
Abstract
Oxidative stress is a pathological process related to not only animal kingdom but also plants. Regarding oxidative stress in plants, heavy metals are frequently discussed as causative stimuli with relevance to ecology. Because heavy metals have broad technological importance, they can easily contaminate the environment. Much of previous effort regarding the harmful impact of the heavy metals was given to their toxicology in the animals and humans. Their implication in plant pathogeneses is less known and remains underestimated.
The current paper summarizes basic facts about heavy metals, their distribution in soil, mobility, accumulation by plants, and initiation of oxidative stress including the decline in basal metabolism. The both actual and frontier studies in the field are summarized and discussed. The major pathophysiological pathways are introduced as well and link between heavy metals toxicity and their ability to initiate an oxidative damage is provided. Mobility and bioaccessibility of the metals is also considered as key factors in their impact on oxidative stress development in the plant. The metals like lead, mercury, copper, cadmium, iron, zinc, nickel, vanadium are depicted in the text.
Heavy metals appear to be significant contributors to pathological processes in the plants and oxidative stress is probably an important contributor to the effect. The most sensitive plant species are enlisted and discussed in this review. The facts presented here outline next effort to investigate pathological processes in the plants.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
In the nature, heavy metals induce serious contamination because of their persistence, high toxicity, and easy transmission through the food chain. It needs to be emphasized that there is no agreement as to what metals cannot be considered heavy. Currently, metals with molecular weight over 20 g/mol and/or density higher than 5 g/mL are designated as heavy metals (Duffus 2002, 2003; Stankovic et al. 2014; Kim et al. 2015).
Cd, Hg, Pb, and Tl are typical metals that meet the definition and fall into the group of heavy metals because of their density and molecular weight. These metals can be found in the nature; however, they have no biological role in animals nor plants. Other metals such as Co, Cu, Fe, Mn, and Zn also meet the definition of heavy metals and they are necessary for keeping homeostasis until their level does not exceed threshold level for toxicity (Van Bussel et al. 2014). Some of the metals are also typical radioisotopes (Pu and U, for instance), hence this fact should be taken into consideration and there should be considered whether the toxicity is mediated by the metal properties or there is a pathological process caused by exposure to ionizing radiation.
Harmful effect of heavy metals is not an easy effect based on an interruption of a single pathway or interaction with a single molecule. On the contrary, heavy metals have impact on multiple processes in the body, resulting in pathological consequences. In humans, association between heavy metals and some neurodegenerative disorders like Alzheimer disease, kidney damage, and cancer are discussed (Pohanka 2014a, b). Apart from the pathologies, heavy metals can be stored in cells. Both plants and microbes have high potential to accumulate heavy metals, which is useful in bioremediation (Wang and Sun 2013; Topolska et al. 2014; Hechmi et al. 2015). On the other hand, heavy metals stored in plants may have toxic effect on animal and human organisms when taken through the food chain.
This review is focused on summarizing the role of heavy metals in plants, pathology mechanisms, and environmental risks caused by the metals. The motivation for writing this paper was triggered by the fact that despite significant recent progress in the research of heavy metals, the issue has not been extensively reviewed. A complex overview of the issue is given in this manuscript, and the impact of heavy metal toxicity, accumulation in plants, chemical assays and the link between heavy metals and oxidative stress in plants are also discussed here.
2 Mobility of Heavy Metals
Metal mobility is influenced by several factors and soil properties such as the content of organic matter, oxides as well as soil structure and profile development (Mehes-Smith et al. 2013a). Transport of metals may be enhanced by several factors including metal association with mobile colloidal size and formation of metal organic and inorganic complexes that do not sorb to soil solid surfaces (Puls et al. 1991). According to Mehes-Smith et al. (2013a), soil topography plays a key role in metal distribution and horizontal mobility. Metals are mobilized by being captured by root cells from soil particles, bound by the cell wall and then transferred across the plasma membrane, driven by ATP-dependent proton pumps that catalyze H+ extrusion across the membrane (Singh et al. 2003). On binding, they displace cations such as Ca2+ and Mg2+ from the cell walls and membranes (Brunner et al. 2008; Flouty and Khalaf 2015). Binding studies with pectins have demonstrated that the binding preferences are: Al3+ > Cu2+ > Pb2+ > Zn2+ = Ca2+ or Cu2+ = Pb2+ > Cd2+ = Zn2 > Ca2+ (Franco et al. 2004), Pb2+ >> Cu2+ > Co2+ > Ni2+ >> Zn2+ > Cd2+ (Kartel et al. 1999) or Pb2+ = Cd2+ (Debbaudt et al. 2004), depending on the origins of the pectin. Pectins are a family of heterogeneous polysaccharides present in the primary cell wall and in the middle lamella of plant tissue (Guo et al. 2015). In enriched heavy metal environments, some plants will elevate the capacity of their cell wall to bind metals by increasing polysaccharides, such as pectins (Colzi et al. 2011). However, the binding capacity can be altered also for other reasons. The main functional groups of pectin are: hydroxyl, carboxyl, amide, and methoxyl, and these have been traditionally associated with heavy metal binding, especially carboxyl groups (which enable the binding of divalent and trivalent heavy metals ions) with great biosorption and heavy metal removal potential (Mata et al. 2009). The quantitative data on adsorption performance of pectins suggest their applicability as food additives or remedies for efficient removal of Pb2+, Cu2+, Co2+, and Ni2+ ions from different biological systems, including human and animal organisms (Kartel et al. 1999).
3 Bioavailability and Bioaccessibility
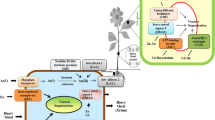
The terms bioaccessibility and bioavailability mean that the chemicals (metals) can interact with the organism and became available in the parts of organism and can take an effect. Assessment of the levels of metal bioavailability and bioaccessibility is critical in understanding the possible effect on soil biota (Ettler et al. 2012). In general, the availability of metals for plants depends on soil pH and on organic matter contents (De Matos et al. 2001; Stankovic et al. 2014), yet there are no standardized protocols for estimating the bioavailable metal content in soil. However, root-colonizing bacteria and mycorrhiza can significantly increase the bioavailability of various heavy metal ions for uptake (Singh et al. 2003). The mobility, bioavailability, and potential toxicity of a metal in the soil depend on its concentration in a soil solution, the nature of its association with other soluble species and the soil’s ability to release the metal from the solid phase to be acquired by the plants (Violante et al. 2010). Plants with a high bioconcentration factor (BCF—metal concentration ratio of plant roots to soil) and low translocation factor (TF—ratio of the concentration of the total amount of metal in leaves compared to the metal content in roots), e.g. Deschampsia cespitosa, have the potential for stabilization of ecological system (Galfati et al. 2011). Although tree fine roots adapt well to conditions with heavy metal contamination, their phytostabilization capabilities seem to be very low (Brunner et al. 2008; Kalubi et al. 2016). A survey of heavy metals interaction with a plant body is depicted in Fig. 1.

Survey of heavy metals interaction with plant body
4 Heavy Metals in Plants
Heavy metals are considered to be an important stress factor for plants. Pathways of heavy metals in plants are briefly summarized in Fig. 2. If a natural amount of heavy metals is present in soils, plants are able to avoid their negative impact (Juknys et al. 2012). However, high concentrations of heavy metals cause harmful effect on cellular and physiological processes in plants (Ma 2005; Dimkpa et al. 2009). The most abundant heavy metals in soils are Fe and Al. Fe is one of essential elements and plays an important role in plant nutrition. Mo and Mn are also important micronutrients: Zn, Ni, Cu, V, Co, W, and Cr belong among trace elements and As, Hg, Ag, Sb, Cd, Pb have no nutritional or physiological function in plants (Schützendübel and Polle 2001). Negative impact of these elements is highly dependent on their concentration in soils as well as in plants (Stankovic et al. 2014). Uptake of heavy metals is influenced by abiotic and biotic conditions. The value of pH is crucial for the solubility of heavy metals, for example Al and Fe become soluble and toxic in pH under 7. Another important parameter of heavy metal uptake are the processes of adsorption, desorption, and complexation, which are associated with individual soil types. Generally, there are two ways of heavy metal uptake: passive uptake, based on the concentration gradient across the membrane and inducible substrate-specific and energy-dependent uptake. Heavy metals uptake from soil to roots and the subsequent behavior of heavy metals on the level of tissue and cells are very complex. However, it is necessary to understand this mechanism to be able to use it in phytoremediation and improve plant tolerance to heavy metals (Schützendübel and Polle 2001). The issue remains unresolved despite great effort which has been made since the introduction of phytoremediation. Still, relevant genes were determined and recent data seem to be promising for future applications (Ahmad et al. 2016). Lolium perenne (Rees et al. 2015), Alyssum montanum (Agrawal et al. 2013), Stanleya albescens (Freeman et al. 2010), Nicotiana tabacum and Brassica juncea (Liang et al. 2009), Senecio coronatus (Mesjasz-Przybyłowicz et al. 2007), Thlaspi arvense and Thlaspi perfoliatum (Freeman et al. 2005) can be mentioned as the typical examples of promising plants (see further text).

Availability and utilization of heavy metals (HM) in plants
There are several strategies plants employ to cope with high concentrations of heavy metals in soils. The first method is the prevention of the intake of heavy metals into plant roots. Heavy metals can be removed from plants by mycorrhizal association, metal sequestration, or complexation by exuding organic compounds from roots (Verbruggen et al. 2009; Antosiewicz et al. 2014). These mechanisms usually immobilize heavy metals. If the plant is not able to avoid heavy metal uptake, tolerance mechanisms for detoxification are activated. These are metal sequestration and compartmentalization in various intracellular compartments, metal ions trafficking, metal binding to cell wall, biosynthesis or accumulation of osmolytes and osmoprotectants. If these strategies fail, antioxidants can help to keep homeostasis (Gajewska et al. 2013; Emamverdian et al. 2015).
