
Abstract
Two key processes constitute the dynamics of planted pine forests (Pinus halepensis, P. brutia) in Mediterranean Israel: pine regeneration and reestablishment of native broad-leaved tree species (broad-leafs). These two processes are regulated mainly by aridity, overstory cover and grazing. Pine forests in Israel have been planted throughout the semiarid to dry-subhumid climate zones (Wetness Index = 0.13–0.50) and over variable geology and topography. They exhibit a range of pine overstory cover levels (Leaf Area Index = 1–3.5) and are subjected to variable grazing by cattle, sheep and goats. Pine regeneration is mainly observed in P. halepensis stands with annual rainfall >350–400 mm. Above this threshold, the extent of pine regeneration is highly unpredictable and strongly influenced by bedrock type, being considerably higher on soft than on hard calcareous bedrock. Pine regeneration is negatively affected by overstory cover and grazing, which limit survival and growth of recruits. Pine regeneration in P. brutia stands is currently limited, though overstory thinning appears effective in promoting this process. Broad-leafs reestablishment occurs where annual rainfall exceeds 450 mm; it increases sharply with rainfall and is higher on north- than on south-facing slopes. Broad-leafs recruitment is positively influenced by overstory cover while the effect of grazing is minor. Aridity, overstory cover and grazing all restrict the growth rate of broad-leaf recruits. Based on our understanding of these processes, management guidelines are proposed for converting the manmade pine monocultures into complex, sustainable mixed-forest ecosystems.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

1 Introduction
Pine plantations were established throughout the Mediterranean zone of Israel during the last century as part of a comprehensive afforestation effort (Osem et al. 2008). These forests were planted as even-aged monocultures based mainly on the local Pinus halepensis Mill. and to a lesser extent, the exotic P. brutia Ten., with unified silvicultural methodologies (e.g. site preparation, planting density and thinning regime) applied throughout a wide geo-climatic range. Establishing even-aged, monospecific plantations based on a few coniferous pioneer species proved efficient in achieving the historical goals and targets of afforestation. However, it resulted in the creation of forest ecosystems characterized by a short life cycle and limited diversity, complexity and ecological integrity (Noy-Meir 1989), and high sensitivity to pest outbreaks, fire, drought and extreme climatic events (snowfall and strong winds, Boneh and Silverstone 1999). However, as these forest systems matured, they underwent a series of vegetation processes depending on a variety of natural- and human-related drivers (Osem et al. 2012, 2017b). Here, we review two key processes that constitute the forest dynamics of pine plantations in Israel and discuss the major factors and interactions that drive them. Based on our understandings of these processes, we propose silvicultural guidelines for gradually turning these simply structured manmade forests into complex, sustainable mixed-forest ecosystems.
Two key parallel processes that occur continuously, in the understory of pine plantations in Mediterranean Israel affect the future of their forest structure: (1) pine regeneration – the spontaneous recruitment and growth of pines and (2) Native broad-leafs reestablishment – the spontaneous recruitment, resprouting and growth of native broadleaved tree species. While these two processes interact with one another, they are also bound to external regulators, mainly: (1) aridity – abiotic habitat factors determining the amount of available water for plants (e.g. rainfall, potential evapotranspiration, soil type and aspect); (2) pine overstory cover – the extent of shading by overstory pine canopies and consequent availability of light, water and nutrients for understory regeneration and growth; and (3) grazing – the consumption by livestock of vegetative material in the forest understory. Despite their critical importance as regulators of forest dynamics, these three factors have not yet been discussed, in an integrative way, as key components of a silvicultural management strategy for developing diverse and sustainable mixed-forest ecosystems in water-limited environments.
2 Regulators
2.1 Aridity
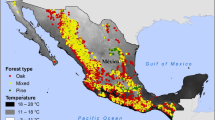
According to the wetness index (WI = annual precipitation/annual potential evapotranspiration) for climatic aridity, pine forests in Israel are distributed along an aridity range of WI = 0.13–0.50 (i.e., semiarid to dry-subhumid climate, Perelman 2018). This is mainly due to the high spatial variation in mean annual rainfall, in the range of 260–850 mm (Wilson 2016), falling mostly during winter (December–March, Fig. 32.1), with some minor additional variation in potential evapotranspiration ranging between 1750 and 1980 mm/year. The development and function of pine forests in water-limited habitats in Israel is known to depend considerably on the amount and distribution of rainfall (Osem et al. 2009, 2012). In addition to climate, habitat aridity is also known to be influenced by edaphic and topographic conditions such as bedrock type, topographic slope and aspect and soil type and depth.

Average monthly rainfall (mm) in 10 pine forest areas in Israel.  Yatir;
Yatir;  Lahav;
Lahav;  Shaharia;
Shaharia;  Eshtaol;
Eshtaol;  Givat-Hamoreh;
Givat-Hamoreh;  Nazereth;
Nazereth;  HaKedoshim;
HaKedoshim;  Yaaranim;
Yaaranim;  Maalot;
Maalot;  Sasa. (Osem et al. 2009)
Sasa. (Osem et al. 2009)
2.2 Pine Overstory Cover
The overstory cover of mature trees in P. halepensis forests in Israel, quantified by leaf area index (LAI), ranges from 1 to 3.5 m m−2 (Perelman 2018). This range corresponds to a stand density of 40–880 trees ha−1, stand stem basal area (BA) of 2.5–28 m2 ha−1 and overstory canopy cover (CC) of 30–100%. Overstory LAI is highly variable across the forests, influenced by forest management (e.g., thinning), as well as by a variety of natural disturbances (e.g., drought, wind-throw, snow). It was found to be linearly and significantly related to the level of aridity (WI), with mean LAI values ranging from 1.5 to 2.5 throughout the climatic gradient of the pine forests’ distribution range in Israel (Perelman 2018). Tree density and overstory cover of mature P. brutia stands in Israel were found to be higher (by ~30% and 15% respectively) than that of P. halepensis stands, and not as closely related to aridity, but more influenced by bedrock type (higher on soft than on hard calcareous bedrock, Wilson 2016).
