
Abstract
Proso millet (Panicum miliaceum L.) is a climate-resilient, ancient, and gluten-free cereal. This allotetraploid crop is healthy for humans and the environment. Its use as human food is steadily increasing, especially for people with diabetes and celiac disease due to its exceptional nutritional properties and gluten-free starch. There is no comprehensive review on proso millet seed nutraceuticals quantity, quality, characteristics, genetics, genomics (omics), and genetic improvement strategies. This review article aims to summarize published research on these aspects of proso millet seed nutrients and nutraceuticals. There are lots of reports on proso millet seed nutrients (macro, micro, and secondary metabolites) and their health benefits. There are a significant number of resources of “omics” tools for proso millet overall genetic improvement. However, no or very little genetic and genomic information (e.g., genes and genetic control mechanisms) and biosynthesis of majority of the nutraceuticals of proso millet seed are available. It may take years to see the full potential of “omics” for proso millet nutraceutomics. Phytochemical analyses and omics technologies are getting more efficient, faster, and cheaper. With this new opportunity, it is essential that proso millet scientists around the world especially food technologists, plant breeders, and geneticists of both public and private sectors work collaboratively for the genetic improvement of proso millet for nutraceutical. This will stimulate industries to use more proso millet in food products for human health and nutritional security under global climate change.
Access provided by Autonomous University of Puebla. Download reference work entry PDF
Similar content being viewed by others

Keywords
1 Introduction
Proso millet (Panicum miliaceum L.), a small-grain, annual cereal, belongs to the Panicoideae subfamily of the Poaceae family. It is one of the earliest cereal crops domesticated by mankind. It was reported that proso millet was domesticated around 8000 BC in the semi-arid regions of northern China (Lu et al. 2009). This minor millet is known by different names in different geographical locations. The word “proso” is a pan-Slavik name for millet (Santra et al. 2019). In the USA, it is commonly referred to as common millet or hog millet. Proso millet is also known as broomcorn millet in China; common millet in Japan, Korea, and other Asia-Pacific countries; Gijang in Korea; Barri in India; Mijo in Spain; Hersey millet in Germany; and French white or panic millet in France (Santra et al. 2019; Das et al. 2019). This millet is globally distributed owing to its wide adaptability to different climatic zones. The most notable countries producing proso millet are China, the USA, India, Ukraine, Russia, Nepal, Belarus, Turkey, and Romania (Das et al. 2019) (Fig. 1). Proso millet seeds were brought to the USA in 1875 by German-Russian immigrants who first cultivated this cereal along the eastern Atlantic coast of the country. Its cultivation later spread westward into the interior of the North American continent (Wietgrefe 1990; Santra 2013; Habiyaremye et al. 2017).

Major proso millet growing countries in the world. (The map was created on mapchart.net (https://www.mapchart.net/world.html))
Proso millet plants are erect, 30–100 cm tall with hollow stems (known as culm) and an adventitious root system (Baltensperger 2002). Proso millet plants develop 10–45 cm long, drooping panicles which can be either open (lax) or compact. The grains (botanically known as caryopsis) are normally 3 mm long and 2 mm wide. A great deal of variation is observed in the panicle’s shape and grain color. The panicle morphology can vary widely from its shape (e.g., compact, open, semi-open), attitude (e.g., erect, semi-erect, dropping), and branches (e.g., lax, medium, and dense). The grains can be white, cream, yellow, red, orange, brown, green, black, or tan (Santra et al. 2019). Proso millet is predominantly self-pollinated with ~10% cross-pollination. It is considered an allotetraploid with nine basic sets of chromosomes (2n = 4x = 36). The proso millet genome is estimated to be 930 megabase pairs (Mbp) (Zou et al. 2019). Hunt et al. (2014) studied the evolution of proso millet species. Proso millet was believed to have evolved through the natural hybridization of two diploid progenitors followed by the natural polyploidization. Hunt et al. reported that diploid witchgrass (Panicum capillare L.) and one diploid parent of the tetraploid torpedo grass (P. repens) are the probable diploid parents of proso millet (Hunt et al. 2014).
Proso millet is primarily cultivated in the predominantly winter wheat-growing semi-arid areas of the High Plains of the USA (Rajput et al. 2016). This region receives an average annual precipitation of ~400 mm. Conserving soil by minimizing water loss and maintaining high organic matter is paramount to sustaining agriculture in this dry region. The usual practice of summer fallow to preserve soil moisture often leads to depletion of soil organic matter (Nielsen and Calderón 2011). Planting wheat following corn and millet and only millet reportedly doubled the productivity in comparison with the wheat-fallow system (Anderson et al. 1999). Even though a wide selection of crops, such as corn, teff, millets, soybean, and peas, are available for crop rotation, proso millet stands out as a rotational crop for the dryland production system. This is because proso millet, being a C4 crop, has exceptional water-use efficiency and can survive harsh conditions like drought, high temperatures, and limited available nutrients (Habiyaremye et al. 2017). This cereal has a short-growing season (60–90) days, which allows it to avoid prolonged exposure to drought conditions. Besides, the shallow root system (90–120 cm) conserves deep soil moisture for the next crop (Das et al. 2019). Cultivation of proso millet lowers weed, disease, and insect pressure on wheat (Habiyaremye et al. 2017). All these attributes make proso millet an excellent choice as a rotation crop in rainfed agricultural systems under water-stress situation.
Millet is considered a nutraceutical and functional food because of its superior seed nutrients such as starch, proteins, vitamins, minerals, dietary fibers, and antioxidant activities. It also has other bioactive compounds such as resistant starch, lipids, phenolic acid, flavonoids, lignin, phytosterols, phytic acid, and tannins present in millets that have health-promoting properties (Tripathi et al. 2021). Proso millet is gluten-free and is abundant in vitamin B, iron, calcium, potassium, zinc, and magnesium (Das et al. 2019). Proso millet could be exploited as nutraceuticals and used in therapeutic applications for chronic diseases such as obesity, cardiovascular diseases, cancer, and diabetes (Majid and Priyadarshini 2020). Millets are grossly neglected as human food despite being highly nutritious primarily due to a lack of awareness. The increasing number of reports on health benefits of millets in recent years generated a renewed interest in using millets for biomedical research. Proso millet grain has protective and preventive effects against old age-related diseases such as diabetes, high blood pressure, cardiovascular diseases, cancer, Parkinson’s disease, and other metabolic syndromes (Kalinová 2007).
The use of proso millet grain for human consumption predates recorded human history. This small grain was widely used for preparing traditional sweet and savory cuisines in Russia, China, and Germany thousands of years ago (Santra and Rose 2013). Proso and other millet-based food are quite common in many countries in Asia and Africa. However, this ancient grain is largely under-utilized in developed countries in Europe and North America despite its remarkable nutritional and health-promoting properties (Das et al. 2019). Proso millet grains are primarily used as birdseed and animal feed in the USA and Europe (Santra et al. 2019). Nevertheless, these grains are primarily used for human consumption in several countries including China, Korea, Japan, India, and Russia. Many traditional cuisines and commercial food products are prepared from whole millet grains. Proso millet grains are used in porridge, rice-based meals, noodles, bread, pasta, flour, pancakes, and alcoholic beverages (Santra et al. 2019; Das et al. 2019). Proso millet grains can be used for ethanol production like corn (Rose and Santra 2013).
The term “omics” is commonly used to refer to a broad field of science and engineering focusing on the interactions among biological information objects pertaining to various “omes” (https://omicstutorials.com/home/). It involves collective quantification and characterization of collections of biological molecules that define the structural and functional attributes of an organism. The primary focuses of omics are on (1) mapping information objects such as genes, proteins, and ligands, (2) finding interaction relationships among the objects, (3) engineering the networks and objects to understand and manipulate the regulatory mechanisms, and (4) integrating various “omes” and “omics” subfields. The five major types of “omics” are (1) genomics, which deals with the analysis of the DNA sequence; (2) transcriptomics, which deals with the analysis of transcribed RNA; (3) proteomics, which focuses on proteins present in a sample; (4) metabolomics, which involves identification and quantification of all metabolites in a sample; and (5) miRNAomics, which studies regulatory mechanisms underlying control of transcription of genes. Here, we introduced a new term called “nutraceutomics” to describe the comprehensive study associated with all phytochemicals present in seeds of food crops (proso millet in this chapter) and their effects on human health and nutrition.
The goal of this chapter is to provide a comprehensive review of available literature for (1) proso millet seed composition and their impacts on human health, (2) variabilities of the seed compositions within proso millet germplasm available across the globe, and (3) genetic improvement for changing proso millet seed compositions through breeding (conventional and modern) and biotechnologies.
2 Proso Millet Seed Nutraceuticals
A nutraceutical is defined as any substance that is a food or part of a food and provides health benefits, including the prevention and treatment of disease. The term nutraceutical is the hybrid of “nutrition” and “pharmaceutical” and was coined by Stephen Defelice in 1989 (Kalra 2003). Nutraceuticals can be of two types: (1) macronutrients such as carbohydrates, proteins, lipids, and fibers (Table 1) and microelements such as minerals (Table 2) and vitamins (Table 3) and (2) secondary metabolites such as polyphenols, flavonoids, etc. These nutraceuticals in proso millet are discussed below.