There are several phytotoxicological impacts of heavy metals: inhibition of enzymes, inactivation of biomolecules, and oxidative stress. Generally, oxidative stress is defined as imbalance between pro-oxidant and antioxidant level which can be also entitled as oxidative homeostasis. Oxidative stress is caused by free radicals and molecules containing activated atoms of oxygen called reactive oxygen species (ROS), and it is associated with a lack of electrons which cause damage to cell compounds (Demidchik 2015). Oxidative stress can be induced by high concentrations of heavy metals, and it causes inhibition of growth. There are several ROS-generating mechanisms where heavy metals participate (Juknys et al. 2012). Redox-active metals (copper and chrome) can produce hydroxyl radicals (OH•) which are the most aggressive type of ROS. Metals without redox capacity such as cadmium, lead, zinc, nickel, etc. can involve singlet oxygen which is able to create another type of ROS superoxide (O2•). ROS can cause unspecific oxidation of proteins and membrane lipids, DNA damage, enzyme inhibition by activation of programmed cell death (Sharma et al. 2012). On the other hand, ROS play an important role in the plant defense system and cannot be totally eliminated from plants (Schützendübel and Polle 2001). The harmful effect of reactive oxygen species is given by their concentration. If the concentration of ROS exceeds the threshold level for defense mechanisms, the oxidative stress will arise (Sharma et al. 2012).
5 Toxicity of Heavy Metals
Soils contaminated with heavy metals represent an escalating problem for all living organisms, such as plants, animals, or humans. In general, mutagenic ability of heavy metals (Knasmüller et al. 1998) and oxidative stress induction via Fenton and Haber-Weiss reactions can be mentioned (Jomova and Valko 2011) and many others can be learned from literature. Adaptive responses of plants to heavy metal-contaminated environments are efficient processes including many physiological, molecular, genetic, and ecological traits, which give certain species the ability to survive or to hyperaccumulate the toxic metals (Sarma 2011).
6 Heavy Metal Phytotoxicity and Oxidative Stress Arise
Heavy metal phytotoxicity may result from alterations of numerous physiological processes caused at cellular/molecular level by inactivating enzymes, blocking functional groups of metabolically important molecules, displacing or substituting for essential elements and disrupting membrane integrity, and the phytotoxicity is an addition of reactive oxygen or nitrogen species releasing in natural pathways like photosynthesis, tricarboxylic acid cycle, and Calvin cycle (Rascio and Navari-Izzo 2011). Fenton reaction can be another pathway where both antioxidants and metals are necessary and interact together. Here, reactive species are results of the interaction. The processes are summarized in Fig. 3. Metal-induced oxidative stress has been strongly linked to early toxicity symptoms (Sharma and Dietz 2009). Complexation with metal-binding peptides, metallothioneins (MTs, gene-encoded polypeptides), and phytochelatins (PCs, enzymatically synthesized peptides) results in formation of non-toxic metabolites protecting cells and the whole organism (Singh et al. 2003). MTs have three characteristic motifs based on their cysteine content and structure: Cysteine-Cysteine, Cysteine-X-Cysteine, and Cysteine-X-X-Cysteine, in which X denotes any amino acid (Mejáre and Bülow 2001). In angiosperms, MTs can be classified into four types based on the conserved positions of Cys residues (Cobbet and Goldsbrough 2002), with some general trends in the expression: type-1 MTs expressed predominantly in roots, type-2 MTs in leaves, type-3 MTs in fruits, and type-4 MTs in seeds (Guo et al. 2008). PCs are tricarboxylic sometimes confusingly described as class III MTs (Cobbet and Goldsbrough 2002) and can be divided into five main classes: canonical PCs, homo-PCs [iso(PC)(β-Ala)], hydroxymethyl-PCs [iso(PC)(Ser)], iso-PCs [iso-PC(Glu)], and desglycine-PCs [des(Gly)PC], containing n γ-Glutamyl-Cysteine repeats capped C-terminally by a Glycine, β-Alanine, Serine, Glutamic acid, or no residue, respectively (Zenk 1996).

Link between oxidative stress and heavy metals in plants
Metal hyperaccumulation typically occurs in over 500 plant species (yet many are still unidentified) and approximately in 0.2% of all angiosperms (Sarma 2011), and metal accumulating plants (of soils in both tropical and temperate zones of all the continents) are model plants for phytoremediation (use of plants to ameliorate contaminated sites) and phytomining. The ratio of metals between soil and plant parts—metal transfer factors from soil (TFS)—is an important criterion for the selection of model plant species for phytoremediation; ratio >1 means higher accumulation of metals in plant parts than soil (Barman et al. 2000). According to Kumar et al. (2013), Parthenium hysterophorus and Spinacia oleracea have TFS above 1 for Cr, Cu, Ni, Pb, and Cd. Impatiens walleriana is able to accumulate a tenfold higher concentration of Cd in the shoot than a typical hyperaccumulator and its TFS values are greater than unity (Wei et al. 2012), while Noccaea caerulescens has TFS >1 for Ni as a consequence of inoculation (Visioli et al. 2014). Farahat and Linderholm (2015) stated that the accumulation of Zn, Mn, Cu, and Cd with transfer factors >1 for wastewater-irrigated trees indicated the ability for metal accumulation of Cupressus sempervirens. Sainger et al. (2011) found out that on the basis of TFS greater than 1, eight plant species for Zn (Achyranthes aspera, Amaranthus viridis, Senna occidentalis, Chenopodium album, Croton bonplandianum, Cynodon dactylon, Saccharum munja, Tephrosia purpurea) and Fe (Vachellia nilotica, A. aspera, A. viridis, S. occidentalis, C. bonplandianum, C. dactylon, Physalis minima, T. purpurea), three plant species for Cu (A. aspera, P. minima, S. munja), and two plant species for Ni (P. minima, T. purpurea) could be considered as hyperaccumulators (HAs) and used in phytoextraction technology. The aforementioned HAs are summarized in Table 1.
HAs are plants that belong to distantly related families, but share the ability to grow on metalliferous soils. These plants are able to accumulate extraordinarily high amounts of heavy metals in the aerial organs, far in excess of the levels found in the majority of species, without suffering phytotoxic effects (Rascio and Navari-Izzo 2011), respectively possesses genetically inherited traits of metals hyperaccumulation and tolerance. These plants are differentiated from non-hyperaccumulating species (NHAs), e.g. Lolium perenne (Rees et al. 2015), Alyssum montanum (Agrawal et al. 2013), Stanleya albescens (Freeman et al. 2010), Nicotiana tabacum and Brassica juncea (Liang et al. 2009), Senecio coronatus (Mesjasz-Przybyłowicz et al. 2007), Thlaspi arvense and Thlaspi perfoliatum (Freeman et al. 2005), by a highly increased ratio of heavy metal uptake (at concentrations 100- to 1000-fold higher than those found in NHAs). An important role is also played by faster translocation of metals from roots to shoots, and a better ability to detoxify and sequester heavy metals in leaves, where constitutive overexpression of genes, e.g. OsZIP4 and AhHMA4 (Antosiewicz et al. 2014), CsHMA3 (Park et al. 2014), AtHMA4 (Verret et al. 2004; Siemianowski et al. 2014), SaMTP1 (Zhang et al. 2011), TcYSL3 (Gendre et al. 2006), encodes transmembrane transporters. Ricachenevsky et al. (2013) revealed that although useful in basic studies of gene function, constitutive over-expression changes metal homeostasis in all organs, and often affects the uptake and distribution of more than one element.
Mechanisms involved in heavy metal increased tolerance and heavy metal distribution in an excluder non-hyperaccumulator and a hyperaccumulator plant: heavy metal binding to the cell walls and/or cell exudates, root uptake, chelation in the cytosol and/or sequestration in vacuoles, root-to-shoot translocation. The spots indicate the plant organ in which the different mechanisms occur and the spot sizes indicate their level. According to the elemental defense hypothesis the high heavy metal concentrations make hyperaccumulator leaves poisonous to herbivores (Rascio and Navari-Izzo 2011).
Initially, the term HAs referred to plants that were able to accumulate more than 1 mg/g of Ni (dry weight) in the shoot, an exceptionally high heavy metal concentration considering that in vegetative organs of most plants Ni toxicity starts from 10 to 15 μg/g (Rascio and Navari-Izzo 2011). As defined by specific phytotoxicity of other heavy metals—HAs are plants that, when growing on native soils, concentrate >10 mg/g (1%) Mn or Zn, >1 mg/g (0.1%) As, Co, Cr, Cu, Pb, Sb, Se or Tl, and >0.1 mg/g (0.01%) Cd in the aerial organs, without suffering phytotoxic damage (Verbruggen et al. 2009). Ni is hyperaccumulated by the largest amount of species (more than 75%) and reaches the highest concentration in a plant, while a low number of HAs have been found for Cd, which is one of the most toxic heavy metals (Rascio and Navari-Izzo 2011). Zn HAs are likewise less numerous. For example, the Thlaspi family are hyperaccumulating plants among which 23 species hyperaccumulate Ni, 10 species hyperaccumulate Zn, just 3 species (T. caerulescens, T. praecox, and T. goesingense) hyperaccumulate Cd and 1 species (T. praecox) hyperaccumulates Pb (Miransari 2011).
To be considered as HAs, plants have to meet also these criteria: shoot/root quotient higher than 1, i.e. level of heavy metal in the shoot divided by the level of heavy metal in the root, and extraction coefficient higher than 1, i.e. level of heavy metal in the shoot divided by the total level of heavy metal in the soil (Mganga et al. 2011).
Plants growing on metalliferous soils can be grouped into three categories: excluders, indicators, and accumulators/HAs (Bhargava et al. 2012). Most HAs are endemic to metalliferous soils, behaving as “strict metallophytes,” whereas some “facultative metallophytes” can live also on non-metalliferous ones, although they are more prevalent on metal-enriched habitats (Assunção et al. 2003). Moreover, there are species that embrace both metallicolous and non-metallicolous populations.
An interesting feature revealed by research is that most key steps in hyperaccumulation do not rely on novel genes, but depend on genes common to HAs and NHAs that are differently expressed and regulated in the two kinds of plants (Verbruggen et al. 2009), such as members of ZIP (Zinc-regulated transporter, Iron-regulated transporter Protein), HMA (Heavy Metal transporting ATPase), MATE (Multidrug And Toxin Efflux), YSL (Yellow Strip 1-Like protein), CAX (Cation Exchanger), NRAMP (Natural Resistance-Associated Macrophage Protein), and CDF (Cation Diffusion Facilitator), resp. MTP (Metal Tolerance Protein) families. Lin et al. (2014) listed in their study also genes belonging to PCR (Plant Cadmium Resistance), PDR (Pleiotropic Drug Resistance protein), and PDF (Plant Defensin) families, as well as transcripts related to metal chelator and metal chelator transporter functions, such as genes of NAS (Nicotianamine Synthase), PCS (Phytochelatin Synthase), MT, ZIF (Zinc Induced Facilitator), and MRP/ABC (Multidrug Resistance-associated Protein/ATP-Binding Cassette transporter). De Abreu-Neto et al. (2013) studied two large families of genes encoding HIPP (Heavy metal-associated Isoprenylated Plant Protein) and HPP (Heavy metal-associated Plant Protein). In addition, Krämer et al. (2007) reported other well-known proteins that mediate the transport of transition metals in plants, which belong to the following families: OPT (Oligopeptide Transporter), MFS (Major Facilitator Superfamily), COPT (Copper Transporter), CCC1 (Ca2+-sensitive Cross Complementer 1), IREG (Iron-Regulated protein) and ATM (ABC Transporters of the Mitochondria). Several cation transporters have been identified in recent years, most of which are also in the SAMS (S-Adenosyl-Methionine Synthetase) and FER (Ferritin Fe (III) binding) families (Bhargava et al. 2012). Yet not every member of all these families listed above has been functionally characterized and therefore, further study is required before their roles and functions are fully understood.