2.3 Grazing
Grazing by domesticated livestock, cattle, sheep and goats is very common in Israel’s pine forests. Based on regional foresters’ knowledge it was estimated that 60% of the forests in the central forest region of Israel and 95% in the northern and southern regions are under some kind of livestock grazing, (Zeidan S et al. pers. comm.). The proportion of forest area grazed by cattle vs. sheep and goats is also variable among the regions. In the north, cattle dominate (80%) the grazed forested area, while in the south and center sheep-goat herds dominate (95% of the grazed forested area). Grazing is considered by local foresters to be the primary tool for fire hazard reduction, seeking maximum herbaceous biomass removal (Ne’eman et al. 1997; Osem et al. 2017b). Grazing in Israeli forest systems is highly variable and detailed information on actual grazing regimes is lacking. Cattle grazing in pine forests typically occurs from mid-winter to late spring or summer (4–6 months) with grazing pressures ranging from 1 to 7 ha per cattle head, depending on forage availability, accessibility and other factors (Seligman et al. 2016). Sheep and goat grazing in the pine forests is usually allowed when the herbaceous vegetation reaches 70–80 g m−2 dry biomass and continue for several months until the remaining dry herbage is not more than 75 g m−2 (Seligman et al. 2016). According to an assessment of carrying capacity of planted pine forests in central Israel, the herbaceous vegetation can support ca. 100–200 sheep grazing days per ha while the woody vegetation can support as much as 200–400 goat grazing days per ha (Evlagon et al. 2012). However, the extent of forage exploitation in these areas is highly variable, ranging from less than 5% in some forests to more than 95% in others (Evlagon et al. 2012).
3 Pine Regeneration
Pine regeneration is the process through which the next conifer forest generation establishes in the understory of mature forests and is essential for pine forest sustainability. This process involves seed production and dispersal, seed germination, seedling survival and growth, tree maturation and production. All these processes are influenced by the abovementioned regulators.
3.1 Aridity
In a study that compared seed rain and seedling emergence within P. halepensis plantations, across three ecoregions differing in rainfall (600, 430 and 280 mm year−1, WI = 0.33, 0.23 and 0.13), large differences were found in seed rain (~250, 150, 25 seeds m−2 year−1), and in emerged seedling density (~13, 2, 0.3 seedlings m−2), corresponding to the variation in aridity level (Osem et al. 2013, Fig. 32.2). Seed rain and seedling emergence were higher on north- vs. south-facing slopes, highlighting the importance of water availability for the natural regeneration potential of pine forests in Israel. A study on the regeneration of a P. brutia plantation in a relatively humid ecoregion (600 mm annual rainfall, WI = 0.33) found seed rain of ~154 seeds m−2 year−1 and emerged seedling density of ~8 seedlings m−2, which corresponded with those found for P. halepensis under similar rainfall amounts (Osem et al. 2015).

Average density of emerged pine seedlings on north- and south-facing slopes, in late winter (March) in Israel: Mishmar-Ha’Emek Forest 1972–1975 (MHF), Shaharia Forest 2008–2009 (SF) and Yatir Forest 2005–2007 (YF). South-facing slopes were not measured in MHF. Error bars indicate standard error (SE). (Osem et al. 2013)
Regarding the density of established pine recruits (i.e., advanced regeneration) in the understory of mature (>30 year old) P. halepensis plantations, along the aridity gradient in Israel, Osem et al. (2009) found a threshold of 400 mm annual rainfall (WI = 0.2) below which pine regeneration is very limited. Thus, under such conditions the ability of natural regeneration as required for sustainable pine forest cover is lacking. However, above 400 mm annual rainfall, a wide range of recruit densities was found (0–1565 recruit ha−1) with no clear relationship to annual rainfall amount nor to topographic aspect (Fig. 32.3).

Regeneration density of Aleppo pine trees in the forest understory along the rainfall gradient in Israel.  North-facing slopes;
North-facing slopes;  South-facing slopes. (Osem et al. 2009)
South-facing slopes. (Osem et al. 2009)
Previously, in his work, Schiller (1978) highlighted the important effect of bedrock type on natural regeneration in three P. halepensis forests in northern and central Israel. Establishment rates were higher on soft (chalk and marl) than on hard (limestone and dolomite) calcareous bedrock types. This effect was attributed to higher soil/rock water holding capacity resulting in better water conditions and to lower competition by annual herbs in habitats on soft bedrock types. A comprehensive survey of advanced regeneration conducted over the long-term monitoring plot setup of Israel’s mature pine forests, provided further support for the great importance of bedrock type, with this factor alone contributing 78% of the explained variance in P. halepensis regeneration (Wilson 2016). In this study, advanced regeneration was also found to be positively related to rainfall amount, but this relationship was prevalent only on soft bedrock and was relatively weak. According to this analysis, natural regeneration of P. halepensis appears above an annual rainfall threshold of 350 mm (WI = 0.18) on soft calcareous bedrock. In contrast, very low regeneration was found in mature P. brutia forests throughout a similar long-term monitoring plot setup, with no clear relationship to rainfall amount or bedrock type.
Support for the importance of herbs in limiting pine regeneration was gained from a study looking at advanced pine regeneration in a semiarid P. halepensis forest (Shaharia Forest) located on the dry margins of the natural regeneration ability of P. halepensis. In this study natural regeneration occurred only in some of the more rainy years, and seedling survival was negatively and significantly related to the biomass of the herbaceous vegetation (Osem et al. 2013). Furthermore, manipulation of herbaceous biomass by ploughing or mowing improved pine seedling survival (Fig. 32.4).

