
Abstract
Synthetic nanoparticles (NPs) have more characteristic properties like high surface area, tunable pore size, and high reactivity, as compared to natural NPs. Therefore, this technology has opened a new epoch of application-based research in plant physiology. With an increase in the use of metal and metal oxide nanoparticles (MNPs/MONPs), they will gradually enter the environment. Regulation of their optimum levels within the soil as nutrients/facilitator/pollutant for sustainable agriculture and crop production is a very challenging and tedious task. Plant development is regulated by diverse environmental factors like nutrient availability, temperature, soil morphology, and light intensity. Therefore, plant-MNP relations concerning the uptake and toxicity of MNPs/MONPs under natural environment have attracted the attention of many researchers. The unrestrained application of MNPs poses a challenge concerning their fate and transport in the natural environment and their hazardous effect on the biotic community. Knowledge about the phytotoxicity of many MNPs/MONPs on plant performance is still meager, and hence it is too early to predict their impacts on plants and on the respective ecosystem.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
The art of manipulating matter on an atomic, molecular, or supramolecular level is known as nanotechnology (Banerjee and Kole 2016). Usually, materials lesser than 100 nm in one dimension, are treated as nanomaterial. Therefore, they can be one dimensional (rod-shaped), two dimensional (films), three dimensional (any shape), or zero dimensional (all dimensions are at nanoscales), based on the modification of matter (Bernhardt et al. 2010; Tiwari et al. 2014; Banerjee and Kole 2016). Because of this, they possess some characteristic features, including the physical and chemical properties that have drawn a general attention to their pivotal application in plant sciences with reference to plant growth and development.
In general, nanoparticles (NPs) can be distinguished into three main groups, viz., natural, incidental, and engineered or manufactured NPs (Nowack and Bucheli 2007; Monica and Cremonini 2009). NPs of the first type are present since the beginning of the earth and are released through natural processes like volcanic eruptions, forest fires, dust storms, and photochemical reactions. The second form of NPs is anthropogenic in nature, which emanates usually from petrol/diesel exhaust, burning of coal, and industrial exhausts (Buzea et al. 2007). The engineered NPs (ENPs) can be categorized as carbon-based NPs (CB-NPs), metal-based NPs (MB-NPs), magnetic NPs, dendrimers, and composite NPs. The metal and metal oxide-based NPs are purposely produced by humans from different metals like gold (Au), silver (Ag), zinc (Zn), nickel (Ni), ferrum (Fe), and copper (Cu) and from metal oxides like titanium dioxide (TiO2), ferroferric oxide (Fe3O4), silicon dioxide (SiO2), cerium oxide (CeO2), aluminum oxide (Al2O3), etc. (Fedlheim and Foss 2001).
During the last two decades, a significant amount of research has been conducted on metal NPs (MNPs) and metal oxide NPS (MONPs) particularly from agricultural perspective, because these NPs can easily slip into the plant system (Tripathi et al. 2011; Husen and Siddiqi 2014a, b; Raliya et al. 2015). Because of their unique properties, NPs are reported to boost plant metabolism (Nair et al. 2011). Excessive use of NPs in the industrial sector, in food and agricultural products, and in remediation technologies has conjured the issue of contamination of ecosystems (Gardea-Torresdey et al. 2014; Nair 2016). This has dragged the attention of many research groups to explore the potential effects of NPs on plants (Monica and Cremonini 2009; Nair et al. 2011; Li et al. 2014; Rico et al. 2015). Being sessile in nature, plants are frequently exposed to NPs. Plants exposed to MNPs and MONPs subsequently accumulate them in their underground and aerial parts. When present into the rhizosphere, NPs can easily enter the epidermis and cortex via the apoplastic route (Rizwan et al. 2016). Translocation to aerial parts is much dependent on exposure time, plant species, and the shape and size of NPs (Li et al. 2014; Rico et al. 2015). On finding their entry into crop plants, NPs also generate a threat to human population through contamination of the food chain. Potential toxicity of NPs toward living organisms is well established. Given this, it becomes imperative to study the interactions among plants and NPs, which determine the mode of the NP uptake and accumulation, and subsequently their fate in the environment (Nair 2016). This chapter provides the latest information related to the interaction of plants with NPs and elucidates the consequent effects on plant growth and development.
2 Metal and Metal Oxide Nanoparticles
Being a new field, nanotechnology has a potential to provide a platform for researchers to design and incorporate new tools for studying the key functioning of NPs into the plant system (Cossins 2014). Metallic NPs are simple to synthesize because of their tunable features like size, shape, composition, structure, and encapsulation, out of different reported NPs (Subbenaik 2016). Of the synthesized NPs, Au and Ag NPs are most frequently used because of their simplicity in preparation, bio-conjugation, and appealing results under various tested systems. Limited size and high density of corner gave exclusive properties to metal oxide NPs also (Picó and Blasco 2012; Raliya and Tarafdar 2013; Subbenaik 2016). Different metals (Au, Ag, Zn, Ni, Fe, and Cu) and metal oxides (TiO2, Fe2O4, SiO2, CeO2, Al2O3, etc.) have been used to design NPs (Fedlheim and Foss 2001) that suit different plant-related processes including protection and fertilization (Gogos et al. 2012). For example, the use of SiO2 and Al2O3 nanoparticles reportedly increases the germination percentage and growth of roots in plants (Lin and Xing 2007; Siddiqui and Al-Whaibi 2014).
However, the rapidly increasing use of MNPs and MONPs in various operations and their presence in the environment has raised the issues of environmental health. Regulation of their optimum levels within the soil as nutrients/facilitators/pollutants for sustainable agriculture and crop production is a tedious task. As plant development is regulated by diverse environmental factors like nutrient availability, temperature, soil morphology, and light intensity, it is important to explore whether MNPs/MONPs also have a potential to influence the plant growth and development and/or create toxicity in the plant system.
