
Abstract
We aimed to elucidate environmental and silvicultural factors that determine the extent of fire-free natural regeneration in east Mediterranean Pinus halepensis forests. The specific aims were to study the potential and identify bottlenecks for natural regeneration and examine the effects of overstory thinning and site preparation treatments. We integrated four experiments conducted in diverse ecoregions in Israel ranging from semiarid to subhumid. Seedling emergence and survival were traced for 2–4 years in Mishmar Ha’Emek Forest (MHF) with annual rainfall of 604 mm; Shaharia Forest (SF)—435 mm; and Yatir Forest (YF)—280 mm. Effects of thinning treatments: clearcut, 100 tree, 200 tree ha−1 and control—310 tree ha−1, were examined in MHF. Effects of site preparation treatments: soil scarification and herb clipping were examined in SF. Density of emerged seedlings varied among ecoregions and was linked to average annual rainfall. Within ecoregions, emerged seedling density was higher on north-facing slopes than on south-facing slopes and was strongly related to stand characteristics (e.g., stem basal area) and seed rain. Seedling survival rate of about 20 % was recorded in MHF while no seedling survival was observed in YF and SF during the study years. Thinning treatments reduced seed rain and emerging seedling density, but increased seedling survival and growth. Recruitment 4 years after thinning was highest in the 100 tree ha−1 treatment and lowest in the control. In SF, seedling survival was limited mainly by herbaceous vegetation and was improved by soil scarification and herb clipping. Silvicultural implications are discussed.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Pinus halepensis is the most widespread pine species in the Mediterranean basin. It occurs naturally in southern Europe, North Africa and, to a much lesser extent, also in the east Mediterranean, i.e., Turkey, Syria, Lebanon, Jordan and Israel, where it usually appears as isolated populations (Barbero et al. 1998; Quezel 2000).
In Israel, native relict populations of P. halepensis are quite restricted and can be found typically in mountainous locations with distinct edaphic conditions (Rabinowitch 1985; Schiller 1985; Weinstein 1989). However, this species has been extensively used for afforestation since the 1920s, and presently, it constitutes as much as 30 % (ca. 30,000 ha, KKL 2007) of the planted forests within the Mediterranean zone of Israel. These forests were established in a variety of habitat types, some of which are clearly beyond the biogeographic range of P. halepensis as recognized today (Liphschitz and Biger 2001). The planted forests in Israel are multifunctional, designated primarily for the provision of recreation opportunities, landscape and other ecosystem services (Osem et al. 2008). Now, because many of these forests are aging, the focus of forest managers is gradually turning toward forest renewal and design of the next forest generation. The possibility of adopting forest renewal strategies based on natural regeneration processes is becoming increasingly relevant as the discussion on sustainable forest management evolves (Osem et al. 2008, 2009).
Pinus halepensis is a short-lived, drought-resistant, pioneer species (Ne’eman and Trabaud 2000). The fire-associated regeneration capacity of this species, which is an obligate seeder, has been quite extensively addressed throughout most of the Mediterranean region (Ne’eman 1997; Pausas et al. 2004; Moya et al. 2007). However, until now, planned renewal of P. halepensis forests via natural regeneration processes has been very little studied (Fernandez et al. 2008; Prevosto and Ripert 2008; Prevosto et al. 2012), and there is a lack of experience of promoting natural regeneration in mature P. halepensis stands; especially, lacking is information regarding the effects on natural regeneration of P. halepensis of stand structure, e.g., tree density and canopy cover and silvicultural treatments, e.g., overstory thinning (regeneration cut) and site preparation. Furthermore, knowledge concerning the regeneration potential of this species and its environmental determinants in the absence of fire is limited. Fire-free regeneration of P. halepensis forests is important for managing a gradual replacement of aging forest stands located in densely populated areas and under high recreational use. In such forests, stand replacing fires is highly undesirable as they pose a real threat to human welfare and are likely to result in rapid destruction of extensive forest area. Following fires, the natural regeneration of P. halepensis is known to be promoted by the co-occurrence of several factors (Ne’eman 1997; Ne’eman et al. 2004). These include massive seed release (Moya et al. 2008), improved soil nutrient availability (Henig-Sever et al. 1996, 2000), reduced granivore activity (Tasker et al. 2011) and release from overstory shading and understory competition (Daskalakou and Thanos 1996; Eshel et al. 2000). However, in the absence of fire, these fundamental factors typically fluctuate in time and space, making simplistic generalities regarding the extent of P. halepensis regeneration practically impossible (Nathan and Ne’eman 2004). The present study aimed to elucidate environmental and silvicultural factors underlying the extent of fire-free natural regeneration in P. halepensis forests under east Mediterranean conditions.
