
Abstract
Sierra Nevada faces the stark challenge of preserving and restoring biodiversity and the provision of ecosystem services in the face of ongoing shifts in natural disturbance regimes. As wildfires become more frequent, severe and widespread under human land-use changes and climate change, there is a need to understand the mechanisms that promote ecological resilience after the fire and the effects of management on such mechanisms. After the 2005 Lanjarón fire in Sierra Nevada, we established an experiment in a burnt pine stand to assess how the ecosystem responded to three management schemes: (a) salvage logging, a common strategy consisting in the felling and extraction of burnt trees; (b) partial cut, where 90% of trees were felled and cut in pieces but left scattered on the ground; and (c) non-intervention, where no action was taken. For 15 years, we monitored how different components of biodiversity and ecosystem processes and services responded to the post-fire treatments, and assessed how management affected the mechanisms that drive natural regeneration. In this chapter, we describe the Lanjarón experiment and its key scientific contributions. We first describe the insights related to the role of dead wood in promoting regeneration. This includes processes such as seed dispersal and predation, fertilization through the gradual decomposition of dead wood, microclimatic amelioration, and herbivory. Second, we portray how the community of birds, vascular plants and soil insects responded to the post-fire treatments. Whereas diversity itself was affected by management, the key responses were related to community composition. And third, we address the functions of the ecosystem related to its capacity to provide benefits to human society. We mainly address regulating ecosystem services but also analyze some provisioning services, including their economic value. In each of these three sections, we end by providing a broader, global view on the effects of salvage logging as derived from reviews that have subsequently been made under international collaborations. Altogether, the Lanjarón experiment in Sierra Nevada constitutes a unique research infrastructure that has broadened our understanding of the role of dead wood in promoting ecological resilience and whose findings have contributed to the integration of knowledge about post-fire dynamics in an international context. Additionally, ongoing monitoring aims to fill the research gap of addressing the long-term effects of a critical post-disturbance management strategy in a world facing novel disturbance regimes.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
Research in Sierra Nevada has helped reveal global challenges for ecosystems resulting from novel climatic conditions (as explained in previous chapters). Shifting temperature and rainfall regimes are producing an unprecedented need for local species to migrate or adapt (Zamora et al. 2015). Climate change also has consequences for environmental management programs, which must increasingly adapt to new conditions for instance by selecting species adapted to projected future conditions for revegetation (Hoegh-Guldberg et al. 2008; Leverkus et al. 2015). But while climate change alone is already expected to produce profound changes in biotic communities worldwide (Trisos et al. 2020), the coupled alterations in the frequency, severity, extent, and seasonality of disturbances such as wildfires and pests (Seidl et al. 2017) demand further measures for adaptation (Leverkus et al. 2021c). To succeed in managing ecosystems under changing climate and disturbance regimes, we need a better understanding of the mechanisms that favor the recovery of ecosystem functioning and biodiversity after disturbances.
Ecosystems are characterized by the occurrence of natural disturbances of particular types, frequencies, and severities—also termed disturbance regimes. In Mediterranean ecosystems, the disturbance regime is generally characterized by wildfires of moderate frequency and severity (Pausas et al. 2008). Many Mediterranean species have evolved adaptations that allow their regeneration after fire, such as the ability to survive under thick protecting bark, to resprout from surviving underground tissues, or to germinate following stimulation from the heat (Pausas and Keeley 2014). However, the capacity of ecosystems to recover after wildfire (which we here term resilience; Nimmo et al. 2015) may be compromised under novel disturbance conditions. Resilience is promoted by the legacies from the affected ecosystem that remain after the fire (Johnstone et al. 2016). These legacies include seed banks, patches with surviving vegetation and animals, resprouting plants, and soil fungi (Swanson et al. 2011; Johnstone et al. 2016). Other organic structures and biologically generated spatial patterns also constitute legacies that set the baseline for regeneration, including remaining organic soil, animal burrows, and—in the case of forests—large amounts of deadwood. However, this deadwood is generally removed from disturbed ecosystems through silvicultural management (Lindenmayer et al. 2008; Müller et al. 2019). The question thus arises whether such additional alteration of the disturbed ecosystem through management that removes and modifies disturbance legacies may compromise ecological resilience.
The practice of removing dead trees after disturbances is named salvage logging. Worldwide, it is conducted primarily for economic motivations (recovering some of the affected forest capital or paying for restoration activities; Müller et al. 2019; Castro 2021). Other frequent justifications include the perception that it helps restore the ecosystem, reduce fuel loads, minimize safety hazards from falling trees, mitigate pest outbreaks, and remove the ugliness of the deadwood (Müller et al. 2019; Castro 2021). Such motivations often lead to unusually large areas being subjected to tree harvesting after large disturbances (Radeloff et al. 2000; Gustafsson et al. 2019) and even protected areas being logged (as happened after the Lanjarón fire here described but also elsewhere, e.g. Schiermeier 2016; Leverkus et al. 2017. Until the early 2000s, there was scant scientific knowledge about the ecological implications of this already common practice (McIver and Starr 2000). A subsequent call for a thorough evaluation of the ecological impacts of salvage logging (Lindenmayer et al. 2004) was followed by intense controversy around the topic (e.g. Karr et al. 2004; DellaSala et al. 2006; Donato et al. 2006; Lindenmayer et al. 2017) and instigated an array of research projects worldwide. Recent advances have highlighted the potential for unforeseen, interacting effects among the natural disturbance and the subsequent disturbance by logging (Buma and Wessman 2011; Leverkus et al. 2018a; Lindenmayer and Sato 2018), and subsequent reviews have aimed to provide broader views on the topic (e.g. Royo et al. 2016; Leverkus et al. 2018b; Thorn et al. 2018).