3 Effects of NPs on Plant Growth and Development
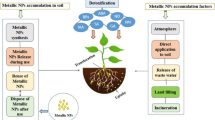
Plant growth and development is a holistic term that starts from the initial stages of seed germination and extends up to the senescence. The effects of MNPs/MONPs on the overall growth process are found to be both positive and negative, possibly depending upon the size, composition, concentration, physical and chemical properties of NPs, and also the nature of plant species (Khodakovskaya et al. 2012; Husen and Siddiqi 2014b; Nair 2016; Siddiqi and Husen 2016, 2017a, b; Husen 2017; Fig. 15.1). This chapter is planned to discuss the plausible role of NPs on overall plant growth and productivity.

Schematic summary depicting the potential impacts of metal and metal oxide nanoparticles on plant growth and productivity. These impacts are affected by size, composition, concentration, physical and chemical properties of NPs, mode of their application, and also on the nature of plant species
3.1 Seed Germination
Effects of NPs on seed germination are both positive and negative (Hong et al. 2015). Nanoparticles synthesized from lead, palladium, gold, and copper have considerably swayed the growth of lettuce (Lactuca sativa) seeds (Shah and Belozerova 2009). The activity of nitrate reductase was increased by the exogenous treatment of nano-SiO2 and nano-TiO2 which results in the better germination of soybean seeds (Lu et al. 2002). Improved seed germination was also noticed in lettuce and cucumber (Cucumis sativus) (Barrena et al. 2009), Indian mustard (Brassica juncea) (Arora et al. 2012), Boswellia ovalifoliolata (Savithramma et al. 2012), and Gloriosa superba (Gopinath et al. 2014) plants when given Au NP treatment. Similarly, application of nano-SiO2 improves seed germination and nutrient’s availability to maize plants (Suriyaprabha et al. 2012). Better germination of seeds was noticed in peanut (Arachis hypogaea) (Prasad et al. 2012), soybean (Glycine max) (Sedghi et al. 2013), wheat (Triticum aestivum) (Ramesh et al. 2014), and onion (Allium cepa) (Raskar and Laware 2014) by the application of Zn nanoparticle.
Negative impacts of NPs on seed germination have also been recorded. Rice, barley, faba bean, and turnip have shown a dose-dependent decrease when treated with Ag NPs (El-Temsah and Joner 2012; Thuesombat et al. 2014; Thiruvengadam et al. 2014). Similarly, NPs of copper oxide (CuO), nickel oxide (NiO), TiO2, iron oxide (Fe2O3), and cobaltosic oxide (Co3O4) also reduced the seed germination of lettuce, radish, and cucumber plants (Wu et al. 2014). Plants growing in soils under natural environment somehow behave differently from those in the lab or greenhouse. Seed germination varied with different soils and was less affected in rye grass, barley, and flax plants when supplied with differential doses of Fe or Ag NPs in soil as compared to water. For instance, it was less obvious in clay soil than in sandy ones (El-Temsah and Joner 2012). In another supporting study, germination of lettuce and radish was less pretentious in soil than in water when treated with Ag NPs (Gruyer et al. 2013).
It is also known that a lower dose of NPs may serve as a seed-priming agent, whereas a higher dose causes phytotoxicity in crops. Moreover, plant response varies significantly with the NPs tested and can be correlated to their dose and size (De Rosa et al. 2013). Although the positive or negative interaction between NPs and seed germination is voraciously studied in plants, the mechanism operative behind the scene is still obscure and needs a comprehensive evaluation.
3.2 Mineral Uptake
Nutrients play a significant role in the process of plant growth, and their deficiency or nonavailability may lead to devastating effects such as stunting, deformity, discoloration, distress, and even death of the plant. The toxic metal ions considerably hamper the uptake of minerals in plants (Zaheer et al. 2015; Rizwan et al. 2016). Foliar spray of Ag NPs at a differential dose range reduced the mineral uptake in tomato seedlings, which led to nutrient deficiency (Shams et al. 2013). Treatment of CeO2 and SiO2 NPs altered the nutrient supply in shoots and roots of transgenic cotton(Gossypium)plant (Le et al. 2014; Li et al. 2014). Similarly, lettuce plants were challenged for the nutrients on application of CuO NPs (Trujillo-Reyes et al. 2014). Zhao et al. (2014) found an increase in the uptake of micronutrients in cucumber plants when treated with ZnO NPs. However, 5, 10, and 20 mg L−1 doses of CuO NPs increased the content of various nutrients, like Cu, P, and S in alfalfa (Medicago sativa) shoots, and lowered the uptake of P and Fe in lettuce shoots (Hong et al. 2015). Such a decrease in mineral uptake by plants on the application of NPs might be because of the release of toxic metal ions from NPs (Dimkpa et al. 2012; Mahmoodzadeh et al. 2013). However, more research is required to know the details of the mechanism involved.
3.3 Photosynthetic Machinery
Photosynthesis is a vital process by which plants convert light energy into chemical energy that can later be used for their growth and development. Of the total solar radiation energy falling on the surface of earth, approximately 2–4% is converted by plants for their growth and development (Kirschbaum 2011). It is important to increase the efficiency of this profit-yielding process for the better growth of plants. One essential modification for increasing the efficiency of photosynthesis in plants is modifying the rubisco activity, an enzyme that catalyzes the conversion of carbon dioxide (CO2) into the biomolecules. Recently, genes of cyanobacterium Synechococcus elongatus were incorporated in tobacco (Nicotiana tabacum) plants by replacing the Rubisco gene for carbon fixation in tobacco plant (Lin et al. 2014). These new engineered plants showed more photosynthetic efficiency than the native ones. It has been reported that the application of SiO2 NPs to plants improves the photosynthetic rate by improving the activity of carbonic anhydrase enzyme (that supplies CO2 to the Rubisco) and synthesis of photosynthetic pigments (Xie et al., 2012, Siddiqui and Al-Whaibi 2014; Siddiqui et al. 2014). Similarly, the use of nano-anatase TiO2 enhances photosynthesis by stimulating the Rubisco activity that could eventually increase the growth rate of plants (Gao et al. 2006; Chand and Siddiqui, 2012). It is also noteworthy that the bulk use of MNPs/MONPs generates the oxidative burst in plants by releasing their metal ions (Rizwan et al. 2016). This oxidative stress may result in the production of ROS that might interfere with many biochemical reactions and reduce the photosynthesis and gas exchange capacity of plants (Adrees et al. 2015; Das et al. 2015). The toxic effect of NPs on photosynthesis and gas exchange in food crops has been widely studied (Mirzajani et al. 2013; Abouzeid and Moustafa 2014; Rao and Shekhawat 2014; Shaw et al. 2014).
