Isurus oxyrinchus) is capable of a maximum speed of 19.44 m/s (Díez et al., 2015). Burst speeds for dolphins have been reported to range from 5.6 m/s up to 15.0 m/s (Fish & Rohr, 1999).
15.4.2 Swimming Mode
The shark, ichthyosaur, and dolphin share a thunniform (i.e., from Thunnus, the generic name for several species of tuna) bauplan with a stiff, streamlined fusiform body, lunate tail, dorsal fin, and extremely narrow necking of the peduncle (Lighthill, 1969; Webb, 1975; Lindsey, 1978; Motani, 2005; Shadwick, 2005; Lingham-Soliar, 2016; Gutarra et al., 2019). Aside from these morphological similarities, the designation of being thunniform (tuna-like) swimmers indicates similar kinematic patterns for highly efficient aquatic propulsion (Aleyev, 1977; Fish et al., 1988; Lindsey, 1978; Webb, 1975; Motani, 2005). Although there is no direct evidence for the swimming kinematics of ichthyosaurs, the morphological similarities with lamnid sharks and dolphins strongly indicate that these modern analogues can be used as a proxy for the swimming mode of the extinct taxon, particularly for highly derived ichthyosaurs (e.g., Ichthyosaurus, Ophthalmosaurus, Stenopterygius).
Thunniform swimming of the extant convergent species, as well as tuna, encompasses undulatory motions of one-half to one full wavelength within the body. Transmission of the undulations to the caudal-most portion of the body produces an oscillatory motion (Fierstine & Walters, 1968; Lighthill, 1969; Lindsey, 1978; Smits, 2019). Significant propulsive movements are confined to the peduncle and caudal fin (Webb, 1975; Lindsey, 1978; Fish, 1998a). However, the peak-to-peak amplitude (A) at the caudal fin is typically about 20% of body length during routine swimming (Webb, 1975), but can be greater than 30% of body length, particularly during rapid accelerations (Fierstine & Walters, 1968; Skrovan et al., 1999; Fish et al., 2014). The peak-to-peak amplitude of the caudal fin remains relatively constant over a range of swimming speeds. This constancy of the oscillatory amplitude at 20% of body length is tied to minimization of the energy required to swim (Saadat et al., 2017).
As the stroke amplitude remains constant, swimming speed (U) increases directly with the frequency of oscillations (f) of the tail (Fish, 1996; Rohr & Fish, 2004; Smits, 2019). Maximum propulsive efficiency is related to the non-dimensional Strouhal number, which combines the three kinematic parameters in the equation:
The Strouhal number is a gauge of the effectiveness of flapping locomotion by indicating the distance a swimmer moves with each tail stroke (Saadat et al., 2017). The Strouhal number predicts that the maximum spatial amplification and optimal creation of thrust-producing jet vortices lies within a narrow range of 0.2–0.4 (Triantafyllou et al., 1993; Triantafyllou & Triantafyllou, 1995; Streitlien & Triantafyllou, 1998). The Strouhal numbers for thunniform swimmers occur within the optimal range and coincide with the maximal propulsive efficiency between 0.76 and 0.98 (Webb, 1975; Triantafyllou & Triantafyllou, 1995; Fish, 1998a; Rohr & Fish, 2004; Fish & Lauder, 2006).
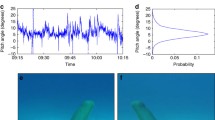
When swimming, the tip of the caudal fin traces out a sinusoidal trajectory for thunniform swimmers (Fig. 15.10). The thunniform tails of the shark and ichthyosaur are heaved (flexed) laterally at the peduncle (Shadwick, 2005), whereas the dolphin flukes are dorsoventrally heaved (Fish & Rohr, 1999). In addition, the base of the caudal fin is pitched so that the angle of inclination (i.e., angle of fin relative to longitudinal movement of swimmer) is maintained at a defined angle throughout the stroke cycle. Without this pitching motion, the fin would sweep through the stroke with a continuously changing angle of inclination. The combination of heave and pitch causes the trailing edge of the caudal fin to lag behind its leading edge, allowing the fin to slice through the water (Van Buren et al., 2020). The addition of heave to a pitching motion can dramatically increase the thrust and propulsive efficiency of an oscillating wing-like hydrofoil. Flexion at the peduncle and base of the caudal fin emulates a double-jointed system, which allows the angle of inclination (α*; Fig. 15.10) of the caudal fin to be adjusted throughout the stroke cycle, maintaining nearly continuous maximum thrust (Parry, 1949; Fierstine & Walters, 1968; Lindsey, 1978; Reif & Weishampel, 1986; Fish et al., 1988).

Path of oscillating dolphin flukes through a stroke cycle. The tips of the flukes move along a sinusoidal path. Sequential fluke positions along the path are illustrated as straight lines. The box on the left shows the relationship between the tangent to the path of the flukes with the angle of attack, α, and the incident angle, α*. Angle of attack is the angle between the tangent of the fluke’s path and the axis of the fluke’s chord; pitch angle is the angle between the fluke axis and the translational movement of the animal. The box on the right shows the relationship between the major forces produced by the motion of the fluke. D is the drag, L is the lift, and T is the thrust resolved from L (from Fish, 1993b)
The thunniform namesake tuna possesses the double-jointed system, with joints at the bases of the anterior end of the peduncle and caudal fin. These two flexion points are separated by a few stiff, laterally keeled vertebrae, which act as pulleys for the tendons connecting with the large anterior muscle mass (Fierstine & Walters, 1968). Despite having keels on the peduncle (Lingham-Soliar & Reif, 1998; Bernal et al., 2001a; Hamilton et al., 2004), the lack of keels on the vertebrae and numbers of vertebrae in the peduncle of the three convergent taxa do not preclude an analogous double-jointed system with the tuna. Instead, the larger number of vertebrae with anteroposteriorly short centra in the peduncle permits increased flexibility for heaving motions (McGowan, 1992; Buchholtz, 2001a, 2001b, 2007; Lingham-Soliar, 2001; Buchholtz & Schur, 2004; Buchholtz et al., 2005; Kim et al., 2013). Flexural pitching of the caudal fin can be actuated by the insertions of long tendons from the anterior musculature (Fierstine & Walters, 1968; Pabst, 1990, 1996; Lingham-Soliar & Reif, 1998; Lingham-Soliar, 2001; Shadwick & Gemballa, 2005; Adams & Fish, 2019). Furthermore to prevent “bowstringing” of the tendons (i.e., lifting away from the joint), fibrous connective tissue in the skin would act like a retinaculum (Fierstine & Walters, 1968; Lingham-Soliar & Reif, 1998; Pabst, 2000). Maximum flexion occurs near the base of the flukes of dolphins at a vertebra known as the “ball vertebra” (Watson & Fordyce, 1993; Tsai, 1998; Fish et al., 2006). This vertebra has convex (rounded) anterior and posterior faces and differs from the flat (acoelous) faces of the other vertebrae. The ability to rotate the flukes about a pitching axis at the ball vertebra allows for control of the angle of the flukes when swimming (Long Jr. et al., 1997; Fish, 1998b). The abrupt dorsal or ventral tailbend of the vertebral column of the shark and ichthyosaur, respectively, indicate this position to be the flexion point for the pitch of the caudal fin.
Aquatic propulsion employing the thunniform mode results from the transfer of momentum from the animal to the water (Webb, 1988). The rate of momentum exchange between the propulsor and the water determines the amount of thrust generated (Daniel et al., 1992). Thrust is produced exclusively by the stiff, high-AR, lunate tail, which acts like an oscillating hydrofoil or wing (Williston, 1914; Lighthill, 1969; Riess, 1986; Webb, 1975; Fish et al., 1988; Massare, 1988; Fish & Hui, 1991; McGowan, 1992; Fish, 1998a, b; Lingham-Soliar, 2004; Shadwick, 2005). The momentum imparted to the water by the oscillating hydrofoil takes the form of a wake with two alternating rows of thrust-type vortices and a posteriorly directed jet stream (Weihs, 1972; Triantafyllou et al., 2000; Fish et al., 2014, 2018; Smits, 2019).
Thrust and efficiency are maximized by the use of lift (circulation)-based oscillating hydrofoils (Lighthill, 1969; Webb, 1975; Sfakiotakis et al., 1999; Fish & Lauder, 2006). Lift-based oscillatory swimming is associated with the radiation into pelagic habitats where steady swimming is required (Webb & de Buffrénil, 1990). Lift is generated as for an airplane wing, and results from differential flow between the surfaces of the hydrofoil (i.e., Bernoulli effect) because the foil is canted at an angle of attack (α; i.e., incident angle to on-coming flow). The heaving and pitching motions of the caudal fins of the thunniform swimmers are responsible for changes of angle of inclination and angle of attack throughout the stroke cycle (Fig. 15.10). As the caudal fin moves along a sinusoidal path through the water, the incident flow encountered by the fin is a combination of the forward movement of the swimmer and the heaving motion of the tail, where the pitch of the fin relative to its path is the angle of attack (Fig. 15.10). Lift is directed perpendicular to the pathway traversed by the caudal fin and can be resolved into an anteriorly directed thrust vector (Lighthill, 1969; Weihs & Webb, 1983; Fish, 1993b). Thrust is derived from a combination of the horizontal component of the lift force and leading-edge suction (Ahmadi & Widnall, 1986). Thrust derived from lift increases directly with increases in angle of attack. To maximize lift, the propulsor is maintained at an angle of attack (<30°) throughout the stroke cycle (Fish et al., 1988; Fish, 1993b). However, low angles of attack increase efficiency while reducing the probability of stalling (i.e., dramatic loss of lift) and decreased thrust production (Chopra, 1976; Van Buren et al., 2020). By restricting bending to the peduncle and base of the caudal fin, this permits rotational motion to maintain a positive angle of attack of the caudal fin to the oncoming flow (Webb, 1975). Thrust is thus generated continuously throughout a stroke cycle.
Some drag is produced by the oscillating fin as a result of skin friction and drag due to lift (i.e., induced drag). The induced drag is small compared to the lift. A high L/D ratio is a function of the high AR of the caudal fin (Bose & Lien, 1989; Liu & Bose, 1993; Fish, 1998a, b). Induced drag is also limited by sweepback of the caudal fin. Sweepback is the angle made by the leading edge with respect to the longitudinal axis of the body. Minimal induced drag is fostered by a swept wing planform with a triangular shape (Küchermann, 1953; Ashenberg & Weihs, 1984).
A tapered wing with sweepback or crescent design can reduce the induced drag by 8.8% compared with a wing with an elliptical planform (van Dam, 1987). The combination of low sweep with high AR allows for high efficiency rapid swimming (Azuma, 1983). Sweep angles of 30° and 40° produce more thrust than a 50° swept fin during the mid-stroke for the caudal fin, but as the fin reverses direction during the oscillation, the 50° sweep produces more thrust (Matta et al., 2019). However, a fin exceeding about 30° of sweep leads to a reduction in efficiency and large sweeps generate less lift (Chopra & Kambe, 1977; Matta et al., 2019). The sweep angle of the symmetrical caudal flukes of delphinids ranges from 4.4° to 41.3° and AR ranges from 3.0 to 5.5 (Fish, unpubl. data). Based on Fig. 1 from Lingham-Soliar (2016), the sweep angle for the hypercaudal and hypocaudal lobes of the caudal fin with AR of 3.8 of a great white shark (Carcharodon carcharias) is 37.8° and 51.0°, respectively. The sweep angle for the hypercaudal and hypocaudal lobes of the caudal fin with AR of 4.2 of the ichthyosaur (Stenopterygius quadriscissus SMF 457) is 33.0° and 50.9°, respectively.