Here, we explain how the post-fire experiment after the 2005 Lanjarón fire, which burned more than 3000 ha of pine stands and shrublands in Sierra Nevada, has contributed to our understanding of the role of deadwood in promoting ecosystem resilience and of the ecological impacts of salvage logging. The experimental setup, with three post-fire management treatments replicated within and across four blocks at different elevations, constitutes one of the most statistically sound experimental designs on post-fire management (Leverkus et al. 2018b) and has produced a great amount of scientific outputs. These include 3 Ph.D. theses, more than 20 papers in scientific journals, several outreach publications and management recommendations, and strong international networks. We also explain how the data and insights from this experiment have subsequently been combined with those from other experiments to produce broader insights into the drivers of the effects of salvage logging on biodiversity and ecosystem services.
2 The Lanjarón Post-fire Experiment
From the 22nd to the 24th of September 2005, the Lanjarón fire burned 3425 ha of shrublands, grasslands and pine afforestations in the Sierra Nevada Natural and National Park. The burnt pines were 35–45 years old at the time of the fire. The climate in the area is Mediterranean, with hot, dry summers and wet, mild winters (Table 1). The area surrounding the burnt site was dominated by shrublands.
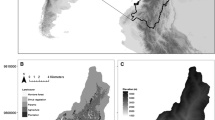
The local forest service planned the removal of the burnt wood across all the burnt area, and in cooperation with this administration, we implemented an experimental design to study the effect of post-fire salvage logging in pine forests on the ecosystem. The experiment was implemented in four blocks of approximately 25 ha each, located across an elevational gradient (Fig. 1, Table 1). The pine species in each block differed according to their ecological requirements along this elevation and moisture gradient. The cluster pine (Pinus pinaster) and black pine (P. nigra) dominated in Block 1, black pine in Block 2, and Scots pine (P. sylvestris) in Blocks 3 and 4. All three species are native in the region, although they were extensively planted in the area.

Location and set-up of the Lanjarón experiment. Top-left, location of the Lanjarón fire in the Iberian Peninsula; top-right, ortho-rectified photo showing the three experimental blocks at different elevations; middle, experimental design with the allocation of post-fire treatments to three plots per block; bottom, diagram of the experimental treatments. In the top-right map, the blue polygon above Block 1 surrounds a population of unburnt holm oaks from which seed dispersal was monitored. PC = Partial Cut, SL = Salvage Logging, NI = Non-Intervention, NI* = Non-Intervention treatment after 6 years, by when all the standing dead trees had naturally collapsed; P = patches of surviving pines in Block 1 in which demographic dynamics of oaks after dispersal were monitored. Block 4, which had one replicate per treatment and was located at the greatest elevation, is not represented here
Within each block, we implemented three replicates (plots) of the following burnt-wood management procedures (treatments hereafter; Fig. 2) in a random spatial distribution: (1) nonintervention (NI), where all burnt trees were left standing; (2) partial cut plus lopping (PC), where 90% of burnt trees were felled and their main branches lopped off, but all the cut biomass was left on the ground; and (3) experimental salvage logging (SL), where trees were cut and the trunks cleaned of branches with chainsaws, trunks were manually piled (groups of 10–15) and the woody debris was chopped with a mechanical chopper. In the salvage logging plots, we planned to extract the trunks with a log forwarder, but this step was canceled by the forest service because of the difficulties of operating machinery within the spatial arrangement of the experiment. Given that the salvage logging treatment could not be carried out with the same intensity as in the area surrounding the experimental blocks, for some of the studies we also included three replicates per block (of about 3 ha each) in the extraction matrix surrounding each of the experimental blocks (matrix salvage logging, MSL; Castro et al. 2010). The treatments were established between 21 April and 28 June 2006. The four blocks were located on siliceous soils, coming from mica schists of the Nevado-Filábride complex (see Table 1 for details, and chapter “Geological Setting of Sierra Nevada”). Within this experimental setup, we measured the effect of post-fire management on different aspects of the regeneration and functioning of the ecosystem, including: woody species regeneration, plant-animal interactions (seed dispersal, seed predation and herbivory), diversity of plants and birds, wood decomposition and its effect on soil properties, soil respiration, carbon exchange with the atmosphere, associated ecosystem services, and the economic and restoration implications of post-fire logging.

Photo of post-fire treatments shortly after their implementation in early 2006. Foreground: Partial Cut treatment, where 90% of burnt trees were felled, cut in pieces, and spread on the ground. Middle: Salvage Logging, where all trees were felled, branches were masticated, and the trunks were piled. Back: Non-Intervention treatment, where burnt trees were left standing
The Lanjarón experiment was among the first where the effect of post-fire salvage logging was assessed in a true experimental design that included random allocation of treatments to spatial units and a size large enough to study processes at ecosystem scale (see chapter “Managing the Uniqueness of Sierra Nevada Ecosystems Under Global Change: The Value of in situ Scientific Research”). The absence of experimental studies has in fact been a major constraint to obtaining robust conclusions on the impacts of post-fire salvage logging, so the design established in the Sierra Nevada was a milestone in this regard.