The use of Ag NPs as seed-priming agent in wheat, soybean, and barley significantly hampered the content of photosynthetic pigments and also quenched the chlorophyll fluorescence (Zhao et al. 2013; Abouzeid and Moustafa 2014; Gorczyca et al. 2015). The excessive application of MNPs/MONPs caused a significant reduction in the total chlorophyll content of some crop plants including Indian mustard, pea, and soybean (Pradhan et al. 2013; Mukherjee et al. 2014; Rao and Shekhawat 2014). Therefore, it is quite now known that the toxic effects of MNPs/MONPs on the photosynthetic machinery depend on the duration, type, and dose of the NPs. Plants have the in-built mechanism to withstand against NP toxicity for certain period of time, and the extended exposure to NPs could hamper the photosynthetic system.
3.4 Plant Morphology
Plants flourishing in suited environment are characterized with better morphology based on the shoot and root lengths, the shoot and root biomass, and the leaf area. Stressful environment has adverse effects on these characters. Asli and Neumann (2009) reported that application of nTiO2 NPs repressed leaf growth and leaf functions in maize seedlings via affecting the water uptake. Implications of the ZnO, Fe2O3, Al2O3, and CuO NPs in modifying the morphological parameters have been assessed in various crop systems (Dimkpa et al. 2012; Mahmoodzadeh et al. 2013; Nair and Chung 2014). It is suggested that the possible reason for the negative effect on plant growth parameters is the plausible release of toxic metal ions from these manufactured NPs. NPs synthesized from Ag significantly inhibited the root growth and biomass of different tested crops such as wheat, rice, sorghum, and tomato (Mazumdar and Ahmed 2011; Dimkpa et al. 2012; Song et al. 2013; Vannini et al. 2014). Exposure of Ag NPs, at a dose of 0.2, 0.5, and 1 mg L−1 for 1 week, remarkably reduced the shoot and root biomass as well as the root elongation in rice seedlings (Nair et al. 2014). The length of wheat seedlings as well as the root growth in soybean and chickpea was noticeably hampered on application of CuO NPs (Adhikari et al. 2012; Dimkpa et al. 2012). Similarly, a dose-dependent decrease in plant height and in the shoot and root biomass of cotton seedlings was caused by the CeO2 and SiO2 NPs added to the growth media (Le et al. 2014). Additionally, occurrence of silver nanoparticles was detected in plasmodesmata and cell wall (Geisler-Lee et al. 2013), which would certainly result in wall seepage and snarl-up of intercellular communication (Geisler-Lee et al. 2014), sequentially distorting the performance of nutrient transporter proteins and intercellular transport and thereby affecting the overall growth of the plants. The anatomical and ultrastructural responses of Capsicum annuum toward Fe NPs have been studied recently by Yuan et al. (2018) who found the responses to be dose-dependent. For instance, improvement in leaf growth, chloroplast number, grana stacking, and development of vascular bundles was observed by light and electron microscopes at low concentrations of Fe NPs. In contrast, at elevated doses, Fe NPs appeared to be aggregated in cell walls and entered into the roots via the apoplastic pathway, thereby blocking the movement of iron.
In addition, many studies have shown the stimulating effects of different NPs toward plant morphology (Wang et al. 2012; Wang et al. 2013a; Antisari et al. 2015). Exposure of tomato seedlings to CeO2 NPs (0.1-10 mg L−1) caused a little increase in plant height and biomass (Wang et al. 2012). Application of nTiO2 to spinach (Spinacia oleracea) significantly improved its growth (Hong et al. 2015); treatment with ZnO at different doses (125, 250, and 500 mg−1 kg−1 soil) enhanced the root length of green peas (Pisum sativum), which was almost doubled in comparison to the control (Mukherjee et al. 2014). Treatment of TiO2 NPs improves the growth and yield of wheat plants in water-deficit condition (Jaberzadeh et al. 2013). Similarly, SiO2 NP application (at a dose of 5–20 kg ha−1) in sandy loam soil appreciably increased the shoot and root length and leaf area of 20-day-old maize seedlings (Suriyaprabha et al. 2012). Tobacco plants exposed to different concentrations (0.1%, 0.5%, and 1%) of Al2O3 NPs exhibited increase in root length and biomass but a drastic decrease in leaf count (Burklew et al. 2012; Verma et al. 2018). Likewise, physiological changes in watermelon were evaluated in vitro after exposure to γ-Fe2O3 NPs (Wang et al. 2016). An optimum dose of NPs was able to recover chlorosis and iron deficiency and promote plant growth.