The Mediterranean zone of Israel, particularly its southern part, represents the xeric limit of the natural distribution of P. halepensis. This limitation is due to both the small amount and the short season of rainfall, which create the long, hot dry season—about 7 months—typical of the east Mediterranean. Information regarding constraints on natural regeneration under these conditions is particularly important for our understanding of P. halepensis biogeography and ecology, as well as for the sustainable management of P. halepensis forests. Furthermore, such information is becoming highly relevant within a much wider geographical context in the light of common scenarios of global climate change. It may generally be assumed that water limitations play a major role in determining P. halepensis recruitment under east Mediterranean conditions. However, P. halepensis is a shade-intolerant species (Higgins and Richardson 1998; Ne’eman et al. 2004); therefore, its recruitment is expected to be influenced also by the availability of light in the forest understory (Fyllas et al. 2008). The inter-play between water and light availability in water limited systems is far from trivial (Riegel et al. 1995; Rodriguez-Calcerrada et al. 2008; Devine and Harrington 2009). Various environmental factors, e.g. climate, soil and bedrock, topography, and silvicultural features, e.g. stand structure, thinning treatments and site preparation treatments may vary in their effects on both water and light availability for the pine recruits and, moreover, many of these factors are known to interact. The nature of these interactions may vary among habitat types (Lortie and Callaway 2006; Gomez-Aparicio et al. 2009), resulting in complex recruitment patterns.
In this work, we addressed P. halepensis regeneration in differing east Mediterranean geo-climatic regions that ranged from semiarid to subhumid. We focused on fire-free natural regeneration within mature planted P. halepensis forests including seed rain, seedling emergence and seedling survival and growth. The general aim was to contribute to the development of silvicultural practices that would promote natural regeneration.
Our specific aims were as follows:
-
i.
To study the potential and identify bottlenecks and thresholds for natural regeneration across an east Mediterranean geo-climatic range.
-
ii.
To study the effects of overstory thinning treatments on natural regeneration.
-
iii.
To study the effects of site preparation treatments on natural regeneration.
Methods
General
This work comprises several experiments conducted as part of three different research projects in distinct forest ecoregions. The experiments were all conducted in mature planted P. halepensis forests managed by the Israeli forest service (KKL) with similar goals and a unified silvicultural strategy with respect to planting techniques and thinning regimes (Osem et al. 2008). The three differing locations were representative of the wide ecological range within which P. halepensis forests thrive in the east Mediterranean region, with water availability assumed to be the overriding factor determining forest performance as well as the extent of natural regeneration.
Research areas
Experiments were conducted in three research areas:
-
i.
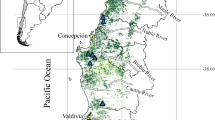
Mishmar Ha’Emek Forest (MHF) located on the northern edge of the Menashe Hills Region (Fig. 1)—average annual rainfall: 604 mm.
Fig. 1 
Location of three forest ecoregions in Israel differing in average annual rainfall: Mishmar Ha’ Emek Forest (MHF)—604 mm; Shaharia Forest (SF)—436 mm and Yatir Forest (YF)—280 mm
-
ii.
Shaharia Forest (SF) located in the Southern Judean Lowland Region (Fig. 1)—average annual rainfall: 435 mm.
-
iii.
Yatir Forest (YF) located on the boundary between the southern Judean Hills and the northern Negev Desert (Fig. 1)—average annual rainfall: 280 mm.

Detailed information regarding environmental and silvicultural characteristics in each site is presented in Table 1.
Experimental design and measurements
The research comprised four experiments:
-
(1)
Pine seedling emergence and survival and pine recruitment in a dry Mediterranean ecoregion (exp. 1). This experiment was carried out during 2008–2009 in SF. Six sites of mature planted P. halepnsis forest were selected, three on north-facing slopes and three on south-facing slopes. In each site, five randomly distributed forest plots were assigned (Altogether 30 plots). Stand characteristics—tree density, average tree stem diameter at breath height (DBH), average tree height, total stem basal area (BA)—were measured within each forest plot by counting and measuring all the planted pine trees. Additionally, measurements of leaf area index (LAI) were conducted in each plot along four evenly distributed 30 m transects using the TRAC devise (RTACS, 3rd Wave engineering, Ontario, Canada, Chen et al. 1997). Seed rain was monitored for 2 years in each plot using plastic containers as seed traps. Five traps were distributed randomly in each plot (altogether 150 traps). The traps were covered with a metal net against birds and rodents, and their sides were spread with a glue paste (Rimifoot paste®) against small granivores (arthropods). Seeds were collected and counted every 2 month. Seedling emergence and survival were monitored in 16 out of the 30 plots, eight on north-facing slopes and eight on south-facing ones, during 2 years within subplots (two subplots in each plot). Emerged seedlings were counted and marked at the end of the winter (March), and their survival was followed in monthly intervals. Five herbaceous biomass samples were taken in each plot. These samples were collected during peak standing crop (April) by clipping above ground biomass within quadrats (herb quadrats). Samples were oven-dried (65°c, 48 h) and weighed. In late spring (June), five randomly distributed soil cores (5 cm in diameter) were collected in each of the 16 plots from two depths: 0–20 cm and 20–40 cm (altogether 160 cores). A metal soil borer was used. Samples were kept in a cooler within sealed bags and taken to the lab. Soil texture, PH, P content, total N content, total and active CaCO2 content and organic matter content were measured in the lab according to Sparks et al. (1996). In 2009, a recruitment survey was conducted. Pine recruits (regenerating pines older than 1 year old) were counted in all 30 plots within the subplots (two subplots in each plot).
-
(2)
Pine seedling emergence and survival in a semiarid forest (exp. 2). This experiment was carried out during 2005–2007 in YF. Ten forest plots of mature planted P. halepensis forest were assigned representing north-facing slopes and south-facing slopes. Seedling emergence was measured during 3 years, and survival was monitored in two of the years (2005, 2007) in each plot within subplots (two subplots in each plot). Emerged seedlings were counted at the end of the winter (March), and their survival was followed in late spring (June) and the following autumn (November). Five herbaceous biomass samples were taken in each subplot in April by clipping above ground biomass within quadrates. Samples were oven-dried and weighed.