2.1 Starch
Starch, a polysaccharide, is the major portion (97.1%) of the total saccharides (64.5 g/100 g) in proso millet (Kalinová 2007). Like in any other cereal, there are two types of starch in proso millet, namely, amylopectin and amylose. Amylose is the straight-chain polymer of glucose, whereas amylopectin is the branched-chain polymer of glucose units. In proso millet, the amylose content varies from 17.21% to 32.6% (dry basis), and amylopectin varies from 67.40% to 82.79%. The amylose content in proso millet starch is a little higher than maize (17–27%) and marginally lower than rice (28%). Native starch of proso millet has the highest digestibility (50%) in comparison to other millets (Kumari and Thayumanavan 1998), and the starch digestibility is similar to that of maize (43%). In general, grain starch has very low solubility in water. The proso millet starch is approximately 6.89% soluble at 90 °C. Moreover, millet starch exhibit a higher water-binding capacity than wheat starch and has a higher tendency to break down (~50 Brabender units - BU). Proso and foxtail millets show lower set back values (about 330 BU)compared to other millets (Kumari and Thayumanavan 1998).
The dietary patterns of many people concerned about healthy eating habit are changing across the world. Proso millet is gradually gaining attention owing to its impressive nutrient profile and gluten-free property. The eating quality and cooking parameters of proso millet grains are attributable to the starch properties. The starch granules in proso millet vary greatly in size and shape. The size of the granules range between 0.3 and 17 μm and can be round or polygonal in shape. Proso millet varieties are categorized as waxy (~1% amylose) and non-waxy (~25% amylose) types. Another important reason for consuming the proso millet is its high resistant starch content, which causes slow starch digestion, particularly in non-waxy varieties. Because of these reasons, the non-waxy starch varieties of proso-millet are widely used in food processing. The variation in amylose content and structural integrity contribute to the pasting and thermal behavior of proso millet starch. The starch of proso millet showed some promising functional properties that make it a suitable alternative to commercially available starch for food and non-food applications. There are several reports on the use of proso millet seeds in the preparation of different food products like gluten-free bread, porridge, pasta, ready-to-eat breakfast cereals, infant foods, and distilleries (Das et al. 2019). It can be concluded that proso millet is an alternative source of starch considering its quality characteristics (Bangar et al. 2021).
2.2 Protein
The seed protein content of proso millet is comparable to maize and wheat. Proso millet seeds contain higher amount of protein than any other millets (Geervani and Eggum 1989). The protein content depends on the variety, environmental conditions, soil nutrients, and weather conditions during grain formation. The average seed protein in proso millet ranges from 11.5% to 13.0% with a maximum of about 17% (Kalinova and Moudry 2006). Varieties with a red seed coat have the lowest quantity and quality of protein (Essential Amino Acid Index, or EAAI) than light-colored grains. Dehulled grains have a little bit higher protein content (12.3–16.3%) due to the removal of the seed coat (hull) that is poor in proteins (Jones et al. 1970; Ravindran 1992). The protein quality (EAAI) of proso millet grain (51–67%) is higher than that in maize.
Proteins are complex biomolecules as they may sometimes comprise several subunits with various physicochemical and functional properties. On the basis of solubility, plant storage proteins are classified as albumin (water-soluble), globulin (saline-soluble), prolamin (alcohol-soluble), and glutelin (alkaline-soluble). Prolamin (31–50%) is the major storage protein in proso millet like in most cereals (Jones et al. 1970; Dendy 1995). Glutinous (waxy) proso millet contains a higher proportion of albumins and globulins (18–31%) than non-glutinous types (13–16%) (Parameswaran and Thayumanavan 1995). Similar to other cereals, proso millet grain protein has inadequate amount of content (3.68%). However, proso millet protein is rich of some essential amino acids (leucine, isoleucine, methionine) compared to wheat protein. Millet protein is believed to have beneficial influence on cholesterol metabolism. Proso millet is well-suited for patients on a gluten-free diet as its grains contain the specific prolamin fraction under the permitted level (Kalinova and Moudry 2006).
The proximate analysis of seed storage proteins is essential for understanding the basic physical and functional properties of proso millet cultivars to form the foundation for developing value-added products. Nine US proso millet varieties were characterized for proximate analysis by Singh et al. (2018). They observed a wide range of values for the physical properties, such as sphericity (0.86–0.91), volume 3.94 to 5.14 mm3, bulk density (765.49–809.67 kg m−3), porosity (42.49–44.20%), and angle of repose (22.98–25.74°). The varieties were also evaluated for their pasting and gelatinization properties. A high correlation was observed between amylose content and onset temperature (r = −0.94), peak gelatinization temperature (r = −0.92), peak viscosity (r = 0.84), final viscosity (r = 0.91), and setback viscosity (r = 0.90) (Singh et al. 2018).
The knowledge of the protein structure-function relationship is necessary to understand the nutritional properties of seed storage proteins. The physicochemical, functional, and structural properties of the storage protein fraction from two defatted proso millet cultivars, viz., Dawn and Plateau, were determined by Akharume et al. (2020). The protein recovery efficiency for the two varieties were determined to be 53.5% (Dawn) and 60.1% (Plateau). The average denaturation temperature of all fractions was 82.1 ± 3.5 °C. Surface hydrophobicity values of Dawn were 11,781, 10,594, 316, and 2225 for albumin, globulin, and glutelin, respectively, and 3415, 2865, 353, and 456 of Plateau fractions, respectively. Most of the protein fractions exhibited the highest solubility at pH 9 and lowest solubilities at pH ≤ 7. The solubility varied from 5.7% to 100%. Emulsifying activity index (EAI) of less than 0.25 m2/g was estimated for most fractions, while the highest emulsion stability index (ESI) was about 60 min. Prolamin fraction showed three major peptide bands of 11, 14, and 24 kDa, while glutelin fraction revealed only a major band of 15 kDa and several minor bands of 11, 22, 24, 78, and 209 kDa. No differences in the electrophoresis pattern were observed for the fraction with or without a reducing agent (Akharume et al. 2020).
Digestibility of seed proteins of proso millet and other millets is reduced significantly after cooking due to conformational changes. Panicin, zein-like, the major seed storage protein fraction in proso millet, produced hydrophobic aggregates upon heating that are extremely resistant to protease digestion (Gulati et al. 2017). Using in vitro digestion as a measure of digestibility, aggregation reduces digestibility from about 80% to <40%. The recalcitrant hydrophobic protein aggregates that form as a result of heating are resistant to both aspartic proteases (pepsin) and serine proteases (chymotrypsin/trypsin). To estimate variation in “digestibility” as a trait, in vitro digestibility across 33 accessions of proso millet from different countries was measured and observed significant variation, ranging from 26% to 57% digestibility after heating (Gulati et al. 2018). Thus, there appears to be ample trait variation for selective breeding.
To further understand the nature of this digestibility trait, liquid chromatography-mass spectrometry (LC-MS)/MS analysis was done on the lines with high and low protein digestibility and identified three peptide sequences from the panicin fraction that were correlated with low digestibility and two that were associated with high digestibility (Gulati et al. 2018). Three peptide sequences of Gulati et al. (2018) were used to query annotated genes Panicum miliaceum genome (Zou et al. 2019). Seven different genes were identified belonging to the alpha family of zein proteins, the major seed storage proteins of maize. BLAST alignments identified a total of 14 members of the zein-alpha family in proso millet based on alignments showing >35% identity to the query. Phylogenetic analysis (Fig. 2a) further divided these zein-like genes into five subfamilies that are related to the PMS-1, P22.1/22A-1, and ZA1/M1 members of the 19 KDa and 22KDa zein alpha family and two members of the 10KDa prolamin-like protein family. The arrangement of the 14 zein-like proteins and the prolamin proteins on the proso millet genome (Fig. 2b) revealed an architecture suggesting that two pairs of highly related zein-like genes have arisen through chromosome duplications (Chr3-Chr4 and Chr9-Chr10), while a combination of five highly related 22 KDa zein-like genes on Chr17-Chr18 appears to have arisen from a combination of ancestral gene duplications within the progenitor of the Chr17/Chr18 chromosomes, followed by duplication of the ancestral chromosome. The four remaining genes across Chr2, Chr8, Chr14, and Chr18 form distinct families that are more distantly related. Structurally, the amino acid composition of predicted proteins from each of these zein-like subfamilies shares features common to zein proteins, including enrichment in polar amino acids, and 9–10 different stretches of repeated sequence containing poly-glutamine (QQQ) separated on average by 11 residues. Structural analysis of the maize zeins has shown the repeated sequences form alpha helices with the polar amino acids (including glutamine residues) interacting with residues on adjacent anti-parallel helices to form a bundle of multiple alpha helices in a compact structure (Argos et al. 1982; Matsushima et al. 1997).

Panicin (zein-like), prolamin gene family in proso millet (Panicum miliaceum). (a) Genes identified using three peptide sequences of prolamin fraction (Gulati et al. 2017) in BLAST alignment of proso millet genome sequence (Zou et al. 2019). Fourteen zein-line genes are divided into five sub-families. Chromosome number is indicated on top of each gene (unpublished). (b) The proso millet genome sequence assembly reported by Zou et al. (2019). This image, licensed under a Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), was used with minor modifications.