Interaction between NPs and rhizosphere is immensely important because of the probable impact of NPs on the root-bacteria symbiosis. ZnO NPs proved perilous to rhizobium-legume symbiosis, as they disrupted early communication between rhizobia and the plant along with the nodule development, and subsequently delayed the onset of nitrogen fixation, an important factor in relation to plant growth and productivity (Huang et al. 2014). Additionally, the presence of NPs also brought other molecular changes in terms of hormonal imbalance in plants. For instance, CuO NP application on cotton and Bt-cotton caused momentous alterations in the intrinsic levels of indole-3-acetic acid (IAA) and abscisic acid (ABA) (Nhan Le et al. 2016). Although the MNPs/MONPs may have synergistic and antagonistic effects on the morphology of plants, their impact depends primarily on size and dose of NPs, duration of exposure, and experimental setup (Pokhrel and Dubey 2013; Thuesombat et al. 2014). It is germane to mention that most of the studies performed on NPs with respect to plant morphology have been of short duration and were conducted in controlled conditions. Therefore, for a better understanding of NP-plant interaction, long-term studies have to be carried out under natural environmental conditions.
3.5 Grain Yield and Quality
Higher yield and excellent quality of seeds signify the best growth of plants. Researchers have applied diverse nanotechnology inputs to get these desired outcomes in plants. It is now well established that plant exposure to NPs alters the uptake of nutrients and ensues biological activities, causing variation in growth and yield in different plant species. For instance, plants raised in the soil with unmodified or modified nano-TiO2 (hydrophobic or hydrophilic coating) for 65 days showed that plant growth, inorganic nutrient uptake (Ca, Mg, P, Cu, Fe, Mn, and Se), enzyme activity, chlorophyll, and carbohydrate production were augmented with coated NPs (Tan et al. 2017). Application of Ag, Fe2O3, and CeO2 NPs increased fruit yield and biomass in cucumber, soybean, and tomato plants (Sheykhbaglou et al. 2010; Wang et al. 2012; Shams et al. 2013). An increased vigor index was observed in fennel seeds exposed to TiO2 NPs, as compared to the control and bulk TiO2-treated plants (Feizi et al. 2013; Verma et al. 2018). Likewise, increased pod biomass and kernel, together with shelling percentage, was observed in peanut, when treated with ZnO NPs (Prasad et al. 2012). Additionally, NPs also affect the nutritional components and dietary value of fruits, and the impact may be trans-generational as observed in tomato plants treated with cerium oxide NPs (Wang et al. 2013b). However, some researchers have encountered contrary effects in other crops like barley (Hordeum vulgare), where exposure of CeO2 NPs prevented seed setting (Rico et al. 2015). Application of the same NP (CeO2) noticeably decreased Fe, sulfur (S), starch, and amino acid content in rice seedlings (Rico et al. 2013). Significant decrease in potassium (K) and phosphorus (P) was observed with the application of TiO2 in cucumber fruits (Servin et al. 2012). The ZnO and CeO2treatments significantly reduce overall yield of maize plants, remarkably altering the quality of corn by disrupting the mineral elements in cobs and kernels (Zhao et al. 2015). In the nutshell, application of different MNPs/MONPs affects the yield and quality of fruits and seeds, which entirely depend on the type and size of NPs and on the mode of treatment. Further illustrative studies need to be conducted to find the optimum dose of NPs. In-depth analysis in terms of long-term exposure, dose-dependent experiments, and molecular studies like proteomics and metabolics can be a handy tool in deciding the exact role of NPs on grain yield and quality.
4 Conclusion and Future Directions
Nanotechnology has evinced immense potential for the growth of agriculture sector and hence used excessively. However, bulk production and inadvertent discharge of NPs into the environment have dragged attention toward the contamination of ecosystems and food supply (Medina-Velo et al. 2016). Being nano in dimension, NPs can easily be inserted into the plant system and translocated to different organs of the plant. Therefore, it becomes imperative to evaluate their interaction on the plant system, irrespective of benefits or hazards. Furthermore, the kinetic studies have revealed the rapid and highly reactive nature of NPs and showed them to be inherently interactive with impurities (Subbenaik 2016). Recently, phytotoxicity and beneficial aspects of these xenobiotic compounds in the plant system have been discussed by several research groups (Sheikh Mohamed and Sakthi Kumar 2016). But the exact picture of their roles and interaction with plants is still blurred. Comprehensive evidence of the toxicity/benefits of different MNPs/MONPs has been presented and discussed in this chapter (Fig. 15.1). It is clear that, from their entry to accumulation, NPs depend upon the plant species, growth stage, NP size, and the mode of treatment (Kole et al. 2013; Raliya et al. 2015).
It is germane to mention that the presence of NPs in plants also poses a threat of human exposure to NPs through the food chain. Therefore, the effects of NPs in plants need to be evaluated from a wider perspective and over several generations of plants to get the better insights on their fate in the ecosystem. A comprehensive research should also be conducted at the molecular level to assess the precise roles of NPs in plants, which could be used as a platform for designing the future research in the field of nano-agriculture.