Molecular technique provides better understanding of the gene regulation systems and plant metal homeostasis. In order to protect themselves from the oxidative stress, plants have several antioxidative defense systems to scavenge toxic radicals. This defensibility is divided into two main classes: low molecular weight antioxidants, which comprise lipid-soluble membrane-associated antioxidants (e.g., α-tocopherol, β-carotene) and the water-soluble reductants (e.g., glutathione, ascorbate); and antioxidative enzymes—superoxide dismutase (SOD), ascorbate peroxidase (APX), catalase (CAT), and glutathione reductase (GR) (Sarma 2011; Zaimoglu et al. 2011; Gupta and Ahmad 2013; Mehes-Smith et al. 2013b; Adrees et al. 2015; Lou et al. 2015).
There are three main strategies involved during the detoxification of metallic ions: phytovolatilization and/or chemical transformation (the chemical conversion of toxic elements into less toxic and volatile compounds results in the removal of harmful elements from plant tissues), efflux from the cytoplasm, and binding or chelation of trace elements (Singh et al. 2003). In particular, chelation is the most widespread intracellular mechanism for the maintenance of low concentrations and detoxification of free ion metals in plant cytoplasm that can be performed by thiol compounds (such as tripeptide glutathione, metallothioneins (MTs), phytochelatins), and also by non-thiol compounds (such as organic acids and amino acids, e.g. histidine, nicotianamine) (Seth et al. 2012; Anjum et al. 2015). GSH, a tripeptide (γ-Glu-Cys-Gly), is recognized as an antioxidant that plays a key role in the defense mechanism of plants (Nahar et al. 2015) and it is a precursor for the synthesis of phytochelatins (family of peptides structurally related to GSH) in metal-exposed plants (Hossain and Komatsu 2013). MTs are low-molecular-weight, cysteine-rich proteins that have high affinity for binding metal cations, and their overexpression can increase plant tolerance to specific metals (Fernandez et al. 2012; Lv et al. 2013). Other compounds like the phenolic one can be involved in the protection and compounds like anthraquinones and flavonoids are important. Phytochelatins (PCs) are also a family of metal-complexing peptides which are rapidly induced on overexposure to metals or metalloids in plants, animals, and some yeasts (Vatamaniuk et al. 2001) and they can bind metals possessing a high affinity to sulfhydryl groups (Anjum et al. 2015).
7 Heavy Metals and Plant Saccharide Metabolism
7.1 Plant Saccharide Metabolism
Plant saccharide metabolism depends on photosynthesis and the Calvin cycle, i.e. two elemental processes that take place in a plant cell, namely inside the chloroplast. Photosynthesis is a process where water and sunlight are absorbed from the air and converted into chemical macroergic compounds (ATP, NADPH) used in subsequent reactions for production of proteins, fats, and carbohydrates. The Calvin cycle is a series of reactions following photosynthesis, where produced ATP and NADPH are utilized for the formation of 3-phosphoglyceraldehyde from carbon dioxide and ribulose-1,5-bisphosphate. Subsequently, d-fructose-6-phosphate as a precursor of d-glucose-6-phosphate and of d-mannose-6-phosphate is enzymatically produced (Fig. 4). These basic saccharides can be converted into other saccharides or can enter protein, fat, or nucleic acid metabolism (Velisek and Cejpek 2005a; Tamoi et al. 2006).

Scheme of basic plant metabolism
Saccharides, of both plant and animal origin, are the primary products of photosynthesis and the Calvin cycle and they can be divided into three groups: monosaccharides, oligosaccharides, and polysaccharides. Monosaccharides are the most basic and the most important part of plant metabolism. Types of monosaccharides are distinguished according to the count of carbon molecules in their structure; five- and six-carbon sugars belong to the most frequently occurring ones in the nature. 3-phosphoglyceraldehyde produced during the first enzymatically catalyzed reaction of the Calvin cycle belongs to three-carbon sugars, and its two molecules are important for the synthesis of d-fructose-6-phosphate, the precursor of all plant sugars. Subsequent monosaccharide metabolic processes are shown in Fig. 5 (Velisek and Cejpek 2005a; Hu et al. 2014).

Scheme of plant saccharide metabolism
Oligosaccharides can be defined as low-molecular-weight carbohydrates consisting of monosaccharide units (at least two but no more than ten of them) held together by a glycosidic bond. Polysaccharides are macromolecules of carbohydrates consisting of monosaccharide units (more than ten) that are also held together by a glycosidic bond. The molecular weight of oligosaccharides ranges between that of monosaccharides and polysaccharides in the nature. All types of sugars have great meaning for the plant itself as well as for animal and human diet and development, structure, and metabolism of these organisms (Velisek and Cejpek 2005b; Mussatto and Mancilha 2007; Kurd and Samavati 2015).
7.2 Important Plant Saccharides
Glucose and fructose are the most important monosaccharides in plant metabolism. They have a storage function (preservation of energy as monosaccharides or as oligosaccharides) and a structural function (structural polysaccharides such as starch and cellulose) (Velisek and Cejpek 2005b; Hieu et al. 2015). Fructose enters glycolysis and respiration and it also serves as a precursor for fat and protein synthesis. Fructose can also be transformed into activated forms of glucose and mannose, precursors of the synthesis of other mono-, oligo-, and polysaccharides (Velisek and Cejpek 2005a). Glucose and fructose have the function of signal molecules; they are able to begin hexose-based metabolism in the membrane or in the cytoplasm (Winter and Huber 2000; Koch 2004). The activated form of glucose (UDP-d-glucose) also serves as a precursor for the synthesis of UDP-d-glucuronic acid, and this conversion is the first step of the synthesis of nucleotide sugars (UDP-d-apiose, UDP-l-arabinose, and UDP-d-xylose) (Reiter and Vanzin 2001).
Sucrose and trehalose belong to one of the few free and the most frequently naturally occurring non-reducing disaccharides in plants. Unlike trehalose, which can only be found in few higher plant species, sucrose occurs even in algae, cyanobacteria, lower plants and all oxygenic photosynthetic organisms (Wingler 2002; Salerno and Curatti 2003). Sucrose, which consists of fructose and glucose units held together by a glycosidic bond, has a major role in many processes like cell growth, development, signaling, etc. (Cumino et al. 2002; Hieu et al. 2015; Rorabaugh et al. 2015). It has an important metabolic role as a donor of glucosyl and fructosyl moiety initiating the synthesis of polysaccharides and nucleotide sugars. Trehalose is composed of two glucose units connected by a glycosidic bond and it has storage, transport, and protective functions during heat stress and dehydration (Wingler 2002; Salerno and Curatti 2003).
Cellulose, callose, and pectin are the most important plant cell wall polysaccharides. Starch and fructans are the major plant storage sugars (Velisek and Cejpek 2005b). Both cellulose, i.e. a homogenous polymer of β-(1,4) glucose, and callose, i.e. a β-(1,3) glucose polymer, are synthesized from UDP-glucose, but cellulose is generated in the plasma membrane, and callose-like pectin originates in the Golgi apparatus (Amor et al. 1995; Perrin 2001; Reiter 2002). Unlike callose and starch, glucose residues of cellulose are repeated in their structural chains (Brown et al. 1996). Cellulose is responsible for cell growth, shape, tissue morphology, and extension (Perrin 2001). Callose is rarely generated within in vivo plant metabolism; however, it can be found in cell plates, sieve tubes and during some stages of mega- and microsporogenesis. Callose is responsible for plants’ wound, stress and infection responses, and it is part of structures involved in cell growth and differentiation (Brown et al. 1996; Kudlicka and Brown 1997). Pectin is composed of α-1,4-glucuronic acid residues occurring in the primary wall of all higher plants except grasses and relative plants, and it plays a major role in cell wall structure, growth, and development (Velisek and Cejpek 2005b).
Starch is a plant polysaccharide consisting of two glucose homopolymers—amylose and amylopectin. Amylopectin is structurally close to the animal polysaccharide of glycogen, it is highly branched and formed of α-(1,4)-linked and α-(1,6)-branched glucose chains (Myers et al. 2000; Li et al. 2015a, b). Amylose has a linear structure consisting of glucose units with α-(1,4) bonds. Starch is generated in leaves during the day when it is stored in chloroplasts, and it is utilized for sucrose synthesis during the night. Fructans serve for the deposition of energy in flowering plants. They are made up of β-(1,2) fructose units linked together, and the end unit of glucose is usually bound (Velisek and Cejpek 2005b; Wasserman et al. 2015).
7.3 Heavy Metal Toxic Effect on Plant Saccharides
Saccharides biosynthesis pathway in plant metabolism includes fixation of carbon dioxide (the Calvin cycle), metabolic transformation of basic monosaccharides into other monosaccharides, and finally the distribution of saccharides throughout the plant and their storage or utilization (Slewinski and Braun 2010). Metals, such as Zn, Fe, Cu, and others, have great influence on plant metabolism and structure and they are required as co-factors in many enzymatically catalyzed reactions. Their major contribution is based on the ability to affect plant photosynthesis, respiration, ethylene perception, circadian clock, and programmed cell death, and they participate on the protection of plant metabolism (Clemens 2006; Garcia et al. 2014). On the other hand, higher concentrations of metals are toxic for plants. Most metals inhibit plant growth due to their interference with elements that are essential for enzymatic functions. Additionally, natural redox activity of heavy metals (Cu, Fe) requested under physiological conditions may increase the plant’s production of reactive oxygen species in high metal concentrations. The synthesis of chlorophyll and other photosynthetic pigments in plant leaves is also decreased by the long-term effect of heavy metals. Reduction of pigments is caused either due to the inhibition of the enzymatic system of chloroplasts or due to insufficiency of essential compounds (Das et al. 1997; Chettri et al. 1998; Fargasova 2004; Clemens 2006; Mera et al. 2016). As a result of human activity, heavy metals begin to be more available in soils and natural sources, ecosystems become damaged, and plant metabolism is disrupted (Das et al. 1997; He et al. 2015).