2.3 Lipids
Dehulled grains of proso millet contain 3.5% to 6.7% of lipid (Jones et al. 1970; Ravindran 1992; Kalinová 2007). Among cereals, only oats have a higher (7.14%) lipid than that in proso millet. The germ contains about 25% of the total lipids. Proso millet gains contain 62.2% of free, 27.8% of bound, and 10% of structural lipids. Unsaturated fatty acids make up 85–89% of proso millet lipids of which 42% are polyunsaturated acids (Kalinová 2007). Polyunsaturated lipids (particularly linoleic acid) play an essential role in cholesterol metabolism. The major fatty acids of proso millet are linolenic (38.4–66.68%), oleic (21.4–22.7%), and palmitic (6.61–11.3%) acids. The physicochemical characteristics of proso millet lipids (consistence, setting point, acid number) are similar to the lipid content of sunflower, maize, and soya (Kalinová 2007).
2.4 Fiber
Cereal grains serve as the most significant source of dietary fiber for the human diet. The fiber content in the human diet is critical for the prevention and treatment of high blood pressure and high cholesterol. The fiber content in dehulled grains of proso millet is like that of oat (0.8–1.2%). The composition of cell wall in proso millet grains is similar to that of tissues of other grass species. The cell wall comprises xyloglucan, arabinoxylan, uronic acid, arabi-nosyl, galactosyl residues, arabinogalactan, and β-D-glucans (Kalinová 2007). The β-D-glucans (glucose units linked to a long polymer chain) play an important role in human nutrition by decreasing total blood cholesterol. The β-glucans content in proso millet (0.5–1.0%) is at the same level of lentils (0.4–1.1%), maize (0.5–1.3%), rice (0.4–0.9%), spelt (0.6–1.2%), or wheat (0.5–1.0%) (Demirbas 2005).
2.5 Minerals
The majority of proso millet seed mineral is present in its pericarp, aleurone layer, and germ. The mineral content varies from 1.5% to 4.2% (Ravindran 1992; Kalinová 2007), which is higher than that in wheat grain (1.5–2.0%). The majority of minerals in proso millet are slightly higher or similar to those of other cereals (Table 2). The common food processing and dehulling reduces the mineral content to around 27–53% (Kalinová 2007). This reduction commensurate with the dehulling intensity. Proso millet is deficient in calcium, but the grain contains a high amount of phosphorus. However, the bondage of phosphorus with phytates considerably interferes with its bioavailability. Proso millet is abundant in potassium, iron, and manganese (19.5–20.6 mg/100 g) (Ravindran 1992). Its grain is almost devoid (0.01 g/100 g) of sodium. Proso millet is an excellent source of zinc, copper, and boron as well (11.6 mg/kg) (Kalinová 2007).
2.6 Vitamins
The dehulled grain of proso millet is rich in vitamin B1, thiamine (0.42–0.80 mg/100 g); B2, riboflavin (0.22–0.40 mg/100 g); B3, niacin (1.55–3.7 mg/100 g); B6, pyridoxin (0.52–0.80 mg/100 g); and E, tocopherol (0.1–2.60 mg/100 g) (Table 3). The levels of vitamins B1 and B2 are twice that of rice, wheat, or barley. Most of the vitamins are deposited in the germ and aleurone layer and therefore, decortication considerably decreases their content. The color of dehulled grains is correlated to carotene and xanthophyll content. The carotene content is relatively stable and does not vary due to growing conditions such as soil type, level of fertilization, and interannual weather changes (Kalinová 2007). The unrefined fat extract of proso millet grain contains 8.3 to 10.5 mg of vitamin A and 87 to 96 mg of vitamin E per 100 g (Tripathi et al. 2021).
2.7 Antinutritional Factors
Knowledge of antinutritional factors such as tannins, phytates, oxalates, and enzyme inhibitors is essential to predict the nutritional value of products. Phytates affect the bioavailability of certain minerals, particularly multivalent cations including P, Ca, Mg Fe, and Zn, by binding with them. Phytic acid forms a complex with most of the free P (~67.3%) in the millets (Ravindran 1992). The phytic acid content in proso millet varies from 0.17 to 0.61 g/100 g, which is higher than that in polished rice but lower than that in wheat. Dehulling reduces the phytate content by 17–24% and thus makes more P bioavailable (Lorenz 1983).
3 Secondary Metabolites in Proso Millet
Like other millets and cereals, proso millet grain contains many secondary metabolites, which reportedly have beneficial effects on human health. The following section summarizes the secondary metabolites in proso millet.
3.1 Phenolic Compounds
Polyphenols are the major group of phytochemicals in plants, and they have numerous health benefits. Polyphenols in cereals have an adverse influence on color, flavor, and nutritional quality. These compounds are mostly localized in the aleurone layers of the grain. The polyphenol content in plants is influenced by factors such as plant part, developmental stage, and environment. Total phenolic compounds in proso millet are about 0.05–0.10 mg per 100 g of catechin equivalents, on dry basis. The bound polyphenols (1% HCl extractable) in proso millet were (2.21 ± 0.01), which was significantly lower than that in other millets such as kodo millet (81.64 ± 0.15), foxtail millet (11.59 ± 0.23), little millet (9.64 ± 0.28), pearl millet (9.14 ± 0.17), and finger millet (3.83 ± 0.18). Bound phenolic compounds have antioxidant, anti-obesity, anti-diabetic, antimutagenic, anticarcinogenic, antimicrobial, and antiviral properties (Tripathi et al. 2021).
3.2 Flavonoids
Flavonoids are a group of plant secondary metabolites with a general structure of a 15-carbon skeleton. Flavonoids such as catechin, quercetin, anthocyanin, tannin, etc. have human health-promoting properties, owing to their pharmacological activities such as radical scavenging. There are no reports of flavonoids in proso millet. However, a few reports are there for other millets. Finger millet leaves are known to contain eight types of flavones: vitexin, isovitexin, saponarin, violanthin, orientin, iso-orientin, lucenin-1, and tricin. Pearl millet contains glucosylvitexin, glucosylorientin, and vitexin in a ratio of 29:11:4 (Kalinová 2007). Finger millet is reportedly the only millet that contains condensed tannins.
3.3 Carotenoids
Carotenoids are well-known for their provitamin A activity. Carotenoids prevent various diseases because they act as antioxidants. A recent report showed that the carotenoid content in edible millet flour varied from 78 to 366 μg/100 g with an average of 366, 199, 78, 173, and 366 μg/100 g in proso, finger, little, and foxtail millets, respectively (Asharani et al. 2010). The carotenoid contents of millets were comparable to that of wheat (150–200 μg/100 g) and sorghum (180–230 μg/100 g) but significantly lower than that in maize (1800–5500 μg/100 g) (Tripathi et al. 2021).
3.4 Phytosterols
Phytosterols, which are desmethyl sterols, are essential structural and functional components of plant cells. Phytosterol esters can potentially reduce the blood serum LDL (low-density lipoprotein) cholesterol levels down to 14% but do not affect HDL (high-density lipoprotein) levels. The risk of heart disease can be reduced by up to 40% by including proso millet in the regular diet depending on age and some other factors. Other than finger millet, there are no reports of phytosterols in proso and other millets. The sterol content of finger millet was reported to be 0.149% on a seed weight basis, whereas other millets contain only a trace amount. Phytosterol content of sorghum and corn was estimated to be 0.5 mg/g and 0.9 mg/g (Singh et al. 2003).
4 Proso Millet Grain for Human Health
Proso millet confer many beneficial effects to human health. Some of these effects are backed by more scientific evidence than others. Several epidemiological studies have shown that regular consumption of millets and their products can ameliorate the affect of chronic diseases such as diabetes, cardiovascular disease, cancers, and all-cause mortality (Table 4).
4.1 Antioxidant
Millets including proso millet are abundant in both soluble and insoluble bound polyphenols, which have high antioxidant potential with metal chelating and reducing powers (Saleh et al. 2013). Several antioxidants present in millets neutralize free radicals (reactive oxygen species) and boost overall immunity and human health. The free radicals can lead to oxidative stress, which in turn are associated with conditions like cancer, arthritis, respiratory, and compromised immunity. Thus, consumption of millets can improve human health by scavenging free radicals from the liver and kidney.
4.2 Antidiabetic
Diabetes mellitus causes several body disorders, characterized by high blood sugar levels with an imbalance in protein, carbohydrate, and lipid metabolism. Consumption of whole millet grains has been clinically proven to lower diabetic effects (Saleh et al. 2013). Rao et al. (2017) reported that regular consumption of millets helps prevent diabetes due to the higher level of magnesium in these small grains, which in turn enhances the efficiency of insulin and glucose receptors. Furthermore, millet-based diets have been reported for their antioxidant potential, nerve growth factor production, and wound healing properties (Rao et al. 2017).
4.3 Cardiovascular Diseases
Millets are abundant in magnesium, potassium, lignans, antioxidants, and fibers, which can reduce the risk of heart strokes, high blood pressure, low-density cholesterol, and atherosclerosis. Rao et al. (2017) reported that the addition of barnyard, proso, and finger millets in the diet increased the blood plasma levels of adiponectin, high-density lipoprotein (HDL), and reduced triglycerides in hyperlipidemia. Therefore, millets, including proso, finger, barnyard, foxtail, and pearl millet, can significantly lower hyperlipidemia, hyperglycemia, triglycerides, and non-HDL and reduce the risk of cardiovascular diseases (Rao et al. 2017).