The relationship between sweep and AR also indicates a structural limitation to the strength and stiffness of the propulsor (van Dam, 1987; Bose et al., 1990). The ability to sustain certain loads without breaking is considered a major constraint on increasing span and AR (Daniel, 1988).
15.5 Porpoising
Highly derived aquatic animals should avoid swimming close to the surface of the water. At the surface, kinetic energy from the motion of the animal is transferred as potential energy to the vertical displacement of water in the formation of waves (Denny, 1993; Vogel, 1994). This transfer increases the resistance on the swimming animals as wave drag. The additional drag on the body can be as much as five times the frictional drag on a fully submerged body (Hertel, 1966). The maximum increased drag occurs when the longitudinal axis of the body is at a depth of half a body diameter. Movement at or near the surface can thus substantially increase the energy expended and limit the speed of swimming (Hertel, 1966; Fish, 1996). For example, sea otters swimming submerged have an oxygen consumption 41% lower than when surface swimming and a maximum swimming speed when submerged that is 74% faster than at the surface (Williams, 1989). When towed at the surface, the drag on harbor seals (Phoca vitulina) was 2.5 times greater than when submerged at the identical velocity (Williams & Kooyman, 1985). Elimination of wave drag occurs at a submergence depth of ≥3 times the body diameter (Hertel, 1966), although wave drag was essentially zero below a depth of only two body diameters for a dolphin (Lagenorhynchus) at a high swimming speed (6.1–9.1 m/s) (Lang & Daybell, 1963). Thus, the locomotor strategy of submerged swimming can result in increased efficiency by the removal of wave drag.
For the shark, gas exchange via the gills means that it can remain submerged indefinitely and thus avoid wave drag, but the ichthyosaur and dolphin are obligate air-breathers and must return to the surface to ventilate the lungs. These aquatic tetrapods can utilize a strategy of swimming below the water surface for prolonged periods. The dolphin, like other marine mammals (cetaceans, pinnipeds), is adapted to extend periods of submergence because of its enhanced oxygen storage capacity resulting from increased blood volume and elevated levels of myoglobin in the muscles (Ridgway & Johnston, 1966; Kooyman, 1989; Elsner & Meiselman, 1995; Noren et al., 2002). Furthermore, the dolphin can use its oxygen reserves sparingly by reducing its metabolism utilizing diving bradycardia, peripheral vasoconstriction, and, if necessary, shifting to an anaerobic metabolism (Scholander, 1940; Kooyman, 1989; Williams et al., 1991, 2015). To prevent increased energy cost when coming to the surface to breathe, these animals limit such times and quickly ventilate the lungs before submerging. Dolphins can ventilate 90% of their lung volume in about 0.3 s (Irving et al., 1941; Ridgway et al., 1969; Kooyman & Cornell, 1981; Hui, 1989).
A variation on the strategy to limit increased drag at the water surface and prolong ventilation time is accomplished by porpoising (Hui, 1987, 1989; Williams, 1987; Fish & Hui, 1991). Porpoising consists of repetitive high-speed motions of rhythmic, long, ballistic jumps alternating with periods of submerged swimming close to the surface (Au et al., 1988; Fish & Hui, 1991; Weihs, 2002). Porpoising is conducted by fast swimmers, including dolphins, penguins, and pinnipeds. The energy required for leaping at low swimming speeds is greater than the energy expended in swimming just below the surface of the water. Porpoising becomes energetically efficient when the energy required to swim a given distance just below the water surface increases with swimming speed faster than the energy to leap that distance (Au & Weihs, 1980; Au et al., 1988; Blake, 1983). Above a critical speed, known as the crossover speed, where the energies of submerged swimming and leaping converge, there is an energetic advantage to swimming by porpoising. For dolphins, various models have indicated a crossover speed of at least 3.9 m/s for porpoising (Fish & Rohr, 1999).
Although energy could be conserved by porpoising, data on porpoising animals contradict the assertions of the models. The models have assumed an emergence angle of 45° to maximize the distance traveled through the air. Gordon (1980) indicated that to maintain forward speed an emergence angle of 30° would be desirable as a compromise between maximum distance and maximum forward speed of a leap. In reality, the emergence angle of the leaps is approximately 39°, with wide variation (Hui, 1989; Weihs, 2002), which lies between the optimal predicted angles. Also bringing into question the energy savings of porpoising are data indicating that the distances covered in the air are interspersed with swimming bouts of about twice the length of the leap. The assumptions of energy savings by porpoising predict that dolphins would spend more time leaping than swimming at speeds greater than crossover speed (Au et al., 1988).
Video data of free-ranging dolphins indicate a graded transition from minimal blowhole exposure at the surface at low swimming speeds to quasi-leaps, in which the dolphin is never completely out of the water at any instant, at medium swimming speeds, and complete porpoising leaps at the highest swimming speeds (Hui, 1989). These observations are consistent with maintaining a minimum blowhole exposure time for respiratory inhalation as swimming speed increases. Consequently, porpoising behavior may be energy conserving only in the sense that it is energetically the cheapest way to breathe, not energetically the cheapest way to swim (Fish & Hui, 1991).
The position of the external nares of ichthyosaurs might have made porpoising a more effective means of breathing while swimming. Unlike dolphins, ichthyosaurs had external nares situated at the end of the rostrum, anterior to the eyes, necessitating that the rostrum be lifted above the water surface to breathe. Although there is no direct evidence for porpoising, ichthyosaurs were envisioned to swim at high enough speeds to enable leaping out of the water and porpoising due to the convergence of design and swimming mode with dolphins. The artist Heinrich Harder (1858–1935) produced a rendering of a group of ichthyosaurs porpoising (Fig. 15.11). Similarly, a color plate in the book Prehistoric Animals by Augusta and Burian (1957) depicted a school of Stenopterygius porpoising like modern dolphins. Massare’s (1988) and Motani’s (2002a, b) calculated swimming speeds for ichthyosaurs were too low to allow porpoising, but their estimates were only for cruising speeds. Cowen (1996) considered that moderately-sized ichthyosaurs could swim at high enough speeds to porpoise, providing the necessary time to breathe while conserving energy. His assertion was even accompanied by a limerick,
Fast swimming air breathers are rare
Some ichthyosaurs did it with flair
They swam up in a leap
(It’s energetically cheap)
And they took a deep breath in mid-air.
Although not known to porpoise, lamnid sharks and some other species of sharks have been observed to make spectacular leaps out of the water (Campana et al., 2005; Schwartz, 2013). Unlike porpoising, such leaping behaviors of sharks are not associated with energy conservation. The white shark will perform a “Polaris breach” to ambush pinnipeds and seabirds from beneath and launch the shark and prey completely out of the water (Martin et al., 2005; Hammerschlag et al., 2012). These vertical leaps require sufficient power to deliver the high escape velocity required to propel the whole body into the air.

Illustration of porpoising ichthyosaurs by Heinrich Harder (1858–1935) from “Tiere der Urwelt” (Animals of the Prehistoric World) created in 1916. The work is in the United States Public Domain
15.6 Thermoregulation
As highly active predators, the three taxa have converged on physiological mechanisms to increase locomotor performance. The lamnid shark, ichthyosaur, and dolphin all are considered to be homeothermic (i.e., maintenance of constant body temperature) with a body temperature that is elevated above that of the environmental temperature. Muscles generally perform better at the elevated stable body temperatures of homeotherms compared to the lower body temperatures that is typical of poikilotherms (i.e., body temperature that is the same as the ambient temperature) (Irschick & Higham, 2016). Elevated body temperatures allow for higher maximum swimming speeds, longer and faster sustained swimming speeds, operation in cold water, migratory habits, enhanced digestion and assimilation rates, brain heating, and enhanced visual acuity (Block & Carey, 1985; Wolf et al., 1988; Bernal et al., 2001a, b; Watanabe et al., 2015).
The dolphin, as a mammal, is an endothermic homeotherm (Castellini & Mellish, 2016). Endothermic animals have an elevated metabolism that generates sufficient internal heat for homeothermy. The excess heat is derived from endogenous biochemical, energy-liberating reactions dedicated to the maintenance of vital functions along with increased muscular work. The body temperature of marine mammals, including dolphins, can be up to 35–40 °C above the temperature of polar waters and even 10 °C higher than tropical waters (Berta et al., 2006). The core body is insulated by the dolphin’s large body size, vascular counter-current thermal exchangers, hypodermal layer of blubber, and low surface-to-volume ratios (Berta et al., 2006; Favilla & Costa, 2020). The FR near the optimal value of 4.5 (see above) maximizes the body volume and minimizes surface area for drag reduction while limiting heat loss (Fish, 1993a; Berta et al., 2006; Gutarra et al., 2019).
The body temperature mirrors the ambient water temperature in most fishes, which are ectothermic poikilotherms. Ectothermy refers to a metabolism that is determined by the external ambient temperature. The lack of any insulating mechanism to retain body heat means that any excess heat generated endogenously or from muscle contractions when swimming is rapidly transferred conductively and convectively to the water across the body surface area and from the blood through the gills (Bernal et al., 2001a, 2012). Lamnid sharks do not have any specific thermogenic tissues but are capable of maintaining an elevated body temperature (Bernal et al., 2001b). Sharks expend mechanical and metabolic energy to generate the propulsive power for swimming. Excess heat devoted to homeothermy in the shark is a by-product of the contraction of aerobic red muscle (RM) resulting from constant activation during swimming (Carey et al., 1971). To swim at high speeds, particularly in cold water, requires high muscle power outputs that are fostered by an elevated stable body temperature (Irschick & Higham, 2016).
Like some scombrid fishes (e.g., tuna), lamnid sharks possess a vascular arrangement, known as the rete mirabile, that acts to circumvent heat loss from the body (Carey, 1973; Carey et al., 1971, 1985). The rete is an array of small arteries and veins in juxtaposition that acts as a countercurrent heat exchanger. The rete acts to return heat back to the muscle. All lamnid sharks have retia for heat exchange to support RM endothermy and support maintenance of a body temperature above ambient (Carey & Teal, 1969a; Carey et al., 1985). Mako and porbeagle sharks maintain body temperature 7–10 °C above ambient (Carey & Teal, 1969a), although Bernal et al. (2001b) found only a 0.3–3.0 °C difference in RM temperature for the mako shark compared to the ambient water temperature. Carey et al. (1982) reported that the muscle temperature of the white shark was 5 °C warmer than the ambient water temperature. An increase of muscle temperature of about 5 °C would provide a three-fold increase in overall speed (Irschick & Higham, 2016). McCosker (1987) and Goldman (1997) measured the stomach temperature as a proxy for core body temperature and found it to be 7.4 °C to 14.3 °C above the water temperature for the white shark. Such temperature differentials indicate that lamnid sharks can inhabit cold water and still be active predators that forage for fast and agile prey (Goldman, 1997).