The toxic effect of heavy metals also leads to inhibition of saccharide metabolism in plants. Four basic metals have been reported as toxic for plant metabolism. Ni is able to affect the mobilization of saccharides in germinating seeds. Pb may decelerate root growth via increased saccharide contents and retard saccharide metabolizing enzymes: α-amylase, β-amylase and invertase as well as Cu and Cd (Solanki and Dhankhar 2011; Sethy and Ghosh 2013).
The effect of Cd on plant metabolism was examined in many studies. Cd is toxic for plants where it becomes accumulated and also for animals that eat plants affected by this metal. Nevertheless, Cd has the most toxic effect on the human body where it gets accumulated in organs after the consumption of food containing Cd (Kirkham 2006).
Hédiji et al. (2010) studied the long-term effect of Cd on the growth and metabolic profile of tomato plants, namely its influence on the metabolism of carotenoids, carbohydrates, organic acids, and amino acids. Tomato plants were treated with two concentrations of Cd (20 and 100 μmol/L). The content of soluble carbohydrates (glucose, fructose, and sucrose) was found out on the basis of saccharide metabolism. Results showed an increase in glucose and sucrose concentrations in mature leaves treated with 100 μmol/L of Cd. Treatment with 20 μmol/L of Cd exhibited an increase in the concentration of sucrose and a decrease in the concentration of glucose and fructose. The mechanism of Cd interference with carbohydrate metabolism, especially the inhibition of the activity of invertase caused by Cd, was proposed as an explanation of soluble saccharides accumulation (Hédiji et al. 2010).
Rahoui et al. (2015) studied the effect of Cd on seedlings of six Medicago truncatula lines with different Cd susceptibility. The content of total soluble sugars, glucose, fructose, and sucrose was determined during 6 days of Cd treatment. Results showed that saccharide metabolism was a key component of Cd stress response. While Cd-tolerant lines of M. truncatula were characterized by high concentrations of glucose and/or sucrose in embryonic axes, high concentrations of fructose were determined in embryonic axes of susceptible lines of M. truncatula. The content of saccharides in susceptible lines was higher than in the tolerant lines. In contrast, tolerant lines showed higher mobilization of total soluble sugars and over-consumption of glucose under the toxic effect of Cd. Both lines affected sugar transport (Rahoui et al. 2015).
An increase in other saccharides (galactose, myoinositol, trehalose, and raffinose) was observed after Cd-treatment of Arabidopsis thaliana in the study of Sun et al. (2010).
Earlier studies suggested some pathways of Cd influence on plant saccharide metabolism: metallo-inhibition of saccharide transport in the bean and rice (Moya et al. 1993), Cd-caused reduction of saccharide transport and altering of α-amylase activity in cotyledon of the pea and the faba bean (Mihoub et al. 2005; Rahoui et al. 2008, 2010). Probably many other Cd-influenced mechanisms are taking place in plant organisms (Rahoui et al. 2015).
Since not only Cd has toxic effect on plant metabolism, studies using other heavy metals or multiple heavy metals together as noxious substances were performed.
The effect of Cd and Pb on growth and biochemical parameters was determined by John et al. (2008) on the water plant of Lemna polyrhiza L., which is known for its great ability to accumulate heavy metals. Plants were observed for their content of soluble sugars during 30 days of treatment with different concentrations of Cd and Pb. The findings showed that lower concentrations of Cd and Pb increased the content of soluble sugars. In contrast, higher concentrations of the metals (Cd >5 mg/kg of soil) decreased sugar content, which was probably caused by photosynthetic inhibition, by over-stimulation of respiration or by possible interaction with the active site of ribulosebisphosphate carboxylase (John et al. 2008).
The effect of Cu on the content of saccharides in 20-day-old cucumber plants was examined by Alaoui-Sossé et al. (2004). The content of starch and sucrose was measured in the first and second leaves and in roots. Starch was increased in both types of leaves when compared to controls, but the content of starch in the first leaves was significantly higher than in the second leaves. Starch was not detected in roots. The content of sucrose was similarly increased in both types of leaves, but it was not affected in roots. Saccharide accumulation may result from Cu inhibition of photosynthesis. However, it may also be a result of a decrease in phloem loading, a problem with the capacity of assimilate transport or it may be caused by a reduction of the utilization of nutrients including saccharides (Alaoui-Sossé et al. 2004).
Saccharides belong to significant nutrients in plant metabolism. They are the first products of CO2 fixation, a precursor of the synthesis of many important organic compounds, they create energy for respiration, protect plants from wounds, infections, and stress situations, and they take care about detoxification pathways. Therefore, the disruption of saccharide metabolism may lead to a loss of the protective ability and structural integrity of the whole plant (Solanki and Dhankhar 2011).
8 Determination of Heavy Metals and Oxidative Stress
8.1 Determination of Heavy Metals in Biological Samples
The contamination of ecosystems and exposure to toxic metals is a significant worldwide burden. Therefore, biomonitoring techniques are getting more relevant because they may help to recognize contaminated area or crop, distribution of metals in the ecosystem and control of potential environmental hazards caused by heavy metal pollution (Ugulu 2015).
Major analytical techniques used to determine heavy metals in environmental matrices are Atomic Absorption Spectrometry (AAS), Inductively Coupled Plasma Atomic Emission Spectrometry (ICP/AES), Inductively Coupled Plasma Mass Spectrometry (ICP/MS), Neutron Activation Analysis (NAA), X-ray fluorescence (XRF), and Ion Chromatography (IC) (Li et al. 2015a, b; Markiewicz et al. 2015; Muller et al. 2015).
A large number of studies implemented so far to evaluate heavy-metal toxicity have focused on the development of analytical methods for the assessment of toxic species in diverse samples, mostly using techniques for elemental analysis, e.g. atomic absorption and emission spectroscopy, and inductively coupled plasma-mass spectrometry being probably the most widely used analytical technique for both multi-elemental analysis and speciation (Luque-Garcia et al. 2011).
In recent years, improvements in protein separation and identification methods and the progress of genomic knowledge have led to an enhancement in the utilization of proteomic techniques to answer biological questions (Isaacson et al. 2006). Plenty of investigations have shown that proteomics, in conjunction with bioinformatics tools, can facilitate the discovery of new, better biomarkers of metal exposure (Zhai et al. 2005).
Various proteomic approaches are accessible, including gel-based and gel-free methods. The most commonly used method in metal-toxicity-related proteomic studies is classical two-dimensional gel electrophoresis (2-DE), which has been predominantly coupled with Edman sequencing or peptide-mass fingerprinting (PMF) by means of a matrix-assisted laser desorption ionization-time-of-flight (MALDI-TOF) mass spectrometer (Luque-Garcia et al. 2011); a less frequent option is to use tandem MS (MS2) {MALDI-TOF/TOF or LC-MS2}, allowing sequencing of peptides and providing more confident protein identification.
A non-gel-based method, e.g. multidimensional protein identification technology (MudPIT), which involves the generation of peptides from a complex protein mixture, followed by two-dimensional chromatographic separation (Visioli et al. 2010), can partly resolve problems that are connected with the use of gel-based proteomic strategies.
According to Luque-Garcia et al. (2011), in the past decade, several quantitative proteomic strategies based on labeling proteins and peptides have been developed, e.g. isotope-coded affinity tag (ICAT), stable-isotope labeling by amino acids in cell culture (SILAC), isobaric tags for relative and absolute quantitation (iTRAQ), isotope-coded protein label (ICPL), N-terminal labeling, labeling with heavy water and even label-free techniques.
In general, the methods of analysis for the determination of heavy metals are changing to meet new or unforeseen conditions, and to ensure precision of measurement. The current analytical methods are precise enough to cover demand for the samples processing. On the other hand, prices of MS devices are still too high to be available for small labs or small companies, and research as well as introduction of simple measuring protocols is still desired. Last but not least important thing in actual literature is the limited number of analyses. As mentioned, modern/actual instrumentation and measurements are very expensive so the precious data for individual experiments are still very limited in number. Actual statistical and scientifically used programs like Statistica or C, C++, R-languages show perfectly values and trends of measured data but need still at least 7 analyses. However, X0 analyses are highly needed for appropriate modeling and better understanding of the real problems. For more details about statistical approach and modern uses of statistics in science, see, for example: Rencher (2002); Ostrouchov et al. (2012); Cass (2016).
8.2 Determination of Oxidative Stress
Three ways can be chosen for the determination of oxidative homeostasis and to judge whether homeostasis was disturbed and oxidative stress arose. Because ROS are not stable, they are not suitable as readily determined markers and therefore stable adducts. Therefore, products of radical mediated reactions and oxidized macromolecules serve as direct markers. Protein carbonyls, malondialdehyde as a product of radical degradation of lipids and 8-oxo-deoxyguanosine can be introduced as typical markers of oxidative damage (Yang et al. 2012; Pohanka 2013; Samsel et al. 2013). Malondialdehyde and protein carbonyls can be simply assayed in biological samples by spectrophotometry (Pohanka 2014c). 8-Oxo-deoxyguanosine can be determined by a competitive immunoassay like Enzyme-Linked Immunosorbent Assay (Gedik et al. 2002). Chromatography techniques for analysis of stress markers are available as well (Al-Rimawi 2015).
The occurrence of oxidative imbalance is followed by the expression of enzymes serving as antioxidants, i.e. enzymes with the ability to detoxify ROS. Such enzymes can be used as markers, and the emergence of oxidative stress can be deduced from their huge presence in plant tissues. Superoxide dismutase is a typical enzyme with significant antioxidant potency, and it is easily used as a marker of oxidative stress in plants (Cui et al. 2015; Jain et al. 2015; Rady and Hemida 2015). Catalase and peroxidase are other enzymes expressed under stress conditions (Naz et al. 2015).