-
(3)
Effect of overstory thinning treatments on seedling emergence, survival and growth in a Mediterranean forest (exp. 3). This experiment was carried out in MHF during 1971–1975 in a mature planted P. halepensis forest. In October 1971, four thinning treatments were applied in 32 forest plots: (1) clear cut, (2) 100 tree ha−1, (3) 200 tree ha−1 and (4) control—300 tree ha−1. Trees were felled using a chain saw and dragged out by a tractor in one piece (i.e., full tree logging). The small amount of slash produced during this operation was left on the forest ground. Treatments were applied in a complete randomized design with eight replicates. Recruit density was measured prior to thinning with no significant differences observed between plots designated for the different thinning treatments. The plots were all located on north-facing slopes. Overstory canopy cover was measured in each plot by means of hemispheric photographs (fish-eye) taken at four randomly distributed locations. Overstory thinning treatments resulted in average canopy cover of 0, 37, 53 and 66 % in the clear cut, 100 tree, 200 tree ha−1 and control treatments, respectively. Seed rain was monitored in each thinning treatment using wood containers as seed traps. Twelve traps were distributed randomly within each of the thinning treatments (altogether 48 traps). The traps were covered with a metal net against birds and rodents, and their sides were spread with a glue-paste (Rimifoot paste®) against small granivores (arthropods). Seeds were collected and counted every month. Seedling emergence and survival and recruit density and growth were monitored for 4 years (1972–1975) in each plot within subplots (two subplots in each plot). Emerged seedlings were counted at the end of each winter (March), and their survival was followed in monthly intervals. Seedling growth was measured in 1 year (1972). In each plot, five randomly selected seedlings that emerged during the winter were collected in the summer (July), oven-dried and weighed. A recruitment survey was conducted during autumn 1975. Recruit (regenerating pines older than 1 year) density was measured in the subplots, and their age was estimated by counting annual shoot growth waves.
-
(4)
Effect of site preparation treatments on seedling emergence and establishment in a dry Mediterranean forest (exp. 4). This experiment was carried out in SF during 2008–2009. A randomized block experimental design was used. Ten forest plots with sparse tree density (100–120 tree ha−1) and lacking pine regeneration were selected. Five plots were located on north-facing slopes and five on south-facing ones. Within each forest plot, three blocks were placed along the slope. Each block was divided into three 2 × 2 m subplots with 1 m distance between them. The subplots were subjected to different site preparation treatments: (1) herb clipping, (2) soil scarification or (3) control. Distribution of treatments within each block was random. In each subplot, 100 pine seeds were sown (1 cm depth) before first rain (October). Herb clipping was done by cutting the herbaceous vegetation close to ground surface (1–2 cm) in early winter (8/12/2008) before pine emergence. Soil scarification was done before first rain (October) using a sharp hoe. It was applied in lines (spacing = 40 cm) perpendicular to the slope. Scarification depth was 10–20 cm. Density of emerged seedlings was measured in late winter (March), and their survival was monitored in monthly intervals.
Detailed information regarding the size and shape of forest plots, regeneration subplots, herb quadrats and seed traps in each experiment is presented in Table 2.
Statistical analyses
Regeneration parameters, i.e., seedling density, survival rate, recruit density and size were analyzed as response variables in all four experiments. Each response variable was examined for normal distribution of errors using the Shapiro–Wilk W statistic and for homogeneity of variance using Levene’s and Bartlett’s tests. No variables met the assumptions for ANOVA even when common transformations were employed. We therefore conducted analyses of variance using rank-transformed data, as outlined by Conover and Iman (1981). The Tukey–Kramer HSD test was used for post hoc comparisons. In each experiment, the response variables were analyzed with respect to different explanatory variables as following: exp. 1, slope aspect (north/south) and planted tree density; exp. 2, slope aspect; exp. 3, thinning treatments; exp 4, site preparation treatment and slope aspect. Two integrative analyses were applied on the different forest ecoregions: (1) one-way ANOVA with winter seedling density on north-facing slopes as the response variable and the forest ecoregion (MHF, SF, YF) as the explanatory variable, (2) two-way ANOVA with winter seedling density as the response variable and ecoregion (SF, YF) and slope aspect (north-facing, south-facing) as the explanatory variables. Linear regression analyses were also applied to examine the relationship between response variables and stand structure parameters (tree height, DBH, BA, LAI), soil characteristics (Ph, N, P, CaCO2, Organic matter, %clay), seed rain and herbaceous biomass. The statistical package JMP (SAS institute 1995) was used.