Pine seedling density in mid-spring (May) as affected by site preparation treatments on south- and north-facing slopes in Shaharia Forest 2009. Different letters above columns indicate significant differences (P < 0.05). Error bars indicate standard error (SE). (Osem et al. 2013)
3.2 Overstory Cover
The first study on the effect of overstory cover on P. halepensis recruitment in Israel, by Schiller (1978), was a silvicultural thinning experiment on mature stands (43-year-old, Mishmar-HaEmek Forest) growing in typically favourable sites for pine regeneration (600 mm annual rainfall, WI = 0.33, north-facing slope and soft chalk bedrock). Thinning forest stands of ca. 300 trees ha−1 (canopy cover = 66%) to 200, 100 and 0 trees ha−1 (CC = 53%, 37% and 0%, respectively) resulted in much lower seed rain and germinating seedling density but increased seedling survival and growth. Overall, the densities of established recruits 6 years after thinning were 3200, 6700, 1300 and 0 recruits ha−1 in the 0, 100, 200 and 300 (non-thinned control) trees ha−1 treatments, respectively (Table 32.1).
Osem et al. (2017a) conducted a similar experiment in P. halepensis stands growing in less favorable sites (540 mm annual rainfall, WI = 0.30, west-facing slopes and hard lime bedrock). In this study, mature forest stands (40 year old, HaKedoshim Forest) of ~600 trees ha−1 (LAI = 2.0; CC = 88%) were thinned to 300, 100 and 0 trees ha−1 (LAI = 1.5, 1.0 and 0; CC = 61%, 48% and 0%, respectively). The responses of pine recruitment and growth were similar to those described by Schiller (1978) but with pine recruit densities being lower by one level of magnitude i.e., 250, 180, 80 and 50 recruit ha−1 in the 0, 100, 300 and 550 (non-thinned control) trees ha−1 treatments, respectively.
Osem et al. (2015) examined the effect of overstory thinning on natural regeneration in P. brutia stands (50-year-old, Mount Horshan Forest) with 600 mm annual rainfall (WI = 0.33) and soft chalk and marl bedrock. Pine seed rain, seedling germination and survival (recruitment) were compared between non-thinned stands of ~230 trees ha−1 (CC = 95%) and thinned stands with ~100 trees ha−1 (CC = 55%). Similarly to the results of Schiller (1978), overstory thinning decreased seed rain (40%) and winter seedling emergence (66%) but increased seedling survival and resulting recruit density by more than threefold. Moreover, in the non-thinned plots most of the recruits were not taller than 20 cm, while in the thinned plots most of them were taller than 20 cm, with many reaching heights of over 1 m and even 2 m. Finally, when plotting recruit height vs. age it became evident that in non-thinned plots the amount of time needed for recruits to reach a certain height was about twice that needed in the thinned plots. We note that in this study, recorded pine recruit densities were extremely high, with 31,000 and 9000 recruits ha−1 in the thinned and non-thinned stands, respectively. Such recruitment levels are not in line with the common observation (as described above) according to which natural regeneration in P. brutia forests of Israel is quite limited compared to that of P. halepensis. This result may indicate exceptionally favorable conditions for pine regeneration prevailing in the specific studied area (e.g., a combination of high annual rainfall and soft calcareous bedrock).
Ashkenazi (2016) examined pine regeneration in P. brutia stands (50-year-old, Begin Park Forests, Judean Mountains) after they were thinned from ~250 trees ha−1 (CC = 78%, control) to 130 (CC = 62%, moderate thinning) and 50 (CC = 12%, heavy thinning) trees ha−1. Seven years after thinning, pine recruit density was 430 and 700 recruits ha−1 in the heavy and moderate thinning treatments, respectively, while in the non-thinned control plots it was zero. This study was conducted in sites with 560 mm annual rainfall (WI = 0.31) and complex soft-hard calcareous bedrock formation. While the reported recruitment levels are much lower than those described in Mount Horshan Forest, they highlight the capacity of P. brutia stands to regenerate naturally in a variety of habitat types in Israel following overstory thinning.
3.3 Grazing
Livestock grazing is widely recognized as a major regulator of vegetation structure in Mediterranean ecosystems as well as a silvicultural tool for fire fuel load control. However, the effects of grazing on vegetation dynamics in the conifer forests of Israel have been hardly studied. In their study, Osem et al. (2015) specifically addressed the effect of cattle grazing on pine recruitment in mature (50-year-old) P. brutia stands (Mount Horshan Forest). Density of newly emerged seedlings, during the winter, was not found to be affected by grazing. However, the density of established pine recruits was significantly reduced by 25% under grazing in non-thinned plots (overstory CC = 95%) and by as much as 85% in thinned ones (CC = 55%). Moreover, in the thinned plots, with no cattle grazing (exclosures), high densities of recruits reaching 101–200 cm in height (~6000 recruit ha−1) and 201–400 cm in height (~2000 recruit ha−1) were found, while under grazing the existence of recruits taller than 100 cm was supressed completely (Fig. 32.5). Thus, grazing reduced pine recruits’ survival and growth; this effect was particularly remarkable in thinned plots with low overstory cover. In contrast, grazing had a significant, positive effect on water availability for young recruits, as measured by predawn shoot water potential of 6-month-old seedlings; this probably indicates reduced water consumption by the competing understory vegetation in grazed areas (Osem et al. 2015, 2017b).