Important information about a plant’s condition and its ability to suppress oxidative stress can be learned from the measurement of the total level of low molecular weight antioxidants. In principle, the total level of antioxidants can be measured in two ways. Firstly, antioxidants can be identified as chemical entities and their amount is typically determined by chromatography (Abdennacer et al. 2015; Wang et al. 2015). Secondly, the total antioxidant capacity is measured by a simple technique. Assays named after the used reagent or principle of assay, such as ABTS [from 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid)], FRAP (from ferric reducing antioxidant power), ORAC (Oxygen Radical Absorbance Capacity), and DPPH (2,2-diphenyl-1-picrylhydrazyl) can be mentioned as examples (Ramirez-Anaya Jdel et al. 2015; Zhang et al. 2015). These approaches are simpler when compared to chromatography, and they achieve complex results. On the other hand, the information as to whether all or an isolated type of antioxidants are depleted remains hidden. Ascorbic acid, epigallocatechin gallate, quercetin, and its derivatives are examples of typical plants’ low molecular weight antioxidants (El-Hawary et al. 2011). Markers of oxidative stress are briefly summarized in Table 2.
9 Conclusions
Heavy metals represent a significant problem for the environment especially in former industrial areas. Long- term persistence of heavy metals is another problem which significantly worsens impact of heavy metals on the environment. Although most attention pertaining to this issue has been focused on their toxicity with respect to humans and animals, plants are not saved from the harmful impact of heavy metals either. Still, plants are potent in protecting themselves from heavy metals because of their ability to cumulate and distribute the metals into their body and store them for a long time. Undoubtedly, differences between certain plants and reasons why some plant species are more sensitive to the presence of metals or better at accumulating them have not been fully understood. The impact on oxidative homeostasis and the development of ROS during the exposure is probably one of significant effects mediated by heavy metals. However, more work on the issue should be done prior to making any definitive conclusions. Further research should be focused on both studying of metals impact on the plants including identification of pathological mechanisms, and establishing of processes where plants can serve as a tool for the metals remove from the environments.
10 Summary
Though toxicity of heavy metals is known in a junction to human health and animal laboratory models, plants appears to be out of the knowledge. It is a little surprising because environmental impact of heavy metals is also mediated through plants. In this paper, basic facts about heavy metals, their distribution in soil, mobility, accumulation by plants, and initiation of oxidative stress including the decline in basal metabolism are presented. The both actual and frontier studies in the field are summarized and discussed. The major pathophysiological pathways are introduced and known relations between heavy metals and their ability to initiate an oxidative damage are outlined for plants. Mobility and bioaccessibility are other factors that should be taken into consideration when heavy metals toxicity is evaluated and the both factors are discussed here. This review contains a wide discussion about metals like lead, mercury, copper, cadmium, iron, zinc, nickel, and vanadium. This survey can be concluded by a statement that heavy metals are significant contributors to pathological processes in most of the known and studied plants and oxidative stress takes place in these processes.
References
Abdennacer B, Karim M, Yassine M, Nesrine R, Mouna D, Mohamed B (2015) Determination of phytochemicals and antioxidant activity of methanol extracts obtained from the fruit and leaves of Tunisian Lycium intricatum Boiss. Food Chem 174:577–584. https://doi.org/10.1016/j.foodchem.2014.11.114
Adrees M, Ali S, Iqbal M et al (2015) Mannitol alleviates chromium toxicity in wheat plants in relation to growth, yield, stimulation of anti-oxidative enzymes, oxidative stress and Cr uptake in sand and soil media. Ecotox Environ Safe 122:1–8. https://doi.org/10.1016/j.ecoenv.2015.07.003
Agrawal B, Czymmek KJ, Sparks DL, Bais HP (2013) Transient influx of nickel in root mitochondria modulates organic acid and reactive oxygen species production in nickel hyperaccumulator Alyssum murale. J Biol Chem 288(10):7351–7362. https://doi.org/10.1074/jbc.M112.406645
Ahmad R, Tehsin Z, Malik ST et al (2016) Phytoremediation potential of hemp (Cannabis sativa L.): identification and characterization of heavy metals responsive genes. Clean-Soil Air Water 44(2):195–201. https://doi.org/10.1002/clen.201500117
Alaoui-Sossé B, Genet P, Vinit-Dunand F, Toussaint ML, Epron D, Badot PM (2004) Effect of copper on growth in cucumber plants (Cucumis sativus) and its relationships with carbohydrate accumulation and changes in ion contents. Plant Sci 166:1213–1218. https://doi.org/10.1016/j.plantsci.2003.12.032
Al-Rimawi F (2015) Development and validation of a simple reversed-phase HPLC-UV method for determination of malondialdehyde in olive oil. J Am Oil Chem Soc 92(7):933–937. https://doi.org/10.1007/s11746-015-2664-x
Amor Y, Haigle CH, Johnson S, Wainscott M, Delmer DP (1995) A membrane-associated form of sucrose synthase and its potential role in synthesis of cellulose and callose in plants. Proc Natl Acad Sci U S A 92(20):9353–9357. https://doi.org/10.1073/pnas.92.20.9353
Anjum NA, Hasanuzzaman M, Hossain MA et al (2015) Jacks of metal/metalloid chelation trade in plants-an overview. Front Plant Sci 6(192):1–17. https://doi.org/10.3389/fpls.2015.00192
Antosiewicz DM, Barabasz A, Siemianowski O (2014) Phenotypic and molecular consequences of overexpression of metal-homeostasis genes. Front Plant Sci 5(80):1–7. https://doi.org/10.3389/fpls.2014.00080
Assunção AGL, Schat H, Aarts MGM (2003) Thlaspi caerulescens, an attractive model species to study heavy metal hyperaccumulation in plants. New Phytol 159(2):351–360. https://doi.org/10.1046/j.1469-8137.2003.00820.x
Barman SC, Sahu RK, Bhargava SK, Chaterjee C (2000) Distribution of heavy metals in wheat, mustard, and weed grown in field irrigated with industrial effluents. Bull Environ Contam Toxicol 64(4):489–496. https://doi.org/10.1007/s001280000030
Bhargava A, Carmona FF, Bhargava M, Srivastava S (2012) Approaches for enhanced phytoextraction of heavy metals. J Environ Manage 105:103–120. https://doi.org/10.1016/j.jenvman.2012.04.002
Brown RM Jr, Saxena IM, Kudlicka K (1996) Cellulose biosynthesis in higher plants. Trends Plant Sci 1:149–156. doi: https://doi.org/10.1016/S1360-1385(96)80050-1
Brunner I, Luster J, Günthardt-Goerg MS, Frey B (2008) Heavy metal accumulation and phytostabilisation potential of tree fine roots in a contaminated soil. Environ Pollut 152(3):559–568. https://doi.org/10.1016/j.envpol.2007.07.006
Cass S (2016) The 2016 top programming languages. http://spectrum.ieee.org/computing/software/the-2016-top-programming-languages
Chettri MK, Cook CM, Vardaka E, Sawidis T, Lanaras T (1998) The effect of Cu, Zn and Pb on the chlorophyll content of the lichens Cladonia convoluta and Cladonia rangiformis. Environ Exp Bot 39(1):1–10. https://doi.org/10.1016/S0098-8472(97)00024-5
Clemens S (2006) Toxic metal accumulation, responses to exposure and mechanisms of tolerance in plants. Biochimie 88(11):1707–1719. https://doi.org/10.1016/j.biochi.2006.07.003
Cobbet C, Goldsbrough P (2002) Phytochelatins and metallothioneins: roles in heavy metal detoxification and homeostasis. Annu Rev Plant Physiol 53:159–182. https://doi.org/10.1146/annurev.arplant.53.100301.135154
Colzi I, Doumett S, Del Bubba M et al (2011) On the role of the cell wall in the phenomenon of copper tolerance in Silene paradoxa L. Environ Exp Bot 72:77–83. https://doi.org/10.1016/j.envexpbot.2010.02.006
Cui LJ, Huang Q, Yan B et al (2015) Molecular cloning and expression analysis of a Cu/Zn SOD gene (BcCSD1) from Brassica campestris ssp chinensis. Food Chem 186:306–311. https://doi.org/10.1016/j.foodchem.2014.07.121
Cumino A, Curatti L, Giarrocco L, Salerno GL (2002) Sucrose metabolism: anabaena sucrose-phosphate synthase and sucrose-phosphate phosphatase define minimal functional domains shuffled during evolution. FEBS Lett 517(1–3):19–23. https://doi.org/10.1016/S0014-5793(02)02516-4
Das P, Samantaray S, Rout GR (1997) Studies on cadmium toxicity in plants: a review. Environ Pollut 98(1):29–36. https://doi.org/10.1016/S0269-7491(97)00110-3
De Abreu-Neto JB, Turchetto-Zolet AC, De Oliveira LF, Zanettini MH, Margis-Pinheiro M (2013) Heavy metal-associated isoprenylated plant protein (HIPP): characterization of a family of proteins exclusive to plants. FEBS J 280(7):1604–1616. https://doi.org/10.1111/febs.12159
De Matos AT, Fontes MPF, Da Costa LM, Martinez MA (2001) Mobility of heavy metals as related to soil chemical and mineralogical characteristics of Brazilian soils. Environ Pollut 111(3):429–435. https://doi.org/10.1016/S0269-7491(00)00088-9