Results
Seedling emergence
In SF (exp. 1), it was found that density of emerged seedling in late winter (March, hereinafter—winter seedling density) was higher on north-facing slopes than on south-facing ones and increased linearly with increased stand tree density (Table 3). Regression analyses revealed that winter seedling density in SF was positively linearly related to stand BA, LAI, annual seed rain and most strongly to autumn (November-December) seed rain (Table 4a), but was not found related to any of the soil characteristics and herbaceous biomass. Site preparation treatments (scarification and clipping) applied in SF (exp. 4) had no significant effect on winter seedling density (Table 5a). In YF (exp. 2), winter seedling density was lower than in SF (Fig. 2; Table 6a). Similarly to SF, in YF, a higher winter seedling density on north-facing slopes than on south-facing slopes was found (Fig. 2; Table 6a). Winter seedling density in YF was not found related to herbaceous biomass. In MHF (exp. 3), average winter seedling density, which was measured only on north-facing slopes, was much higher than on the north-facing slopes of SF and YF (Fig. 2; Table 6b). When plotting winter seedling density against ecoregion average annual rainfall, an exponential relationship emerged (N = 3, R 2 = 0.998, p = 0.026). However, within ecoregions, trends of inter-year variations in winter seedling density did not match those of inter-year variations in rainfall amount. Overstory thinning treatments applied in MHF had a significant effect on winter seedling density which decreased with decreasing tree density (Table 7). Similarly to SF, winter seedling density in MHF was found positively linearly related to annual seed rain (N = 8, R 2 = 0.95, p < 0.0001, Table 7).

Average density of emerged pine seedlings in late winter (March) In Mishmar Ha’Emek Forest 1972–1975 (MHF), Shaharia Forest 2008–2009 (SF) and Yatir Forest 2005–2007 (YF). In MHF, south-facing slopes were not measured. Error bars indicate standard error (SE)
Seedling survival
Seedling survival rate was defined as the proportion of the seedlings that had emerged in the winter that had survived the dry season and were recorded alive in the field in the following autumn (October–November). In both YF and SF where annual rainfall amounts during the study years were below 320 mm (Table 2), seedling survival was zero. However, in MHF, where annual rainfall during the study years was 600 mm year−1 in average (Table 2), an average seedling survival of 20.2 ± 3.46 % was recorded. Overstory thinning treatments applied in MHF had a significant effect on seedling survival rate (Table 7). The highest survival was found in forest stands thinned to 100 tree ha−1; slightly lower survival was found under clear-cut stands; and the lowest survival rates were found in stands thinned to 200 trees ha−1 and in the control.
In SF, during the research years, seedling survival extended only up to late spring (June). Therefore, the seedling survival rate at mid-spring (May) was used as a measure for the spatial variation in the potential for seedling survival to be expected in years with higher rainfall. Mid-spring seedling survival was found to be positively linearly related to stand basal area, LAI and average tree height and negatively related to understory herbaceous biomass (Table 4b). It was not found related to any of the soil characteristics. Soil scarification and herb clipping applied in SF had significant positive effects on mid-spring seedling survival rate (Table 5b) and resulting mid-spring seedling density (Table 5c; Fig. 3), but no seedlings survived the summer.

The effect of site preparation treatments on pine seedling density in mid-spring (May) on south- and north-facing slopes in Shaharia Forest 2009. Different letters above columns indicate significant differences (p < 0.05). Error bars indicate standard error (SE)
Recruitment
Recruitment was defined as the density of recruits (regenerating pines older than 1 year) in the forest understory. In SF (exp. 1), in spite of the lack of any recorded seedling survival during the research years, recruitment had occurred in previous years, and recruit density varied widely among the forest plots, from 0 to 0.34 seedlings m−2. No effect of slope aspect and tree density was found. Additionally, no relationships were found by regression analyses between variations in stand characteristics (BA, height, LAI), or soil characteristics, on the one hand, and the variation in recruit density, on the other hand. Nevertheless, the spatial variation in recruit density was found to be negatively related to the spatial variation in herbaceous biomass, as recorded during the research years (Table 4c).
Recruitment as measured in MHF 4 years after overstory thinning was significantly affected by the thinning treatments (Table 7). Highest density was found in plots thinned to 100 and 200 trees ha−1, slightly lower density was in the unthinned control and lowest in the clear-cut plots. However, when the recruit-age distribution was measured, it was found that the density of 4–6-years-old recruits (seedlings that have germinated shortly before or right after the thinning) was highest in the 100-trees ha−1 plots followed by clear-cut plots, much lower in 200-trees ha−1 plots and zero in the unthinned control plots (Table 7). The effect of overstory thinning treatments on seedling growth during their first year was also assessed in MHF. Thinning treatments had a significant effect on seedling growth that increased consistently with increasing overstory canopy openness.
Discussion
The density of emerged seedlings in the forest understory differed widely among the three studied ecoregions—about 0.3 seedlings m−2 in YF, 2 seedlings m−2 in SF and 13 seedlings m−2 in MHF. These seedling densities are well within the range of about 0.2–20 seedlings m−2 reported for post-fire emergence in various P. halepensis forests throughout the east Mediterranean region (for example, Ne’eman et al. 1995; Evengalia and Daskalakou 2004 and others). Winter seedling density was strongly linked to the variation among ecoregions in average annual rainfall amount indicating water availability as an important limiting factor. The relationship between water availability and seedling emergence was also manifested in the greater winter seedling density recorded on north-facing slopes than on south-facing slopes in SF and YF. Higher water availability and vegetation productivity on north-facing slopes than on south-facing slopes is a commonly observed phenomenon in this region, as well as in other water limited environments of the northern hemisphere (Sternberg and Shoshany 2001; Pausas et al. 2004; Osem et al. 2009).