Density of Pinus brutia saplings, by height classes, as affected by cattle grazing in (a) thinned and (b) non-thinned plots in Mount Horshan Forest, Israel, November 2009. Bars indicate SE, N = 20. (Osem et al. 2015)
Cohen (2019) examined the effect of heavy goat grazing pressure on post-fire P. halepensis recruits (5000–10,000 recruits ha−1) in the Mount Carmel region (600 mm annual rainfall, complex limestone, chalk and marl bedrock). The herd was introduced into the regenerating forest area 5 years after the fire (grazing regime: 2000 grazing days per ha during the spring) for three consecutive years (2016–2018). In this study, grazing did not affect pine recruit density, that is, it did not increase recruit mortality, but it restricted recruit growth as well as cone production with respect to the ungrazed control.
4 Broad-Leafs Reestablishment
Regeneration of Broad-leaved tree species in the understories of conifer plantations may take place either by seed germination, from external or internal seed sources, or by resprouting. Assessing this process is more complicated as it involves a variety of tree species differing in their ecological requirements.
4.1 Aridity
Osem et al. (2009) examined the natural recruitment of broad-leaved trees in P. halepensis forests distributed along the aridity gradient of Israel (280–900 mm annual rainfall), on north and south-facing slopes. This survey focused on ungrazed, mature stands (40–50 year old) with overstory tree densities corresponding to accepted thinning practices (300–350 trees ha−1). Broadleaved trees appeared in the conifer forest understories in areas of at least 450 and 500 mm annual rainfall on north- and south-facing slopes, respectively. Above these thresholds, the density of naturally regenerating broad-leaved trees increased sharply with increasing rainfall amount reaching up to ~2000 and 8000 trees ha−1 in the most humid forests (>700 mm annual rainfall) on south- and north-facing slopes, respectively (significant rainfall×aspect interaction, Fig. 32.6). No effect of climatic aridity on the size of the regenerating trees was found; average tree height was typically in the range 1–2 m. One variable that was found to be the most strongly related to rainfall amount (R2 > 0.8) was the species richness of the regenerating broad-leaved trees, which reached ~5 coexisting species per 200 m2 in the most humid forests (Fig. 32.7). The effect of topographic aspect on tree species richness was minor. Overall, 12 species of broad-leaved trees were recorded throughout the climatic range. Among the broad-leaved tree species regenerating in the forest understory, the most dominant was common oak (Quercus calliprinos Webb, 40–90% of total broad-leaved tree density) which appeared from 500 mm annual rainfall and increased linearly with increasing rainfall amount up to ~1000 and 2000 trees ha−1 in the most humid forests, on north- and south-facing slopes, respectively. In several sites of the most humid forests, the dominance of Q. calliprinos was overtaken by other species, namely, Arbutus andrachne L., Q. boissieri Reut. and Laurus nobilis L.. These species were restricted to forest sites where average annual rainfall exceeded 600 mm. In contrast, Ceratonia siliqua L. was found to be relatively important in forests receiving 450–600 mm annual rainfall, but decreased in importance at higher rainfall levels. One other important tree species was Pistacia palaestina Boiss., which was quite common from 500 mm annual rainfall and above.

Total density of regenerating trees in the forest understory on north-facing slopes ( , R2=0.6, P=0.01) and south-facing slopes (
, R2=0.6, P=0.01) and south-facing slopes ( , R2=0.77, P=0.002) along the rainfall gradient in Israel. (Osem et al. 2009)
, R2=0.77, P=0.002) along the rainfall gradient in Israel. (Osem et al. 2009)

Species richness of regenerating trees in the forest understory on north-facing slopes ( , R2=0.82, P=0.0009) and south-facing slopes (
, R2=0.82, P=0.0009) and south-facing slopes ( , R2=0.83, P=0.0007) along the rainfall gradient in Israel. (Osem et al. 2009)
, R2=0.83, P=0.0007) along the rainfall gradient in Israel. (Osem et al. 2009)
Wilson (2016) conducted a more extensive understory survey over the long-term monitoring plot setup of Israel’s mature pine forests. He examined broad-leaved tree reestablishment in P. halepensis (48 stands) and P. brutia (32 stands) forests, along the rainfall gradient (250–850 mm annual rainfall) and across bedrock type (hard vs. soft calcareous bedrock), topographic aspect (north vs. south) and elevation (70–820 m). Broad-leafs reestablishment was highly variable throughout the climatic range with Q. calliprinos being the most dominant tree species. The extent of broad-leafs reestablishment was positively and linearly related to annual rainfall amount in both P. halepensis and P. brutia forests. There were no significant effects of forest type (P. halepensis vs. P. brutia), topographic aspect, bedrock type or elevation level.
4.2 Overstory Cover
Osem et al. (2015) examined the effect of overstory thinning on broad-leafs reestablishment in P. brutia stands (50 year old, Mount Horshan Forest) with 600 mm annual rainfall and soft chalk bedrock. The density and size of regenerating trees in the forest understory were compared between non-thinned stands of ~230 trees ha−1 (CC = 95%) and thinned stands of ~100 trees ha−1 (CC = 55%). Native broad-leaved tree species including Q. ithaburensis Decne., Q. calliprinos, P. palaestina, Rhamnus alaternus L., Styrax officinalis L., Phillyrea latifolia L. and C. siliqua were found throughout the forest understory, with recruit densities of 1000–2400 trees ha−1. Recruit density of Q. ithaburensis, the most dominant of the regenerating broad-leaved tree species in this forest, was nearly four-fold higher in the non-thinned than in the thinned forest plots. No significant effect of overstory thinning was found on the recruit density of the other broad-leaved tree species. Moreover, no effect of overstory thinning was found on the height distribution, canopy cover or species richness of broad-leaf recruits.