4.4 Anticancer
Cancer is characterized by changes at the cellular level due to uncontrolled cell growth and division. Millets generally have high levels of phenolic compounds such as tannins, fibers, and other phytonutrients than other cereals. These compounds improve health status and lower the risks of colon, breast, and esophageal cancers (Saleh et al. 2013). It is recommended that a daily ingestion of 30 g of fiber may reduce the risk of breast cancer chances by 50% (Rao et al. 2017).
4.5 Antiaging
Millets, including proso millet, are nutrient-rich functional food, enriched with high levels of micronutrients, vitamins, minerals, phenolic compounds, fibers, and antioxidants. These compounds are helpful in inhibiting glycation and cross-linking of collagen which is responsible for aging (Saleh et al. 2013).
4.6 Gluten Sensitivity
Celiac disease and gluten sensitivity are caused by gluten in organisms that are genetically susceptible to these conditions. The disease affects 0.6–1% of the population worldwide. Other food grains are amaranth, buckwheat, quinoa, corn, and sorghum functional and nutraceutical food. Millets have a lot of potential in the production of gluten-free food and beverages targeting people suffering from gluten sensitivity and celiac disease (Saleh et al. 2013).
5 Genetic Variation of Seed Components
Genetic variation is crucial for the adaptability and survivability of any species. From a breeding perspective, genetic variation ensures the selection of superior individuals (plants or animals) with improved characteristics from a population. A very limited number of studies have been conducted to assess the variation in seed composition in proso millet. Ravindran (1992) studied the amino acid composition and a few other traits of six proso millet varieties. He reported small differences in amino acid content among the varieties (Ravindran 1992). Kalinova and Moudry (2006) assessed the crude protein and amino acid content in seven proso millet varieties. In contrast to Ravindran’s findings, they observed significant varietal differences in seed content of the amino acid lysine, valine, isoleucine, phenylalanine, aspartate, glycine, tyrosine, histidine, and arginine (Kalinova and Moudry 2006). In another study, a varietal difference in phenolic compounds and antioxidant capacity in whole millet grains was reported (Chandrasekara and Shahidi 2011). Vetriventhan and Upadhyaya (2018) studied 200 proso millet accessions to assess the diversity of grain nutritional traits including seed protein, calcium (Ca), iron (Fe), and zinc (Zn). Large variations for all four grain-nutritional traits were observed in their study. More research on the genetic variation of seed components, including secondary metabolites, needs to be conducted for generating resources for genome-wide association studies (GWAS) and developing varieties with improved nutrient profiles (Vetriventhan and Upadhyaya 2018).
6 Breeding for Grain Quality Improvement
Plant breeding is the process of creating and selecting superior genotypes with enhanced yield, abiotic and biotic stress tolerance, and improved nutritional quality. Breeding programs dedicated to developing genetically improved varieties of proso millet were established in several countries. China, the USA, India, Russia, and Kenya are notable countries that have been instrumental in continuing efforts for the genetic improvement of proso millet (Gomashe 2017). Considerable progress has been made in generating high-yielding varieties with desirable maturity, seed color, plant height, and other yield-related morpho-agronomic traits. However, efforts to develop superior proso millet cultivars enriched with nutrients and health-promoting metabolites are very limited. With the growing demand for proso millet in the ever-expanding human healthy food market, it is imperative that breeding programs focused on improving nutrient and metabolite profiles of the high-yielding varieties are established to promote this crop as a functional food. Common breeding approaches and a couple of biotechnological tools to complement conventional breeding in accelerating variety development efforts are discussed further down.
6.1 Conventional and Wide Hybridization
The majority of proso millet varieties of North America were developed through conventional breeding methods (crossing followed by selection of progenies) using a limited pool of parental lines through conventional hybridization. The resulting narrow genetic base may lead to diminished performance of the varieties in terms of resistance to abiotic and biotic stresses over time. Besides, the variation of seed quality traits is not very large, which makes genetic improvement of seed composition through conventional breeding approaches even tougher. Nevertheless, the conventional method was still useful in developing specialty proso millet varieties for human food uses. One such example was ‘Plateau’, the first and only waxy proso millet variety in North America (Santra et al. 2015). The variety was developed via artificial pollination between PI 436626 (Lung Shu 18), a Chinese waxy accession, and ‘Huntsman’, a high-yielding US variety. Similar market-driven strategies can be adopted to develop new high-yielding cultivars enriched with essential nutrients and healthy metabolites to market them as nutraceuticals.
The gene pool of wild relatives of proso millet can be a large repository of novel traits or genes that can be transferred via introgression into the existing cultivars (Santra et al. 2019). Wide hybridization is a promising prospect for improving grain nutritional quality by introducing genes from the wild relatives such as P. sumatrense and P. repens to improve the metabolic profile of the cultivars.
6.2 Mutation Breeding
Mutation breeding involves deliberate induction of mutation by artificial mutagenesis and breeding for genetic variations that do not exist in the gene pool. Mutation-inducing agents (mutagens) can be physical, ionizing (Χ-rays and γ-rays), and non-ionizing radiations (ultraviolet rays) or chemical agents such as ethylmethane sulfonate (EMS), methyl methane sulfonate (MMS), diethylsulfate (DES), and nitrosoguanidine (NS). EMS is the most widely used chemical mutagen due to its effectiveness and ease of use. Proso millet seeds treated with 0.2% EMS was found to improve panicle filling, grain shattering tolerance, and grain yield in the M2 progeny (Singode et al. 2018). Recently, Francis et al. (2022) used different doses of gamma radiation to develop phenotypic mutations for various traits, including plant height, panicle shape, compactness, seed color, and lodging resistance. They were able to isolate eight high-yielding mutant families with significantly improved grain yield (GY) and fodder yield (FY) (Francis et al. 2022). More research on artificial mutagenesis is absolutely required in proso millet. Mutation breeding is a viable technique to overcome poor combining ability and narrow genetic based for improving grain nutrient and metabolite profile (Santra et al. 2019).
6.3 Doubled Haploids and Transgenics
Doubled haploid (DH) and transgenic technologies are two commonly used tissue culture-based biotechnology tools for the genetic improvement of crops. The DH technology produces 100% homozygous genotypes using haploid gametes (pollen or egg). Transgenic technology uses genetic engineering to insert one or two specific genes from other plants. Conventional plant breeding via pure-line selection normally takes 11–13 years to develop a new variety as it usually involves 5–6 generations of inbreeding to generate homogeneous breeding lines with >98% homozygosity before multi-year yield assessment at multiple sites. The desirable 100% homozygosity can be achieved in a much shorter time by implementing doubled haploid technology (Khound et al. 2013; Santra et al. 2019). Only a few reports on successful tissue regeneration have been reported in proso millet. In 1982, Heyser and Nobors reported successful callus induction and shoot regeneration from a variety of explants by manipulating auxin levels in Linsmaier and Skoog (L and S) medium (Heyser and Nabors 1982). Bobkov and Suvorova (2012) studied the efficiency of anther culture technique in proso millet for embryogenic callus induction and regeneration. Heat (32 °C) and cold (4 °C) were observed to successfully induce callus formation and regeneration of the explants (Bobkov and Suvorova 2012). For several decades, genetic engineering has been used to introduce foreign genes expressing novel traits into a host genome or to knock out genes with deleterious effects. Genetic improvement of crops through genetic engineering relies on the development of an efficient regeneration method and a robust transformation system (Kumar et al. 2016). There has been no report on the generation of transgenic proso millet via genetic transformation so far. The DH and genetic engineering technologies can be used for developing proso millet varieties enriched with health-promoting bioactive compounds and no or minimum antinutrients. Therefore, more studies on efficient tissue culture and transformation systems are warranted for facilitating genetic engineering-assisted genetic improvement of proso millet.
7 Omics for Improving Grain Nutritional Quality
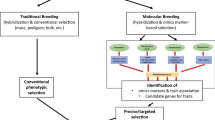
Biology works as a “system” rather than as an individual part separately. All the parts of biology work together in harmony to manifest overall biological functions. Many biochemical compounds are involved in this intrinsic process. Similarly, all the nutrients and antinutrients in food work together in harmony to manifest their effect on human health and nutrition. Therefore, it is very important to study proso millet seed nutrients quality and quantity, genes, biosynthesis pathways, regulation, and interactions together rather than separately. That means all aspects of proso millet seed nutraceuticals must be studied following the “omics” approach. The term “neutrocetomics” is used to indicate the combined study of nutraceuticals and omics. Proso millet being a minor crop, its “omics” areas of research are not as rich as other major crops such as corn, wheat, rice, and soybean. However, in recent years a lot of omics research has been done in proso millet, and more is being conducted due to its increasing importance for human health. Details of proso millet “omics” research were published in a recent comprehensive review paper (Khound and Santra 2020). The following section summarizes various fields of “omics” in proso millet and their relevance to its “nutraceutomics” (Fig. 3).