3 The Role of Burnt Wood as a Disturbance Legacy: Mechanisms that Promote Resilience
From an ecological standpoint, extracting burnt wood implies the removal of large amounts of disturbance legacies—i.e. the organisms, organic materials and biologically derived patterns that persist after disturbance and set the baseline for regeneration (Franklin et al. 2000; Johnstone et al. 2016). As a first logical consideration, extracting burnt wood implies the removal of an important pool of nutrients accumulated over the lifetime of trees. At the Lanjarón site, we found that the macro- and micronutrient content in burnt pine trunks was similar to that in live specimens (Marañón-Jiménez et al. 2013a). Leaves and fine twigs are usually consumed during wildfires and their nutrients are released to the atmosphere or to the ground as ashes. However, coarse branches and trunks are isolated from the heat, and a considerable pool of nutrients may remain in the wood. These nutrients are protected from sudden losses from runoff or wind after the fire and are gradually released into the soil during decomposition, thereby enhancing soil fertility during succession. For instance, the nutrient pool remaining in the burnt wood four years after the fire was 2–9 times higher than the pool in the first 10 cm of the soil for elements such as Na, Mn, Fe, Zn or Cu (Marañón-Jiménez and Castro 2013), and microbiological parameters such as dissolved organic carbon, dissolved organic nitrogen, microbial biomass and microbial N and P content also benefited from the presence of deadwood. At a greater time scale, the content of K and P—which are particularly limiting at the study site—were still higher where the burnt wood was left in situ 10.5 years after the fire (Juan-Ovejero et al. 2021).
Higher soil nutrient pools were transferred to pine (Pinus pinaster Ait.) saplings regenerating from seed (Fig. 3a). These grew more in the treatments with abundant decomposing wood while keeping a similar nutrient concentration as in salvaged areas (Marañón-Jiménez et al. 2013b), suggesting that a higher nutrient availability led to a higher total nutrient pool in the plants. The reproductive status of the pines after six years was also improved by the presence of deadwood, and the analysis of stable isotopes pointed to greater water-use efficiency in pines growing in the presence of burnt wood due to enhanced nutritional status (Marañón-Jiménez et al. 2013b). Long-lasting effects reported in recent studies (Bowd et al. 2019) reinforce the role of burnt wood as a biological legacy that improves soil fertility.

a Pinus pinaster seedlings regenerating under the shelter of fallen burnt trees; b naturally regenerating plant community in the non-intervention treatment; contrast this with c plant community in the salvage logging treatment; d extraction of soil cores for macroarthropod sampling
The burnt wood may also act as a nurse object. Mediterranean ecosystems are characterized by an abundance of facilitative interactions among plants (Gómez-Aparicio et al. 2004; Siles et al. 2010), which are driven by the mitigation of abiotic stress as seedlings grow sheltered under other plants, especially shrubs. Facilitation in Mediterranean-type ecosystems arises primarily from the reduction of high radiation intensity by the canopy of the nurse plant, which improves water status in the beneficiary plants by reducing temperature and wind speed and increasing soil moisture and air–water vapour content. However, these positive aspects are at least partly counterbalanced by the impact of competition for resources between the nurse plant and the facilitated seedling (Maestre et al. 2003; Gómez-Aparicio et al. 2005; Rey Benayas et al. 2005). The burnt wood can act as a nurse object in the sense that it reduces water stress but without producing competition (Castro et al. 2011; Ginzburg and Steinberger 2012; Marcolin et al. 2019; Fig. 2b). This constitutes a mechanism through which burnt wood may improve post-fire regeneration (Castro et al. 2011; Marzano et al. 2013; Taboada et al. 2018; Urretavizcaya and Defosse 2019). Moreover, as mentioned above, the wood provides nutrients during decomposition and it can retain moisture, organic matter, and fine sediments by acting as a barrier against runoff. Burnt trees also reduced wind speed at the study site (Serrano-Ortiz et al. 2011), and acted as a physical barrier against herbivores (Castro 2013). Thus, the burnt wood acted as a nurse object that provided better conditions for two critical factors that limit the regeneration of woody species in Mediterranean-type environments: summer drought and herbivore pressure.
The presence of burnt wood can also favour mutualistic interactions for the natural regeneration of the forest after fire. Many frugivorous birds use standing burnt trees, felled trees or piles of branches as perches (e.g. Castro et al. 2010; Rost et al. 2010). This may increase the entry of tree and shrub seeds of intermediate stages of succession—many of them producing fleshy fruits—with respect to areas where burnt wood is removed (e.g. Rost et al. 2010; Cavallero et al. 2013). A relevant case of mutualism for post-fire regeneration is the dispersal of acorns by the Eurasian jay (Garrulus glandarius). Jays disperse acorns in temperate forests of the Holarctic, as they scatterer-hoard seeds across the landscape for later consumption. A fraction of the acorns remains unrecovered, providing the opportunity for seed germination and tree recruitment (Pesendorfer et al. 2016). The Eurasian jay dwells in relatively dense forests and avoids open landscapes; pine forests, including relatively dense plantations, are among the habitats most used by jays to hide acorns (Mosandl and Kleinert 1998; Gómez 2003). In the Lanjarón area we documented that jays still used the burnt pine forest to cache acorns as long as the trees remained standing (Castro et al. 2012). Thus, standing dead trees constituted a crucial structural element for the colonisation of the burnt area by oaks (Leverkus et al. 2016), which in turn represents an ecosystem service with quantifiable economic value (Leverkus and Castro 2017).
The plethora of mechanisms through which dead wood may improve regeneration suggest that a major ecological impact of salvage logging may be its potential to affect forest regeneration and thereby compromise resilience (Van Nieuwstadt et al. 2001; Noss and Lindenmayer 2006; Donato et al. 2006). To assess this, we conducted a meta-analysis on regeneration data from Lanjarón and other European and North American studies after wildfires, windthrows, and insect outbreaks. This study found that there is no generalized response of regeneration to salvage logging across all tree species, and that initial impacts of salvage logging on tree regeneration density tend to decrease over time (Leverkus et al. 2021b). However, the lack of overall impacts was accompanied by the finding that more related tree species tend to respond more similarly to salvage logging than more distantly related species, and that the measured effects are quite specific to study sites (Leverkus et al. 2021b). This highlights the importance of understanding the mechanisms through which salvage logging may influence tree natural regeneration under local set of circumstances and species, adding value to detailed studies such as those described in this section for the development of local mitigation strategies.
