
Abstract
The volume closes with a brief summary of highlights and pointers to further directions for the physics of biological oscillators.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others
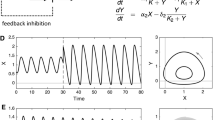
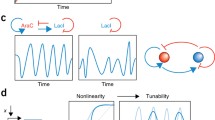
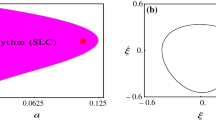
To understand living systems requires more than just traditional dynamical systems theory.
Living systems are subject to a time-varying environment, so a key first step is to use and further develop the theory of non-autonomous dynamical systems. Also, they do not last forever, so practical versions of dynamical systems concepts like limit cycles are required that make sense for long but finite time-intervals.
Secondly, a lot of the environment may not be known explicitly, so it is appropriate to model the residual effects by a stochastic process. There are many candidates, however, and it is unlikely to be enough to consider only Weiner processes. Many processes in Nature that serve as perturbations of living systems are in fact oscillatory.
Thirdly, for systems where discreteness plays a role, for example at the level of biomolecules or at the level of individuals in a population, the dynamics of the system itself may require a stochastic formulation. Alternatively, individuals in a population can be treated as oscillatory systems and analysed as networks of coupled non-autonomous oscillators.
Finally, living systems operate far from thermodynamic equilibrium. Principles are required to treat this realm of physics. The minimum entropy production rate principle of Glansdorff and Prigogine is valid only near equilibrium. Maes and Netocny [4] have come up with an extension that is valid for all Markov processes, namely that the stationary distribution maximises the Donsker-Varadhan large deviation rate function for fluctuations in the occupation probabilities, but more work is required to make this physically meaningful. See [3] for a recent survey.
Another principle, that goes back to Helmholtz, then Ashby in the days of cybernetics, and has been extended by Friston [2], is the free energy principle. It states that systems act to minimise “surprise”: the discrepancy between observations and beliefs. See the Wikipedia page for more. On the other hand, a totally opposite principle has been promoted, the future state maximisation principle, that individuals act to keep their options open as much as possible (for an application, see [1]).
Although various of the above phenomena in living organisms are describable within the realm of statistical mechanics and thermodynamics, they cannot be described in terms of the Boltzmann–Gibbs theory. This is because, in one way or another, the latter requires the microscopic dynamics to be strongly chaotic, i.e., basically mixing and ergodic, being consistently based on the usual additive entropic functional. Living and living-like organisms and systems (economics, linguistics, geophysics, astrophysics), with their subtle nature, usually evolve within weak chaos, at the edge of chaos, between strong chaos and order (typically with vanishing Lyapunov exponents). This feature usually mandates nonadditive entropic functionals, and, consequently, an entire reformulation and mesoscopic interpretation of thermodynamics itself (see, for instance, [5]).
This volume has concentrated on the physics of biological oscillations. The pervasiveness of oscillations in living systems suggest that none of the above principles will really get to the heart of how living systems function. Their dynamical nature must be taken into account. So also must the continuous exchange of energy and matter, which is the key characteristic of living systems. It occurs on all scales ranging from exchange across the cell membrane to exchanges at organ and system levels. So, conservation of matter (mass) is not an appropriate principle. It is the exchange of matter that results in the continuous sustained oscillations that are observed, with frequencies that are non-constant. Perhaps one can focus on the interactions between different parts of the system to develop new principles based on the phase differences between non-autonomous oscillatory systems, or networks of such systems. This scenario seems realistic in view of the studies that have already pointed to the stabilising effect of non-autonomicity.
References
H.J. Charlesworth, M.S. Turner, Intrinsically motivated collective motion. PNAS 116, 15362–15367 (2019)
K. Friston, The free-energy principle: a unified brain theory? Nat. Rev. Neurosci. 11, 127–138 (2010)
C. Maes, Frenesy. https://arxiv.org/abs/1904.10485
C. Maes, K. Netocny, Minimum entropy production principle from a dynamical fluctuation law. J. Math. Phys. 48, 053306 (2007)
C. Tsallis, Beyond Boltzmann-Gibbs-Shannon in physics and elsewhere. Entropy 21, 696 (2019)
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2021 Springer Nature Switzerland AG
About this chapter

Cite this chapter
MacKay, R.S., Stefanovska, A., Tsallis, C. (2021). Physics of Biological Oscillators: Outlook. In: Stefanovska, A., McClintock, P.V.E. (eds) Physics of Biological Oscillators. Understanding Complex Systems. Springer, Cham. https://doi.org/10.1007/978-3-030-59805-1_27
Download citation
DOI: https://doi.org/10.1007/978-3-030-59805-1_27
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-59804-4
Online ISBN: 978-3-030-59805-1
eBook Packages: Physics and AstronomyPhysics and Astronomy (R0)