Schematic representation of the applications of traditional breeding methods and tools, as well as omics-based and post-genomic breeding strategies to the genetic improvement of proso millet
7.1 Genomics
Genomics is the branch of omics that tackles large-scale studies on the structural and functional aspects of the genome to understand the genetic and molecular underpinning of various biological processes. Initial genomic studies were primarily focused on genome size and physical as well as genetic mapping of organisms. Next-generation sequencing (NGS) marked a new era of genomics by enabling researchers to sequence, assemble, and analyze the genomes of important crops at lower costs and in a shorter time (Khound and Santra 2020).
7.1.1 Molecular Markers
Molecular markers are important genomic resources that have been widely used in genetic mapping, genetic diversity, and taxonomic and population genetic studies of crops. A limited number of molecular markers were developed in proso millet as this crop is grossly under-researched. The molecular markers identified in proso millet so far include amplified fragment length polymorphism (AFLP), random amplified polymorphic DNA (RAPD), inter-simple sequence repeat (ISSR), cleaved amplified polymorphic DNA (CAP), simple sequence repeat (SSR), and single nucleotide polymorphism (SNP) markers. These molecular markers were primarily used for evaluating the genetic diversity of the proso millet germplasm (Santra et al. 2019).
SSRs are preferable for genetic studies as they are abundant, evenly distributed, multi-allelic, codominant, highly polymorphic, easy to score, and highly reproducible (Khound and Santra 2020). The earliest SSR-based genetic studies were conducted using SSRs from other species such as rice, wheat, oat, barley, and switchgrass. The first proso millet-specific SSRs were identified by Cho and coworkers (Cho et al. 2010). Twenty-five polymorphic SSRs were developed from the genomic DNA of 50 diverse proso millet genotypes. These SSRs were used to evaluate the genetic diversity of Chinese proso millet accessions (Liu et al. 2016).
SNP markers are ideal for genetic studies, genetic map constriction, and genomic selection as they occur abundantly in the genome (Khound and Santra 2020). These markers are especially well suited for constructing linkage maps as they are cost-effective, high-throughput, and more efficient compared to other DNA markers (Rajput et al. 2016). Several reports on SNP identification in proso millet have been published. Rajput et al. identified 833 high-quality biallelic genotype-by-sequencing (GBS)-SNPs from genotyping of 93 recombinant inbred lines (RILs). These GBS-SNPs were used for constructing the first proso millet linkage map (details later in the quantitative trait locus (QTL) mapping section) (Rajput et al. 2016). Wang and her coworkers evaluated the allelic diversity of two waxy genes, viz., Wx-L and Wx-S, in 132 proso millet accessions from 12 provinces of China. In their study, Wang et al. (2018) identified six SNPs at the Wx-L locus after sequencing the PCR amplicons of all the accessions. Johnson and his team used GBS to generate 1882 filtered SNPs to conduct genome-wide population genetic studies in proso millet. The SNP markers were found to accurately represent genetic variation within the population (Johnson et al. 2019). In 2021, Boukail et al. (2021) developed 2412 SNPs from 88 proso millet accessions with RAD-seq. The identified SNPs were used for conducting GWAS to detect marker-trait associations for several agronomic and seed morphology traits (more details are covered in Sect. 8) (Boukail et al. 2021). In another recently published report, 126,822 filtered SNPs were identified using specific-locus amplified fragment sequencing (SLAF-seq) of 106 accessions (Li et al. 2021). More recently, Khound et al. (2022) developed 972,863 high-quality biallelic SNPs from low-pass genome sequencing of 85diverse proso millet genotypes of the USDA gene bank. They employed those SNPs to study the population structure and phylogenetic relationships among the genotypes (Khound et al. 2022).
7.1.2 QTL Mapping
Quantitative trait locus (QTL) mapping is routinely used in many crops for detecting quantitative trait loci (QTLs) associated with key traits due to the abundance of versatile DNA markers and statistical models and methods. QTLs for a wide array of traits have been reported in many major and minor crops including wheat, rice, maize, sorghum, millet, amaranth, quinoa, oat, and rye (Yabe and Iwata 2020). Among millets, pearl millet and foxtail millet have the major share of published reports on QTL mapping. In proso millet, very little research has been done on genetic mapping of QTLs compared to major crops such as rice, wheat, maize, soybean, and sorghum. This is possibly because of the minor crop status and inadequate resources available to proso millet geneticists. The first-ever genetic linkage map of proso millet was published in 2016 by Rajput et al. They used 833 GBS-SNPs and 93 RILs. Several QTLs and linked SNP markers for a few morpho-agronomic traits were identified (Rajput et al. 2016). However, no QTLs for proso millet seed components such as starch, protein, minerals, vitamins, and other bioactive compounds have been identified in proso millet to date.
7.1.3 Whole-Genome Assembly
Developing a whole-genome assembly (WGA) is a crucial step in genome-wide molecular studies as this creates opportunities for exploring complex molecular structures and functions in an organism. Information on genome assembly is an invaluable resource for genomic-assisted breeding for crop improvement. Proso millet was the fourth millet to have a published whole-genome assembly after foxtail millet, pearl millet, and finger millet (Khound and Santra 2020). In 2019, Zou et al. developed the complete proso millet genome sequence using various NGS tools. They also identified 55,930 protein-coding and 339 microRNA genes (Zou et al. 2019). In the same year, Shi et al. reported a near-complete assembly of the proso millet genome. The authors generated 18 super scaffolds covering approximately 96% of the estimated genome. Moreover, they were able to annotate 63,671 protein-coding genes in the proso millet genome (Shi et al. 2019).
7.1.4 TILLING
Another modern approach that could be applied for improving grain nutritional quality of proso millet is the TILLING (targeted induced local lesions in genomes) method. This is an advanced method of detecting beneficial mutations or alleles that allows rapid identification of induced gene mutations within a mutagenized population via heteroduplex analysis. The initial step of TILLING involves creating a mutagenized population, which is subsequently assessed for detecting useful gene mutations that could be linked to important phenotypes. The mutations within the mutagenized population are usually detected by one of the three methods, viz., LI-COR method (uses CEL 1 enzyme), high-resolution melting (HRM) method, and NGS method. TILLING has been used for detecting useful mutations in a wide array of crops including wheat, rice, sorghum, and maize (Irshad et al. 2020). Tilling has been successfully employed to identify mutations in genes associated with starch synthesis, plant architecture, and disease resistance in several crop species. Another related method, namely, EcoTILLING, is used to detect naturally occurring polymorphisms within a population. The polymorphisms identified could be used to study the phylogenetic diversity within the population. This technique can also be used to detect novel allelic variations that can be exploited for genetic improvement. EcoTILLING has been utilized in different crops including Arabidopsis, chickpea (seed weight), wheat (disease susceptibility), rice (drought tolerance, starch synthesis), and soybean (seed protein) (Irshad et al. 2020).
7.1.5 Allele Mining
A large number of beneficial alleles are present in plant genetic resources, which remain unutilized as they were abandoned during evolution and domestication. Introgression of these natural genetic variations has the potential to improve the performance of available cultivars. The recent surge of genomic data accelerated the discovery and annotation of novel genes and loci linked to essential agronomic traits in many crop species. The idea of identifying alleles in the annotated genes led to the concept of allele mining in plants. Allele mining refers to the approach of dissecting naturally occurring variations at candidate genes or loci associated with major agronomic traits with the help of modern genomic tools (Kumar et al. 2010). Two primary approaches for identifying alleles are EcoTILLING and sequence-based allele mining. As discussed in the previous section, EcoTILLING is used to identify naturally occurring allelic variants present in the primary and secondary gene pools. In a sequence-based approach, alleles of diverse genotypes are amplified by PCR and subsequently sequenced. Allele mining was used in identifying alleles for important seed quality traits including Amy32b(α-amylase gene) and Gpc-B1 (gene for grain protein content) in barley, wx locus (waxy gene) in rice, and Wx-A1 (waxy gene) in wheat (Kumar et al. 2010). The prospect of using allele mining for detecting novel alleles of genes is relatively unexplored in millets. The availability of whole-genome assemblies, high-quality molecular markers, and the gradually reducing cost of sequencing is expected to result in more studies for identifying novel genes and loci. That would encourage more efforts for the mining of candidate genes for some key agronomic and grain quality traits leading to the detection of useful alleles for application in molecular breeding.
7.2 Transcriptomics
A handful of reports on transcriptome studies investigating biological processes have been published in proso millet. Yue et al. (2016) used Illumina sequence reads from two proso millet accessions, viz., Yumi No.2 and Yumi No.3, to assemble a proso genome transcriptome. From the 113,643 unigenes assembled, 62,543 contigs were assigned to 315 gene ontology (GO) categories. Additionally, 15,514 unigenes could be mapped into 202 Kyoto Encyclopedia of Genes and Genomes (KEGG) clusters, and 51,020 unigenes were mapped to 25 clusters of orthologous groups (COG) categories. The most represented KEGG pathways included metabolic pathways (25.65%), biosynthesis of secondary metabolites (10.71%), and biosynthesis of amino acids (3.57%) (Yue et al. 2016). In 2017, Hou et al. developed a transcriptome assembly using Illumina sequence reads from a single genotype of proso millet named Neimenggu-Y1. They were able to identify 25,341 unigenes out of which 5170 (20.4%) could be mapped to 146 KEGG pathways. GO annotation with 2936 tissue-specific genes resulted in three subcategories- biological processes, molecular functions, and cellular components (Hou et al. 2017). Zhang et al. (2019) published a comparative analysis of transcriptome associated with drought tolerance in two genotypes, viz., Neimi 5 and Jinshu 6. Drought stress resulted in 833 and 2166 DEGs in Jinshu 6 and Neimi 5, respectively. Some of the DEGs could be mapped to some key KEGG pathways including carbon metabolism, phenylpropanoid biosynthesis, and amino acid biosynthesis (Zhang et al. 2019). The unigenes that were mapped to the key KEGG pathways in the above reports should be explored further to identify their roles in those pathways. This information could be useful in targeting specific genes for enriching proso millet with useful nutraceuticals or inhibiting the biosynthesis of antinutrients.