Winter seedling density was found strongly positively related to stand BA, LAI and seed rain indicating that winter seedling density is closely linked to stand condition most probably through seed production. This pattern was consistent across spatial scales, i.e., among the different forest ecoregions and also between plots within each ecoregion. Interestingly, within-site year-to-year variations in winter seedling density were not found closely related to the inter-year variations in rainfall amount. We offer two complementary explanations for this inconsistency, which are both related to seed rain: (1) yearly amount of ripen seeds (i.e., seed production) is influenced also by conditions of previous years, and (2) amount and timing of seed release from cones are determined by climatic factors other than rainfall (Natan and Ne’eman 2004). Thus, emerging seedling density is more clearly related to long-term site conditions and resulting stand characteristics than to inter-year fluctuations in rainfall amount.
It should be emphasized that, although winter seedling density was highly variable among ecoregions, the amount of emerged seedlings—about 3,000 seedlings ha−1 year−1 even in the driest region studied, YF—could still be amply sufficient for natural regeneration, if we assume that even a minor proportion of these seedlings establish every year and grow. However, our results point toward seedling establishment as a fundamental process limiting natural regeneration as discussed hereinafter.
In MHF, seedling growth and survival trough the first dry season were clearly enhanced by overstory thinning. Furthermore, the positive relationship between canopy openness and recruit density became stronger with increasing recruit age showing that the effect of thinning on recruit survival has gone beyond the first dry season following emergence. As a small-seeded shade-intolerant species (Higgins and Richardson 1998; Ne’eman et al. 2004), P. halepensis relies strongly on light availability to ensure that photosynthetic activity and growth during the humid season are sufficient to enable survival through the dry season—May–October (Jose et al. 1998). Interestingly, in MHF, recruitment was highest in the 100 tree ha−1 thinning treatment in which the seedling survival during the first dry season was even better than that in the clear-cut plots. The observed positive effect of moderate shading on seedling survival may have been related to water regime. This should be further investigated as overstory effect on understory water regime is known to be complex (Bellot et al. 2004; Molina and del-Campo 2012). The results of this work point toward 100 tree ha−1 (ca. 30 % canopy cover) as an optimum thinning level for natural regeneration in Pinus halepensis forests. However, higher resolution of thinning level and site specific adjustments may still be required.
In both YF and SF, no seedling survival was recorded during the years of the present study. This is particularly interesting in the case of SF, where dense recruitment that had occurred in previous years was observed in some of the research plots. This apparent contradiction may be attributed to year-to-year variations in rainfall. In SF, the annual rainfall during the study years 2007–2008 and 2008–2009—292 and 314 mm, respectively—was about 30 % below the average of 435 mm. Thus, we suggest that under east Mediterranean conditions natural regeneration in P. halepensis forests should not be expected in areas or years with annual rainfall below about 350 mm. This finding corresponds to the currently recognized natural distribution of P. halepensis (Barbero et al. 1998; Quezel 2000) and is also consistent with the findings of Osem et al. (2009) regarding the extent of P. halepensis recruitment along the rainfall gradient in Israel.
Although the annual rainfall limit for P. halepensis recruitment has become quite clear, variations in the level of recruitment are poorly accounted for by rainfall variations (Osem et al. 2009). Present results regarding seedling survival in SF highlight the overriding importance of competition with neighboring herbaceous vegetation in determining the ability of newly emerged seedlings to survive their first dry season and to establish themselves. This finding is consistent with that of Prevosto et al. (2012) regarding fire-free natural regeneration of P. halepensis in a west Mediterranean environment (see also, Castro et al. 2002). Competition with herbs was also shown to influence post-fire regeneration (Pausas et al. 2002). The herbaceous vegetation which, in the studied region is mostly composed of annual species, precedes the pines in germination and growth, and thereby exploits the available resources (Kolb and Robberecht 1996). Grasses (Graminae), which constitute an important fraction of this annual vegetation, were previously shown to exert, especially, high competitive pressure on pine seedlings (Hoveland and Buchanan 1973; Sheley et al. 1993). The high importance of competition with annual herbaceous vegetation is relevant to several puzzling observations concerning P. halepensis recruitment, of which the most common are poor relationships between recruitment and annual rainfall (Osem et al. 2009) and soil nutrient content (as shown in SF in the present study), positive effects of disturbances such as grazing (Osem et al. 2011), fire (Ne’eman et al. 1995), soil disturbance (Prevosto and Ripert 2008) and rock cracks (Lavi et al. 2005).
The importance of competition with neighboring understory vegetation is highly relevant with regard to formulation of silvicultural guidelines concerning forest renewal based on natural regeneration (Sternberg et al. 2001; Castro et al. 2002; Boydak 2004; Prevosto and Ripert 2008; Prevosto et al. 2012). In Mediterranean habitats with typically high abundance of annual species which can rapidly colonize available space, the window of opportunity for pine seedling establishment following overstory thinning may not last long enough unless supplementary silvicultural means are implemented. In the present study, soil scarification and to a lesser extent herb clipping improved seedling survival in SF, though this was not sufficient in the light of the rainfall limitation that prevailed during the study year. Similarly, Prevosto and Ripert (2008) demonstrated positive effects of site preparation treatments, particularly soil scarification, on P. halepensis seedling establishment in southern France. The effectiveness of soil scarification in encouraging pine seedling establishment should be largely attributed to the suppression of competing herbaceous vegetation. Nevertheless, additional factors such as seed accessibility to the mineral soil surface, water infiltration and root zone aeration may have been promoted through this silvicultural practice. In a recent study, Hibsher et al. (2013) demonstrated the positive effect of plowing on post-fire pine seedling survival during the first dry season following the fire. It may be assumed that if more rainfall had occurred during the present study, soil scarification might have substantially enhanced pine recruitment. This assumption is strengthened by the negative relationship that was found between herbaceous biomass and the extent of pine recruitment from previous years recorded in SF. Nevertheless, because in such marginal habitats, regeneration conditions may prevail only once in several years, i.e., as regeneration pulses (Savage et al. 1996), more efficient long-lasting means to control competing understory vegetation should be considered.