Ashkenazi (2016) looked at broad-leafs reestablishment in P. brutia stands (50 year old, Begin Park Forests, Judean Mountains) with 560 mm annual rainfall and complex soft-hard calcareous bedrock formation. In this study trees were thinned from ~250 trees ha−1 (CC = 78%, control) to 130 (CC = 62%, moderate thinning) and 50 (CC = 12%, heavy thinning) trees ha−1. Seven years after thinning the density of regenerating broadleaved tree species was similar among treatments with an average of ~1250 trees ha−1. Regenerating tree species were mainly Q. calliprinos and P. palaestina that were equally abundant, with an additional minor contribution of Crataegus aronia (L.) DC.
In another study conducted by Osem et al. (2017a), broad-leafs reestablishment was examined in P. halepensis stands with 540 mm annual rainfall on west-facing slopes and hard calcareous bedrock (lime). In this study, mature forest stands (40 year old, HaKedoshim Forest) of ~550 trees ha−1 (LAI = 2.0; CC = 88%) were thinned to 300, 100 and 0 trees ha−1 (LAI = 1.5, 1.0 and 0; CC = 61%, 48% and 0%, respectively). Naturally regenerating broadleaved tree species included Q. calliprinos, P. palaestina, C. siliqua, P. latifolia and Olea europaea L.. Among these, Q. calliprinos was highly dominant and the only species responding to overstory thinning. While prior to thinning the density of regenerating oaks was similar among the forest plots with an average of ~400 trees ha−1, 7 years after thinning it continued to increase in the non-thinned and moderately thinned plots (550 and 300 trees ha−1) but slightly decreased in the heavily thinned and clear-cut plots (100 and 0 trees ha−1). In contrast, the growth rate of oak recruits was positively affected by overstory thinning (i.e., relative growth rate increased three-fold following complete removal of overstory cover).
Cooper et al. (2014) specifically examined the effect of overstory cover on the performance of young Q. ithaburensis trees growing in the understory of a mature P. brutia forest (Metzer Forest, 45 year old, ~600 mm annual rainfall, soft to semi-hard chalk bedrock). This study compared carbon assimilation, water use and growth rate of artificially sown oaks (of identical age) along an overstory cover gradient (irradiance range of 20–100%). Ten years after the creation of the overstory gradient, the size of the oaks was strongly affected by overstory cover, with oaks growing under no cover (irradiance = 100%) reaching more than 10 times the height and 300 times the stem basal area of those growing under dense overstory cover (irradiance≈20%, Fig. 32.8). Correspondingly, irradiance level had a positive effect not only on carbon assimilation rate but also on water use efficiency.

Tree height (a, y = 0.30x–19.58, R2 = 0.85, P < 0.0001) and stem basal area (b, y = 0.39e0.005x, R2 = 0.81, P < 0.0001) of 20-year-old oaks (Quercus ithaburensis) as a function of daily average irradiance in Metzer Forest, Israel, 2009. (Cooper et al. 2014)
4.3 Grazing
Osem et al. (2015) studied the effects of cattle grazing on broad-leafs reestablishment in mature (50 year.old) P. brutia stands (Mount Horshan Forest). Recruits of native broad-leaved tree species including Q. ithaburensis, Q. calliprinos, P. palaestina, R. alaternus, S. officinalis, P. latifolia and C. siliqua were found throughout the forest understory. Recruit density of Q. ithaburensis, the most abundant of these species, was nearly three-fold higher in ungrazed than in grazed areas. However, no significant effect of grazing was found on the recruit density of any of the other broad-leaved tree species. When examining total recruit density by height class it became evident that the density of larger recruits (20 cm to 4 m in height) was higher in the ungrazed than in the grazed areas, while the density of smaller recruits (<20 cm) was higher in grazed than in ungrazed areas (Fig. 32.9). This resulted in the total cover of broad-leaved tree recruits being more than three-fold higher in ungrazed than in grazed areas. Furthermore, the species richness of the broad-leaved tree recruits was higher in ungrazed than in grazed areas.

Density of broadleaved tree species saplings by height classes as affected by cattle grazing in thinned (a) and non-thinned (b) plots. Mount Horshan Forest, Israel, November 2009. Bars indicate SE, N = 20. (Osem et al. 2015)
5 Integrative Discussion
The ways by which aridity, overstory cover and livestock grazing regulate tree regeneration in the understory of Israel’s pine forests may be summarized as follows:
Natural pine recruitment in the understories of Israel’s pine forests (i.e., advance regeneration) occurs in P. halepensis stands in forests receiving more than ~350–400 mm annual rainfall depending on bedrock formation (i.e., lower threshold on soft- than on hard-calcareous bedrock). Above these thresholds, the relationship between aridity and pine recruitment is weak and observed mainly on soft bedrock formations. This difficulty is most likely related to a negative feedback by competing annual herbs that normally increase at higher rainfall amounts and on hard vs. soft bedrock formations. In addition, pine recruitment is negatively affected by overstory cover and livestock grazing which consistently hamper seedling survival. Moreover, aridity, overstory cover and livestock grazing all restrict the growth (size gain) of pine recruits.
Currently, the extent of advance regeneration occurring in P. brutia stands is very limited. This may be partly explained by the fact that mature P. brutia stands in Israel are considerably denser with higher overstory cover than P. halepensis stands (Wilson 2016). In addition, P. brutia seedlings are probably more sensitive to drought. Nevertheless, this pine species has proven its capacity to regenerate naturally under some habitat conditions following overstory thinning. More research is needed to determine the environmental thresholds for natural regeneration of P. brutia in Israel.
broad-leafs recruitment appears in the understory of Israel’s pine forests at locations exceeding 450–500 mm of annual rainfall with a linear increase in tree recruit density and species diversity with increasing rainfall. This effect of climatic aridity is accompanied by an additional positive effect of north- vs. south-facing slopes, further illustrating the importance of water availability for broad-leafs recruitment. In contrast to pine recruitment, broad-leafs recruitment is positively influenced by overstory cover, while the effect of livestock grazing on this process appears to be minor with the exception of Q. ithaburensis which appears to be rather sensitive to grazing. When considering the growth of broad-leaf recruits, the effects of aridity, overstory cover and grazing are all consistently negative as observed for pine recruits.