4 Biodiversity Response to Post-fire Management
Salvage logging can affect biodiversity through a range of mechanisms. These include the direct destruction of organisms during working operations, the removal of larvae and propagules in the dead trunks, the elimination of substrate and food in the form of dead wood, the change in structural properties of the habitat, and a reduction in the variety of niches (Lindenmayer et al. 2008; Thorn et al. 2015, 2020b). Still, it can be expected that some species also benefit from an open habitat structure (e.g. Leverkus et al. 2013; Hagge et al. 2019), so the overall impact on biodiversity ultimately results from the balance between winners and losers.
In Lanjarón, biodiversity studies have addressed plant, bird, and soil macroarthropod communities. Vascular plants, sampled two years after the fire, showed a reduction in species richness, Shannon diversity and ground cover after salvage logging (Fig. 3b, c; Leverkus et al. 2014). But the composition of communities also changed. The abundance of species that regenerate through seeds was most negatively affected, likely because by the time salvage logging was conducted those species were already emerging; trampling by people and machinery therefore depleted the seedling bank and compromised regeneration. Tree and shrub species were also negatively affected, which could suggest a slowing down of succession (Leverkus et al. 2014). Re-sampling of vegetation is planned ~17 years after the fire to assess whether these impacts are long-lasting.
Bird communities (Castro et al. 2010), sampled in two years after the fire, showed a higher diversity and abundance in the treatments where the wood was left than in the salvage logging treatment. Moreover, the composition of the community changed depending on the treatment. Species that typically inhabit forests were more abundant in the non-intervention treatment, species from open landscapes and shrublands were most abundant in the salvage-logging treatment, and a mixed species composition was observed in the treatment where 90% of trees were cut and felled (Castro et al. 2010). Of particular relevance was that the non-intervention treatment had the highest abundance of species that disperse mid- and late-successional shrubs and trees that produce fleshy fruits, such as thrushes and Eurasian jays.
Finally, soils were sampled 10 years after the fire to assess the effect of remaining wood on the diversity and composition of soil macroarthropods (Fig. 3d). The salvage logging treatment resulted in an overall reduction in the abundance and richness of macroarthropod communities compared to the partial-cut treatment, in which decomposing dead trunks were still abundant (Molinas-González et al. 2019). This suggests that the legacy of dead wood also affects this hidden—albeit conspicuous—community in the soil, and potentially the ecological functions that it performs.
Besides the Lanjarón biodiversity data serving as an individual case study, international collaborations have allowed learning from this and other experiments worldwide about the extent to which the biodiversity responses to salvage logging are generalized, stochastic, or dependent on factors such as taxonomic group and local conditions. A meta-analysis involving 134 original species abundance datasets found that salvage logging after natural disturbances (including wildfire, insect outbreaks and storms) decreased the species richness of 8 out of 24 taxonomic groups that were assessed, and that significant alterations in community composition occurred in 40% of the groups (Thorn et al. 2018). This impact was mostly observed on saproxylic species; on the contrary, species groups typically associated with open habitats profited from salvage logging (Thorn et al. 2018). More detailed analysis on bird species assemblages found that dissimilarities in the occurrence of species between salvaged and unsalvaged areas are strongest, and most persistent, for rare species (Georgiev et al. 2020). This suggests that common and dominant species are comparatively less affected by salvage logging, and that assessments focusing only on these groups are likely to underestimate impacts on species of high conservation value. Finally, the global biodiversity dataset was used to ask what proportion of an area affected by a natural disturbance needs to be protected from salvage logging to preserve a certain proportion of the biodiversity specific to disturbed yet unsalvaged forest (Thorn et al. 2020a). Newly developed statistical tools helped estimate that 75% of disturbed areas are required to maintain 90% of their unique species assemblages, whereas retaining 50% of a disturbed area would protect 73% of the species unique to disturbed forests. The analyses allow estimating the proportion of disturbed areas that needs to remain unlogged to conserve different proportions of particular taxonomic groups, according to the needs of local managers (Thorn et al. 2020a).
5 Effect of Post-fire Management on Ecosystem Services
Ecosystems produce benefits to society, including the provision of materials, the regulation of biogeochemical cycles, and scientific and spiritual values—i.e. ecosystem services (MA 2005). Natural disturbances disrupt the functioning of ecosystems, including a temporary reduction in primary production, a change from subsurface water flow to surface runoff, and an alteration of aesthetic values, among others. Contrary to overall increases in biodiversity, disturbances may thus result in a reduction in the provision of ecosystem services (Thom and Seidl 2016). In fact, a global assessment—which included the case of Sierra Nevada—revealed that the reasons that motivate salvage logging in the first place include mitigating the expected reduction in key ecosystem services such as fire and pest control, timber production, and scenic value (Müller et al. 2019). However, key ecosystem services are not always reduced by natural disturbances (Beudert et al. 2015), and the capacity of post-disturbance management to mitigate potential ecosystem service losses is unclear (e.g. Donato et al. 2006). As explained in this section, the Lanjarón experiment, along with subsequent international reviews, has provided insights into how several classes of ecosystem services (Haines-Young and Potschin 2018) may be affected by salvage logging.