Debbaudt AL, Ferreira ML, Gschaider ME (2004) Theoretical and experimental study of M2+ adsorption on biopolymers. III. Comparative kinetic pattern of Pb, Hg and Cd. Carbohydr Polym 56(3):321–332. https://doi.org/10.1016/j.carbpol.2004.02.009
Demidchik V (2015) Mechanisms of oxidative stress in plants: from classical chemistry to cell biology. Environ Exp Bot 109:212–228. https://doi.org/10.1016/j.envexpbot.2014.06.021
Dimkpa CHO, Merten D, Svatos A, Büchel G, Kothe E (2009) Metal-induced oxidative stress impacting plant growth in contaminated soil is alleviated by microbial siderophores. Soil Biol Biochem 41(1):154–162. https://doi.org/10.1016/j.soilbio.2008.10.010
Duffus JH (2002) “Heavy metals” a meaningless term? IUPAC Tech Rep Pure Appl Chem 74(5):793–807. https://doi.org/10.1351/pac200274050793
Duffus JH (2003) Errata “heavy metals” a meaningless term? IUPAC Tech Rep Pure Appl Chem 75(9):1357. https://doi.org/10.1351/pac200375091357
El-Hawary SA, Sokkar NM, Ali ZY, Yehia MM (2011) A profile of bioactive compounds of Rumex vesicarius L. J Food Sci 76(8):1195–1202. https://doi.org/10.1111/j.1750-3841.2011.02370.x
Emamverdian A, Ding Y, Mokhberdoran F, Xie Y (2015) Heavy metal stress and some mechanisms of plant defense response. Sci World J 2015:1–18. https://doi.org/10.1155/2015/756120
Ettler V, Kribek B, Majer V, Knesl I, Mihaljevic M (2012) Differences in the bioaccessibility of metals/metalloids in soils from mining and smelting areas (Copperbelt, Zambia). J Geochem Explor 113:68–75. https://doi.org/10.1016/j.gexplo.2011.08.001
Farahat E, Linderholm HW (2015) The effect of long-term wastewater irrigation on accumulation and transfer of heavy metals in Cupressus sempervirens leaves and adjacent soils. Sci Total Environ 512–513:1–7. https://doi.org/10.1016/j.scitotenv.2015.01.032
Fargasova A (2004) Toxicity comparison of some possible toxic metals (Cd, Cu, Pb, Se, Zn) on young seedlings of Sinapis alba L. Plant Soil Environ 50(1):33–38
Fernandez LR, Vandenbussche G, Roosens N, Govaerts C, Goormaghtigh E, Verbruggen N (2012) Metal binding properties and structure of a type III metallothionein from the metal hyperaccumulator plant Noccaea caerulescens. Biochim Biophys Acta 1824(9):1016–1023. https://doi.org/10.1016/j.bbapap.2012.05.010
Flouty R, Khalaf G (2015) Role of Cu and Pb on Ni bioaccumulation by Chlamydomonas reinhardtii: validation of the biotic ligand model in binary metal mixtures. Ecotox Environ Safe 113:79–86. https://doi.org/10.1016/j.ecoenv.2014.11.022
Franco CR, Chagas AP, Jorge RA (2004) Ion-exchange equilibria with aluminum pectinates. Colloid Surf A 204(1–3):183–192. https://doi.org/10.1016/S0927-7757(01)01134-7
Freeman JL, Garcia D, Kim D, Hopf A, Salt DE (2005) Constitutively elevated salicylic acid signals glutathione-mediated nickel tolerance in Thlaspi nickel hyperaccumulators. Plant Physiol 137(3):1082–1091. https://doi.org/10.1104/pp.104.055293
Freeman JL, Tamaoki M, Stushnoff C et al (2010) Molecular mechanisms of selenium tolerance and hyperaccumulation in Stanleya pinnata. Plant Physiol 153(4):1630–1652. https://doi.org/10.1104/pp.110.156570
Gajewska E, Głowacki R, Mazur J, Skłodowska M (2013) Differential response of wheat roots to Cu, Ni and Cd treatment: oxidative stress and defense reactions. Plant Growth Regul 71(1):13–20. https://doi.org/10.1007/s10725-013-9803-x
Galfati I, Bilal E, Sassi AB, Abdallah H, Zaier A (2011) Accumulation of heavy metals in native plants growing near the phosphate treatment industry, Tunisia. Carpath J Earth Env 6(2):85–100
Garcia L, Welchen E, Gonzalez DH (2014) Mitochondria and copper homeostasis in plants. Mitochondrion 19:269–274. https://doi.org/10.1016/j.mito.2014.02.011
Gedik CM, Boyle SP, Wood SG, Vaughan NJ, Collins AR (2002) Oxidative stress in humans: validation of biomarkers of DNA damage. Carcinogenesis 23(9):1441–1446. https://doi.org/10.1093/carcin/23.9.1441
Gendre D, Czernic P, Conéjéro G et al (2006) TcYSL3, a member of the YSL gene family from the hyperaccumulator Thlaspi caerulescens, encodes a nicotianamine-Ni/Fe transporter. Plant J 49(1):1–15. https://doi.org/10.1111/j.1365-313X.2006.02937.x
Guo WJ, Meetam M, Goldsbrough PB (2008) Examining the specific contributions of individual Arabidopsis metallothioneins to copper distribution and metal tolerance. Plant Physiol 146:1697–1706. https://doi.org/10.1104/pp.108.115782
Guo X, Meng H, Zhu S, Zhang T, Yu S (2015) Purifying sugar beet pectins from non-pectic components by means of metal precipitation. Food Hydrocoll 51:69–75. https://doi.org/10.1016/j.foodhyd.2015.05.009
Gupta AK, Ahmad M (2013) Effect of refinery waste effluent on tocopherol, carotenoid, phenolics and other antioxidants content in Allium cepa. Toxicol Ind Health 29(7):652–661. https://doi.org/10.1177/0748233712436639
He F, Liu Q, Zheng L, Cui Y, Shen Z, Zheng L (2015) RNA-Seq analysis of rice roots reveals the involvement of post-transcriptional regulation in response to cadmium stress. Front Plant Sci 6(1136):1–16. https://doi.org/10.3389/fpls.2015.01136
Hechmi N, Ben Aissa N, Abdenaceur H, Jedidi N (2015) Uptake and bioaccumulation of pentachlorophenol by emergent wetland plant Phragmites australis (common reed) in cadmium co-contaminated soil. Int J Phytoremediation 17:109–116. https://doi.org/10.1080/15226514.2013.851169
Hédiji H, Djebali W, Cabasson C et al (2010) Effects of long-term cadmium exposure on growth and metabolomic profile of tomato plants. Ecotox Environ Safe 73(8):1965–1974. https://doi.org/10.1016/j.ecoenv.2010.08.014
Hieu HC, Li H, Miyauchi Y, Mizutani G, Fujita N, Nakamura Y (2015) Wetting effect on optical sum frequency generation (SFG) spectra of D-glucose, D-fructose, and sucrose. Spectrochim Acta A Mol Biomol Spectrosc 138:834–839. https://doi.org/10.1016/j.saa.2014.10.108
Hossain Z, Komatsu S (2013) Contribution of proteomic studies towards understanding plant heavy metal stress response. Front Plant Sci 3(310):1–12. https://doi.org/10.3389/fpls.2012.00310
Hu Y, Wang T, Yang X, Zhao Y (2014) Analysis of compositional monosaccharides in fungus polysaccharidesby capillary zone electrophoresis. Carbohydr Polym 102:481–488. https://doi.org/10.1016/j.carbpol.2013.11.054
Isaacson T, Damasceno CMB, Saravanan RS et al (2006) Sample extraction techniques for enhanced proteomic analysis of plant tissues. Nat Protoc 1(2):769–774. https://doi.org/10.1038/nprot.2006.102
Jain R, Chandra A, Venugopalan VK, Solomon S (2015) Physiological changes and expression of SOD and P5CS genes in response to water deficit in sugarcane. Sugar Tech 17(3):276–282. https://doi.org/10.1007/s12355-014-0317-2
John R, Ahmad P, Gadgil K, Sharma S (2008) Effect of cadmium and lead on growth, biochemical parameters and uptake in Lemna polyrrhiza L. Plant Soil Environ 54(6):262–270
Jomova K, Valko M (2011) Advances in metal-induced oxidative stress and human disease. Toxicology 283(2–3):65–87. https://doi.org/10.1016/j.tox.2011.03.001
Juknys R, Vitkauskaité G, Račaité M, Vencloviené J (2012) The impacts of heavy metals on oxidative stress and growth of spring barley. Cent Eur J Biol 7(2):299–306. https://doi.org/10.2478/s11535-012-0012-9
Kalubi KN, Mehes-Smith M, Omri A (2016) Comparative analysis of metal translocation in red maple (Acer rubrum) and trembling aspen (Populus tremuloides) populations from stressed ecosystems contaminated with metals. Chem Ecol 32(4):312–323. https://doi.org/10.1080/02757540.2016.1142978
Kartel MT, Kupchik LA, Veisov BK (1999) Evaluation of pectin binding of heavy metal ions in aqueous solutions. Chemosphere 38(11):2591–2596. https://doi.org/10.1016/S0045-6535(98)00466-4
Kim RY, Yoon JK, Kim TS, Yang JE, Owns G, Kim KR (2015) Bioavailability of heavy metals in soils: definitions and practical implementation—a critical review. Environ Geochem Health 37(6):1041–1061. https://doi.org/10.1007/s10653-015-9695-y
Kirkham MB (2006) Cadmium in plants on polluted soils: effects of soil factors, hyperaccumulation, and amendments. Geoderma 137(1):19–32. https://doi.org/10.1016/j.geoderma.2006.08.024
Knasmüller S, Gottmann E, Steinkellner H et al (1998) Detection of genotoxic effects of heavy metal contaminated soils with plant bioassays. Mutat Res 420(1–3):37–48. https://doi.org/10.1016/S1383-5718(98)00145-4
Koch K (2004) Sucrose metabolism: regulatory mechanisms and pivotal roles in sugar sensing and plant development. Curr Opin Plant Biol 7(3):235–246. https://doi.org/10.1016/j.pbi.2004.03.014
Krämer U, Talke IN, Hanikenne M (2007) Transition metal transport. FEBS Lett 581(12):2263–2272. https://doi.org/10.1016/j.febslet.2007.04.010
Kudlicka K, Brown RM Jr (1997) Cellulose and Callose biosynthesis in higher plants. Plant Physiol 115(2):643–656. https://doi.org/10.1104/pp.115.2.643
Kumar N, Bauddh K, Kumar S, Dwivedi N, Singh DP, Barman SC (2013) Accumulation of metals in weed species grown on the soil contaminated with industrial waste and their phytoremediation potential. Ecol Eng 61:491–495. https://doi.org/10.1016/j.ecoleng.2013.10.004
Kurd F, Samavati V (2015) Water soluble polysaccharides from Spirulina platensis: extraction and in vitro anti-cancer activity. Int J Biol Macromol 74:498–506. https://doi.org/10.1016/j.ijbiomac.2015.01.005