The present results highlight the importance of early seedling establishment, i.e., seedling survival through the dry seasons during the first few years, which represents the main bottleneck impeding fire-free natural regeneration in east Mediterranean P. halepensis forests. This is also in agreement with results obtained from studies on post-fire regeneration of P. halepensis woodlands (Evengalia and Daskalakou 2004). In this climatic region, the dry season is long and hot; therefore, pine seedling establishment is limited primarily by water availability. Water availability for the pine recruits is constrained by several factors, among which rainfall regime, which determines water input, and topography, which affects water loss are important. It is worth mentioning, however, that apart from rainfall amount and slope aspect, water availability is influenced by several additional factors, among which bedrock type, rock cover and soil depth have been shown to be relevant (Tsitsoni 1997; Pausas et al. 2004). Thus, when assessing the potential for natural regeneration and prescribing silvicultural treatments at the site level, these characteristics should also be taken into consideration. Pausas et al. (2004) has previously demonstrated the importance of water-related factors in determining the extent of post-fire regeneration of P. halepensis woodlands of the west Mediterranean. Nevertheless, in his study, rainfall amount was less important than other local and microclimatic factors. We propose that the positive effects of higher water input, i.e., increased rainfall, and/or lower water loss, e.g., north-facing slopes compared with south-facing slopes, on the water availability for pine recruits are often counterbalanced by their parallel effects on the neighboring—and competing—understory vegetation. These effects combine to create a negative feedback loop which, above a certain threshold—rainfall of 400–450 mm year−1—makes the habitat water regime weak predictor for the level of fire-free natural regeneration of P. halepensis forests. Moreover, the development of seedlings during the winter and spring, which enables sufficient root growth and, consequently, access to essential deeper water resources, is critically limited by light availability, which depends on the overstory characteristics. Although increased stand canopy cover should result in larger seed rain and, consequently, higher density of newly emerged seedlings, low to zero canopy cover, which ensures high light availability, would be favorable for pine seedling development and long-term survival. Similarly to water availability, increased light availability may also activate a negative feedback loop, through the enhancement of competing understory vegetation. The described inter-play between habitat water regime, forest overstory and understory structure and resource availability for pine recruitment is presented by a conceptual model in Fig. 4.

Interactions and feedback loops determining resource availability for recruitment in water limited Pinus halepensis forests. Full-line arrows indicate positive relationships, while broken-line arrows indicate negative relationship. Hence, opposing full and broken arrows indicate a negative feedback loop. Habitat water regime determines the availability of water, which is the main limiting factor for the forest primary production. The leaf area index (LAI) of the forest overstory (canopy) and understory represents the performance of each of these components. Higher LAI = higher primary production, water consumption and light interception and therefore stronger competitive pressure. Increased water availability is counterbalanced by increased water consumption and light interception by the forest overstory. Reduction of overstory LAI by thinning is counterbalanced by increased understory LAI. If understory growth is slow, such as in the case of typical Mediterranean woody vegetation, then a window of opportunity for pine recruitment is opened. In contrast, if understory growth is rapid, such as in the case of annual herbaceous vegetation, then the window of opportunity for natural regeneration may not be sufficient unless site preparation treatments are applied
Conclusions
In the light of our findings, we propose that management for natural regeneration in east Mediterranean P. halepensis forests should focus simultaneously on the achievement of high overstory canopy openness and low understory competition. Silvicultural practice may, therefore, vary among ecoregions as follows.
-
i.
Mediterranean—average annual rainfall more than 450 mm. Overstory thinning (regeneration cut) should leave canopy cover of 0–30 %, i.e., up to about 100 trees ha−1. This should be accompanied by short-term suppression of herbaceous vegetation, when required. Short-term herb suppression may be achieved by soil scarification applied during summer or early autumn.
-
ii.
Dry Mediterranean—annual rainfall, 350–450 mm. Natural regeneration is limited by year-to-year rainfall fluctuations. Thus, overstory thinning should be applied with emphasis on retaining seed trees (Smith 1986; Boydak 2004) to ensure continuous seed input. In such ecoregions, long-term suppression of herbaceous vegetation may also be needed, in order to guarantee regeneration success in case of a year of good rainfall.
-
iii.
Semiarid—annual rainfall less than 350 mm. Meaningful regeneration should not be expected under these conditions unless intervention involves water supplementation or rainfall redistribution, e.g., by harvesting water runoff.
Several gaps still remain in our knowledge regarding the overwhelming variability in the extent of P. halepensis natural regeneration under seemingly comparable conditions. With regard to silvicultural practice, overstory thinning and suppression of understory competing vegetation were shown to enhance pine recruitment. However, more information is required regarding fine adjustment of the overstory thinning regime under varied water conditions. Furthermore, the development of practical, sustainable means for temporary suppression of competing vegetation is required, particularly interesting are prescribed fire and grazing.