While temperature measurements and anatomical studies of the circulatory specializations can be performed on the extant shark and dolphin to determine their thermoregulatory capabilities, comparative analysis is not directly possible for the extinct ichthyosaur. The inability to take direct temperature and metabolic measurements has made the arguments about homeothermy versus poikilothermy and endothermy versus ectothermy for ichthyosaurs controversial (Motani, 2010). Given the phylogenetic relationship of ichthyosaurs with reptiles the null hypothesis would be that ichthyosaurs were ectothermic poikilotherms. However, the convergence of body plan with the shark and dolphin indicates a large, fast-swimming, highly active ocean predator that would have at least been homeothermic. The analogous ecology of ichthyosaurs, lamnid sharks and oceanic dolphins and the paleoclimatic distribution of ichthyosaurs showing them to have inhabited cold marine waters would indicate adaptations associated with homeothermy (Bernard et al., 2010). Whether this homeothermy was derived endogenously, as in the dolphin, or from RM endothermy, as in the lamnid shark, is subject to debate (Bernard et al., 2010; Motani, 2010).
Based on the bone histology of highly derived ichthyosaurs, de Buffrénil and Mazin (1990) considered that their pattern of bone remodeling was associated with rapid postnatal growth, as indicated in the skeleton of medium and large mammals. Remodeling of the cortical bone was found to be characterized by greater bone reabsorption than bone reconstruction, resulting in a relative loss of bone mass. Similar to remodeling of bone in the dolphin (Delphinus delphis), the compact cortical bone is secondarily reconstructed through ontogeny into cancellous bone (de Buffrénil & Schoevaert, 1988; de Buffrénil & Mazin, 1990). The pattern of bone growth and remodeling was argued to be associated with high endothermic metabolism, incipient endothermy, or gigantothermic metabolism (de Buffrénil & Mazin, 1990). Gigantothermy is defined as the maintenance of a constant high body temperature due to thermal inertia accompanying large body size (Houssaye, 2013) and would indicate homeothermy but not necessarily endothermy.
Evidence for homeothermy in ichthyosaurs was supported by Bernard et al. (2010). When comparing the oxygen isotope (δ18O) compositions of teeth of ichthyosaurs with coexisting fish (i.e., same sedimentary bed). Their results indicated that the body temperature of ichthyosaurs did not vary significantly with seawater temperature, even when the water temperature was as low as 12 °C. The body temperature of ichthyosaurs was considered to be as high as 35 °C. This high body temperature supported the idea that ichthyosaurs had some kind of endothermy. RM endothermy, if not a mammal-like endogenous metabolism, would have been sufficient for homeothermy if there was a large body size (i.e., gigantothermy) and heat conservation mechanisms (e.g., specialized circulatory patterns, blubber) (Motani, 2010). Based on hydrodynamically-derived estimates of optimal swimming speeds, Motani (2002a, b) estimated that the basal metabolic rate of Stenopterygius was between reptilian and mammalian levels (Motani, 2005). Gigantothermy associated with homeothermy is observed in the leatherback sea turtle (Dermochelys coriacea), and also tunas, which have specialized retia for temperature control and RM-generated endothermy (Carey & Teal, 1969b; Carey et al., 1971, 1984; Carey, 1973; Neill et al., 1974; Dizon & Brill, 1979; Paladino et al., 1990; Holland et al., 1992; Lutcavage et al., 1992; Dewar et al., 1994; Holland & Sibert, 1994; Bostrom & Jones, 2007; Casey et al., 2014). Tunas also possess a thick layer of adipose tissue just under the skin, which has been inferred to reduce conductive heat transfer (Carey et al., 1984). Similarly, ichthyosaurs had a layer of adipose tissue under the skin like the blubber layer of dolphins that would help retain body heat for homeothermy (Lindgren et al., 2018; Delsett et al., 2022).
15.7 Conclusions
As evolution is a probabilistic rather than random process that can be directed by similar selection pressures, there is a high probability that multiple lines of divergent taxa will arrive at common solutions to shared environmental challenges and converge on a distinct morphology. The physical nature of the aquatic environment places huge restrictions on the design and functioning of organisms that originally evolved to inhabit a particular niche space. Occupying the same niche space in the aquatic realm could only have occurred in independent clades at separate times or locations. Despite different phylogenies, the iconic example of lamnid sharks, post-Triassic ichthyosaurs, and oceanic dolphins all converged on a homoplasious design in concert with analogous kinematic and physiological mechanisms. The shape of the body and appendages when measured according to fineness ratio, shoulder position, and aspect ratio indicate that all three taxa evolved designs that are optimal for minimizing drag when swimming. This thunniform design, in concert with the use of caudal fin/fluke as an oscillating hydrofoil, allowed these swimmers to efficiently generate thrust for high-speed cruising.
The evolutionary trajectories for each of the three convergent taxa, with their distinct body plans and physiologies, originated for the sharks in the Paleozoic, ichthyosaurs in the Mesozoic, and dolphins in the Cenozoic. The temporal separation allowed each group to develop into highly mobile, fast swimming, epipelagic, apex predators, but the interaction with the physical environment dictated and constrained their eventual convergence. This convergence was not an exact duplication because each taxon independently derived its morphological design, construction, and physiology within the constraints imposed by their divergent phylogenetic trajectories. The necessity for high swimming speed with minimal energy expenditure in a dense, viscous, thermally conductive environment demanded a fusiform body shape, common stabilizing control surfaces, homeothermy, and a lift-based thunniform type of propulsion.
Other animals exhibit convergent designs and adaptations for high-speed swimming performance. Swordfish, sailfish, marlin, tuna and squid are among the fastest swimmers in the ocean which possess fusiform bodies with fineness ratios in the optimal region (Aleyev, 1977; Fish, 2020). The massive great whales (e.g., blue whale, fin whale, sperm whale) are not bulky, but possess highly streamlined bodies (Fish, 1993a). Likewise, other marine mammals like phocid seals, otariid sea lions, and even the slow swimming manatee have converged on the optimal body design (Webb, 1975; Aleyev, 1977; Fish et al., 1988; Fish, 1993a; Kojeszewski & Fish, 2007). Fast swimmers have adapted their caudal propulsor as a rigid lunate fin or flukes to generate lift-based thrust that functions with a high propulsive efficiency (Lighthill, 1969; Webb, 1975; Lindsey, 1978; Fish, 1993a, 1996, 1998a; Fish et al., 2021). The convergence on lift-based propulsion using wing-like structures occurs in sea lions, which use oscillations of their foreflippers for propulsion (Feldkamp, 1987). The speed and power generated by the flippers provides sea lions with the capability to porpoise (Leahy et al., 2021). Such cases of analogy and homoplasy associated with morphologies and mechanics reflective of adaptation to the marine environment give even greater primacy to the convergence of the shark, ichthyosaur, and dolphin as the quintessential example for understanding the relationship between physics and the environment in directing evolutionary change.
References
Abbott, I. H., & von Doenhoff, A. E. (1959). Theory of wing sections. Dover.
Adam, P. J. (2009). Hind limb anatomy. In W. F. Perrin, B. Würsig, & J. G. M. Thewissen (Eds.), Encyclopedia of Marine Mammals (pp. 562–565). Academic Press.
Adams, D. S., & Fish, F. E. (2019). Odontocete peduncle tendons for possible control of fluke orientation and flexibility. Journal of Morphology, 280(9), 1323–1331.
Ahlborn, B. K., Blake, R. W., & Chan, K. H. S. (2009). Optimal fineness ratio for minimum drag in large whales. Canadian Journal of Zoology, 87, 124–131.
Ahmadi, A. R., & Widnall, S. E. (1986). Energetics and optimum motion of oscillating lifting surfaces of finite span. Journal of Fluid Mechanics, 162, 261–282.
Alexander, R. M. N. (1965). The lift produced by the heterocercal tails of selachii. The Journal of Experimental Biology, 43, 131–138.
Aleyev, Y. G. (1977). Nekton. Junk.
Andrews, C. ddW. (1910). A descriptive catalogue of the marine reptiles of the Oxford Clay. British Museum (Natural History), London.
Andrews, R. C. (1921). A remarkable case of external hind limbs in a humpback whale. American Museum Novitates, 9, 1–6.
Ashenberg, J., & Weihs, D. (1984). Minimum induced drag of wings with curved planform. Journal of Aircraft, 21, 89–91.
Au, D., & Weihs, D. (1980). At high speeds dolphins save energy by leaping. Nature, 284(5756), 548–550.
Au, D., Scott, M. D., & Perryman, W. L. (1988). Leap-swim behavior of ‘porpoising’ dolphins. Cetus, 8(1), 7–10.
Augusta, J., & Burian, Z. (1957). Prehistoric animals. Spring Books.
Azuma, A. (1983). Biomechanical aspects of animal flying and swimming. In H. Matsui & K. Kobayashi (Eds.), Biomechanics VIII-A: International series on biomechanics (Vol. 4A, pp. 35–53). Human Kinetics Publishers.
Bandyopadhyay, P. R. (1989). Viscous drag reduction of a nose body. AIAA Journal, 27(3), 274–282.
Bandyopadhyay, P. R., Castano, J. M., Rice, J. Q., Philips, R. B., Nedderman, W. H., & Macy, W. K. (1997). Low-speed maneuvering hydrodynamics of fish and small underwater vehicles. Transactions of the ASME, 119, 136–144.
Bejder, L., & Hall, B. K. (2002). Limbs in whales and limblessness in other vertebrates: Mechanisms of evolutionary and developmental transformation and loss. Evolution and Development, 4, 445–458.
Ben-Zvi, M., & Shadwick, R. E. (2013). Exploring the mechanics of thunniform propulsion: A model study. Canadian Journal of Zoology, 91(10), 741–755.
Bernal, D., Dickson, K. A., Shadwick, R. E., & Graham, J. B. (2001a). Analysis of the evolutionary convergence for high performance swimming in lamnid sharks and tunas. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 129(2–3), 695–726.
Bernal, D., Sepulveda, C., & Graham, J. B. (2001b). Water-tunnel studies of heat balance in swimming mako sharks. The Journal of Experimental Biology, 204(23), 4043–4054.
Bernal, D., Carlson, J. K., Goldman, K. J., & Lowe, C. G. (2012). Energetics, metabolism, and endothermy in sharks and rays. In J. C. Carrier, J. A. Musick, & M. R. Heithaus (Eds.), Biology of Sharks and Their Relatives (2nd ed., pp. 211–237). CRC Press.
Bernard, A., Lécuyer, C., Vincent, P., Amiot, R., Bardet, N., Buffetaut, E., Cuny, G., Fourel, F., Martineau, F., Mazin, J. M., & Prieur, A. (2010). Regulation of body temperature by some Mesozoic marine reptiles. Science, 328(5984), 1379–1382.