Li E, AC W, Li J, Liu Q, Gilbert RG (2015a) Improved understanding of rice amylose biosynthesis from advanced starch structural characterization. Rice 8(20):1–8. https://doi.org/10.1186/s12284-015-0055-4
Li ZM, Yu Y, Li ZL (2015b) A review of biosensing techniques for detection of trace carcinogen contamination in food products. Anal Bioanal Chem 407(10):2711–2726. https://doi.org/10.1007/s00216-015-8530-8
Liang HM, Lin TH, Chiou JM, Yeh KC (2009) Model evaluation of the phytoextraction potential of heavy metal hyperaccumulators and non-hyperaccumulators. Environ Pollut 157(6):1945–1952. https://doi.org/10.1016/j.envpol.2008.11.052
Lin YF, Severing EI, Te Lintel Hekkert B, Schijlen E, Aarts MGM (2014) A comprehensive set of transcript sequences of the heavy metal hyperaccumulator Noccaea caerulescens. Front Plant Sci 5(261):1–15. https://doi.org/10.3389/fpls.2014.00261
Lou Y, Yang Y, Hu L, Liu H, Xu Q (2015) Exogenous glycinebetaine alleviates the detrimental effect of Cd stress on perennial ryegrass. Ecotoxicology 24(6):1330–1340. https://doi.org/10.1007/s10646-015-1508-7
Luque-Garcia JL, Cabezas-Sanchez P, Camara C (2011) Proteomics as a tool for examining the toxicity of heavy metals. Trends Anal Chem 30(5):703–716. https://doi.org/10.1016/j.trac.2011.01.014
Lv Y, Deng X, Quan L, Xia Y, Shen Z (2013) Metallothioneins BcMT1 and BcMT2 from Brassica campestris enhance tolerance to cadmium and copper and decrease production of reactive oxygen species in Arabidopsis thaliana. Plant and Soil 367(1–2):507–519. https://doi.org/10.1007/s11104-012-1486-y
Ma JF (2005) Plant root responses to three abundant soil minerals: silicon, aluminum and iron. Crit Rev Plant Sci 24(4):267–281. https://doi.org/10.1080/07352680500196017
Markiewicz B, Komorowicz I, Belter M, Baralkiewicz D (2015) Chromium and its speciation in water samples by HPLC/ICP-MS—technique establishing metrological traceability: a review since 2000. Talanta 132:814–828. https://doi.org/10.1016/j.talanta.2014.10.002
Mata YN, Blázquez ML, Ballester A, González F, Muñoz JA (2009) Sugar-beet pulp pectin gels as biosorbent for heavy metals: preparation and determination of biosorption and desorption characteristics. Chem Eng J 150(2–3):289–301. https://doi.org/10.1016/j.cej.2009.01.001
Mehes-Smith M, Nkongolo KK, Narendrula R, Cholewa E (2013a) Mobility of heavy metals in plants and soil: a case study from a mining region in Canada. Am J Environ Sci 9(6):483–493. https://doi.org/10.3844/ajessp.2013.483.493
Mehes-Smith M, Nkongolo KK, Cholewa E (2013b) Coping mechanisms of plants to metal contaminated soil. In: Silvern S, Young S (eds) Environmental change and sustainability. Chapter 3, pp 53–90. InTech. ISBN: 978-953-51-1094-1. doi: https://doi.org/10.5772/55124
Mejáre M, Bülow L (2001) Metal binding proteins and peptides in bioremediation and phytoremediation of heavy metals. Trends Biotechnol 19(2):67–73. https://doi.org/10.1016/S0167-7799(00)01534-1
Mera R, Torres E, Abalde J (2016) Influence of sulphate on the reduction of cadmium toxicity in the microalga Chlamydomonas moewusii. Ecotox Environ Safe 128:236–245. https://doi.org/10.1016/j.ecoenv.2016.02.030
Mesjasz-Przybyłowicz J, Barnabas A, Przybyłowicz W (2007) Comparison of cytology and distribution of nickel in roots of Ni-hyperaccumulating and non-hyperaccumulating genotypes of Senecio coronatus. Plant and Soil 293:61–78. https://doi.org/10.1007/s11104-007-9237-1
Mganga N, Manoko MLK, Rulangaranga ZK (2011) Classification of plants according to their heavy metal content around North Mara gold mine, Tanzania: implication for phytoremediation. Tanz J Sci 37:109–119
Mihoub A, Chaoui A, El Ferjani E (2005) Biochemical changes associated with cadmium and copper stress in germinating pea seeds (Pisum sativum L.) C R Biol 328(1):33–41. https://doi.org/10.1016/j.crvi.2004.10.003
Miransari M (2011) Hyperaccumulators, arbuscular mycorrhizal fungi and stress of heavy metals. Biotechnol Adv 29(6):645–653. https://doi.org/10.1016/j.biotechadv.2011.04.006
Moya JL, Ros R, Picazo I (1993) Influence of cadmium and nickel on growth, photosynthesis and carbohydrate distribution in rice plants. Photosynth Res 36(2):75–80. https://doi.org/10.1007/BF00016271
Muller AL, Oliveira JS, Mello PA, Muller EI, Flores EM (2015) Study and determination of elemental impurities by ICP-MS in active pharmaceutical ingredients using single reaction chamber digestion in compliance with USP requirements. Talanta 136:161–169. https://doi.org/10.1016/j.talanta.2014.12.023
Mussatto SI, Mancilha IM (2007) Non-digestible oligosaccharides: a review. Carbohydr Polym 68(3):587–597. https://doi.org/10.1016/j.carbpol.2006.12.011
Myers AM, Morell MK, James MG, Ball SG (2000) Recent progress toward understanding biosynthesis of the amylopectin crystal. Plant Physiol 122:989–997. https://doi.org/10.1104/pp.122.4.989
Nahar K, Hasanuzzaman M, Alam MM, Fujita M (2015) Exogenous glutathione confers high temperature stress tolerance in mung bean (Vigna radiata L.) by modulating antioxidant defense and methylglyoxal detoxification system. Environ Exp Bot 112:44–54. https://doi.org/10.1016/j.envexpbot.2014.12.001
Naz FS, Yusuf M, Khan TA, Fariduddin Q, Ahmad A (2015) Low level of selenium increases the efficacy of 24-epibrassinolide through altered physiological and biochemical traits of Brassica juncea plants. Food Chem 185:441–448. https://doi.org/10.1016/j.foodchem.2015.04.016
Ostrouchov G, Chen W-C, Schmidt D, Patel P (2012) Programming with big data in R. http://r-pbd.org/
Park W, Feng Y, Ahn SJ (2014) Alteration of leaf shape, improved metal tolerance, and productivity of seed by overexpression of CsHMA3 in Camelina sativa. Biotechnol Biofuels 7(96):1–17. https://doi.org/10.1186/1754-6834-7-96
Perrin RM (2001) Cellulose: how many cellulose synthases to make a plant? Curr Biol 11(6):213–216. https://doi.org/10.1016/S0960-9822(01)00108-7
Pohanka M (2013) Role of oxidative stress in infectious diseases. A review. Folia Microbiol 58(6):503–513. https://doi.org/10.1007/s12223-013-0239-5
Pohanka M (2014a) Alzheimer’s disease and oxidative stress. A review. Curr Med Chem 21(3):356–364. https://doi.org/10.2174/09298673113206660258
Pohanka M (2014b) Copper, aluminum, iron and calcium inhibit human acetylcholinesterase in vitro. Environ Toxicol Pharmacol 37(1):455–459. https://doi.org/10.1016/j.etap.2014.01.001
Pohanka M (2014c) Caffeine alters oxidative homeostasis in the body of BALB/c mice. Bratisl Med J 115(11):699–703. https://doi.org/10.4149/BLL_2014_135
Puls RW, Powell RM, Clark D, Eldred CJ (1991) Effects of pH, solid/solution ratio, ionic strength and organic acids on Pb and Cd sorption on kaolinite. Water Air Soil Pollut 57–58(1):423–430. https://doi.org/10.1007/BF00282905
Rady MM, Hemida KA (2015) Modulation of cadmium toxicity and enhancing cadmium-tolerance in wheat seedlings by exogenous application of polyamines. Ecotoxic Environ Safe 119:178–185. https://doi.org/10.1016/j.ecoenv.2015.05.008
Rahoui S, Chaoui A, El Ferjani E (2008) Differential sensitivity to cadmium in germinating seeds of three cultivars of faba bean (Vicia faba L.) Acta Physiol Plant 30(4):451–456. https://doi.org/10.1007/s11738-008-0142-x
Rahoui S, Chaoui A, El Ferjani E (2010) Reserve mobilization disorder in germinating seeds of Vicia faba exposed to cadmium. J Plant Nutr 33(5–8):809–817. https://doi.org/10.1080/01904161003654055
Rahoui S, Chaoui A, Ben C, Rickauer M, Gentzbittel L, El Ferjani E (2015) Effect of cadmium pollution on mobilization of embryo reserves in seedlings of six contrasted Medicago truncatula lines. Phytochemistry 111:98–106. https://doi.org/10.1016/j.phytochem.2014.12.002
Ramirez-Anaya Jdel P, Samaniego-Sanchez C, Castañeda-Saucedo MC, Villalon-Mir M, López-García de la Serrana H (2015) Phenols and the antioxidant capacity of Mediterranean vegetables prepared with extra virgin olive oil using different domestic cooking techniques. Food Chem 188:430–438. https://doi.org/10.1016/j.foodchem.2015.04.124
Rascio N, Navari-Izzo F (2011) Heavy metal hyperaccumulating plants: how and why do they do it? And what makes them so interesting? Plant Sci 180(2):169–181. https://doi.org/10.1016/j.plantsci.2010.08.016
Rees F, Germain C, Sterckeman T, Morel JL (2015) Plant growth and metal uptake by a non-hyperaccumulating species (Lolium perenne) and a Cd-Zn hyperaccumulator (Noccaea caerulescens) in contaminated soils amended with biochar. Plant Soil 395(1–2):57–73. https://doi.org/10.1007/s11104-015-2384-x
Reiter WD (2002) Biosynthesis and properties of the plant cell wall. Curr Opin Plant Biol 5(6):536–542. https://doi.org/10.1016/S1369-5266(02)00306-0
Reiter WD, Vanzin GF (2001) Molecular genetics of nucleotide sugar interconversion pathways in plants. Plant Mol Biol 47(1–2):95–113. https://doi.org/10.1007/s11104-015-2384-x
Rencher AC (2002) Methods of multivariate analysis. Wiley series in probability and mathematical statistics. Wiley, Hoboken, NJ
Ricachenevsky FK, Menguer PK, Sperotto RA, Williams LE, Fett JP (2013) Roles of plant metal tolerance proteins (MTP) in metal storage and potential use in biofortification strategies. Front Plant Sci 4(144):1–16. https://doi.org/10.3389/fpls.2013.00144