References
Barbero M, Loisel R, Quezel P, Richardson DM, Romane F (1998) Pines of the Mediterranean Basin. Ecology and biogeography of Pinus, Ed Richardson, DM., 153–170, Cambridge University Press. Cambridge
Bellot J, Maestre FT, Chirino E, Hernandez N, de Urbina JO (2004) Afforestation with Pinus halepensis reduces native shrub performance in a Mediterranean semiarid area. Acta Oecologia-Int J Ecol 25:7–15
Boydak M (2004) Silvicultural characteristics and natural regeneration of Pinus brutia ten. A review. Plant Ecol 171:153–163
Castro J, Zamora R, Hódar JA (2002) Mechanisms blocking Pinus sylvestris colonization of Mediterranean mountain meadows. J Veg Sci 13:725–731
Chen JM, Rich PM, Gower TS, Norman JM, Plumer S (1997) Leaf area index of boreal forests: theory, techniques and measurements. J Geophys Res—Atmos 102:29475–29482
Conover WJ, Iman RL (1981) Rank transformation as a bridge between parametric and nonparametric statistics. Am Stat 35:124–129
Daskalakou EN, Thanos CA (1996) Aleppo pine (Pinus halepensis) postfire regeneration: the role of canopy and soil seed banks. Int J Wildland Fire 6:59–66
Devine WD, Harrington TB (2009) Belowground competition from overstory trees influences Douglas-fir sapling morphology in thinned stands. New For 37:137–153
Eshel A, Henig-Sever N, Ne’eman G (2000) Spatial variation of seedling distribution in an east Mediterranean pine woodland at the beginning of post-fire succession. Plant Ecol 148:175–182
Evengalia ND, Daskalakou AT (2004) Post-fire regeneration of Aleppo pine—the temporal pattern of seedling recruitment. Plant Ecol 171:81–89
Fei S, Steiner KC (2008) Relationships between advance oak regeneration and biotic and abiotic factors. Tree Physiol 28:1111–1119
Fernandez C, Voiriot S, Mevy JP et al (2008) Regeneration failure of Pinus halepensis Mill: the role of autotoxicity and some abiotic environmental parameters. For Ecol Manag 255:2928–2936
Fyllas NM, Dimitrakopoulos PG, Troumbis AY (2008) Regeneration dynamics of a mixed Mediterranean pine forest in the absence of fire. For Ecol Manag 256:1552–1559
Gomez-Aparicio L, Zavala MA, Bonet FJ (2009) Are pine plantations valid tools for restoring Mediterranean forests? An assessment along abiotic and biotic gradients. Ecol Appl 19:2124–2141
Henig-Sever N, Eshel A, Neeman G (1996) pH and osmotic potential of pine ash as post-fire germination inhibitors. Physiol Plant 96:71–76
Henig-Sever N, Eshel A, Ne’eman G (2000) Regulation of the germination of Aleppo pine (Pinus halepensis) by nitrate, ammonium, and gibberellin, and its role in post-fire forest regeneration. Physiol Plant 108:390–397
Hibsher N, Moshe Y, Bney-Moshe E, Ben-Moshe E, Zangi E, Zuck A, Osem Y (2013) Post-fire regeneration in Mediterranean reforested sites as affected by mechanical site preparation: lessons for restoration. Appl Veg Sci. doi:10.1111/AVSC.12030
Higgins SI, Richardson DM (1998) Pine invasions in the southern hemisphere: modeling interactions between organisms, environment and disturbance. Plant Ecol 135:79–93
Hoveland CS, Buchanan GA (1973) Weed seed germination under simulated drought. Weed Sci 21:322–324
José MB, Riba M, Retana J (1998) Seed germination and seedling performance of two Mediterranean tree species, holm oak (Quercus ilex L.) and Aleppo pine (Pinus halepensis Mill.): a multifactor experimental approach. Plant Ecol 138:17–26
KKL (2007). Green information: forest statistics. The department of forest management and GIS, the forest department. Keren Kayemeth LeIsrael—Jewish National Fund
Kolb PF, Robberecht R (1996) Pinus ponderosa seedling establishment and the influence of competition with the bunchgrass Agropyron spicatum. Int J Plant Sci 157:509–515
Lavi A, Perevolotsky A, Kigel J, Noy-Meir I (2005) Invasion of Pinus halepensis from plantations into adjacent natural habitats. Appl Veg Sci 8:1–85
Liphschitz N, Biger G (2001) Past distribution of Aleppo pine (Pinus halepensis) in the mountains of Israel (Palestine). Holocene 11:427–436
Lortie CJ, Callaway RM (2006) Re-analysis of meta-analysis: support for the stress-gradient hypothesis. J Ecol 94:7–16
Mediterranean Basin. Backhuys Publishers, Leiden, the Netherlands, pp. 37–50
Molina AJ, del Campo AD (2012) The effects of experimental thinning on throughfall and stemflow: a contribution towards hydrology-oriented silviculture in Aleppo pine plantations. For Ecol Manag 269:206–213
Moya D, Espelta JM, Verkaik I et al (2007) Tree density and site quality influence on Pinus halepensis Mill. reproductive characteristics after large fires. Ann For Sci 64:649–656
Moya D, Saracino A, Salvatore R, Lovreglio R, de las Heras J, Leone V (2008) Anatomic basis and insulation of serotinous cones in Pinus halepensis Mill. Trees Struct Funct 22:511–519