References
Abou-Zeid HM, Moustafa Y (2014) Physiological and cytogenetic responses of wheat and barley to silver nanopriming treatment. Int J Appl Biol PharmTechnol 5:265–278
Adhikari T, Kundu S, Biswas AK, Tarafdar JC, Rao AS (2012) Effect of copper oxide nanoparticle on seed germination of selected crops. J Agric Sci Technol 2:815–823
Adrees M, Ali S, Rizwan M, Ibrahim M, Abbas F, Farid M, Rehman MZ, Irshad MK, Bharwana SA (2015) The effect of excess copper on growth and physiology of important food crops: a review. Environ Sci Pollut Res 22:8148–8162
Antisari LV, Carbone S, Gatti A, Vianello G, Nannipieri P (2015) Uptake and translocation of metals and nutrients in tomato grown in soil polluted with metal oxide (CeO2, Fe3O4, SnO2, TiO2) or metallic (Ag, Co Ni) engineered nanoparticles. Environ Sci Pollut Res 22(3):1841–1853
Arora S, Sharma P, Kumar S, Nayan R, Khanna PK, Zaidi MGH (2012) Gold-nanoparticle induced enhancement in growth and seed yield of Brassica juncea. Plant Growth Regul 66:303–310
Asli S, Neumann PM (2009) Colloidal suspensions of clay or titanium dioxide nanoparticles can inhibit leaf growth and transpiration via physical effects on root water transport. Plant Cell Environ 32:577–584
Banerjee J,Kole C (2016) Plant nanotechnology: an overview on concepts strategies and tools. Chittaranjan, Kole, D. Sakthi Kumar and Mariya V. Khodakovskaya, Plant Nanotechnology- principals and practices, Springer International Publishing Switzerland, pp 1–14
Barrena R, Casals E, Colon J, Font X, Sanchez A, Puntes V (2009) Evaluation of the ecotoxicity of model nanoparticles. Chemosphere 75:850–857
Bernhardt ES, Colman BP, Hochella JMF, Cardinale BJ, Nisbet RM, Richardson CJ, Yin L (2010) An ecological perspective on nanomaterial impacts in the environment. J Environ Qual 39:1954–1965
Burklew CE, Ashlock J, Winfrey WB, Zhang B (2012) Effects of aluminum oxide nanoparticles on the growth, development, and microRNA expression of tobacco (Nicotiana tabacum). PLoS One 7:e34783
Buzea C, Pacheco II, Robbie K (2007) Nanomaterials and nanoparticles: sources and toxicity. Biointerphases 2:Mr17–Mr71
Chand N, Siddiqui N (2012) Improvement in thermo mechanical and optical properties of in situ synthesized PMMA/TiO2 nanocomposite. Compos Interfaces 19:51–58
Cossins D (2014) Next generation: nanoparticles augment plant functions. The incorporation of synthetic nanoparticles into plants can enhance photosynthesis and transform leaves into biochemical sensors. The Scientist, News & Opinion. http://www.the-scientist.com/?articles.view/articleNo/39440/title/Next-Generation–Nanoparticles-Augment-Plant-Functions/
Das S, Wolfson BP, Tetard L, Tharkur J, Bazata J, Santra S (2015) Effect of N-acetyl cysteine coated CdS:Mn/ZnS quantum dots on seed germination and seedling growth of snow pea (Pisum sativum L.): imaging and spectroscopic studies. Environ Sci Nano 2(2):203–212
De Rosa G, Lopez-Moreno ML, De Haro D, Botez CE, Peralta-Videa JR, Gardea-Torresdey J (2013) Effects of ZnO nanoparticles in alfalfa, tomato, and cucumber at the germination stage: root development and X-ray absorption spectroscopy studies. Pure Appl Chem 85(12):2161–2174
Dimkpa CO, McLean JE, Latta DE, Manangon E, Britt DW, Johnson WP et al (2012) CuO and ZnO nanoparticles: phytotoxicity, metal speciation, and induction of oxidative stress in sand-grown wheat. J Nanopart Res 14:1–15
El-Temsah YS, Joner EJ (2012) Impact of Fe and Ag nanoparticles on seed germination and differences in bioavailability during exposure in aqueous suspension and soil. Environ Toxicol 27:42–49
Feizi H, Kamali M, Jafari L, Rezvani MP (2013) Phytotoxicity and stimulatory impacts of nanosized and bulk titanium dioxide on fennel (Foeniculum vulgare Mill). Chemosphere 91:506–511
Fedlheim DL, Foss CA (2001) Metal Nanoparticles: Synthesis, Characterization, and Applications. CRC Press, Boca Raton, FL, USA
Gao FQ, Hong FH, Liu C, Zheng L, Su MY, Wu X, Yang F, Wu Yang P (2006) Mechanism of nano-anatase TiO2 on promoting photosynthetic carbon reaction of spinach–including complex of Rubisco-Rubisco activase. Biol Trace Elem Res 111:239–253
Gardea-Torresdey JL, Rico CM, White JC (2014) Trophic transfer, transformation, and impact of engineered nanomaterials in terrestrial environments. Environ Sci Technol 48:2526–2540
Geisler-Lee J, Wang Q, Yao Y, Zhang W, Geisler M, Li K, Huang Y, Chen Y, Kolmakov A, Ma X (2013) Phytotoxicity, accumulation and transport of silver nanoparticles by Arabidopsis thaliana. Nanotoxicology 7:323–337
Geisler-Lee J, Brooks M, Gerfen J, Wang Q, Fotis C, Sparer A, Ma X, Berg R, Geisler M (2014) Reproductive toxicity and life history study of silver nanoparticle effect, uptake and transport in Arabidopsis thaliana. Nano 4:301–318
Gogos A, Knauer K, Bucheli TD (2012) Nanomaterials in plant protection and fertilization: current state, foreseen applications, and research priorities. J Agric Food Chem 60:9781–9792
Gopinath K, Karthika V, Gowri S, Senthil Kumar V, Kumaresan S, Arumugam A (2014) Antibacterial activity of ruthenium nanoparticles synthesized using Gloriosa superba L. leaf extract. J Nanostruct Chem 4:83