Berta, A., Sumich, J. L., & Kovacs, K. M. (2006). Marine mammals: Evolutionary biology. Academic.
Blake, R. W. (1983). Energetics of leaping in dolphins and other aquatic animals. Journal of the Marine Biological Association of the UK, 63(1), 61–70.
Blake, R. W. (1991). Efficiency and economy in animal physiology. Cambridge University Press.
Block, B. A., & Carey, F. G. (1985). Warm brain and eye temperature in sharks. Journal of Comparative Physiology. B, 156, 229–236.
Bonfil, R., Meyër, M. A., Scholl, M. C., Johnson, R. L., O’Brian, S., Oosthuizen, W. H., Swanson, S., Kotze, D., & Patterson, M. (2005). Transoceanic migration, spatial dynamics, and population linkages of white sharks. Science, 310, 100–103.
Bonfil, R., Francis, M. P., Duffy, C., Manning, M. J., & O’Brien, S. (2010). Large-scale tropical movements and diving behavior of white sharks Carcharodon carcharias tagged off New Zealand. Aquatic Biology, 8(2), 115–123.
Bose, N., & Lien, J. (1989). Propulsion of a fin whale (Balaenoptera physalus): Why the fin whale is a fast swimmer. Proceedings of the Royal Society of London B, 237, 175–200.
Bose, N., Lien, J., & Ahia, J. (1990). Measurements of the bodies and flukes of several cetacean species. Proceedings of the Royal Society of London B, 242, 163–173.
Bostrom, B. L., & Jones, D. R. (2007). Exercise warms adult leatherback turtles. Comparative Biochemistry and Physiology A, 147(2), 323–331.
Brawn, V. M. (1962). Physical properties and hydrostatic function of the swimbladder of herring (Clupea harengus L.). Journal of the Fisheries Research Board of Canada, 19, 635–656.
Brooks, D. R. (1996). Explanations of homoplasy at different levels of biological organization. In M. J. Sanderson & L. Hufford (Eds.), Homoplasy: The Recurrence of Similarity in Evolution (pp. 3–36). Academic Press.
Buchholtz, E. A. (2001a). Swimming styles in Jurassic ichthyosaurs. The Journal of Vertebrate Paleontology, 21, 61–73.
Buchholtz, E. A. (2001b). Vertebral osteology and swimming style in living and fossil whales (Order: Cetacea). Journal of Zoology, 253(2), 175–190.
Buchholtz, E. A. (2007). Modular evolution of the cetacean vertebral column. Evolution and Development, 9(3), 278–289.
Buchholtz, E. A., & Schur, S. A. (2004). Vertebral osteology in Delphinidae (cetacea). Zoological Journal of the Linnean Society, 140(3), 383–401.
Buchholtz, E. A., Wolkovich, E. M., & Cleary, R. J. (2005). Vertebral osteology and complexity in Lagenorhynchus acutus (Delphinidae) with comparison to other delphinoid genera. Marine Mammal Science, 21(3), 411–428.
Burcher, R., & Rydill, L. (1994). Concepts in submarine design. Cambridge University Press.
Caldwell, M. W. (2002). From fins to limbs to fins: Limb evolution in fossil marine reptiles. The American Journal of Medical Genetics, 112, 236–249.
Campana, S. E., Marks, L., & Joyce, W. (2005). The biology and fishery of shortfin mako sharks (Isurus oxyrinchus) in Atlantic Canadian waters. Fisheries Research, 73, 341–352.
Carey, F. G. (1973). Fishes with warm bodies. Scientific American, 288(2), 36–44.
Carey, F. G., & Teal, J. M. (1969a). Mako and porbeagle: Warm–bodied sharks. Comparative Biochemistry and Physiology, 28, 199–204.
Carey, F. G., & Teal, J. M. (1969b). Regulation of body temperature by the bluefin tuna. Comparative Biochemistry and Physiology, 28(1), 205–213.
Carey, F. G., Teal, J. M., Kanwisher, J. W., Lawson, K. D., & Beckett, J. S. (1971). Warm-bodied fish. American Zoologist, 11, 137–143.
Carey, F. G., Kanwisher, J. W., Brazier, O., Gabrielson, G., Casey, J. G., & Pratt, H. L., Jr. (1982). Temperature and activities of a white shark, Carcharodon carcharias. Copeia, 1982, 254–260.
Carey, F. G., Kanwisher, J. W., & Stevens, E. D. (1984). Bluefin tuna warm their viscera during digestion. The Journal of Experimental Biology, 109, 1–20.
Carey, F. G., Casey, J. G., Pratt, H. L., Urquhart, D., & McCosker, J. E. (1985). Temperature, heat production and heat exchange in lamnid sharks. Memoirs of the Southern California Academy of Sciences, 9, 92–108.
Casey, J. P., James, M. C., & Williard, A. S. (2014). Behavioral and metabolic contributions to thermoregulation in freely swimming leatherback turtles at high latitudes. The Journal of Experimental Biology, 217(13), 2331–2337.
Castellini, M. A., & Mellish, J.-A. (2016). Thermoregulation. In M. A. Castellini & J.-A. Mellish (Eds.), Marine mammal physiology: Requisites for ocean living (pp. 193–216). CRC Press.
Chang, J. T., & Kim, J. (1996). The measurement of homoplasy: A stochastic view. In M. J. Sanderson & L. Hufford (Eds.), Homoplasy: The recurrence of similarity in evolution (pp. 189–203). Academic Press.
Chopra, M. G. (1976). Large amplitude lunate-tail theory of fish locomotion. Journal of Fluid Mechanics, 74, 161–182.
Chopra, M. G., & Kambe, T. (1977). Hydrodynamics of lunate-tail swimming propulsion. Part 2. Journal of Fluid Mechanics, 79, 49–69.
Cooper, L. N., Dawson, S. D., Reidenberg, J. S., & Berta, A. (2007). Neuromuscular anatomy and evolution of the cetacean forelimb. The Anatomical Record, 290, 1121–1137.
Cooper, L. N., Sedano, N., Johannson, S., May, B., Brown, J., Holliday, C., Kot, W., & Fish, F. E. (2008). Hydrodynamic performance of the minke whale (Balaenoptera acutorostrata) flipper. The Journal of Experimental Biology, 211, 1859–1867.
Cowen, R. (1996). Locomotion and respiration in marine air-breathing vertebrates. In D. Jablonski, D. H. Erwin, & J. H. Lipps (Eds.), Evolutionary biology (pp. 337–352). University of Chicago Press.
Crofts, S. B., Shehata, R., & Flammang, B. E. (2019). Flexibility of heterocercal tails: What can the functional morphology of shark tails tell us about ichthyosaur swimming? Integrative Organismal Biology, 1, obz002.
Daniel, T. (1988). Forward flapping flight from flexible fins. Canadian Journal of Zoology, 66, 630–638.
Daniel, T. L., & Webb, P. W. (1987). Physics, design, and locomotor performance. In P. Dejours, L. Bolis, C. R. Taylor, & E. R. Weibel (Eds.), Comparative physiology: Life in water and on land (pp. 343–369). Liviana Press, Springer.
Daniel, T. L., Jordon, C., & Grunbaum, D. (1992). Hydromechanics of swimming. In R. M. N. Alexander (Ed.), Advances in comparative and environmental physiology (Vol. 11, pp. 17–49). Springer.
Darwin, C. (1859). On the origin of species by means of natural selection. John Murray.
de Buffrénil, V., & Mazin, J. M. (1990). Bone histology of the Ichthyosaurs: Comparative data and functional interpretation. Paleobiology, 16(4), 435–447.
de Buffrénil, V., & Schoevaert, D. (1988). On how the periosteal bone of the delphinid humerus becomes cancellous; ontogeny of a histological specialization. Journal of Morphology, 198, 149–164.
Delair, J. B. (1966). Unusual preservation of fibrous elements in an ichthyosaur skull. Nature, 212(5062), 575–576.
Delsett, L. L., Friis, H., Kölbol-Ebert, M., & Hurum, J. H. (2022). The soft tissue and skeletal anatomy of two Late Jurassic ichthyosaur specimens from the Solnhofen archipelago. PeerJ. https://doi.org/10.7717/peerj.13173
Denny, M. W. (1993). Air and water. Princeton University Press.
Dewar, H., Graham, J., & Brill, R. (1994). Studies of tropical tuna swimming performance in a large water tunnel-thermoregulation. The Journal of Experimental Biology, 192(1), 33–44.
Díaz-Berenguer, E., Badiola, A., Moreno-Azanza, M., & Canudo, J. I. (2018). First adequately-known quadrupedal sirenian from Eurasia (Eocene, Bay of Biscay, Huesca, northeastern Spain). Scientific Reports, 8(1), 1–13.
Díaz-Berenguer, E., Houssaye, A., Badiola, A., & Canudo, J. I. (2019). The hind limbs of Sobrarbesiren cardieli (Eocene, Northeastern Spain) and new insights into the locomotion capabilities of the quadrupedal sirenians. Journal of Mammalian Evolution, 1–27.
Díez, G., Soto, M., & Blanco, J. M. (2015). Biological characterization of the skin of shortfin mako shark Isurus oxyrinchus and preliminary study of the hydrodynamic behaviour through computational fluid dynamics. Journal of Fish Biology, 87, 123–137.
Dizon, A. E., & Brill, R. W. (1979). Thermoregulation in tunas. American Zoologist, 19(1), 249–265.
Domning, D. P. (2000). The readaptation of Eocene sirenians to life in water. Historical Biology, 14, 115–119.
Domning, D. P. (2001). The earliest known fully quadrupedal sirenian. Nature, 413, 625–627.
Donley, J. M., Sepulveda, C. A., Konstantinidis, P., Gemballa, S., & Shadwick, R. E. (2004). Convergent evolution in mechanical design of lamnid sharks and tunas. Nature, 429, 61–65.
Elsner, R., & Meiselman, H. J. (1995). Splenic oxygen storage and blood viscosity in seals. Marine Mammal Science, 11, 93–95.
Favilla, A. B., & Costa, D. P. (2020). Thermoregulatory strategies of diving air-breathing marine vertebrates: A review. Frontiers in Ecology and Evolution, 8, 555509.
Fedak, T. J., & Hall, B. K. (2004). Perspectives on hyperphalangy: Patterns and processes. Journal of Anatomy, 204, 151–163.
Feldkamp, S. D. (1987). Foreflipper propulsion in the California sea lion Zalophus californianus. Journal of Zoology (London), 212, 43–57.
Felts, W. J. L. (1966). Some functional and structural characteristics of cetacean flippers and flukes. In K. S. Norris (Ed.), Whales, dolphins, and porpoises (pp. 255–276). University of California Press.
Fierstine, H. L., & Walters, V. (1968). Studies of locomotion and anatomy of scombrid fishes. Memoirs of the Southern California Academy of Sciences, 6, 1–31.
Fischer, V., Bardet, N., Benson, R. B. J., Arkhangelsky, M. S., & Friedman, M. (2016). Extinction of fish-shaped marine reptiles associated with reduced evolutionary rates and global environmental volatility. Nature Communications, 7, 10825.