Nathan R, Ne’eman G (2004) Spatiotemporal dynamics of recruitment in Aleppo pine (Pinus halepensis Mill.). 2nd International Conference on Mediterranean Pines, Chania. Greece Plant Ecol 171:123–137
Ne’eman G (1997) Regeneration of natural pine forest—review of work done after the 1989 fire in Mount Carmel, Israel. Int J Wildland Fire 7:295–306
Ne’eman G and Trabaud L (2000) Ecology, biogeography and management of Pinus Halepensis and P. Brutia forest ecosystems in the Mediterranean basin. 1st ed. Backhuys, Leiden, Netherland
Ne’eman G, Lahav H, Izhaki I (1995) Recovery of vegetation in a natural east Mediterranean pine forest on Mount Carmel, Israel as affected by management strategies. For Ecol Manag 75:17–26
Ne’eman G, Goubitz S, Nathan R (2004) Reproductive traits of Pinus halepensis in the light of fire—a critical review. Plant Ecol 171:69–79
Osem Y, Ginsberg P, Tauber I, Atzmon N, Perevolotsky A (2008) Sustainable management of Mediterranean planted coniferous forests: an Israeli definition. J For 106:38–46
Osem Y, Zangy E, Bney-Moshe E (2009) The potential of transforming simple structured pine plantations into mixed Mediterranean forests through natural regeneration along a rainfall gradient. For Ecol Manag 259:14–23
Osem Y, Lavi A, Rosenfeld A (2011) Colonization of Pinus halepensis in Mediterranean habitats: consequences of afforestation, grazing and fire. Biol Invas 13:485–498
Pausas JG, Ribeiro E, Vallejo R (2004) Post-fire regeneration variability of Pinus halepensis in the eastern Iberian Peninsula. For Ecol Manag 203:251–259
Prevosto B, Ripert C (2008) Regeneration of Pinus halepensis stands after partial cutting in southern France: impacts of different ground vegetation, soil and logging slash treatments. For Ecol Manag 256:2058–2064
Prevosto B, Amandier L, Quesney T, de Boisgelin G, Ripert C (2012) Regenerating mature Aleppo pine stands in fire-free conditions: site preparation treatments matter. For Ecol Manag 282:70–77
Quezel P, (2000) Taxonomy and biogeography of Mediterranean pines (Pinus halepensis and P. brutia). In: Ne’eman G, Trabaud L. (Eds.), Ecology, Biogeography and Management of Pinus halepensis and Pinus brutia Forest Ecosystems in the Mediterranean Basin, Backhuys, Leiden, Netherlands, pp. 1–12
Rabinowitch A (1985) Arboreal plant communities of the Mediterranean ecosystems in Israel. Rotem 18:5–12 in Hebrew, English abstract
Riegel GM, Miller RF, Krueger WC (1995) The effects of aboveground and belowground competition on understory species composition in a Pinus ponderosa forest. For Sci 41:864–889
Rodriguez-Calcerrada J, Pardos JA, Gil L et al (2008) Ability to avoid water stress in seedlings of two oak species is lower in a dense forest understory than in a medium canopy gap. For Ecol Manag 255:421–430
Savage M, Brown PM, Feddema J (1996) The role of climate in a pine forest regeneration pulse in the southwestern United States. Ecoscience 3:310–318
Schiller G (1985) Natural Pinus halepensis: distribution and genetic relations. Rotem 18:69–78 in Hebrew, English abstract
Sheley RL, Larson LL, Johnson DE (1993) Germination and root dynamics of range weeds and forage species. Weed Technol 7:234–237
Smith DM (1986) The Practice of Silviculture. John Wiley and Sons, New York
Sparks DL, Page AL, Helmke PA, Leoppert RH, Soltanpour PN, Tabatabai MA, Johnston CT, Sumner ME (1996) Methods of soil analysis—soil science society of America book series. ASA and SSA, Madison, WI, USA
Sternberg M, Shoshany M (2001) Influence of slope aspect on Mediterranean woody formations: comparison of a semiarid and an arid site in Israel. Ecol Res 16:335–345
Sternberg M, Danin A, Noy-Meir I (2001) Effects of clearing and herbicide treatments on coniferous seedling establishment and growth in newly planted Mediterranean forests. For Ecol Manag 148:179–184
Tasker EM, Denham AJ, Taylor E et al (2011) Post-fire seed predation: does distance to unburnt vegetation matter? Austral Ecol 36:755–766
Tsitsoni T (1997) Conditions determining natural regeneration after wildfires in the Pinus halepensis (Miller, 1768) forests of Kassandra Peninsula (North Greece). For Ecol Manag 92:199–208
Weinstein A (1989) Geographical variation and phenology of Pinus halepensis, P. brutia and P. eldarica in Israel. For Ecol Manag 27:99–108
Acknowledgments
We thank Eitan Bney-Moshe and Ezra Ben-Moshe for their great assistance in the field work. We thank the Israeli Forest Service—the KKL—for supporting the research.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by A. Weiskittel.
Rights and permissions
About this article
Cite this article
Osem, Y., Yavlovich, H., Zecharia, N. et al. Fire-free natural regeneration in water limited Pinus halepensis forests: a silvicultural approach. Eur J Forest Res 132, 679–690 (2013). https://doi.org/10.1007/s10342-013-0704-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-013-0704-3