7.3 Proteomics
There is an abundance of reports on proteomics research for the major crops including rice, corn, soybean, wheat, and potato. However, like other omics disciplines, the number of published reports pertaining to proteomics in proso millet and other millet is very meager (Khound and Santra 2020). Perhaps the first proteomics analysis in proso millet was done on a 2500-year-old starch food in China. Comparative proteomics analysis between the ancient sour dough bread and a few other reference cereals identified proso millet and barley as the ingredients of the ancient bread (Shevchenko et al. 2014). In 2017, Roy and co-workers reported the seed protein analysis of four Korean proso millet varieties using two-dimensional (2-D) electrophoresis and mass fingerprinting to map the seed proteins and determine their functional properties. They detected 1152 differentially expressed proteins, out of which 26 reproducible proteins were further analyzed using matrix-assisted laser desorption/ionization time-of-flight/time-of-flight mass spectrometry (MALDI-TOF-TOF/MS). Two of the 26 proteins were found to be upregulated in all the cultivars, while 13 were upregulated and 11 were downregulated in 2 proso millet cultivars. The authors opined that the differential expression of the proteins in the four proso millet cultivars was possibly variety-specific (Roy et al. 2017).
7.4 Metabolomics
There are only a couple of available reports on the metabolomics of proso millet. The first published study on proso millet metabolomics used gas chromatography-time-of-flight mass spectrometry (GC-TOFMS) to evaluate gain quality. Kim et al. (2013) studied the primary metabolites and phenolic acids of the matured grains of three Korean proso millet varieties, viz., ‘Joongback’, ‘Joongjuk’, and ‘Hwangguem’. They were able to identify 48 metabolites from the grains, which included 43 primary metabolites and 5 phenolic acids. The mature grains of the variety ‘Joongjuk’ contained significantly higher levels of phenolic acids than the other two varieties. This makes this variety a suitable candidate for further evaluations and genetic improvement as a nutraceutical (Kim et al. 2013).
In a relatively recent publication, 172 metabolites and 3 cooking quality traits were compared between conventionally and organically grown seeds of two proso millet varieties. There was no difference in the metabolite profiles between the conventionally and organically grown gains, except in the levels of some carbohydrates such as glucose and fructose, which were higher in the organically grown grain. The variations observed in the metabolite content could be primarily attributed to the variety (Liang et al. 2018a). These findings emphasize the importance of variety selection for developing proso millet varieties for nutraceutical use.
7.5 Phenomics
A phenome can be described as the complete set of phenotypic traits expressed by a cell, tissue, or organism, and the field of studies on the phenome is known as phenomics. Establishing the relationship between the genotypes and phenotypes is a major breeding objective of any crop improvement program. Traditional phenotyping approaches are usually laborious, time-consuming, expensive, and mostly destructive (Santra et al. 2019). It is important to determine more phenotype-to-genotype relationships to develop reliable predictive models for predicting a full array of phenotypes of a genotype (Gustin and Settles 2015). In recent years, high-throughput phenotyping techniques and tools including high-resolution imaging, spectroscopy, robotics, and powerful algorithms have been developed to push plant phenotyping to the next level. Different high-throughput phenotyping approaches have been used to study a variety of phenotypic traits such as plant growth, biomass, leaf morphology, maturity, and nutrient status (Santra et al. 2019). To the best of our knowledge, the first-ever report on high-throughput phenotyping in proso millet was reported by Zhao et al. who used UAV-based imaging for heading percentage detection in proso millet (Zhao et al. 2022).
A more niche area of phenomics, for example, “seed phenomics,” could be developed in proso millet to specifically study various seed characteristics (Gustin and Settles 2015). This will require the integration of various imaging technologies, spectroscopy, and multiple omics such as genomics, transcriptomics, proteomics, and metabolomics. Various imaging techniques and rapid, nondestructive spectroscopic techniques, such as near-infrared (NIR) spectroscopy and nuclear magnetic resonance (NMR) spectroscopy, are routinely used to determine various seed traits such as shape, size, color, and chemical composition. To the best of our knowledge, there is no report or ongoing research on proso millet seed phenome, which is very important for genetic improvement of nutraceuticals values in proso millet.
8 Post-Genomic Approaches for Improving Seed Nutritional Quality
The following sections summarize the current status of proso millet “omics” resources available to the scientists working in the field of proso millet nutraceutomics. The following sections address post-genomic omics approaches for genetic manipulation of proso millet seed nutraceuticals.
8.1 Genome-Wide Association Studies (GWAS)
Genome-wide association study (GWAS) or association mapping (AM) involves the detection of an association between DNA marker(s) and a trait of interest based on the principle of linkage disequilibrium (LD). This is achieved through large-scale genotyping of germplasm panels or breeding populations exhibiting contrasting phenotypes across different environments. Several GWAS have been conducted in millets to uncover the marker-trait association for some important traits. However, only a few reports of GWAS for seed nutritional traits are available. The only available report on GWAS in proso millet to date was published in 2021 by Boukail et al. They used a global collection of 88 varieties and landraces to identify marker trait associations (MTAs) for seed morphological traits. They identified 2412 high-quality SNPs using restriction site-associated DNA sequencing (RAD-seq). These SNPs were used for GWAS for seed traits, such as seed length (SL), seed width (SW), seed perimeter (SP), and seed color (RGB) as well as agronomical traits. They identified MTAs for the seed and agronomic traits (Boukail et al. 2021). The SNPs that were found to have a strong association with the agronomic and seed traits could be strong candidates for marker-assisted selection (MAS) in proso millet breeding programs. Having said that, no report on GWAS for nutraceutical traits is available in proso millet yet. This warrants the initiation of genome-scale studies to identify MTAs for some key seed quality traits to accelerate the development of proso millet varieties with impeccable agronomic qualities and healthy seed components.
8.2 Genomic Selection (GS)
Genomic selection (GS) is the breeding approach of using genome-wide high-density markers to facilitate rapid selection of suitable candidates for breeding (Srivastava et al. 2020). GS is still in the nascent stage in millets as very few GS studies have been conducted so far due to the limited availability of genomic resources. Varshney et al. (2017) utilized whole-genome resequencing (WGRS) data for performing GS to predict the grain yield of pearl millet under four different stress scenarios across environments. They used to analyze the grain yield of 64 hybrids with 302,110 SNPs to identify promising hybrid combinations for hybrid production (Varshney et al. 2017). In another study, Liang and his coauthors (2018b) evaluated four genomic selection schemes using two genotyping strategies, namely, RAD-seq and tunable genotyping by sequencing (tGBS) in pearl millet. The authors observed that for traits with significant mid-parent heterosis, the inbred phenotypic data moderately improved genomic predication of the hybrid genomic estimated breeding values when the trait values of the inbred and hybrid lines were scored relative to the mean trait values of the corresponding populations (Liang et al. 2018b). Similar GS studies can be conducted in proso millet for major morpho-agronomic and seed quality traits to accelerate genetic improvement of breeding populations in a time-efficient and cost-effective manner.
8.3 Genome Editing
Genome editing is a relatively newer technique adopted by plant breeders to develop new and improved varieties of crops. Rather than introducing transgene(s) randomly into the host genome, genome editing used sequence-specific nucleases (SSNs) to induce targeted and precise nucleotide sequence changes to the genome (Santra et al. 2019). Genome editing tools have been successfully used to introduce genes into major cereals including rice, wheat, and maize (Ceasar 2022). The most commonly used genome editing tools are clustered regularly interspersed short palindromic repeats-CRISPR-associated nucleases (CRISPR-Cas) system, transcription activator-like effector nucleases (TALENs), and zinc-finger nucleases (ZFNs) (Santra et al. 2019; Ceasar 2022).
There are numerous reports on genome editing in major cereals, especially rice, wheat, maize, and barley. However, genome editing is still a relatively unexplored territory in millets. One example of the successful use of genome editing tools in improving grain quality is the knock-out of three key genes associated with phytic acid (PA) biosynthesis in maize. PA naturally occurs in the grains of many cereal crops, including proso millet. It is considered an antinutrient as it is largely indigestible and may cause environmental pollution. TALEN and CRISPR-Cas9 systems were used to induce mutations in the genes, viz., ZmIPK, ZmIPK1A, and ZmMRP4, encoding enzymes that catalyze three steps in PA biosynthesis (Liang et al. 2014). There are only two reports on successful CRISPR-Cas9-mediated genome modification in foxtail millet (Ceasar 2022). Similar strategies can be implemented in proso millet for improving grain quality by reducing the levels of antinutrients and enriching with health-promoting compounds such as carotenoids. However, reliable, reproducible, and robust micropropagation and transformation systems need to be developed for this crop in order to accomplish this.