In Mediterranean climate regions which are undergoing climate change (i.e., higher temperatures, reduced precipitation), a lack of natural regeneration is considered to be one of the main limitations threatening the sustainability of forest ecosystems (Benavides et al. 2013; Manso et al. 2014; Benavides et al. 2016; Vergarechea et al. 2019). This threat is further exacerbated by other human-related factors such as inappropriate forest management practices (Martin-Alcon et al. 2015; Ruano et al. 2015; Simões et al. 2016) and overgrazing (Plieninger et al. 2011; Lempesi et al. 2017). Climatic aridity leading to water scarcity, particularly summer drought during the early seedling establishment phase, is frequently viewed as the main bottleneck for natural regeneration in Mediterranean forests (Vizcaino-Palomar et al. 2014; Benavides et al. 2016; Principe et al. 2019; Vergarechea et al. 2019). This constraint may be either strengthened or mitigated by local conditions such as topography and soil conditions (González-Martínez and Bravo 2001; Principe et al. 2019), light regime (Prevosto et al. 2012; Adili et al. 2013; Calama et al. 2013, 2015; Benavides et al. 2013, 2016; Martin-Alcon et al. 2015; Vergarechea et al. 2019) and the characteristics of the neighboring vegetation (Benavides et al. 2013, 2016; Simões et al. 2016; Vergarechea et al. 2019). The effect of overstory cover on natural regeneration in Mediterranean forests varies across forest types and tree species. While overstory shading can have positive (Calama et al. 2013, 2015, Martin-Alcon et al. 2015 – with regard to broad-leafs, Vergarechea et al. 2019) or negative (Martin-Alcon et al. 2015 – with regard to pines) effects on early seedling survival, it usually has negative effects on the growth rate of recruits (Adili et al. 2013; Benavides et al. 2013, 2016; Martin-Alcon et al. 2015; Calama et al. 2015). In addition to its direct effect on light availability for recruits, overstory cover may influence natural regeneration through indirect effects on the neighboring understory vegetation which may compete with (Prevosto et al. 2012; Benavides et al. 2013, 2016; Vergarechea et al. 2019) or facilitate (Benavides et al. 2013, 2016) tree recruits. In contrast to overstory cover, the effects of livestock grazing on natural regeneration in Mediterranean forests appear to be quite consistent, with commonly negative effects on recruit survival and growth observed across various forest types (González-Martínez and Bravo 2001; Cierjacks and Hensen 2004; Alias et al. 2010; Plieninger et al. 2011; Lempesi et al. 2017). However, this outcome is usually associated with intense grazing regimes, while moderate and carefully managed livestock grazing may facilitate natural regeneration in water-limited Mediterranean forests through the reduction of competing vegetation (Tyler et al. 2008).
6 Management Guidelines for Developing Mixed Forests
Considering these conclusions, I present management guidelines for turning simply structured monoculture pine forests, in Mediterranean Israel, into sustainable mixed-forest ecosystems.
6.1 Aridity
-
In forest habitats with <350 mm annual rainfall (WI < 0.17), neither pine forest renewal nor broad-leafs establishment should rely on natural regeneration processes.
-
In forest habitats with >400 mm annual rainfall (WI ≥ 0.21), on soft calcareous bedrock, natural pine regeneration will most probably suffice for forest renewal. In such habitats, management of pine regeneration should focus on early age thinning to avoid the development of excessively dense forest stands. The target density of regenerating pine stands should decrease with increasing aridity. For example, recent management guidelines in Israel recommend the final density of P. halepensis stands to be set before the age of 30 year at 300, 250 and 200 trees ha−1 for annual rainfall levels of 250–350, 351–500 and >500 mm, respectively.
-
In habitats with >400 mm annual rainfall, on hard calcareous bedrock as well as on various other soft-hard bedrock complexes, the extent of natural pine regeneration is highly unpredictable. Advance regeneration assessments may assist in reducing this uncertainty in such habitats, and monitoring successes following regeneration treatments (e.g., regeneration thinning) is necessary.
-
When pine recruitment appears insufficient, herb control may be considered for promoting natural regeneration, although effective implementation of such a practice has not yet been developed and demonstrated in Israel’s pine forests.
-
Broad-leafs reestablishment should be expected in habitats with >450 mm annual rainfall (WI ≥ 0.23), with the density and species diversity of broadleaved tree recruits increasing with higher water availability.
6.2 Overstory Cover
-
High pine overstory cover (≥50%, LAI ≥ 2) promotes the recruitment of native broad-leaved trees.
-
Low overstory cover (≤20%, LAI ≤ 1) is required to allow successful pine recruitment and growth.
-
Overstory thinning for pine regeneration should limit overstory cover to the range 0–20% cover, or LAI = 0–1, depending on target stand structure: 0–5% coverage for single and 6–20% coverage for multi-age structure.
-
Decreased overstory cover will enhance the growth rate of both pine and native broad-leaved trees.
-
Selective thinning may be applied in regenerated mixed stands focusing on regulating tree cover according to habitat aridity, creating variable age structures and promoting tree species diversity. This requires further study.
6.3 Grazing
-
Grazing exclusion is required to allow survival of young pine recruits and enhance the growth of both pine and broad-leaf recruits.
-
Grazing should be excluded for a sufficient time period (approximately 10 year), allowing tree recruits to reach a minimal height of 3 m.