Gorczyca A, Pociecha E, Kasprowicz M, Niemiec M (2015) Effect of nanosilver in wheat seedlings and Fusarium culmorum culture systems. Eur J Plant Pathol 142(2):251–261
Gruyer N, Dorais M, Bastien C, Dassylva N, Triffault-Bouchet G (2013) Interaction between silver nanoparticles and plant growth. In: International symposium on new technologies for environment control, energy-saving and crop production in green house and Plant 1037:795–800
Hong J, Rico CM, Zhao L, Adeleye AS, Keller AA, Videa JRP, Gardea-Torresdey JL (2015) Toxic effects of copper-based nanoparticles or compounds to lettuce (Lactuca sativa) and alfalfa (Medicago sativa). Environ Sci Processes Impacts 17:177–185
Huang YC, Fan R, Grusak MA, Sherrier JD, Huang CP (2014) Effects of nano-ZnO on the agronomically relevant Rhizobium-legume symbiosis. Sci Total Environ 466-467:503–512
Husen A, Siddiqi KS (2014a) Carbon and fullerene nanomaterials in plant system. J Nanobiotechnol 12:16
Husen A, Siddiqi KS (2014b) Phytosynthesis of nanoparticles: concept, controversy and application. Nano Res Lett 9:229
Husen A (2017) Gold nanoparticles from plant system: synthesis, characterization and application. In: Ghorbanpourn M, Manika K, Varma A (eds) Nanoscience and plant–soil systems, vol 48. Springer International Publishing AG, Cham, Switzerland, pp 455–479
Jaberzadeh A, Moaveni P, Moghadam HRT, Zahedi H (2013) Influence of bulk and nanoparticles titanium foliar application on some agronomic traits, seed gluten and starch contents of wheat subjected to water deficit stress. Notulae Bot Hort Agrobo 41:201–207
Khodakovskaya M, Dervishi E, Mahmood M, Xu Y, Li ZR, Watanabe F, Biris AS (2012) Carbon nanotubes are able to penetrate plant seed coat and dramatically affect seed germination and plant growth (retraction of vol 3, pg 3221, 2009). ACS Nano 6:7541–7541
Kirschbaum MUF (2011) Does enhanced photosynthesis enhance growth? Lessons learned from CO2 enrichment studies. Plant Physiol 155:117–124
Kole C, Kole P, Randunu KM, Choudhary P, Podila R, Ke PC, Rao AM, Marcus RK (2013) Nanobiotechnology can boost crop production and quality: first evidence from increased plant biomass, fruit yield and phytomedicine content in bitter melon (Momordica charantia). BMC Biotechnol 13:37
Le VN, Rui Y, Gui X, Li X, Liu S, Han Y (2014) Uptake, transport, distribution and bio-effects of SiO2 nanoparticles in Bt-transgenic cotton. J Nanobiotechnol 12:50
Li X, Gui X, Rui Y, Ji W, Yu Z, Peng S (2014) Bt-transgenic cotton is more sensitive to CeO2 nanoparticles than its parental non-transgenic cotton. J Hazard Mater 274:173–180
Lin D, Xing B (2007) Phytotoxicity of nanoparticles: inhibition of seed germination and root growth. Environ Pollut 150(2):243–250
Lin MT, Occhialini A, Andralojc PJ, Parry MAJ, Hanson MR (2014) A faster Rubisco with potential to increase photosynthesis in crops. Nature 513:547–550
Lu CM, Zhang CY, Wen JQ, Wu GR, Tao MX (2002) Research on the effect of nanometer materials on germination and growth enhancement of Glycine max and its mechanism. Soybean Sci 21:68–172
Mahmoodzadeh H, Aghili R, Nabavi M (2013) Physiological effects of TiO2 nanoparticles on wheat (Triticum aestivum). Tech J Eng Appl Sci 3:1365–1370
Mazumdar H, Ahmed GU (2011) Phytotoxicity effect of silver nanoparticles on Oryza sativa. Int J Chem Technol Res 3:1494–1500
Medina-Velo IA, Zuverza-Mena N, Tan W, Hernandez-Viezcas JA, Peralta-Videa JR, Gardea-Torresdey JL (2016) Biophysical methods of detection and quantification of uptake, translocation, and accumulation of nanoparticles. In: Chittaranjan K, Sakthi Kumar D, Khodakovskaya MV (eds) Plant nanotechnology- principals and practices. Springer International Publishing, Switzerland, pp 29–64. https://doi.org/10.1007/978-3-319-42154-4
Mirzajani F, Askari H, Hamzelou S, Farzaneh M, Ghassempour A (2013) Effect of silver nanoparticles on Oryza sativa L. and its rhizosphere bacteria. Ecotoxicol Environ Saf 88:48–54
Monica RC, Cremonini R (2009) Nanoparticles and higher plants. Caryologia Int J Cytol Cytosyst Cytogenet 62(2):161–165
Mukherjee A, Pokhrel S, Bandyopadhyay S, Mädler L, Peralta-Videa JR, Gardea-Torresdey JL (2014) A soil mediated phyto-toxicological study of iron doped zinc oxide nanoparticles (Fe@ ZnO) in green peas (Pisum sativum L.). Chem Eng J 258:394–401
Nair PMG, Chung IM (2014) Impact of copper oxide nanoparticles exposure on Arabidopsis thaliana growth, root system development, root lignification, and molecular level changes. Environ Sci Pollut Res 21(22):12709–12722
Nair PMG, Kim SH, Chung IM (2014) Copper oxide nanoparticle toxicity in mung bean (Vigna radiata L.) seedlings: physiological and molecular level responses of in vitro grown plants. Acta Physiol Plant 36:2947–2958
Nair R (2016) Effects of nanoparticles on plant growth and development. In: Chittaranjan K, Sakthi Kumar D, Khodakovskaya MV (eds) Plant Nanotechnology- principals and practices. Springer International Publishing, Switzerland, pp 95–118
Nair R, Poulose AC, Nagaoka Y, Yoshida Y, Maekawa T, Kumar DS (2011) Uptake of FITC labeled silica nanoparticles and quantum dots by rice seedlings: effects on seed germination and their potential as biolabels for plants. J Fluoresc 21:2057–2068