9 Conclusion and Future Prospects
Compared to the major crops such as rice, wheat, corn, and soybean, there is a dearth of genomic resources available for the genetic improvement of proso millet. Especially, little to no progress has been made in exploring genes or QTLs linked to grain nutraceutical traits. Therefore, there is a need for devising strategies for developing and harnessing genomic resources in the identification of genes and QTLs associated with seed components in proso millet. This will facilitate omics-assisted breeding of this ancient crop thereby enabling rapid and precise genetic improvement for various agronomic and seed quality traits. Therefore, the global millet researchers must work together to take advantage of such technological advancement in the “omics” research to advance the field of “nutraceutomics” in proso millet. Interdisciplinary research, extension, and promotion are essential for using proso millet in the human food market.
The future prospects of proso millet along with other climate-resilient millets are vast for the food and nutritional security of the global population in the current century and future. The global climate is deteriorating exponentially with the acute pressure of population increase and farmland reduction. All seven millets have great potential to address the mammoth global challenge. A strong collaboration among the millet geneticists and breeders across the globe will be mandatory for the successful implementation of the “omics” in proso millet “nutraceutomics” research and applications for human health and nutritional security in the changing climate, especially in the climate-fragile countries in the world.
References
Akharume F, Santra D, Adedeji A (2020) Physicochemical and functional properties of proso millet storage protein fractions. Food Hydrocoll 108:105497. https://doi.org/10.1016/j.foodhyd.2019.105497
Anderson RL, Bowman RA, Nielsen DC et al (1999) Alternative crop rotations for the central Great Plains. J Prod Agric 12:95–99. https://doi.org/10.2134/jpa1999.0095
Argos P, Pedersen K, Marks MD, Larkins BA (1982) A structural model for maize zein proteins. J Biol Chem 257:9984–9990. https://doi.org/10.1016/S0021-9258(18)33974-7
Asharani VT, Jayadeep A, Malleshi NG (2010) Natural antioxidants in edible flours of selected small millets. Int J Food Prop 13:41–50. https://doi.org/10.1080/10942910802163105
Baltensperger DD (2002) Progress with proso, pearl and other millets. In: Janick J, Whipkey A (eds) Trends in new crops and new uses. ASHS Press, Alexandria, VA, pp 100–103
Bangar SP, Ashogbon AO, Dhull SB et al (2021) Proso-millet starch: properties, functionality, and applications. Int J Biol Macromol 190:960–968. https://doi.org/10.1016/j.ijbiomac.2021.09.064
Bobkov S, Suvorova G (2012) Temperature stress in anther culture of proso millet (Panicum miliaceum L.). In: Yan C, Baili F (eds) Proceedings of the 1st international broomcorn millet symposium: advances in broomcorn millet research, pp 82–88
Boukail S, Macharia M, Miculan M et al (2021) Genome wide association study of agronomic and seed traits in a world collection of proso millet (Panicum miliaceum L.). BMC Plant Biol 21:330. https://doi.org/10.1186/s12870-021-03111-5
Ceasar A (2022) Genome-editing in millets: current knowledge and future perspectives. Mol Biol Rep 49:773–781. https://doi.org/10.1007/s11033-021-06975-w
Chandrasekara A, Shahidi F (2011) Inhibitory activities of soluble and bound millet seed phenolics on free radicals and reactive oxygen species. J Agric Food Chem 59:428–436. https://doi.org/10.1021/jf103896z
Cho Y-I, Chung J-W, Lee G-A et al (2010) Development and characterization of twenty-five new polymorphic microsatellite markers in proso millet (Panicum miliaceum L.). Genes Genom 32:267–273. https://doi.org/10.1007/s13258-010-0007-8
Das S, Khound R, Santra M, Santra D (2019) Beyond bird feed: proso millet for human health and environment. Agriculture 9:64. https://doi.org/10.3390/agriculture9030064
Demirbas A (2005) Glucan and mineral nutrient contents of cereals grown in Turkey. Food Chem 90:773–777. https://doi.org/10.1016/j.foodchem.2004.06.003
Dendy DAV (1995) Sorghum and Millets: Chemistry and Technology. AACC, St. Paul, Minnesota, 406 pp.
Francis N, Rajasekaran R, Krishnamoorthy I et al (2022) Gamma irradiation to induce beneficial mutants in proso millet (Panicum miliaceum L.): an underutilized food crop. Int J Radiat Biol 98:1277–1288. https://doi.org/10.1080/09553002.2022.2024292
Geervani P, Eggum BO (1989) Nutrient composition and protein quality of minor millets. Plant Foods Hum Nutr 39:201–208. https://doi.org/10.1007/BF01091900
Gomashe SS (2017) Proso millet, Panicum miliaceum (L.): genetic improvement and research needs. In: Patil JV (ed) Millets and sorghum. John Wiley & Sons, Ltd, Chichester, pp 150–169
Gulati P, Li A, Holding D et al (2017) Heating reduces proso millet protein digestibility via formation of hydrophobic aggregates. J Agric Food Chem 65:1952–1959. https://doi.org/10.1021/acs.jafc.6b05574
Gulati P, Jia S, Li A et al (2018) In vitro pepsin digestibility of cooked proso millet (Panicum miliaceum L.) and related species from around the world. J Agric Food Chem 66:7156–7164. https://doi.org/10.1021/acs.jafc.8b02315
Gustin JL, Settles AM (2015) Seed phenomics. In: Fritsche-Neto R, Borém A (eds) Phenomics. Springer International Publishing, Cham, pp 67–82
Habiyaremye C, Matanguihan JB, D’Alpoim Guedes J et al (2017) Proso millet (Panicum miliaceum L.) and its potential for cultivation in the Pacific Northwest, U.S.: a review. Front Plant Sci 7. https://doi.org/10.3389/fpls.2016.01961
Heyser JW, Nabors MW (1982) Regeneration of proso millet from embryogenic Calli derived from various plant Parts1. Crop Sci 22:1070–1074. https://doi.org/10.2135/cropsci1982.0011183X002200050043x
Hou S, Sun Z, Li Y et al (2017) Transcriptomic analysis, genic SSR development, and genetic diversity of proso millet (Panicum miliaceum; Poaceae). Appl Plant Sci 5:1600137. https://doi.org/10.3732/apps.1600137
Hunt HV, Badakshi F, Romanova O et al (2014) Reticulate evolution in Panicum (Poaceae): the origin of tetraploid broomcorn millet, P miliaceum. J Exp Bot 65:3165–3175. https://doi.org/10.1093/jxb/eru161
Irshad A, Guo H, Zhang S, Liu L (2020) TILLING in cereal crops for allele expansion and mutation detection by using modern sequencing technologies. Agronomy 10:405. https://doi.org/10.3390/agronomy10030405
Johnson M, Deshpande S, Vetriventhan M et al (2019) Genome-wide population structure analyses of three minor millets: Kodo millet, little millet, and proso millet. Plant Genome 12:190021. https://doi.org/10.3835/plantgenome2019.03.0021
Jones RW, Beckwith AC, Khoo U, Inglett GE (1970) Protein composition of proso millet. J Agric Food Chem 18:37–39. https://doi.org/10.1021/jf60167a003
Kalinová J (2007) Nutritionally important components of proso millet (Panicum miliaceum L.). FoodReview 1:91–100
Kalinova J, Moudry J (2006) Content and quality of protein in proso millet (Panicum miliaceum L.) varieties. Plant Foods Hum Nutr 61:43–47. https://doi.org/10.1007/s11130-006-0013-9
Kalra EK (2003) Nutraceutical-definition and introduction. AAPS PharmSci 5:27–28. https://doi.org/10.1208/ps050325
Khound R, Santra DK (2020) Omics for proso millet genetic improvement. Nucleus 63:241–247. https://doi.org/10.1007/s13237-020-00339-8
Khound R, Santra M, Baenziger PS, Santra DK (2013) Effect of cold-mediated pretreatment on microspore culture in winter and spring wheat. Am J Plant Sci 04:2259–2264. https://doi.org/10.4236/ajps.2013.411278
Khound R, Sun G, Mural RV et al (2022) SNP discovery in proso millet (Panicum miliaceum L.) using low-pass genome sequencing. Plant Direct 6(9):e447. https://doi.org/10.1002/pld3.447
Kim JK, Park S-Y, Yeo Y et al (2013) Metabolic profiling of millet (Panicum miliaceum) using gas chromatography–time-of-flight mass spectrometry (GC-TOFMS) for quality assessment. Plant Omics J 6:73–78
Kumar GR, Sakthivel K, Sundaram RM et al (2010) Allele mining in crops: prospects and potentials. Biotechnol Adv 28:451–461. https://doi.org/10.1016/j.biotechadv.2010.02.007
Kumar A, Metwal M, Kaur S et al (2016) Nutraceutical value of finger millet [Eleusine coracana (L.) Gaertn.], and their improvement using omics approaches. Front Plant Sci 7:934. https://doi.org/10.3389/fpls.2016.00934
Kumari SK, Thayumanavan B (1998) Characterization of starches of proso, foxtail, barnyard, kodo, and little millets. Plant Foods Hum Nutr 53:47–56
Ladabaum U, Mannalithara A, Myer PA et al (2014) Obesity, abdominal obesity, physical activity, and caloric intake in US adults: 1988 to 2010. Am J Med 127(8):717–727.