-
Reintroduction of grazing following the satisfactory development of tree recruits would facilitate further tree development through herbaceous and shrub biomass removal and reduced competition, fire hazard reduction and pruning effect.
-
More research is required to examine the various grazing regimes and their effects on forest dynamics.
Using these guidelines for prescribing overstory thinning and livestock grazing regimes according to site water conditions should facilitate the conversion of first generation, even-aged pine monocultures into mixed multi-aged forests of variable stand structure, and promote the diversity and complexity of forest ecosystems in Mediterranean Israel.
6.4 Concluding Remarks
The diversification of forest systems is broadly considered to be a desirable course (Schuler et al. 2017; Sánchez de Dios et al. 2019) promoting biodiversity (Felton et al. 2016; Pukkala 2018; Wang et al. 2019), productivity (Pretzsch et al. 2012, de-Dios-Garcia et al. 2015), and multiple ecosystem services (Felton et al. 2016; Schuler et al. 2017; Pukkala 2018), as well as forest health (Felton et al. 2016; Aoki et al. 2018), resistance (Pretzsch et al. 2013, de-Dios-Garcia et al. 2015), and resilience (Pukkala 2018). Nonetheless, currently, very little is known about these processes in the recently evolving mixed forests of Mediterranean Israel. Furthermore, silvicultural strategies to optimize ecosystem services and promote the resistance and resilience of these semi-natural ecosystems, in the face of climate change, are yet to be developed.
References
Adili B, El Aouni MH, Balandier P (2013) Unravelling the influence of light, litter and understorey vegetation on Pinus pinea natural regeneration. Forestry 86:297–307
Alias S, Bianchi L, Calamini G et al (2010) Shrub facilitation of Quercus ilex and Quercus pubescens regeneration in a wooded pasture in Central Sardinia (Italy). IFOREST 3:16–22
Aoki CF, Cook M, Dunn J et al (2018) Old pests in new places: effects of stand structure and forest type on susceptibility to a bark beetle on the edge of its native range. For Ecol Manag 419:206–219
Ashkenazi M (2016) Fuelbreak treaments and their effect vegetation structure and fire behaviour in Mediterranean coniferous forests. MSc thesis submitted to the Hebrew University of Jerusalem, Istrael
Benavides R, Rabasa SG, Granda E et al (2013) Direct and indirect effects of climate on demography and early growth of Pinus sylvestris at the rear edge: changing roles of biotic and abiotic factors. PLoS One 8:e59824
Benavides R, Escudero A, Coll L et al (2016) Recruitment patterns of four tree species along elevation gradients in Mediterranean mountains: not only climate matters. For Ecol Manag 360:287–296
Boneh O, Silverstone A (1999) Forest renewal in the northern region. Scientific report submitted to the Forestry Division, Keren Kayemeth Le’Israel (KKL), Eshtaol, Israel. (in Hebrew)
Calama R, Puertolas J, Madrigal G et al (2013) Modeling the environmental response of leaf net photosynthesis in Pinus pinea L. natural regeneration. Ecol Model 251:9–21
Calama R, Puertolas J, Manso R et al (2015) Defining the optimal regeneration niche for Pinus pinea L. through physiology-based models for seedling survival and carbon assimilation. Trees-Struct Funct 29:1761–1771
Cierjacks A, Hensen I (2004) Variation of stand structure and regeneration of Mediterranean holm oak along a grazing intensity gradient. Plant Ecol 173:215–223
Cohen N (2019) Effect of goat grazing and pine seedling removal on the plant community, in general, and pine seedlings, in particular, following the Carmel fire. MSc thesis submitted to the Hebrew University of Jerusalem, Israel
Cooper A, Shapira O, Zaidan S et al (2014) Oak restoration in water-limited pine plantations: interactive effects of overstory light interception and water availability on understory oak performance. Eur J For Res 133:661–670. https://doi.org/10.1007/s10342-014-0794-6
de Dios-Garcia J, Pardos M, Calama R (2015) Interannual variability in competitive effects in mixed and monospecific forests of Mediterranean stone pine. For Ecol Manag 358:230–239
Evlagon D, Kommisarchik S, Gurevich B et al (2012) Estimating normative grazing capacity of planted Mediterranean forests in a fire-prone environment. Agric Ecosyst Environ 155:133–141
Felton A, Nilsson U, Sonesson J et al (2016) Replacing monocultures with mixed-species stands: ecosystem service implications of two production forest alternatives in Sweden. Ambio 45:s124–s139
González-Martínez SC, Bravo F (2001) Density and population structure of the natural regeneration of Scots pine (Pinus sylvestris L.) in the High Ebro Basin (Northern Spain). Ann For Sci 58:277–288
Lempesi A, Eleftheriadou A, Delivasi Z et al (2017) Effects of grazing intensity on the regeneration of woody species in an oak woodland. Not Bot Horti Agrobot Cluj Napoca 45:597–601
Manso R, Pukkala T, Pardos M et al (2014) Modelling Pinus pinea forest management to attain natural regeneration under present and future climatic scenarios. Can J For Res 44:250–262
Martin-Alcon S, Coll L, Salekin S (2015) Stand-level drivers of tree-species diversification in Mediterranean pine forests after abandonment of traditional practices. For Ecol Manag 353:107–117
Ne’eman G, Perevolotsky A, Schiller G (1997) The management implications of the Mt. Carmel research project. Int J Wildl Fire 17:343–350
Noy-Meir I (1989) An ecological viewpoint on afforestation in Israel: past and future. Allg Forst Zeitsch 24–26:614–618
Osem Y, Ginsberg P, Tauber I et al (2008) Sustainable management of Mediterranean planted coniferous forests: an Israeli definition. J For 106:38–46