Fish, F. E. (1992). Aquatic locomotion. In T. Tomasi & T. Horton (Eds.), Mammalian energetics: Interdisciplinary views of metabolism and reproduction (pp. 34–64). Cornell University Press.
Fish, F. E. (1993a). Influence of hydrodynamic design and propulsive mode on mammalian swimming energetics. Australian Journal of Zoology, 42, 79–101.
Fish, F. E. (1993b). Power output and propulsive efficiency of swimming bottlenose dolphins (Tursiops truncatus). The Journal of Experimental Biology, 185, 179–193.
Fish, F. E. (1996). Transitions from drag-based to lift-based propulsion in mammalian aquatic swimming. American Zoologist, 36, 628–641.
Fish, F. E. (1998a). Comparative kinematics and hydrodynamics of odontocete cetaceans: Morphological and ecological correlates with swimming performance. The Journal of Experimental Biology, 201, 2867–2877.
Fish, F. E. (1998b). Biomechanical perspective on the origin of cetacean flukes. In J. G. M. Thewissen (Ed.), The emergence of whales: Evolutionary patterns in the origin of Cetacea (pp. 303–324). Plenum Press.
Fish, F. E. (2000). Biomechanics and energetics in aquatic and semiaquatic mammals: Platypus to whale. Physiological and Biochemical Zoology, 73(6), 683–698.
Fish, F. E. (2001). Mechanism for evolutionary transition in swimming mode by mammals. In J.-M. Mazin, P. Vignaud, & V. de Buffrénil (Eds.), Secondary adaptation of tetrapods to life in water (pp. 261–287). Verlag Dr. Friedrich Pfeil.
Fish, F. E. (2002). Balancing requirements for stability and maneuverability in cetaceans. Integrative and Comparative Biology, 42, 85–93.
Fish, F. E. (2004). Structure and mechanics of nonpiscine control surfaces. IEEE Journal of Oceanic Engineering, 28, 605–621.
Fish, F. E. (2015). Estimation of shape of the sea lion for hydrodynamic analysis. Response to ‘Cambered profile of a California sea lion’s body’. The Journal of Experimental Biology, 218, 1271–1272.
Fish, F. E. (2018). Streamlining. In W. F. Perrin, B. Würsig, & J. G. M. Thewissen (Eds.), Encyclopedia of marine mammals (3 edn, pp. 951–954). Academic.
Fish, F. E. (2020). Advantages of aquatic animals as models for bio-inspired drones over present AUV technology. Bioinspiration & Biomimetics, 15, 025001.
Fish, F. E., & Battle, J. M. (1995). Hydrodynamic design of the humpback whale flipper. Journal of Morphology, 225, 51–60.
Fish, F. E., & Beneski, J. T. (2014). Evolution and bio-inspired design: Natural limitations. In A. Goel, D. A. McAdams, & R. B. Stone (Eds.), Biologically inspired design: Computational methods and tools (pp. 287–312). Springer.
Fish, F. E., & Hui, C. A. (1991). Dolphin swimming: A review. Mamm. Rev., 21, 181–196.
Fish, F. E., & Lauder, G. V. (2006). Passive and active flow control by swimming fishes and mammals. Annual Review of Fluid Mechanics, 38, 193–224.
Fish, F. E., & Lauder, G. V. (2017). Control surfaces of aquatic vertebrates in relation to swimming modes. The Journal of Experimental Biology, 220, 4351–4363.
Fish, F. E., & Rohr, J. (1999). Review of dolphin hydrodynamics and swimming performance. SPAWARS Systems Center Technical Repeort. 1801, San Diego, CA.
Fish, F. E., & Shannahan, L. D. (2000). The role of the pectoral fins in body trim of sharks. Journal of Fish Biology, 56, 1062–1073.
Fish, F. E., Innes, S., & Ronald, K. (1988). Kinematics and estimated thrust production of swimming harp and ringed seals. The Journal of Experimental Biology, 137, 157–173.
Fish, F. E., Smelstoys, J., Baudinette, R. V., & Reynolds, P. S. (2002). Fur doesn’t fly, it floats: Buoyancy of hair in semi-aquatic mammals. Aquatic Mammals, 28(2), 103–112.
Fish, F. E., Nusbaum, M. K., Beneski, J. T., & Ketten, D. R. (2006). Passive cambering and flexible propulsors: Cetacean flukes. Bioinspiration & Biomimetics, 1, S42–S48.
Fish, F. E., Beneski, J. T., & Ketten, D. R. (2007). Examination of the three-dimensional geometry of cetacean flukes using CT-scans: hydrodynamic implications. The Anatomical Record, 290, 614–623.
Fish, F. E., Howle, L. E., & Murray, M. M. (2008). Hydrodynamic flow control in marine mammals. Integrative and Comparative Biology, 211, 1859–1867.
Fish, F. E., Legac, P., Williams, T. M., & Wei, T. (2014). Measurement of hydrodynamic force generation by swimming dolphins using bubble DPIV. The Journal of Experimental Biology, 217(2), 252–260.
Fish, F. E., Williams, T. M., Sherman, E., Moon, Y. E., Wu, V., & Wei, T. (2018). Experimental measurement of dolphin thrust generated during a tail stand using DPIV. Fluids, 2018(3), 33.
Fish, F. E., Rybczynski, N., Lauder, G. V., & Duff, C. M. (2021). The evolution of the tail and lack thereof for aquatic propulsion. Integrative and Comparative Biology, 64, 398–413. https://doi.org/10.1093/icb/icab021
Flammang, B. E. (2010). Functional morphology of the radialis muscle in shark tails. Journal of Morphology, 271, 340–352.
Flammang, B. E., Lauder, G. V., Troolin, D. R., & Strand, T. (2011). Volumetric imaging of shark tail hydrodynamics reveals a three-dimensional dual-ring vortex wake structure. Proceedings of the Royal Society B: Biological Sciences, 278(1725), 3670–3678.
Fox, R. W., Prichard, P. J., & McDonald, A. T. (2009). Introduction to fluid mechanics. Wiley.
Gabler, U. (2000). Submarine design. Bernard & Graefe.
Gans, C. (1974). Biomechanics: An approach to vertebrate biology. Lippincott.
Garten, J. L., & Fish, F. E. (2020). Comparative histological examination of the integument of cetacean flukes. Aquatic Mammals, 46(4), 367–381.
Gertler, M. (1950). Resistance experiments on a systematic series of streamlined bodies of revolution-for application to the design of high-speed submarines. Rep. C-297. David W. Taylor Model Basin.
Gingerich, P. D. (2015). Evolution of whales from land to sea. In K. P. Dial, N. Shubin, & E. L. Brainerd (Eds.), Great transformation in vertebrate evolution (pp. 239–256). University of Chicago Press.
Gingerich, P. D., Smith, B. H., & Simons, E. L. (1990). Hind limbs of Eocene Basilosaurus: Evidence of feet in whales. Science, 249, 154–157.
Gingerich, P. D., Raza, S. M., Arif, M., Anwar, M., & Zhou, X. (1994). New whale from the Eocene of Pakistan and the origin of cetacean swimming. Science, 368, 844–847.
Gingerich, P. D., ul Haq, M., Zalmout, I. S., Khan, I. H., & Malkani, M. S. (2001). Origin of whales from early artiodactyls: hands and feet of Eocene Protocetidae from Pakistan. Science, 293, 2239–2242.
Goldbogen, J. A., Cade, D. E., Wisniewska, D. M., Potvin, J., Segre, P. S., Savoca, M. S., Hazen, E. L., Czapanskiy, M. F., Kahane-Rapport, S. R., DeRuiter, S. L., & Gero, S. (2019). Why whales are big but not bigger: Physiological drivers and ecological limits in the age of ocean giants. Science, 366, 1367–1372.
Goldman, K. J. (1997). Regulation of body temperature in the white shark, Carcharodon carcharias. Journal of Comparative Physiology B, 167(6), 423–429.
Gordon, C. N. (1980). Leaping dolphins. Nature, 287, 759.
Gough, W. T., Fish, F. E., Wainwright, D. K., & Bart-Smith, H. (2018). Morphology of the core fibrous layer of the cetacean tail fluke. Journal of Morphology, 279(6), 757–765.
Gould, S. J. (2002). The structure of evolutionary theory. Belknap Press.
Gutarra, S., Moon, B. C., Rahman, I. A., Palmer, C., Lautenschlager, S., Brimacombe, A. J., & Benton, M. J. (2019). Effects of body plan evolution on the hydrodynamic drag and energy requirements of swimming in ichthyosaurs. Proceedings of the Royal Society B, 286(1898), 20182786.
Haas, O., & Simpson, G. G. (1946). Analysis of some phylogenetic terms, with attempts at redefinition. Proceedings. American Philosophical Society, 90, 319–349.
Hall, B. K. (1984). Developmental mechanisms underlying the formation of atavisms. Biological Reviews, 59, 89–122.
Hamilton, J. L., Dillaman, R. M., McLellan, W. A., & Pabst, D. A. (2004). Structural fiber reinforcement of keel blubber in harbor porpoise (Phocoena phocoena). Journal of Morphology, 261, 105–117.
Hammerschlag, N., Martin, R. A., Fallows, C., Collier, R. S., & Lawrence, R. (2012). Investigatory behavior toward surface objects and nonconsumptive strikes on seabirds by white sharks, Carcharodon carcharias, at Seal Island, South Africa (1997–2010). In M. L. Domeier (Ed.), Global perspectives on the biology and life history of the white shark (pp. 91–103).
Harris, J. E. (1936). The role of the fins in the equilibrium of the swimming fish. I. Wind-tunnel tests on a model of Mustelus canis (Mitchill). The Journal of Experimental Biology, 13, 476–493.
Harris, J. E. (1938). The role of the fins in the equilibrium of the swimming fish. II. The role of the pelvic fins. The Journal of Experimental Biology, 15, 32–47.
Hertel, H. (1966). Structure, form, movement. Reinhold.
Hildebrand, M. (1995). Analysis of vertebrate structure. Wiley.
Hoerner, S. F. (1965). Fluid-dynamic drag. Published by Author.
Hoffman, S. L., Buser, T. J., & Porter, M. E. (2020). Comparative morphology of shark pectoral fins. J. Morph., 2020, 1–16.
Holland, K. N., & Sibert, J. R. (1994). Physiological thermoregulation in bigeye tuna, Thunnus obesus. Environmental Biology of Fishes, 40(3), 319–327.
Holland, K. N., Brill, R. W., Chang, R. K., Sibert, J. R., & Fournier, D. A. (1992). Physiological and behavioural thermoregulation in bigeye tuna (Thunnus obesus). Nature, 358(6385), 410–412.
Houssaye, A. (2013). Bone histology of aquatic reptiles: What does it tell us about secondary adaptation to an aquatic life? Biological Journal of the Linnean Society, 108(1), 3–21.
Howell, A. B. (1930). Aquatic Mammals. Charles C Thomas.