Nhan LV, Ma C, Shang J, Rui Y, Liu S, Xing B (2016) Effects of CuO nanoparticles on insecticidal activity and phytotoxicity in conventional and transgenic cotton. Chemosphere 144:661–670
Nowack B, Bucheli TD (2007) Occurrence, behavior and effects of nanoparticles in the environment. Environ Pollut 150:5–22
Picó Y, Blasco C (2012) Nanomaterials in food, which way forward? Analys Risk Nanomater Environ Food Samp 59:305
Pokhrel LR, Dubey B (2013) Evaluation of developmental responses of two crop plants exposed to silver and zinc oxide nanoparticles. Sci Total Environ 452:321–332
Pradhan S, Patra P, Das S, Chandra S, Mitra S, Dey KK, Goswami A (2013) Photochemical modulation of biosafe manganese nanoparticles on Vigna radiata: a detailed molecular biochemical, and biophysical study. Environ Sci Technol 47:13122–13131
Prasad TNVK, Sudhakar P, Sreenivasulu Y, Latha P, Munaswamy V, Reddy KR, Sreeprasad TS, Sajanlal PR, Pradeep T (2012) Effect of nanoscale zinc oxide particles on the germination, growth and yield of peanut. J Plant Nutr 35:905–927
Raliya R, Nair R, Chavalmane S, Wang W-N, Biswas P (2015) Mechanistic evaluation of translocation and physiological impact of titanium dioxide and zinc oxide nanoparticles on the tomato (Solanum lycopersicum L.) plant. Metallomics 7:1584–1594
Raliya R, Tarafdar J (2013) ZnO nanoparticle biosynthesis and its effect on phosphorous-mobilizing enzyme secretion and gum contents in Cluster bean (Cyamopsis tetragonoloba L.). Agric Res 2:48–57
Ramesh M, Palanisamy K, Babu K, Sharma NK (2014) Effects of bulk and nano-titanium dioxide and zinc oxide on physio-morphological changes in Triticum aestivum L. J Glob Biosci 3:415
Rao S, Shekhawat GS (2014) Toxicity of ZnO engineered nanoparticles and evaluation of their effect on growth, metabolism and tissue specific accumulation in Brassica juncea. J Environ Chem Eng 2:105–114
Raskar SV, Laware SL (2014) Effect of zinc oxide nanoparticles on cytology and seed germination in onion. Int J Curr Microbiol App Sci 3:467–473
Rico CM, Barrios AC, Tan W, Rubenecia R, Lee SC, Ramirez AV (2015) Physiological and biochemical response of soil-grown barley (Hordeum vulgare L.) to cerium oxide nanoparticles. Environ Sci Pollut Res 22(1):10551–10558
Rico CM, Morales MI, Barrios AC, McCreary R, Hong J, Lee WY, Nunez J, Peralta-Videa JR, Gardea-Torresdey JL (2013) Effect of cerium oxide nanoparticles on the quality of rice (Oryza sativa L.) grains. J Agric Food Chem 67:11278–11285
Rizwan M, Ali S, Adrees M, Rizvi H, Rehman MZ, Hannan F, Qayyum MF, Hafeez F, Ok YS (2016) Cadmium stress in rice: toxic effects, tolerance mechanisms and management: a critical review. Environ Sci Pollut Res 23(18):17859–17879
Savithramma N, Ankanna S, Bhumi G (2012) Effect of nanoparticles on seed germination and seedling growth of Boswellia ovalifoliolata – an endemic and endangered medicinal tree taxon. Nano Vis 2:61–68
Sedghi M, Hadi M, Toluie SG (2013) Effect of nano zinc oxide on the germination of soybean seeds under drought stress. Ann West Univ Timisoara 16(2):73–78
Servin AD, Michel HC, Viezcas JAH, Diaz BC, Videa JRP, Torresdey JLG (2012) Synchrotron micro-XRF and micro-XANES confirmation of the uptake and translocation of TiO(2) nanoparticles in cucumber (Cucumis sativus) plants. Environ Sci Technol 46:7637–7643
Shah V, Belozerova I (2009) Influence of metal nanoparticles on the soil microbial community and germination of lettuce seeds. Water Air Soil Pollut 197:143–148
Shams G, Ranjbar M, Amiri AA, Khodarahmpour Z (2013) The effect of 35 nm silver nanoparticles on antagonistic and synergistic mineral elements in leaves and fruit of tomato (Lycopersicon esculentum Mill.). Int J Agric Crop Sci 5:439–500
Shaw AK, Ghosh S, Kalaji HM, Bosa K, Brestic M, Zivcak M, Hossain Z (2014) Nano-CuO stress induced modulation of antioxidative defense and photosynthetic performance of Syrian barley (Hordeum vulgare L.). Environ Exp Bot 102:37–47
Sheikh Mohamed M, Sakthi Kumar D (2016) Methods of using nanoparticles. In: Chittaranjan K, Sakthi Kumar D, Khodakovskaya MV (eds) Plant Nanotechnology: Principles and Practices. Springer International Publishing, Cham
Sheykhbaglou R, Sedghi M, Shishevan MT, Sharifi RS (2010) Effects of nano-iron oxide particles on agronomic traits of soybean. Notulae Scientia Biologicae 2:112–113
Siddiqui MH, Al-Whaibi MH (2014) Role of nano-SiO2 in germination of tomato (Lycopersicum esculentum Mill.) seeds. Saud J Biol Sci 21:13–17
Siddiqui MH, Al-Whaibi MH, Faisal M, Al Sahli AA (2014) Nano-silicon dioxide mitigates the adverse effects of salt stress on Cucurbita pepo L. Environ Toxicol Chem 33:2429–2437
Siddiqi KS, Husen A (2016) Engineered gold nanoparticles and plant adaptation potential. Nano Res Lett 11:400
Siddiqi KS, Husen A (2017a) Recent advances in plant-mediated engineered gold nanoparticles and their application in biological system. J Trace Elem Med Biol 40:10–23
Siddiqi KS, Husen A (2017b) Plant response to engineered metal oxide nanoparticles. Nano Res Lett 12:92