Provisioning ecosystem services are at the core of the salvage logging question. This is greatly because wildfires and other disturbances produce a sudden shift from a forest gradually increasing its economic value (as trees increase their volume through growth) to the value rapidly decreasing (as the remaining wood decays after tree death). Although disturbance-affected wood is usually of lesser quality and value, and wood prices commonly drop after large disturbances due to pulses in offer that saturate local markets (Peter and Bogdanski 2010), the economic incentive for salvaging the deadwood is still usually high—hence the term salvage logging (Lindenmayer et al. 2008). Another economic incentive involves the cost of subsequent restoration activities such as revegetation, which are assumed to be easier, and hence less expensive, to conduct. However, an assessment after the Lanjarón fire revealed that the high management costs associated with salvage logging operations were neither compensated by the benefits from selling the wood nor by the reduction in the cost of revegetation (Leverkus et al. 2012). A subsequent assessment showed that salvage logging produced additional losses through the reduction of natural regeneration, which increased the potential need for reforestation (Leverkus and Castro 2017). Of course, several stakeholders such as forestry operators do obtain economic gains from salvage logging, yet the economic assessment in Lanjarón challenged the overall view that salvage logging is generally profitable, particularly in unproductive Mediterranean forests or if considering the value of non-wood ecosystem services such as natural regeneration. This situation may change with ongoing developments such as the growing market for biomass pellets, which can make salvaged wood more profitable (Pons and Rost 2016). But also new forest growth models in more productive forests support that refraining from salvage logging does not necessarily produce the feared detrimental economic impacts (Knoke et al. 2021).
Many of the undesired impacts of salvage logging fall in the realm of regulating ecosystem services. As indicated in Sect. 4 above, a major effect of extracting deadwood is the elimination of food, substrate, and habitat for a plethora of deadwood-dependent species (Thorn et al. 2020b), which fall in the realm of lifecycle maintenance services (Haines-Young and Potschin 2018). But also the structural attributes of salvaged and non-salvaged ecosystems define habitat suitability; the Lanjarón experiment has demonstrated opposing outcomes of such changes for different guilds of mammals (Puerta-Piñero et al. 2010; Leverkus et al. 2013). Effects on habitat provision can carry over to other ecosystem services produced by animals such as seed dispersal by birds (Castro et al. 2012; Leverkus et al. 2016), and to disservices such as seed predation (Puerta-Piñero et al. 2010; Leverkus et al. 2013). Additionally, services of soil quality and formation are improved by dead wood, as described above (Marañón-Jiménez and Castro 2013). Ultimately, the effects of salvage logging on vegetation, microclimate and soil functioning may influence the ecosystem service of climate regulation at larger spatial scales. The measurement of soil respiration and carbon fluxes with eddy-covariance towers (Marañón-Jiménez et al. 2011; Serrano-Ortiz et al. 2011) helped reveal that the areas subjected to salvage logging acted as net carbon sources, whereas areas devoid of salvage logging acted as carbon sinks. The mechanisms underlying this counterintuitive result are likely related to the increase in primary production in nonintervention areas for the reasons described in Sect. 3, which overcompensated the larger carbon emissions from deadwood decomposition. These results, combined with those from several other studies in a global meta-analysis (Leverkus et al. 2020), showed that salvage logging produces a negative overall impact of moderate magnitude on regulating ecosystem services. Further, the meta-analysis produced the key insight that waiting before logging after natural disturbances can mitigate this negative effect—a conclusion that is unfortunately difficult to reconcile with the desire to quickly recover the deadwood (Leverkus et al. 2020).
One key concern about disturbed forests is the risk of further disturbances that may produce additional impacts on either the ecosystem or human populations and infrastructures. This is because natural disturbances can modify the likelihood of occurrence, extent and severity of further disturbances, thereby producing what is termed a linked disturbance (Buma 2015). As salvage logging produces additional alterations to the disturbed ecosystem, it can both mitigate some additional disturbances and trigger others (Leverkus et al. 2018a). In Lanjarón, an evident example of the latter is drought stress. The removal of the deadwood, which otherwise produced shade and retained moisture, increased the physiological stress suffered by plant seedlings, as measured by the ratios of stable isotopes in leaves (Marañón-Jiménez et al. 2013b). Also browsing disturbance was greater in salvaged areas, as these were devoid of the scattered trunks and branches that otherwise impeded access by large herbivores (Puerta-Piñero et al. 2010; Castro 2013). On the contrary, salvage logging removed the hazard from falling dead trees. However, we point out that the collapse of dead trees occurred mostly during stormy events and along a relatively narrow period of time (Molinas-González et al. 2017), so that the risk related to falling trees was generally small except for those events. In terms of larger-scale disturbances, salvage logging frequently aims to reduce ecosystem fuels as well as breeding grounds for pest insects (Müller et al. 2019). However, an additional meta-analysis combining data from multiple studies around the world revealed that the effectiveness of salvage logging in reducing fuel loads depends on how particular types of fuels respond to salvage logging over time (Leverkus et al. 2020). Salvage logging was found to be most effective in reducing the amount of large downed logs at intermediate to long timeframes, while it produced strong increases in small—and thus more flammable—fuels in the first years after logging (Leverkus et al. 2020). A broader international review highlighted that predicting the effects of post-disturbance management on subsequent disturbance risk requires evaluating the effects of management on multiple, not always evident ecosystem properties. For instance, predicting the temporal trajectories of fuel loads also requires understanding the response of vegetation to management (Leverkus et al. 2021a). The review highlights that managing disturbed ecosystems to avoid additional disturbances already implies imposing an additional disturbance (namely logging) and may also result in other, unforeseen disturbances (such as hydrologic disturbances and mass flows); it proposed a decision-making framework to avoid hasty decision-making after natural disturbances for the sake of reducing hazards (Leverkus et al. 2021a).