Rorabaugh JM, Stratford JM, Zahniser NR (2015) Differences in bingeing behavior and cocaine reward following intermittent access to sucrose, glucose or fructose solutions. Neuroscience 301:213–220. https://doi.org/10.1016/j.neuroscience.2015.06.015
Sainger PA, Dhankhar R, Sainger M, Kaushik A, Singh RP (2011) Assessment of heavy metal tolerance in native plant species from soils contaminated with electroplating effluent. Ecotox Environ Safe 74(8):2284–2291. https://doi.org/10.1016/j.ecoenv.2011.07.028
Salerno GL, Curatti L (2003) Origin of sucrose metabolism in higher plants: when, how and why? Trends Plant Sci 8(2):63–69. https://doi.org/10.1016/S1360-1385(02)00029-8
Samsel M, Dzierzbicka K, Trzonkowski P (2013) Adenosine, its analogues and conjugates. Postepy Hig Med Dosw 67:1189–1203. https://doi.org/10.5604/17322693.1078588
Sarma H (2011) Metal hyperaccumulation in plants: a review focusing on phytoremediation technology. J Environ Sci Technol 4(2):118–138. https://doi.org/10.3923/jest.2011.118.138
Schützendübel A, Polle A (2001) Plant responses to abiotic stresses: heavy metal induced oxidative stress and protection by mycorrhization. J Exp Bot 53(372):1351–1365. https://doi.org/10.1093/jexbot/53.372.1351
Seth CS, Remans T, Keunen E et al (2012) Phytoextraction of toxic metals: a central role for glutathione. Plant Cell Environ 35(2):334–346. https://doi.org/10.1111/j.1365-3040.2011.02338.x
Sethy SK, Ghosh S (2013) Effect of heavy metals on germination of seeds. J Nat Sci Biol Med 4(2):272–275. https://doi.org/10.4103/0976-9668.116964
Sharma SS, Dietz KJ (2009) The relationship between metal toxicity and cellular redox imbalance. Trends Plant Sci 14(1):43–50. https://doi.org/10.1016/j.tplants.2008.10.007
Sharma P, Jha AB, Dubey RS, Pessarakli M (2012) Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J Bot 2012:1–26. https://doi.org/10.1155/2012/217037
Siemianowski O, Barabasz A, Kendziorek M et al (2014) HMA4 expression in tobacco reduces Cd accumulation due to the induction of the apoplastic barrier. J Exp Bot 65(4):1125–1139. https://doi.org/10.1093/jxb/ert471
Singh OV, Labana S, Pandey G, Budhiraja R (2003) Phytoremediation: an overview of metallic ion decontamination from soil. Appl Microbiol Biotechnol 61(5–6):405–412. https://doi.org/10.1007/s00253-003-1244-4
Slewinski TL, Braun DM (2010) Current perspectives on the regulation of whole-plant carbohydrate partitioning. Plant Sci 178(4):341–349. https://doi.org/10.1016/j.plantsci.2010.01.010
Solanki R, Dhankhar R (2011) Biochemical changes and adaptive strategies of plants under heavy metal stress. Biologia 66(2):195–204. https://doi.org/10.2478/s11756-011-0005-6
Stankovic S, Kalaba P, Stankovic AR (2014) Biota as toxic metal indicators. Environ Chem Lett 12(1):63–84. https://doi.org/10.1007/s10311-013-0430-6
Sun X, Zhang J, Zhang H et al (2010) The responses of Arabidopsis thaliana to cadmium exposure explored via metabolite profiling. Chemosphere 78(7):840–845. https://doi.org/10.1016/j.chemosphere.2009.11.045
Tamoi M, Nagaoka M, Miyagawa Y, Shigeoka S (2006) Contribution of fructose-1,6-bisphosphatase and sedoheptulose-1,7-bisphosphatase to the photosynthetic rate and carbon flow in the Calvin cycle in transgenic plants. Plant Cell Physiol 47(3):380–390. https://doi.org/10.1093/pcp/pcj004
Topolska J, Latowski D, Kaschabek S, Manecki M, Merkel BJ, Rakovan J (2014) Pb remobilization by bacterially mediated dissolution of pyromorphite Pb5(PO4)3Cl in presence of phosphate-solubilizing Pseudomonas putida. Environ Sci Pollut R 21(2):1079–1089. https://doi.org/10.1007/s11356-013-1968-3
Ugulu I (2015) Determination of heavy metal accumulation in plant samples by spectrometric techniques in Turkey. Appl Spectrosc Rev 50(2):113–151. https://doi.org/10.1080/05704928.2014.935981
Van Bussel CGJ, Schroeder JP, Mahlmann L, Schulz C (2014) Aquatic accumulation of dietary metals (Fe, Zn, Cu, Co, Mn) in recirculating aquaculture systems (RAS) changes body composition but not performance and health of fuvenile turbot (Psetta maxima). Aquacult Eng 61:35–42. https://doi.org/10.1016/j.aquaeng.2014.05.003
Vatamaniuk OK, Bucher EA, Ward JT, Rea PA (2001) A new pathway for heavy metal detoxification in animals. Phytochelatin synthase is required for cadmium tolerance in Caenorhabditis elegans. J Biol Chem 276:20817–20820. https://doi.org/10.1074/jbc.C100152200
Velisek J, Cejpek K (2005a) Biosynthesis of food constituents: saccharides. 1. Monosaccharides, oligosaccharides, and related compounds—a review. Czech J Food Sci 23(4):129–144
Velisek J, Cejpek K (2005b) Biosynthesis of food constituents: saccharides. 2. Polysaccharides—a review. Czech J Food Sci 23(5):173–183
Verbruggen N, Hermans C, Schat H (2009) Molecular mechanisms of metal hyperaccumulation in plants. New Phytol 181(4):759–776. https://doi.org/10.1111/j.1469-8137.2008.02748.x
Verret F, Gravot A, Auroy P et al (2004) Overexpression of AtHMA4 enhances root-to-shoot translocation of zinc and cadmium and plant metal tolerance. FEBS Lett 576(3):306–312. https://doi.org/10.1016/j.febslet.2004.09.023
Violante A, Cozzolino V, Perelomov L, Caporale AG, Pigna M (2010) Mobility and bioavailability of heavy metals and metalloids in soil environments. J Soil Sci Plant Nutr 10(3):268–292. https://doi.org/10.4067/S0718-95162010000100005
Visioli G, Marmiroli M, Marmiroli N (2010) Two-dimensional liquid chromatography technique coupled with mass spectrometry analysis to compare the proteomic response to cadmium stress in plants. J Biomed Biotechnol 2010:1–10. https://doi.org/10.1155/2010/567510
Visioli G, D’Egidio S, Vamerali T, Mattarozzi M, Sanangelantoni AM (2014) Culturable endophytic bacteria enhance Ni translocation in the hyperaccumulator Noccaea caerulescens. Chemosphere 117:538–544. https://doi.org/10.1016/j.chemosphere.2014.09.014
Wang T, Sun H (2013) Biosorption of heavy metals from aqueous solution by UV-mutant Bacillus subtilis. Environ Sci Pollut R 20(10):7450–7463. https://doi.org/10.1007/s11356-013-1767-x
Wang C, Hua D, Yan C (2015) Structural characterization and antioxidant activities of a novel fructan from Achyranthes bidentata Blume, a famous medicinal plant in China. Ind Crop Prod 70:427–434. https://doi.org/10.1016/j.indcrop.2015.03.051
Wasserman LA, Sergeev AI, Vasil’ev VG et al (2015) Thermodynamic and structural properties of tuber starches from transgenic potato plants grown in vitro and in vivo. Carbohydr Polym 125:214–223. https://doi.org/10.1016/j.carbpol.2015.01.084
Wei JL, Lai HY, Chen ZS (2012) Chelator effects on bioconcentration and translocation of cadmium by hyperaccumulators, Tagetes patula and Impatiens walleriana. Ecotox Environ Safe 84:173–178. https://doi.org/10.1016/j.ecoenv.2012.07.004
Wingler A (2002) The function of trehalose biosynthesis in plants. Phytochemistry 60(5):437–440. https://doi.org/10.1016/S0031-9422(02)00137-1
Winter H, Huber SC (2000) Regulation of sucrose metabolism in higher plants: localization and regulation of activity of key enzymes. Crit Rev Biochem Mol Biol 35(4):253–289. https://doi.org/10.1080/10409230008984165
Yang L, Chen JH, Xu T, Zhou AS, Yang HK (2012) Rice protein improves oxidative stress by regulating glutathione metabolism and attenuating oxidative damage to lipids and proteins in rats. Life Sci 91(11–12):389–394. https://doi.org/10.1016/j.lfs.2012.08.003
Zaimoglu Z, Koksal N, Basci N, Kesici M, Gulen H, Budak F (2011) Antioxidative enzyme activities in Brassica juncea L. and Brassica oleracea L. plants under chromium stress. J Food Agric Environ 9(1):676–679
Zenk MH (1996) Heavy metal detoxification in higher plants—a review. Gene 179(1):21–30. https://doi.org/10.1016/S0378-1119(96)00422-2
Zhai R, Su S, Lu X et al (2005) Proteomic profiling in the sera of workers occupationally exposed to arsenic and lead: identification of potential biomarkers. Biometals 18(6):603–613. https://doi.org/10.1007/s10534-005-3001-x
Zhang M, Senoura T, Yang X, Nishizawa NK (2011) Functional analysis of metal tolerance proteins isolated from Zn/Cd hyperaccumulating ecotype and non-hyperaccumulating ecotype of Sedum alfredii Hance. FEBS Lett 585(16):2604–2609. https://doi.org/10.1016/j.febslet.2011.07.013
Zhang S, Cui Y, Li L et al (2015) Preparative HSCCC isolation of phloroglucinolysis products from grape seed polymeric proanthocyanidins as new powerful antioxidants. Food Chem 188:422–429. https://doi.org/10.1016/j.foodchem.2015.05.030
Acknowledgments
This work was supported by the Ministry of Education, Youth and Sports of the Czech Republic under the CEITEC 2020 (LQ1601) and NAZV QJ1320122 projects. We thank all reviewers for their constructive reviews of this manuscript, and Prof. Pim de Voogt for editorial comments that led to improvements of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer International Publishing AG
About this chapter
Cite this chapter
Fryzova, R. et al. (2017). Oxidative Stress and Heavy Metals in Plants. In: de Voogt, P. (eds) Reviews of Environmental Contamination and Toxicology Volume 245. Reviews of Environmental Contamination and Toxicology, vol 245. Springer, Cham. https://doi.org/10.1007/398_2017_7
Download citation
DOI: https://doi.org/10.1007/398_2017_7
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-75036-1
Online ISBN: 978-3-319-75037-8
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)