Huelsmann, M., Hecker, N., Springer, M. S., Gatesy, J., Sharma, V., & Hiller, M. (2019). Genes lost during the transition from land to water in cetaceans highlight genomic changes associated with aquatic adaptations. Science Advances, 5(9), eaaw6671.
Hui, C. A. (1987). The porpoising of penguins: An energy-conserving behavior for respiratory ventilation? Canadian Journal of Zoology, 65, 209–211.
Hui, C. A. (1989). Surfacing behavior and ventilation of free-ranging dolphins. Journal of Mammalogy, 70, 833–835.
Irving, L. (1966). Elective regulation of the circulation in diving animals. In K. S. Norris (Ed.), Whales, dolphins, and porpoises (pp. 381–395). University of California Press.
Irving, L., Scholander, P., & Grinnell, S. (1941). The respiration of the porpoise Tursiops truncatus. Journal of Cellular and Comparative Physiology, 17, 145–168.
Irschick, D. J., & Higham, T. E. (2016). Animal athletes: An ecological and evolutionary approach. Oxford University Press.
Kabasakal, H., & Kabasakal, Ö. (2013). First record of a shortfin mako shark, Isurus oxyrinchus Rafinesque, 1810 (Chondrichthyes: Lamnidae) from the Bay of Saroz (NE Aegean Sea). Annales, Series Historia Naturalis, 23(1), 27–32.
Kardong, K. V. (2019). Vertebrates: Comparative anatomy, function, evolution. New York.
Kelley, N. P., & Motani, R. (2015). Trophic convergence drives morphological convergence in marine tetrapods. Biology Letters, 11, 20140709.
Kelley, N. P., & Pyenson, N. D. (2015). Evolutionary innovation and ecology in marine tetrapods from Triassic to Anthropocene. Science, 348(6232), aaa3716.
Kemp, N. E. (1977). Banding pattern and fibrillogenesis of ceratotrichia in shark fins. Journal of Morphology, 154(2), 187–203.
Kim, S. H., Shimada, K., & Rigsby, C. K. (2013). Anatomy and evolution of heterocercal tail in lamniform sharks. The Anatomical Record, 296(3), 433–442.
Kojeszewski, T., & Fish, F. E. (2007). Swimming kinematics of the Florida manatee (Trichechus manatus latirostris): Hydrodynamic analysis of an undulatory mammalian swimmer. The Journal of Experimental Biology, 210, 2411–2418.
Kooyman, G. L. (1973). Respiratory adaptations in marine mammals. American Zoologist, 13, 457–468.
Kooyman, G. L. (1989). Diverse divers: Physiology and behavior. Springer.
Kooyman, G., & Cornell, L. (1981). Flow properties of expiration and inspiration in a trained bottle-nosed porpoise. Physiological Zoology, 54, 55–61.
Küchermann, D. (1953). The distribution of lift over the surface of swept wings. Aeronautical Quarterly, 4, 261–278.
Lambert, O., Bianucci, G., Salas-Gismondi, R., Di Celma, C., Steurbaut, E., Urbina, M., & de Muizon, C. (2019). An amphibious whale from the middle Eocene of Peru reveals early South Pacific dispersal of quadrupedal cetaceans. Current Biology, 29, 1352–1359.
Lang, T. G. (1963). Porpoise, whale, and fish: Comparison of predicted and observed speeds. Naval Engineers Journal, 75, 437–441.
Lang, T. G. (1966). Hydrodynamic analysis of dolphin fin profiles. Nature, 209, 1110–1111.
Lang, T. G., & Daybell, D. A. (1963). Porpoise performance tests in a seawater tank. Naval Ordnance Test Station Technical Report 3063.
Lang, A. W., Motta, P., Hidalgo, P., & Westcott, M. (2008). Bristled shark skin: A microgeometry for boundary layer control? Bioinspiration & Biomimetics, 3(4), 046005.
Larson, A., & Losos, J. B. (1996). Phylogenetic systematics of adaptation. In M. R. Rose & G. V. Lauder (Eds.), Adaptation (pp. 187–220). Academic Press.
Leahy, A. M., Fish, F. E., Kerr, S. J., Zeligs, J. A., Skrovan, S., Cardenas, K. L., Leftwich, M. C. (2021). The role of California sea lion (Zalophus californianus) hindflippers as aquatic control surfaces for maneuverability. J. Exp. Biol. JEXBIO/2021/242388 (In revision).
Liem, K. F. (1990). Aquatic versus terrestrial feeding modes: Possible impacts on the trophic ecology of vertebrates. American Zoologist, 30, 209–221.
Liem, K. F., Bemis, W. E., Walker, W. F., Jr., & Grande, L. (2001). Functional anatomy of the vertebrates: An evolutionary perspective. Harcourt.
Lighthill, J. (1969). Hydrodynamics of aquatic animal propulsion – A survey. Annual Review of Fluid Mechanics, 1, 413–446.
Lighthill, J. (1970). Aquatic animal propulsion of high hydromechanical efficiency. Journal of Fluid Mechanics, 44, 265–301.
Lindgren, J., Sjövall, P., Thiel, V., Zheng, W., Ito, S., Wakamatsu, K., Hauff, R., Kear, B. P., Engdahl, A., Alwmark, C., Eriksson, M. E., Jarenmark, M., Sachs, S., Ahlberg, P. E., Marone, F., Kuriyama, T., Gustafsson, O., Malmberg, P., Thomen, A., Rodríguez-Meizoso, I., Uvdal, P., Ojika, M., & Schweitzer, M. H. (2018). Soft-tissue evidence for homeothermy and crypsis in a Jurassic ichthyosaur. Nature, 564, 359–365.
Lindsey, C. C. (1978). Form, function, and locomotory habits in fish. In W. S. Hoar & D. J. Randall (Eds.), Fish physiology: Locomotion (Vol. 7, pp. 1–100). Academic Press.
Lingham-Soliar, T. (1999). Rare soft tissue preservation showing fibrous structures in an ichthyosaur from the Lower Lias (Jurassic) of England. Proceedings Royal Society London Series B: Biological Science, 266, 2367–2373.
Lingham-Soliar, T. (2001). The ichthyosaur integument: Skin fibers, a means for a strong, flexible and smooth skin. Lethaia, 34(4), 287–302.
Lingham-Soliar, T. (2004). Caudal fin allometry in the white shark Carcharodon carcharias: Implications for locomotory performance and ecology. Naturwissenschaften, 92, 231–236.
Lingham-Soliar, T. (2005a). Dorsal fin in the white shark, Carcharodon carcharias: A dynamic stabilizer for fast swimming. Journal of Morphology, 260, 1–11.
Lingham-Soliar, T. (2005b). Caudal fin in the white shark, Carcharodon carcharias: A dynamic propeller for fast, efficient swimming. Journal of Morphology, 264, 233–252.
Lingham-Soliar, T. (2016). Convergence in thunniform anatomy in lamnid sharks and Jurassic ichthyosaurs. Integrative and Comparative Biology, 56, 1323–1336.
Lingham-Soliar, T., & Plodowski, G. (2007). Taphonomic evidence for high-speed adapted fin in thunniform ichthyosaurs. Naturwissenschaften, 94, 65–70.
Lingham-Soliar, T., & Reif, W.-E. (1998). Taphonomic evidence for fast tuna-like swimming in Jurassic and Cretaceous ichthyosaurs. Neues Jahrbuch für Geologie und Paläontologie Abh, 207(2), 171–183.
Lingham-Soliar, T., & Wesley-Smith, J. (2008). First investigation of the collagen D-band ultrastructure in fossilized vertebrate integument. Proceedings Royal Society London Series B: Biological Science, 275(1648), 2207–2212.
Liu, P., & Bose, N. (1993). Propulsive performance of three naturally occurring oscillating propeller planforms. Ocean Engineering, 20, 57–75.
Long, J. H., Jr., Pabst, D. A., Shepherd, W. R., & McLellan, W. A. (1997). Locomotor design of dolphin vertebral columns: Bending mechanics and morphology of Delphinus delphis. The Journal of Experimental Biology, 200, 65–81.
Lovvorn, J. R., Croll, D. A., & Liggins, G. A. (1999). Mechanical versus physiological determinants of swimming speeds in diving Brunnich’s guillemots. The Journal of Experimental Biology, 202, 1741–1752.
Lutcavage, M. E., Bushnell, P. G., & Jones, D. R. (1992). Oxygen stores and aerobic metabolism in the leatherback sea turtle. Canadian Journal of Zoology, 70(2), 348–351.
Martin, R. A., Hammerschlag, N., Collier, R. S., & Fallows, C. (2005). Predatory behaviour of white sharks (Carcharodon carcharias) at Seal Island, South Africa. Journal of the Marine Biological Association of the UK, 85, 1121–1136.
Massare, J. A. (1988). Swimming capabilities of Mesozoic marine reptiles: Implications for method of predation. Paleobiology, 14, 187–205.
Massare, J. A., & Lomax, D. R. (2019). Hindfins of Ichthyosaurus: Effects of large sample size on ‘distinct’ morphological characters. Geological Magazine, 156, 725–744.
Matta, A., Bayandor, J., Battaglia, F., & Pendar, H. (2019). Effects of fish caudal fin sweep angle and kinematics on thrust production during low-speed thunniform swimming. Biology Open, 8, bio040626.
Maxwell, E. E. (2012). Unraveling the influences of soft-tissue flipper development on skeletal variation using an extinct taxon. The Journal of Experimental Zoology Part B, 318, 545–554.
McClain, C. R., Balk, M. A., Benfield, M. C., Branch, T. A., Chen, C., Cosgrove, J., Dove, A. D., Gaskins, L. C., Helm, R. R., Hochberg, F. G., & Lee, F. B. (2015). Sizing ocean giants: Patterns of intraspecific size variation in marine megafauna. PeerJ, 3, e715.
McCosker, J. E. (1987). The white shark, Carcharodon carcharias, has a warm stomach. Copeia, 1987(1), 195–197.
McGowan, C. (1991). Dinosaurs, Spitefires, & Sea Dragons. Harvard University Press.
McGowan, C. (1992). The ichthyosaurian tail: Sharks do not provide an appropriate analogue. Palaeontology, 35, 555–570.
Mitani, Y., Andrews, R. D., Sato, K., Kato, A., Naito, Y., & Costa, D. P. (2010). Three-dimensional resting behaviour of northern elephant seals: Drifting like a falling leaf. Biology Letters, 6, 163–166.
Miyazaki, N., Kasuya, T., & Nishiwaki, M. (1974). Distribution and migration of two species of Stenella in the Pacific coast of Japan. The Scientific Reports of the Whales Research Institute, Tokyo, 26, 227–243.
Moore, M. J., Hammar, T., Arruda, J., Cramer, S., Dennison, S., Montie, E., & Fahlman, A. (2011). Hyperbaric computed tomographic measurement of lung compression in seals and dolphins. The Journal of Experimental Biology, 214, 2390–2397.
Motani, R. (2000). Rulers of the Jurassic seas. Scientific American, 283, 52–59.
Motani, R. (2002a). Scaling effects in caudal fin propulsion and the speed of ichthyosaurs. Nature, 415, 309–312.