6 Conclusions
The Lanjarón experiment has played a fundamental role in understanding the implications of post-fire management for ecosystem resilience. Extracting the deadwood from a recently disturbed ecosystem produces an array of impacts that ultimately affect the capacity for natural regeneration, biodiversity, and the ecosystem services produced by burnt forests. As found in subsequent global reviews, some of these effects are generalized, yet others require detailed assessment of local species and conditions to fully understand—and properly manage—the drivers of management effects. New studies ought to transcend the plot scale to address the landscape-scale implications of different types of management, and integrate that information to obtain optimal landscape configurations that maximize the delivery of different ecosystem service and biodiversity elements. Also, long-term monitoring is needed to assess successional trajectories across succession under multiple combinations of natural and anthropogenic disturbances. The broad conclusion of the Lanjarón experiment is that post-disturbance management strategies that avoid superimposing additional disturbances may help promote resilience and thereby reduce the risk of ecosystem collapse in a world facing shifting disturbance regimes.

Author: Luis Ordoñez
References
Beudert B, Bässler C, Thorn S et al (2015) Bark beetles increase biodiversity while maintaining drinking water quality. Conserv Lett 8:272–281
Bowd EJ, Banks SC, Strong CL, Lindenmayer DB (2019) Long-term impacts of wildfire and logging on forest soils. Nat Geosci 12:113–118
Buma B (2015) Disturbance interactions: characterization, prediction, and the potential for cascading effects. Ecosphere 6:Art70
Buma B, Wessman CA (2011) Disturbance interactions can impact resilience mechanisms of forests. Ecosphere 2:art64
Castro J (2021) Post-fire restoration of Mediterranean pine forests. In: Ne’eman G OY (ed) Pines and their mixed forest ecosystems in the Mediterranean Basin. Springer, p In press
Castro J (2013) Postfire burnt-wood management affects plant damage by ungulate herbivores. Int J for Res 2013:965461
Castro J, Allen CD, Molina-Morales M et al (2011) Salvage logging versus the use of burnt wood as a nurse object to promote post-fire tree seedling establishment. Restor Ecol 19:537–544
Castro J, Moreno-Rueda G, Hódar JA (2010) Experimental test of postfire management in pine forests: Impact of salvage logging versus partial cutting and nonintervention on bird-species assemblages. Conserv Biol 24:810–819
Castro J, Puerta-Piñero C, Leverkus AB et al (2012) Post-fire salvage logging alters a key plant-animal interaction for forest regeneration. Ecosphere 3:art90. 1890/ES12-00089.1
Cavallero L, Raffaele E, Aizen MA (2013) Birds as mediators of passive restoration during early post-fire recovery. Biol Conserv 158:342–350
DellaSala DA, Karr JR, Schoennagel T et al (2006) Post-fire logging debate ignores many issues. Science (80-) 314:51–52
Donato DC, Fontaine JB, Campbell JL et al (2006) Post-wildfire logging hinders regeneration and increases fire risk. Science (80-) 311:352
Franklin JF, Lindenmayer D, MacMahon JA et al (2000) Threads of continuity. Conserv Pract 1:8–17
Georgiev KB, Chao A, Castro J et al (2020) Salvage logging changes the taxonomic, phylogenetic and functional successional trajectories of forest bird communities. J Appl Ecol 57:1103–1112
Ginzburg O, Steinberger Y (2012) Salvage logging versus natural regeneration post-fire practices in a forest: soil chemical and microbial aspects. Open J Ecol 2:29–37
Gómez-Aparicio L, Gómez JM, Zamora R, Boettinger JL (2005) Canopy vs soil effects of shrubs facilitating tree seedlings in Mediterranean montane ecosystems. J Veg Sci 16:191–198
Gómez-Aparicio L, Zamora R, Gómez JM et al (2004) Applying plant facilitation to forest restoration: a meta-analysis of the use of shrubs as nurse plants. Ecol Appl 14:1128–1138
Gómez JM (2003) Spatial patterns in long-distance dispersal of Quercus ilex acorns by jays in a hetereneous landscape. Ecography (Cop) 5:573–584
Gustafsson L, Berglind M, Granström A et al (2019) Rapid ecological response and intensified knowledge accumulation following a north European mega-fire. Scand J for Res 34:234–253
Hagge J, Müller J, Bässler C et al (2019) Deadwood retention in forests lowers short-term browsing pressure on silver fir saplings by overabundant deer. For Ecol Manage 451:117531
Haines-Young R, Potschin MB (2018) Common international classification of ecosystem services (CICES) V5.1 and guidance on the application of the revised structure. www.cices.eu
Hoegh-Guldberg O, Hughes L, McIntyre S et al (2008) Assisted colonization and rapid climate change. Science (80-) 321:345–346
Johnstone JF, Allen CD, Franklin JF et al (2016) Changing disturbance regimes, ecological memory, and forest resilience. Front Ecol Environ 14:369–378
Juan-Ovejero R, Molinas-González CR, Leverkus AB et al (2021) Decadal effect of salvage logging on the soil nutrient stocks of a wildfire-affected Mediterranean forest. For Ecol Manage 498:119570
Karr JR, Rhodes JJ, Minshall GW et al (2004) The effects of postfire salvage logging on aquatic ecosystems in the American West. Bioscience 54:1029
Knoke T, Gosling E, Thom D et al (2021) Economic losses from natural disturbances in Norway spruce forests—a quantification using Monte-Carlo simulations. Ecol Econ 185:107046