Song U, Jun H, Waldman B, Roh J, Kim Y, Yi J, Lee EJ (2013) Functional analysis of nanoparticle toxicity: a comparative study of the effects of TiO2 and Ag on tomatoes (Lycopersicon esculentum). Ecotoxicol Environ Saf 93:60–67
Subbenaik SC (2016) Physical and chemical nature of nanoparticles. In: Chittaranjan K, Sakthi Kumar D, Khodakovskaya MV (eds) Plant nanotechnology- principals and practices. Springer International Publishing, Switzerland, pp 15–28
Suriyaprabha R, Karunakaran G, Yuvakkumar R, Rajendran V, Kannan N (2012) Silica nanoparticles for increased silica availability in maize (Zea mays L.) seeds under hydroponic conditions. Curr Nanosci 8:902–908
Tan W, Du W, Barrios AC, Jr AR, Zuverzamena N, Ji Z, Chang CH, Zink JI, Hernandezviezcas JA, Peraltavidea JR (2017) Surface coating changes the physiological and biochemical impacts of nano-TiO2 in basil (Ocimum basilicum) plants. Environ Pollut 222:64–72
Thiruvengadam M, Gurunathan S, Chung IM (2014) Physiological, metabolic, and transcriptional effects of biologically-synthesized silver nanoparticles in turnip (Brassica rapa ssp. rapa L.). Protoplasma 252(4):1031–1046
Thuesombat P, Hannongbua S, Akasit S, Chadchawan S (2014) Effect of silver nanoparticles on rice (Oryza sativa L cv. KDML 105) seed germination and seedling growth. Ecotoxicol Environ Saf 104:302–309
Tiwari DK, Dasgupta-Schubert N, Cendejas LMV, Villegas J, Montoya LC, Garcia SEB (2014) Interfacing carbon nanotubes (CNT) with plants: enhancement of growth, water and ionic nutrient uptake in maize (Zea mays) and implications for nanoagriculture. Appl Nanosci 4:577–591
Tripathi S, Sonkar SK, Sarkar S (2011) Growth stimulation of gram (Cicer arietinum) plant by water soluble carbon nanotubes. Nanoscale 3:1176–1181
Trujillo-Reyes J, Majumdar S, Botez CE, Videa JRP, Gardea-Torresdey JL (2014) Exposure studies of core–shell Fe/Fe3O4 and Cu/CuO NPs to lettuce (Lactuca sativa) plants: are they a potential physiological and nutritional hazard. J Hazard Mater 267:255–263
Vannini C, Domingo G, Onelli E, De Mattia F, Bruni I, Marsoni M, Bracale M (2014) Phytotoxic and genotoxic effects of silver nanoparticles exposure on germinating wheat seedlings. J Plant Physiol 171:1142–1148
Verma SK, Das AK, Patel MK, Shah A, Kumar V, Gantait S (2018) Engineered nanomaterials for plant growth and development: a perspective analysis. Sci Total Environ 630:1413–1435
Wang P, Menzies NW, Lombi E, McKenna BA, Johannessen B, Glover CJ et al (2013a) Fate of ZnO nanoparticles in soils and cowpea (Vigna unguiculata). Environ Sci Technol 47:13822–13830
Wang Q, Ebbs SD, Chen Y, Ma X (2013b) Trans-generational impact of cerium oxide nanoparticles on tomato plants. Metallomics 5:753–759
Wang Q, Ma X, Zhang W, Pei H, Chen Y (2012) The impact of cerium oxide nanoparticles on tomato (Solanum lycopersicum L.) and its implications for food safety. Metallomics 4:1105–1112
Wang Z, Xu L, Zhao J, Wang X, White JC, Xing B (2016) CuO nanoparticle interaction with Arabidopsis thaliana: toxicity, parent-progeny transfer, and gene expression. Environ Sci Technol 50:6008
Wu SG, Huang L, Head J, Ball M, Tang YJ, Chen DR (2014) Electrospray facilitates the germination of plant seeds. Aerosol Air Qual Res 14:632–641
Xie Y, Li B, Zhang Q, Zhang C (2012) Effects of nano-silicon dioxide on photosynthetic fluorescence characteristics of Indocalamus barbatus McClure. J Nanjing For Univ (Natl Sci Ed) 2:59
Yuan J, Chen Y, Li H, Lu J, Zhao H, Liu M, Nechitaylo GS, Glushchenko NN (2018) New insights into the cellular responses to iron nanoparticles in Capsicum annuum. Sci Rep 8:3228
Zaheer IE, Ali S, Rizwan M, Farid M, Shakoor MB, Gill RA, Najeeb U, Iqbal N, Ahmad R (2015) Citric acid assisted phytoremediation of copper by Brassica napus L. Ecotoxicol Environ Saf 120:310–317
Zhao L, Hernandez-Viezcas JA, Videa JRP, Bandyopadhyay S, Peng B, Munoz B (2013) ZnO nanoparticle fate in soil and zinc bioaccumulation in corn plants (Zea mays) influenced by alginate. Environ Sci Processes Impacts 15:260–266
Zhao L, Peralta-Videa JR, Rico CM, Hernandez-Viezcas JA, Sun Y, Niu G, Servin A, Nunez JE, Duarte-Gardea M, Gardea-Torresdey JL (2014) CeO2 and ZnO nanoparticles change the nutritional qualities of cucumber (Cucumis sativus). J Agric Food Chem 62(13):2752–2759
Zhao L, Sun Y, Viezcas JRP, Hong J, Majumdar S (2015) Monitoring the environmental effects of CeO2 and ZnO nanoparticles through the life cycle of corn (Zea mays) plants and in situ -XRF mapping of nutrients in kernels. Environ Sci Technol 49(5):2921–2928
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2019 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Kanwar, M.K., Sun, S., Chu, X., Zhou, J. (2019). Impacts of Metal and Metal Oxide Nanoparticles on Plant Growth and Productivity. In: Husen, A., Iqbal, M. (eds) Nanomaterials and Plant Potential. Springer, Cham. https://doi.org/10.1007/978-3-030-05569-1_15
Download citation
DOI: https://doi.org/10.1007/978-3-030-05569-1_15
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-05568-4
Online ISBN: 978-3-030-05569-1
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)