Motani, R. (2002b). Swimming speed estimation of extinct marine reptiles: Energetic approach revisited. Paleobiology, 28(2), 251–262.
Motani, R. (2005). Evolution of fish-shaped reptiles (Reptilia: Ichthyopterygia) in their physical environments and constraints. Annual Review of Earth and Planetary Sciences, 33, 395–420.
Motani, R. (2010). Warm-blooded “sea dragons”? Science, 328(5984), 1361–1362.
Motani, R., You, H., & McGowan, C. (1996). Eel-like swimming in the earliest ichthyosaurs. Nature, 382(6589), 347–348.
Neill, W. H., Stevens, E. D., Carey, F. G., Lawson, K. D., Mrosovsky, N., & Frair, W. (1974). Thermal inertia versus thermoregulation in “warm” turtles and tunas. Science, 184(4140), 1008–1010.
Nesteruk, I. (2020). Fastest fish shapes and optimal supercavitating and hypersonic bodies of revolution. Innovative Biosystems and Bioengineering, 4(4), 169–178.
Noren, S. R., Lacave, G., Wells, R. S., & Williams, T. M. (2002). The development of blood oxygen stores in bottlenose dolphins (Tursiops truncatus): implications for diving capacity. Journal of Zoology London, 258, 105–113.
Nursall, J. R. (1962). Swimming and the origin of paired appendages. American Zoologist, 2, 127–141.
Oeffner, J., & Lauder, G. V. (2012). The hydrodynamic function of shark skin and two biomimetic applications. The Journal of Experimental Biology, 215(5), 785–795.
Ohsumi, S. (1965). A dolphin (Stenella caeruleoalba) with protruded rudimentary hind limbs. The Scientific Reports of the Whales Research Institute, 19, 135–136.
Pabst, D. A. (1990). Axial muscles and connective tissues of the bottlenose dolphin. In S. Leatherwood & R. R. Reeves (Eds.), The Bottlenose dolphin (pp. 51–67). Academic Press.
Pabst, D. A. (1996). Morphology of the subdermal connective tissue sheath of dolphins: A new fibre-wound, thin-walled, pressurized cylinder model for swimming vertebrates. Journal of Zoology London, 238, 35–52.
Pabst, D. A. (2000). To bend a dolphin: Convergence of force transmission designs in cetaceans and scombrid fishes. American Zoologist, 40, 146–155.
Paladino, F. V., O’Connor, M. P., & Spotila, J. R. (1990). Metabolism of leatherback turtles, gigantothermy, and thermoregulation of dinosaurs. Nature, 344(6269), 858–860.
Parry, D. A. (1949). The swimming of whales and a discussion of Gray’s paradox. The Journal of Experimental Biology, 26, 24–34.
Patterson, C. (1988). Homology in classical and molecular biology. Molecular Biology and Evolution, 5, 603–625.
Pennycuick, C. J. (1992). Newton rules biology: A physical approach to biological problems. Oxford University Press.
Pershin, S. V. (1983). Hydrobionic features for optimizing the exterior forms of the propeller system in cetaceans. Bionika, 17, 13–24. (translated from Russian).
Pershin, S. V., Chernyshov, L. F., Kozlov, L. F., Koval, A. P., & Zayets, V. A. (1976). Patterns in the integuments of fast-swimming fishes. Bionika, 10, 3–21.
Powell, R. (2007). Is convergence more than an analogy? Homoplasy and its implications for macroevolutionary predictability. Biology and Philosophy, 22, 565–578.
Radinsky, L. B. (1987). The evolution of vertebrate design. University of Chicago.
Rayner, J. M. V. (1985). Vorticity and propulsion mechanics in swimming and flying animals. In J. Riess & E. Frey (Eds.), Konstruktionsprinzipen lebender und ausgestorbener Reptilien (pp. 89–118). University of Tubingen.
Rayner, J. M. (1986). Pleuston: Animals which move in water and air. Endeavour, 10(2), 58–64.
Reidy, L. W. (1987). Flat plate drag reduction in a water tunnel using riblets. NOSC Technical Report 1169.
Reif, W. E. (1978). Protective and hydrodynamic function of the dermal skeleton of elasmobranchs. Neues Jahrbuch für Geologie und Paläontologie, 157, 133–141.
Reif, W. E. (1985). Squamation and ecology of sharks. Senckenbergische Naturforschende Gesellschaft.
Reif, W. E., & Dinkelacker, A. (1982). Hydrodynamics of the squamation in fast swimming sharks. Neues Jahrbuch für Geologie und Paläontologie, 164, 184–187.
Reif, W. E., & Weishampel, D. B. (1986). Anatomy and mechanics of the lunate tail in lamnid sharks. Zoologische Jahrbücher Anatomie, 114, 221–234.
Ridgway, S. H., & Howard, R. (1979). Dolphin lung collapse and intramuscular circulation during free diving: Evidence from nitrogen washout. Science, 206, 1182–1183.
Ridgway, S. H., & Johnston, D. G. (1966). Blood oxygen and ecology of porpoises of three genera. Science, 151, 456–457.
Ridgway, S. H., Scronce, B. L., & Kanwisher, J. (1969). Respiration and deep diving in the bottlenose porpoise. Science, 166, 1651–1654.
Riess, J. (1986). Fortbewegunsweise, Schwimmbiophysik und Phylogenie der Ichthyosaurier. Palaeontographica A, 192, 93–155.
Rohr, J. J., & Fish, F. E. (2004). Strouhal numbers and optimization of swimming by odontocete cetaceans. The Journal of Experimental Biology, 207(10), 1633–1642.
Saadat, M., Fish, F. E., Domel, A., Di Santo, V., Lauder, G. V., & Haj-Hariri, H. (2017). On the rules for aquatic locomotion. Physical Review Fluids, 2, 083102.
Schmidt-Nielsen, K. (1972). Locomotion: Energy cost of swimming, flying, and running. Science, 177, 222–228.
Scholander, P. F. (1940). Experimental investigations on the respiratory function in diving mammals and birds. Hvalradets Skrifter, 22, 1–131.
Schwartz, F. J. (2013). Jumping and spinning by carcharhinid sharks: Another view. The Journal of the North Carolina Academy of Science, 129(3), 107–110.
Sfakiotakis, M., Lane, D. M., & Davies, J. B. C. (1999). Review of fish swimming modes for aquatic locomotion. IEEE Journal of Oceanic Engineering, 24(2), 237–252.
Shadwick, R. E. (2005). How tuna and lamnid sharks swim: An evolutionary convergence. American Scientist, 93, 524–531.
Shadwick, R. E., & Gemballa, S. (2005). Structure, kinematics, and muscle dynamics in undulatory swimming. Fish Physiology, 23, 241–280.
Shoemaker, P. A., & Ridgway, S. H. (1991). Cutaneous ridges in odontocetes. Marine Mammal Science, 7(1), 66–74.
Skrovan, R. C., Williams, T. M., Berry, P. S., Moore, P. W., & Davis, R. W. (1999). The diving physiology of bottlenose dolphins (Tursiops truncatus). II. Biomechanics and changes in buoyancy at depth. The Journal of Experimental Biology, 202(20), 2749–2761.
Smits, A. J. (2019). Undulatory and oscillatory swimming. Journal of Fluid Mechanics, 874, P1. https://doi.org/10.1017/jfm.2019.284
Standen, E. M. (2008). Pelvic fin locomotor function in fishes: Three-dimensional kinematics in rainbow trout (Oncorhynchus mykiss). The Journal of Experimental Biology, 211, 2931–2942.
Stayton, C. T. (2015). The definition, recognition, and interpretation of convergent evolution, and two new measures for quantifying and assessing the significance of convergence. Evolution, 69(8), 2140–2153.
Sternes, P. C., & Shimada, K. (2020). Body forms in sharks (Chondrichthyes: Elasmobranchii) and their functional, ecological, and evolutionary implications. Zoology, 140, 125799.
Streitlien, K., & Triantafyllou, G. S. (1998). On thrust estimates for flapping foils. The Journal of Fluids and Structures, 12, 47–55.
Struthers, J. (1893). On the rudimentary hind-limb of a great fin-whale (Balœnoptera musculus) in comparison with those of the humpback whale and the Greenland right-whale. Journal of Anatomy and Physiology, 27(3), 291–335.
Sun, Q., Morikawa, H., Kobayashi, S., Ueda, K., Miyahara, H., & Nakashima, M. (2010a). Structure and mechanical properties on tail flukes of dolphin. Journal of Aero Aqua Bio-Mechanics, 1(1), 45–50.
Sun, Q., Morikawa, H., Kobayashi, S., Ueda, K., Miyahara, H., & Nakashima, M. (2010b). Structure and bending properties of central part of tail fin of dolphin. Journal of Biomechanical Science and Engineering, 5, 388–398.
Sun, Q., Morikawa, H., Ueda, K., Miyahara, H., & Nakashima, M. (2011). Bending properties of tail flukes of dolphin. Journal of Biomechanical Science and Engineering, 6(1), 15–25.
Taylor, M. A. (1987). A reinterpretation of ichthyosaur swimming and buoyancy. Palaeontology, 30, 531–535.
Taylor, A. R., Schacke, J. H., Speakman, T. R., Castleberry, S. B., & Chandler, R. B. (2016). Factors related to common bottlenose dolphin (Tursiops truncatus) seasonal migration along South Carolina and Georgia coasts, USA. Animal Migration, 1, 14–26.
Thewissen, J. G. M. (2018). Highlights of cetacean embryology. Aquatic Mammals, 44, 591–602.
Thewissen, J. G., & Bajpai, S. (2001). Whale origins as a poster child for macroevolution. Bioscience, 51, 1037–1049.
Thewissen, J. G. M., & Fish, F. E. (1997). Locomotor evolution in the earliest cetaceans: Functional model, modern analogues, and paleontological evidence. Paleobiology, 23(4), 482–490.
Thewissen, J. G. M., & Nummela, S. (2008). Introduction: On becoming aquatic. In J. G. M. Thewissen & S. Nummela (Eds.), Sensory evolution on the threshold: Adaptations in secondarily aquatic vertebrates (pp. 1–25). University of California Press.
Thewissen, J. G. M., Cohn, M. J., Stevens, L. S., Bajpai, S., Heyning, J., & Horton, W. E. (2006). Developmental basis for hind-limb loss in dolphins and origin of the cetacean bodyplan. Proceedings of the National Academy of Sciences of the United States of America, 103(22), 8414–8418.
Thewissen, J. G., Cooper, L. N., Clementz, M. T., Bajpai, S., & Tiwari, B. N. (2007). Whales originated from aquatic artiodactyls in the Eocene epoch of India. Nature, 450(7173), 1190–1194.
Thewissen, J. G. M., Cooper, L. N., George, J. C., & Bajpai, S. (2009). From land to water: The origin of whales, dolphins, and porpoises. Evolution: Education and Outreach, 2, 272–288.
Thomson, K. S., & Simanek, E. D. (1977). Body form and locomotion of sharks. American Zoologist, 17, 343–354.