Leverkus AB, Buma B, Wagenbrenner J et al (2021a) Tamm review: does salvage logging mitigate subsequent forest disturbances? For Ecol Manage 481:118721
Leverkus AB, Gustafsson L, Lindenmayer DB et al (2020) Salvage logging effects on regulating ecosystem services and fuel loads. Front Ecol Environ 18:391–400
Leverkus AB, Lindenmayer DB, Thorn S, Gustafsson L (2018a) Salvage logging in the world’s forests: interactions between natural disturbance and logging need recognition. Glob Ecol Biogeogr 27:1140–1154
Leverkus AB, Polo I, Baudoux C et al (2021b) Resilience impacts of a secondary disturbance: meta-analysis of salvage logging effects on tree regeneration. J Ecol in Press
Leverkus AB, Rey Benayas JM, Castro J et al (2016) Shifting demographic conflicts across recruitment cohorts in a dynamic post-disturbance landscape. Ecology 97:2628–2639
Leverkus AB, Rey Benayas JM, Castro J et al (2018b) Salvage logging effects on regulating and supporting ecosystem services—a systematic map. Can J for Res 48:983–1000
Leverkus AB, Thorn S, Gustafsson L et al (2021c) Environmental policies to cope with novel disturbance regimes—steps to address a world scientists’ warning to humanity. Environ Res Lett 16:021003
Leverkus AB, Castro J (2017) An ecosystem services approach to the ecological effects of salvage logging: valuation of seed dispersal. Ecol Appl 27:1057–1063
Leverkus AB, Jaramillo-López PF, Brower LP et al (2017) Mexico’s logging threatens butterflies. Science 358:1008
Leverkus AB, Lorite J, Navarro FB et al (2014) Post-fire salvage logging alters species composition and reduces cover, richness, and diversity in Mediterranean plant communities. J Environ Manage 133:323–331
Leverkus AB, Puerta-Piñero C, Guzmán-Álvarez JR et al (2012) Post-fire salvage logging increases restoration costs in a Mediterranean mountain ecosystem. New for 43:601–613
Leverkus AB, Castro J, Delgado-Capel MJ et al (2015) Restoring for the present or restoring for the future: enhanced performance of two sympatric oaks (Quercus ilex and Quercus pyrenaica) above the current forest limit. Restor Ecol 23:936–946
Leverkus AB, Castro J, Puerta-Piñero C, Rey Benayas JM (2013) Suitability of the management of habitat complexity, acorn burial depth, and a chemical repellent for post-fire reforestation of oaks. Ecol Eng 53:15–22
Lindenmayer D, Thorn S, Banks S (2017) Please do not disturb ecosystems further. Nat Ecol Evol 1:art31
Lindenmayer DB, Burton PJ, Franklin JF (2008) Salvage logging and its ecological consequences. Island Press, Washington, D.C.
Lindenmayer DB, Foster DR, Franklin JF et al (2004) Salvage harvesting policies after natural disturbance. Science 303:1303
Lindenmayer DB, Sato C (2018) Hidden collapse is driven by fire and logging in a socioecological forest ecosystem. Proc Natl Acad Sci 115:5181–5186
MA [Millennium Ecosystem Assessment] (2005) Ecosystems and human well-being: synthesis. Island Press, Washington, DC
Maestre FT, Bautista S, Cortina J (2003) Positive, negative, and net effects in grass-shrub interactions in Mediterranean semiarid grasslands. Ecology 84:3186–3197
Marañón-Jiménez S, Castro J (2013) Effect of decomposing post-fire coarse woody debris on soil fertility and nutrient availability in a Mediterranean ecosystem. Biogeochemistry 112:519–535
Marañón-Jiménez S, Castro J, Fernández-Ondoño E, Zamora R (2013a) Charred wood remaining after a wildfire as a reservoir of macro- and micronutrients in a Mediterranean pine forest. Int J Wildl Fire 22:681–695
Marañón-Jiménez S, Castro J, Kowalski AS et al (2011) Post-fire soil respiration in relation to burnt wood management in a Mediterranean mountain ecosystem. For Ecol Manage 261:1436–1447
Marañón-Jiménez S, Castro J, Querejeta JI et al (2013b) Post-fire wood management alters water stress, growth, and performance of pine regeneration in a Mediterranean ecosystem. For Ecol Manage 308:231–239
Marcolin E, Marzano R, Vitali A et al (2019) Post-fire management impact on natural forest regeneration through altered microsite conditions. Forests 10:1014
Marzano R, Garbarino M, Marcolin E et al (2013) Deadwood anisotropic facilitation on seedling establishment after a stand-replacing wildfire in Aosta Valley (NW Italy). Ecol Eng 51:117–122
McIver JD, Starr L (2000) Environmental effects of postfire logging: literature review and annotated bibliography. USDA For Serv Gen Tech Rpt PNW-GTR 72
Molinas-González CR, Castro JJ, González-Megías A, Leverkus AB (2019) Effects of post-fire deadwood management on soil macroarthropod communities. Forests 10:art1046
Molinas-González CR, Leverkus AB, Marañón-Jiménez S, Castro J (2017) Fall rate of burnt pines across an elevational gradient in a Mediterranean mountain. Eur J For Res 136
Mosandl R, Kleinert A (1998) Development of oaks (Quercus petraea (Matt.) Liebl.) emerged from bird-dispersed seeds under old-growth pine (Pinus silvestris L.) stands. For Ecol Manage 106:35–44
Müller J, Noss RF, Thorn S et al (2019) Increasing disturbance demands new policies to conserve intact forest. Conserv Lett 12:e12449
Nimmo DG, Mac Nally R, Cunningham SC et al (2015) Vive la résistance: reviving resistance for 21st century conservation. Trends Ecol Evol 30:516–523
Noss RF, Lindenmayer DB (2006) The ecological effects of salvage logging after natural disturbance. Conserv Biol 20:946–948