Li C, Liu M, Sun F et al (2021) Genetic divergence and population structure in weedy and cultivated broomcorn millets (Panicum miliaceum L.) revealed by specific-locus amplified fragment sequencing (SLAF-Seq). Front Plant Sci 12:688444. https://doi.org/10.3389/fpls.2021.688444
Liang Z, Zhang K, Chen K, Gao C (2014) Targeted mutagenesis in Zea mays using TALENs and the CRISPR/Cas system. JGG 41:63–68.
Liang K, Liang S, Lu L et al (2018a) Metabolic variation and cooking qualities of millet cultivars grown both organically and conventionally. Food Res Int 106:825–833. https://doi.org/10.1016/j.foodres.2018.01.023
Liang Z, Gupta SK, Yeh C-T et al (2018b) Phenotypic data from inbred parents can improve genomic prediction in pearl millet hybrids. G3 (Bethesda) 8:2513–2522. https://doi.org/10.1534/g3.118.200242
Liu M, Xu Y, He J et al (2016) Genetic diversity and population structure of broomcorn millet (Panicum miliaceum L.) cultivars and landraces in China based on microsatellite markers. Int J Mol Sci 17:370. https://doi.org/10.3390/ijms17030370
Lorenz K (1983) Tannins and phytate content in proso millets (Panicum miliaceum). Cereal Chem 60:424–426
Lu H, Zhang J, Liu K et al (2009) Earliest domestication of common millet (Panicum miliaceum) in East Asia extended to 10,000 years ago. Proc Natl Acad Sci U S A 106:7367–7372. https://doi.org/10.1073/pnas.0900158106
Majid A, Priyadarshini CGP (2020) Millet derived bioactive peptides: a review on their functional properties and health benefits. Crit Rev Food Sci Nutr 60:3342–3351. https://doi.org/10.1080/10408398.2019.1686342
Malik S (2015) Pearl millet-nutritional value and medicinal uses. IJARIIE 1(3):414–418
Matsushima N, Danno G, Takezawa H, Izumi Y (1997) Three-dimensional structure of maize a-zein proteins studied by small-angle X-ray scattering. Biochim Biophys Acta 1339:14–22
Nielsen DC, Calderón FJ (2011) Fallow effects on soil. In: Hatfield JL, Sauer TJ (eds) Soil management: building a stable base for agriculture. Soil Science Society of America, Madison, pp 287–300
Parameswaran KP, Thayumanavan B (1995) Homologies between prolamins of different minor millets. Plant Foods Hum Nutr 48:119–126. https://doi.org/10.1007/BF01088307
Rajput SG, Santra DK, Schnable J (2016) Mapping QTLs for morpho-agronomic traits in proso millet (Panicum miliaceum L.). Mol Breed 36:37. https://doi.org/10.1007/s11032-016-0460-4
Rao BD, Bhaskarachary K, Arlene Christina GD et al (2017) Nutritional and health benefits of millets, 1st edn. ICAR- Indian Institute of Millets Research, Hyderabad
Ravindran G (1992) Seed protein of millets: amino acid composition, proteinase inhibitors and in-vitro protein digestibility. Food Chem 44:13–17. https://doi.org/10.1016/0308-8146(92)90251-V
Rose DJ, Santra DK (2013) Proso millet (Panicum miliaceum L.) fermentation for fuel ethanol production. Ind Crop Prod 43:602–605. https://doi.org/10.1016/j.indcrop.2012.08.010
Roy SK, Kwon S-J, Yu J-H et al (2017) Comparison of protein profiles of proso millet (Panicum miliaceum) seeds of various Korean cultivars. Korean J Crop Sci 62:40–50. https://doi.org/10.7740/KJCS.2016.62.1.040
Saleh ASM, Zhang Q, Chen J, Shen Q (2013) Millet grains: nutritional quality, processing, and potential health benefits. Compr Rev Food Sci Food Saf 12:281–295. https://doi.org/10.1111/1541-4337.12012
Santra DK (2013) Proso millet varieties for Western Nebraska. University of Nebraska-Lincoln NebGuide, Scottsbluff, Nebraska
Santra DK, Rose DJ (2013) Alternative uses of proso millet. University of Nebraska-Lincoln Extension, p G2218
Santra DK, Heyduck RF, Baltensperger DD et al (2015) Registration of ‘plateau’ waxy (amylose-free) proso millet. J Plant Regist 9:41–43. https://doi.org/10.3198/jpr2013.11.0067crc
Santra DK, Khound R, Das S (2019) Proso millet (Panicum miliaceum L.) breeding: progress, challenges and opportunities. In: Al-Khayri JM, Jain SM, Johnson DV (eds) Advances in plant breeding strategies: cereals. Springer International Publishing, Cham, pp 223–257
Shevchenko A, Yang Y, Knaust A et al (2014) Proteomics identifies the composition and manufacturing recipe of the 2500-year old sourdough bread from Subeixi cemetery in China. J Proteome 105:363–371. https://doi.org/10.1016/j.jprot.2013.11.016
Shi J, Ma X, Zhang J et al (2019) Chromosome conformation capture resolved near complete genome assembly of broomcorn millet. Nat Commun 10:464. https://doi.org/10.1038/s41467-018-07876-6
Singh V, Moreau RA, Hicks KB (2003) Yield and phytosterol composition of oil extracted from grain sorghum and its wet-milled fractions. Cereal Chem 80:126–129. https://doi.org/10.1094/CCHEM.2003.80.2.126
Singh M, Adedeji A, Santra D (2018) Physico-chemical and functional properties of nine proso millet cultivars. Trans ASABE 61:1165–1174. https://doi.org/10.13031/trans.12553
Singode A, Balakrishna D, Bhat V et al (2018) Improving yield using EMS mutation in proso millet (Panicum miliaceum L). In: Santra DK, Johnson JJ (eds) The 3rd international broomcorn millet symposium. Fort Collins, Colorado, USA
Srivastava RK, Singh RB, Pujarula VL et al (2020) Genome-wide association studies and genomic selection in pearl millet: advances and prospects. Front Genet 10:1389. https://doi.org/10.3389/fgene.2019.01389
Tripathi MK, Mohapatra D, Jadam RS et al (2021) Nutritional composition of millets. In: Kumar A, Tripathi MK, Joshi D, Kumar V (eds) Millets and millet technology. Springer Singapore, Singapore, pp 101–119
Varshney RK, Shi C, Thudi M et al (2017) Pearl millet genome sequence provides a resource to improve agronomic traits in arid environments. Nat Biotechnol 35:969–976. https://doi.org/10.1038/nbt.3943
Vetriventhan M, Upadhyaya HD (2018) Diversity and trait-specific sources for productivity and nutritional traits in the global proso millet (Panicum miliaceum L.) germplasm collection. Crop J 6:451–463. https://doi.org/10.1016/j.cj.2018.04.002
Wang R, Wang H, Liu X et al (2018) Waxy allelic diversity in common millet (Panicum miliaceum L.) in China. Crop J 6:377–385.
Wietgrefe GW (1990) How to produce proso millet (a Farmer’s guide). Wietgrefe, Gary W., Pierre, South Dakota
Yabe S, Iwata H (2020) Genomics-assisted breeding in minor and pseudo-cereals. Breed Sci 70:19–31. https://doi.org/10.1270/jsbbs.19100
Yue H, Wang L, Liu H et al (2016) De novo assembly and characterization of the transcriptome of broomcorn millet (Panicum miliaceum L.) for gene discovery and marker development. Front Plant Sci 7:1083. https://doi.org/10.3389/fpls.2016.01083
Zhang Y, Gao X, Li J et al (2019) Comparative analysis of proso millet (Panicum miliaceum L.) leaf transcriptomes for insight into drought tolerance mechanisms. BMC Plant Biol 19:397. https://doi.org/10.1186/s12870-019-2001-x
Zhao B, Khound R, Ghimire D et al (2022) Heading percentage estimation in proso millet (Panicum miliaceum L.) using aerial imagery and deep learning. Plant Phenome J. https://doi.org/10.1002/ppj2.20049
Zou C, Li L, Miki D et al (2019) The genome of broomcorn millet. Nat Commun 10:436. https://doi.org/10.1038/s41467-019-08409-5
Conflict of Interest
The authors declare that they have no conflict of interest.
Funding Acknowledgment
The project was supported by “Research State Aided” internal funds: 21-6243-1001.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2023 Springer Nature Singapore Pte Ltd
About this entry

Cite this entry
Khound, R., Mathivanan, R.K., Santra, D.K. (2023). Proso Millet Nutraceutomics for Human Health and Nutritional Security. In: Kole, C. (eds) Compendium of Crop Genome Designing for Nutraceuticals. Springer, Singapore. https://doi.org/10.1007/978-981-19-4169-6_10
Download citation
DOI: https://doi.org/10.1007/978-981-19-4169-6_10
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-19-4168-9
Online ISBN: 978-981-19-4169-6
eBook Packages: Biomedical and Life SciencesReference Module Biomedical and Life Sciences