Triantafyllou, M. S. (2017). Tuna fin hydraulics inspire aquatic robotics. Science, 357, 251–252.
Triantafyllou, G. S., & Triantafyllou, M. S. (1995). An efficient swimming machine. Scientific American, 272, 40–48.
Triantafyllou, G. S., Triantafyllou, M. S., & Grosenbaugh, M. A. (1993). Optimal thrust development in oscillating foils with applications to fish propulsion. The Journal of Fluids and Structures, 7, 205–224.
Triantafyllou, M. S., Triantafyllou, G. S., & Yue, D. K. P. (2000). Hydrodynamics of fishlike swimming. Annual Review of Fluid Mechanics, 32, 33–53.
Tsai, W-L. (1998). Dorsoventral bending of the tail and functional morphology of the caudal vertebrae in the bottlenose dolphin, Tursiops truncatus. Master’s thesis. Oklahoma State University.
Tucker, V. A. (1970). Energetic cost of locomotion in animals. Comparative Biochemistry and Physiology, 34, 841–846.
Tucker, V. A. (1975). The energetic cost of moving about. American Scientist, 63, 413–419.
Uhen, M. D. (2010). The origin(s) of whales. Annual Review of Earth and Planetary Sciences, 38, 189–219.
Van Buren, T., Floryan, D., & Smits, A. J. (2020). Bioinspired underwater propulsion. In W. Soboyejo & L. Daniel (Eds.), Bioinspired structures and design (pp. 113–139). Cambridge University Press.
van Dam, C. P. (1987). Efficiency characteristics of crescent-shaped wings and caudal fins. Nature, 325, 435–437.
Videler, J. J. (1993). Fish swimming. Chapman & Hall.
Vogel, S. (1981). Life in moving fluids. Willard Grant Press.
Vogel, S. (1994). Life in moving fluids. Princeton University Press.
von Mises, R. (1945). Theory of flight. Dover.
Wainwright, S. A., Vosburgh, F., & Hebrank, J. H. (1978). Shark skin: Function in locomotion. Science, 202, 747–749.
Wainwright, D. K., Fish, F. E., Ingersoll, S., Williams, T. M., & St. Leger, J., Smits, A.J., Lauder, G.V. (2019). How smooth is a dolphin? The ridged skin of odontocetes. Biology Letters, 15, 20190103.
Wake, D. B. (1991). Homoplasy: The result of natural selection, or evidence of design limitations? The American Naturalist, 138(3), 543–567.
Wake, D. B., Wake, M. H., & Specht, C. D. (2011). Homoplasy: From detecting pattern to determining process and mechanism of evolution. Science, 331, 1032–1035.
Walsh, M. J. (1990). Riblets. Progress in Astronautics and Aeronautics, 123, 203–261.
Walters, V. (1962). Body form and swimming performance in scombrid fishes. American Zoologist, 2, 143–149.
Watanabe, Y. Y., Goldman, K. J., Caselle, J. E., Chapman, D. D., & Papastamatiou, Y. P. (2015). Comparative analyses of animal-tracking data reveal ecological significance of endothermy in fishes. Proceedings of the National Academy of Sciences of the United States of America, 112, 6104–6109.
Watson, A. G., & Fordyce, R. E. (1993). Skeleton of two minke whales, Balaenoptera acutorostrata, stranded on the south-east coast of New Zealand. New Zealand Natural Sciences, 20, 1–14.
Webb, P. W. (1975). Hydrodynamics and energetics of fish propulsion. Bulletin Fisheries Research Board of Canada, 190, 1–158.
Webb, P. W. (1988). Simple physical principles and vertebrate aquatic locomotion. American Zoologist, 28, 709–725.
Webb, P. W. (2004). Maneuverability-general issues. IEEE Journal of Ocean Engineering, 29, 547–555.
Webb, P. W. (2006). Stability and maneuverability. In R. E. Shadwick & G. V. Lauder (Eds.), Fish biomechanics (pp. 281–332). Academic Press.
Webb, P. W., & de Buffrénil, V. (1990). Locomotion in the biology of large aquatic vertebrates. Transactions of the American Fisheries Society, 119, 629–641.
Webb, P. W., LaLiberte, G. D., & Schrank, A. J. (1996). Does body and fin form affect the maneuverability of fish traversing vertical and horizontal slits? Environmental Biology of Fishes, 46, 7–14.
Weber, P. W., Murray, M. M., Howle, L. E., & Fish, F. E. (2009a). Comparison of real and idealized cetacean flippers. Bioinspiration & Biomimetics, 4, 046001.
Weber, P. W., Howle, L. E., Murray, M. M., & Fish, F. E. (2009b). Lift and drag performance of odontocete cetacean flippers. The Journal of Experimental Biology, 212, 2149–2158.
Weber, P. W., Howle, L. E., Murray, M. M., Reidenberg, J. S., & Fish, F. E. (2014). Hydrodynamic performance of the flippers of large-bodied cetaceans in relation to locomotor ecology. Marine Mammal Science, 30, 413–432.
Wegner, P. P. (1991). What makes airplanes fly? Springer.
Weihs, D. (1972). Semi-infinite vortex trails, and their relation to oscillating airfoils. Journal of Fluid Mechanics, 54, 679–690.
Weihs, D. (1989). Design features and mechanics of axial locomotion in fish. American Zoologist, 29, 151–160.
Weihs, D. (1993). Stability of aquatic animal locomotion. Contemporary Mathematics, 141, 443–461.
Weihs, D. (2002). Dynamics of dolphin porpoising revisited. Integrative and Comparative Biology, 42(5), 1071–1078.
Weihs, D., & Webb, P. W. (1983). Optimization of locomotion. In P. W. Webb & D. Weihs (Eds.), Fish biomechanics (pp. 339–371). Praeger.
Weng, K. C., Boustany, A. M., Pyle, P., Anderson, S. D., Brown, A., & Block, B. A. (2007). Migration and habitat of white sharks (Carcharodon carcharias) in the eastern Pacific Ocean. Marine Biology, 152(4), 877–894.
Weng, K. C., Foley, D. G., Ganong, J. E., Perle, C., Shillinger, G. L., & Block, B. A. (2008). Migration of an upper trophic level predator, the salmon shark Lamna ditropis, between distant ecoregions. Marine Ecology Progress Series, 372, 253–264.
Wicander, R., & Monroe, J. S. (2012). Historical geology: Evolution of earth and life through time. Brooks/Cole.
Wilga, C. D., & Lauder, G. V. (2000). Three-dimensional kinematics and wake structure of the pectoral fins during locomotion in Leopard sharks, Triakis semifasciata. The Journal of Experimental Biology, 203, 2261–2278.
Wilga, C. D., & Lauder, G. V. (2001). Functional-morphology of the pectoral fins in bamboo sharks, Chiloscyllium plagiosum—benthic vs. pelagic station-holding. Journal of Morphology, 249, 195–209.
Wilga, C. D., & Lauder, G. V. (2004). Hydrodynamic function of the shark’s tail. Nature, 430, 850.
Williams, T. M. (1987). Approaches for the study of exercise physiology and hydrodynamics in marine mammals. In A. C. Huntley, D. P. Costa, G. A. J. Worthy, & M. A. Castellini (Eds.), Approaches to marine mammal energetics (Vol. No. 1, pp. 127–145). Special Publication of the Society for Marine Mammalogy.
Williams, T. M. (1989). Swimming by sea otters: Adaptations for low energetic cost locomotion. Journal of Comparative Physiology. A, 164, 815–824.
Williams, T. M. (1999). The evolution of cost efficient swimming in marine mammals: Limits to energetic optimization. Philosophical Transactions of the Royal Society London B, 353, 1–9.
Williams, T. M. (2001). Intermittent swimming by mammals: A strategy for increasing energetic efficiency during diving. American Zoologist, 41, 166–176.
Williams, T. M., & Kooyman, G. L. (1985). Swimming performance and hydrodynamic characteristics of harbor seals Phoca vitulina. Physiological Zoology, 58, 576–589.
Williams, T. M., Kooyman, G. L., & Croll, D. A. (1991). The effect of submergence on heart rate and oxygen consumption of swimming seals and sea lions. Journal of Comparative Physiology. B, 160, 637–644.
Williams, T. M., Friedl, W. A., & Haun, J. E. (1993). The physiology of bottlenose dolphins (Tursiops truncatus): Heart rate, metabolic rate and plasma lactate concentration during exercise. The Journal of Experimental Biology, 179, 31–46.
Williams, T. M., Davis, R. W., Fuiman, L. A., Francis, J., Le, B. J., Horning, M., Calambokidis, J., & Croll, D. A. (2000). Sink or swim: Strategies for cost-efficient diving by marine mammals. Science, 288, 133–136.
Williams, T. M., Fuiman, L. A., Kendall, T., Berry, P., Richter, B., Noren, S. R., Thometz, N., Shattock, M. J., Farrell, E., Stamper, A. M., & Davis, R. W. (2015). Exercise at depth alters bradycardia and incidence of cardiac anomalies in deep-diving marine mammals. Nature Communications, 6(1), 1–9.
Williston, S. W. (1914). Water reptiles of the past and present. University of Chicago Press.
Withers, P. C. (1992). Comparative animal physiology. Saunders.
Wolf, N. G., Swift, P. R., & Carey, F. G. (1988). Swimming muscle helps warm the brain of lamnid sharks. Journal of Comparative Physiology. B, 157, 709–715.
Yazdi, P., Kilian, A., & Culik, B. (1999). Energy expenditure of swimming bottlenose dolphins (Tursiops truncatus). Marine Biology, 134, 601–607.
Yurchenko, N. F., & Babenko, V. V. (1980). Stabilization of the longitudinal vortices by skin integuments of dolphins. Biophysics, 25, 309–315.
Zhang, J.-D., Sung, H. J., & Huang, W.-X. (2020). Specialization of tuna: A numerical study on the function of caudal keels. Physics of Fluids, 32, 111902.
Acknowledgements
I am grateful for the assistance of Danielle Adams, William Gough, Griffin Lewis, and Kelsey Tennett with research on the structure and swimming performance of highly-derived aquatic vertebrates. I also appreciate the fruitful discussions with George Lauder, Judy Massare, Anthony Nicastro, Hans Thewissen, Paul Webb, and Terrie Williams in formulating some of the ideas presented in this chapter. Judy Massare is also gratefully acknowledged for contributing images of ichthyosaurs, and Samrithaa Balakrishnan, Anthony Russell and Alex Werth for useful comments on the manuscript. The work was supported in part with a grant from the Office of Naval Research (N000141410533).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2023 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this chapter

Cite this chapter
Fish, F.E. (2023). Aquatic Locomotion: Environmental Constraints That Drive Convergent Evolution. In: Bels, V.L., Russell, A.P. (eds) Convergent Evolution. Fascinating Life Sciences. Springer, Cham. https://doi.org/10.1007/978-3-031-11441-0_15
Download citation
DOI: https://doi.org/10.1007/978-3-031-11441-0_15
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-11440-3
Online ISBN: 978-3-031-11441-0
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)