Osem Y, Zangy E, Bney-Moshe E et al (2009) The potential of transforming simple structured pine plantations into mixed Mediterranean forests through natural regeneration along a rainfall gradient. For Ecol Manag 259:14–23. https://doi.org/10.1016/j.foreco.2009.09.034
Osem Y, Zangy E, Bney-Moshe E et al (2012) Understory woody vegetation in manmade Mediterranean pine forests: variation in community structure along a rainfall gradient. Eur J For Res 131:693–704. https://doi.org/10.1007/s10342-011-0542-0
Osem Y, Yavlovich H, Zecharia N et al (2013) Fire-free natural regeneration in water limited Pinus halepensis forests: a silvicultural approach. Eur J For Res 132:679–690
Osem Y, Fogel T, Moshe Y et al (2015) Managing cattle grazing and overstorey cover for the conversion of pine monocultures into mixed Mediterranean woodlands. Appl Veg Sci 18:261–271. https://doi.org/10.1111/avsc.12152
Osem Y, Zangi E, Ashkenazi M et al (2017a) Long term ecological research site for studying the management of planted forests in Israel: the effects of overstory and understory silvicultural treatments on forest vitality and health, natural regeneration and biodiversity in Pinus halepensis forest. Scientific report submitted to the Forestry Division, Keren Kayemeth Le’Israel (KKL), Eshtaol, Israel. (in Hebrew)
Osem Y, Fogel T, Moshe Y et al (2017b) Understory structure and function following release from cattle grazing and overstory thinning in Mediterranean conifer plantations. Ann For Sci 74:22. https://doi.org/10.1007/s13595-017-0622-5
Perelman Y (2018) Leaf area organization in Pinus halepensis forests as related to abiotic environmental factors. MSc thesis submitted to the Hebrew University of Jerusalem, Israel
Plieninger T, Schaich H, Kizos T (2011) Land-use legacies in the forest structure of silvopastoral oak woodlands in the Eastern Mediterranean. Reg Environ Chang 11:603–615
Pretzsch H, Dieler J, Seifert T et al (2012) Climate effects on productivity and resource-use efficiency of Norway spruce (Picea abies [L.] Karst.) and European beech (Fagus sylvatica [L.]) in stands with different spatial mixing patterns. Trees-Struct Funct 26:1343–1360
Pretzsch H, Schutze G, Uhl E (2013) Resistance of European tree species to drought stress in mixed versus pure forests: evidence of stress release by inter-specific facilitation. Plant Biol 15:483–495
Prevosto B, Amandier L, Quesney T et al (2012) Regenerating mature Aleppo pine stands in fire-free conditions: site preparation treatments matter. For Ecol Manag 282:70–77
Principe A, Matos P, Sarris D et al (2019) In Mediterranean drylands microclimate affects more tree seedlings than adult trees. Ecol Indic 106:105476
Pukkala T (2018) Effect of species composition on ecosystem services in European boreal forest. J For Res 29:261–272
Ruano I, Manso R, Fortin M et al (2015) Extreme climate conditions limit seed availability to successfully attain natural regeneration of Pinus pinaster in sandy areas of Central Spain. Can J For Res 45:1795–1802
Sánchez de Dios R, Velazquez JC, Ollero HS (2019) Classification and mapping of Spanish Mediterranean mixed forests. IFOREST 12:480–487
Schiller G (1978) The effect of environmental factors on the natural regeneration of a Pinus halepensis forest. PhD thesis submited to the University of Tel-Aviv, Israel
Schuler LJ, Bugmann H, Snell RS (2017) From monocultures to mixed-species forests: is tree diversity key for providing ecosystem services at the landscape scale? Landsc Ecol 32:1499–1516
Seligman N, Unger E, Henkin Z et al (eds) (2016) About vegetation, animals and people: rangeland management in Israel. Nekudat Chen, Jerusalem
Simões MP, Belo AF, Fernandes M et al (2016) Regeneration patterns of Quercus suber according to montado management systems. Agrofor Syst 90:107–115
Tyler CM, Davis FW, Mahall BE (2008) The relative importance of factors affecting age-specific seedling survival of two co-occurring oak species in southern California. For Ecol Manag 255:3063–3074
Vergarechea M, Calama R, Fortin M et al (2019) Climate-mediated regeneration occurrence in Mediterranean pine forests: a modeling approach. For Ecol Manag 446:10–19
Vizcaino-Palomar N, Revuelta-Eugercios B, Zavala MA et al (2014) The role of population origin and microenvironment in seedling emergence and early survival in Mediterranean maritime pine (Pinus pinaster Aiton). PLoS One 9:e109132
Wang XY, Hua FY, Wang L et al (2019) The biodiversity benefit of native forests and mixed-species plantations over monoculture plantations. Divers Distrib 25:1721–1735
Wilson T (2016) Principal abiotic factors influencing the structure and function of mature pine forests in Israel. MSc thesis submited to Texas A&M University
Acknowledgements
The studies reported in this review were all supported by the forest service of Israel – KKL. I thank Ira Mor, Michael Sprintsin, Chanoch Zoref and Avi Perevolotsky for critical reading of the MS.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2021 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Osem, Y., Moshe, Y. (2021). From First Generation of Pine Monocultures to Mixed-Forest Ecosystems: Biotic and Abiotic Determinants of Pine Forests’ Dynamics in Mediterranean Israel. In: Ne'eman, G., Osem, Y. (eds) Pines and Their Mixed Forest Ecosystems in the Mediterranean Basin. Managing Forest Ecosystems, vol 38. Springer, Cham. https://doi.org/10.1007/978-3-030-63625-8_32
Download citation
DOI: https://doi.org/10.1007/978-3-030-63625-8_32
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-63624-1
Online ISBN: 978-3-030-63625-8
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)