Pausas JG, Keeley JE (2014) Evolutionary ecology of resprouting and seeding in fire-prone ecosystems. New Phytol 204:55–65
Pausas JG, Llovet J, Anselm R, Vallejo R (2008) Are wildfires a disaster in the Mediterranean basin?—a review. Int J Wildl Fire 17:713–723
Pesendorfer MB, Sillett TS, Koenig WD, Morrison SA (2016) Scatter-hoarding corvids as seed dispersers for oaks and pines: a review of a widely distributed mutualism and its utility to habitat restoration. Condor 118:215–237
Peter B, Bogdanski B (2010) The economics of salvage harvesting and reforestation in British Columbia’s mountain pine beetle-affected forests. Nat Res Canada, Canada
Pons P, Rost J (2016) The challenge of conserving biodiversity in harvested burned forests. Conserv Biol 31:226–228
Puerta-Piñero C, Sánchez-Miranda A, Leverkus A, Castro J (2010) Management of burnt wood after fire affects post-dispersal acorn predation. For Ecol Manage 260:345–352
Radeloff VC, Mladenoff DJ, Boyce MS (2000) Effects of interacting disturbances on landscape patterns: Budworm defoliation and salvage logging. Ecol Appl 10:233–247
Rey Benayas JM, Navarro J, Espigares T et al (2005) Effects of artificial shading and weed mowing in reforestation of Mediterranean abandoned cropland with contrasting Quercus species. For Ecol Manage 212:302–314
Rost J, Clavero M, Bas JM, Pons P (2010) Building wood debris piles benefits avian seed dispersers in burned and logged Mediterranean pine forests. For Ecol Manage 260:79–86
Royo AA, Peterson CJ, Stanovick JS, Carson WP (2016) Evaluating the ecological impacts of salvage logging: can natural and anthropogenic disturbances promote coexistence? Ecology 97:1566–1582
Schiermeier Q (2016) Pristine forest at risk. Nature 530:393
Seidl R, Thom D, Kautz M et al (2017) Forest disturbances under climate change. Nat Clim Chang 7:395–402
Serrano-Ortiz P, Marañón-Jiménez S, Reverter BR et al (2011) Post-fire salvage logging reduces carbon sequestration in Mediterranean coniferous forest. For Ecol Manage 262:2287–2296
Siles G, Rey PJ, Alcántara JM (2010) Post-fire restoration of Mediterranean forests: testing assembly rules mediated by facilitation. Basic Appl Ecol 11:422–431
Swanson ME, Franklin JF, Beschta RL et al (2011) The forgotten stage of forest succession: early-successional ecosystems on forest sites. Front Ecol Environ 9:117–125
Taboada A, Fernández-García V, Marcos E, Calvo L (2018) Interactions between large high-severity fires and salvage logging on a short return interval reduce the regrowth of fire-prone serotinous forests. For Ecol Manage 414:54–63
Thom D, Seidl R (2016) Natural disturbance impacts on ecosystem services and biodiversity in temperate and boreal forests. Biol Rev 91:760–781
Thorn S, Bässler C, Bernhardt-Römermann M et al (2015) Changes in the dominant assembly mechanism drives species loss caused by declining resources. Ecol Lett 19:109–215
Thorn S, Bässler C, Brandl R et al (2018) Impacts of salvage logging on biodiversity—a meta-analysis. J Appl Ecol 55:279–289
Thorn S, Chao A, Georgiev KB et al (2020a) Estimating retention benchmarks for salvage logging to protect biodiversity. Nat Commun 11:4762
Thorn S, Seibold S, Leverkus AB et al (2020b) The living dead–acknowledging life after tree death to stop forest degradation. Front Ecol Environ 18:505–512
Trisos CH, Merow C, Pigot AL (2020) The projected timing of abrupt ecological disruption from climate change. Nature 580:496–501
Urretavizcaya MF, Defosse GE (2019) Restoration of burned and post-fire logged Austrocedrus chilensis stands in Patagonia: effects of competition and environmental conditions on seedling survival and growth. Int J Wildl Fire 28:365–376
Van Nieuwstadt MGL, Sheil D, Kartawinata K (2001) The ecological consequences of logging in the burned forests of East Kalimantan, Indonesia. Conserv Biol 15:1183–1186
Zamora RJ, Pérez Luque AJ, Bonet García FJ et al (eds) (2015) La huella del Cambio Global en Sierra Nevada: Retos para la conservación. Consejería de Medio Ambiente y Ordenación del Territorio, Junta de Andalucía
Acknowledgements
We are grateful to numerous colleagues and students who helped conceptualize and conduct this experiment. The Direction of the Natural Park of Sierra Nevada and the Consejería de Medio Ambiente from the Andalusian Regional Government provided invaluable support. Key funding was provided by grants 10/2005 from Organismo Autónomo de Parques Nacionales, SUM2006-00010-00-00 from INIA, CGL2008-01671 from Ministerio de Ciencia e Innovación, P12-RNM-2705 from Junta de Andalucía, and an ongoing ERIC LifeWatch Mountain Thematic Center, supplemented by several other grants and personal fellowships that are acknowledged in individual publications.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2022 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this chapter

Cite this chapter
Leverkus, A.B., Castro, J. (2022). Restoration of Mediterranean Forest Ecosystems After Major Disturbances: The Lanjarón Post-fire Experiment Over 15 Years of Succession. In: Zamora, R., Oliva, M. (eds) The Landscape of the Sierra Nevada. Springer, Cham. https://doi.org/10.1007/978-3-030-94219-9_14
Download citation
DOI: https://doi.org/10.1007/978-3-030-94219-9_14
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-94218-2
Online ISBN: 978-3-030-94219-9
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)