
Abstract
Lysosomes participate within the cells in the degradation of organelles, macromolecules, and a wide variety of substrates. In any study on specific roles of lysosomes, both under physiological and pathological conditions, it is advisable to include methods that allow their reproducible and reliable isolation. However, purification of lysosomes is a difficult task, particularly in the case of cultured cells. This is mainly because of the heterogeneity of these organelles, along with their low number and high fragility. Also, isolation methods, while disrupting plasma membranes, have to preserve the integrity of lysosomes, as the breakdown of their membranes releases enzymes that could damage all cell organelles, including themselves. The protocols described below have been routinely used in our laboratory for the specific isolation of lysosomes from rat liver, NIH/3T3, and other cultured cells, but can be adapted to other mammalian tissues or cell lines.
Access provided by CONRICYT – Journals CONACYT. Download protocol PDF
Similar content being viewed by others



Key words
1 Introduction
Lysosomes are a group of organelles with varying sizes, forms, content, and densities. Lysosomes enclose a wide variety of acid hydrolases, including proteases (cathepsins ), lipases , glucosidases , and nucleases [1, 2]. Macromolecules and other substrates reach the lysosomes by diverse mechanisms, including endocytosis , crinophagy, and different kinds of autophagy (macroautophagy , microautophagy , and chaperone-mediated autophagy ) [3]. Once inside the lysosomes, the sequestered materials are degraded and their building blocks are recycled. Lysosomes are involved in a large variety of cell processes, including differentiation, development, aging and cell death, and they also play an important role in many pathological disorders, such as cancer and neurological diseases.
Since the discovery of lysosomes 60 years ago [4] and their initial implication in the so-called lysosomal storage diseases [5, 6], the studies on these organelles, which usually include their isolation, have grown exponentially. For example, isolation of lysosomes followed by proteomic analysis has been used to identify new lysosomal membrane proteins (e.g., [7–9]). These and other studies where isolation of lysosomes was employed have been useful to progress our knowledge on the physiology and pathology of lysosomes .
Subcellular fractionation has allowed researchers to study the characteristics and function of different cellular components. Lysosomes were first identified by de Duve when trying to find the structure where glucose 6-phosphatase was localized [10]. This was done by differential centrifugation of liver homogenates to obtain a light mitochondrial (LM) fraction, followed by a sucrose gradient that separated lysosomes from other cell components in this fraction by their different density. Because of the above-mentioned heterogeneity of lysosomes, purification of a pure fraction of lysosomes is a difficult task and it is common to find these fractions contaminated, mainly with mitochondria and peroxisomes but also with other cell components. This caveat was first overcome by the injection of detergent Triton WR-1339 [11, 12]. The detergent is selectively taken up by lysosomes , reducing the lysosomal density and therefore allowing a better separation from mitochondria. Other strategy for achieving a better separation, for example, is the loading of lysosomes with colloidal gold [13, 14], producing an increase in the lysosomal density. These methods are currently in use (e.g., [7]) and allow a better purification of the lysosomal fraction . However, they imply a change in the lysosomal composition, as they are loaded with external agents and this could affect the lysosomal function. In fact, the organelles purified with both procedures are tritosomes and aurosomes , which are not strictly lysosomes.
The extraction of pure “intact” lysosomes is therefore a critical step for researchers in this field. The isolation method has been improved by the use of gradients of Percoll, Metrizamide, Nycodenz, and other reagents (e.g., [9, 15]). Although these methods have been widely used for the isolation of lysosomes from animal tissues , this is much more arduous when using cultured cells, particularly because the difficulty of disrupting their plasma membranes without affecting the lysosomal membranes. Anyway, there are several reports in the literature for the isolation of lysosomes or their subpopulations from different sources (e.g., [7, 14, 16–19]). Here, we describe the protocols that we have followed for many years in our laboratory to isolate these organelles.
2 Materials
2.1 Isolation of Lysosomes from Rat Liver
Prepare all solutions using ultrapure water and centrifugation or analytical grade reagents. Solutions must be freshly made or stored at –20 °C, unless otherwise stated. All experiments involving animals should be conducted in compliance with approved Institutional Animal Use Committee protocols.
-
1.
Male Wistar rats weighing 200–250 g starved during 16–24 h (only with water ad libitum).
-
2.
Dissection instruments (scissors, clamps, and tweezers).
-
3.
Glassware: 250 mL Erlenmeyer flask, 250 mL beakers, and 100 mL graduated cylinder.
-
4.
Gauze (two layers) or cheesecloth.
-
5.
Thomas Pestle Tissue Grinder homogenizer (55-mL, Thomas Scientific, catalog number 3431E55, Swedesboro, NJ, USA) attached to an IKA RW-20 digital dual-range mixer (Cole-Parmer, catalog number EW-50705-00, Vernon Hills, IL, USA).
-
6.
Refractometer (Carl Zeiss, Jena, Germany).
-
7.
Cold finger (10 × 150 mm test tube filled with ice).
-
8.
Pasteur pipettes (plastic and glass) and pipette rubber bulbs.
-
9.
Homogenization medium: 0.3 M sucrose. This solution can be prepared the previous day and stored at 4 °C.
-
10.
85.6 % Metrizamide in water at pH 7.0. Preparation of this solution is extremely tedious (see Note 1 ).
-
11.
Heraeus centrifuge (Biofuge 28RS) equipped with a 3745 rotor or equivalent (Hanau, Germany) and 50-mL polycarbonate tubes.
-
12.
Sorvall centrifuge (Evolution RL) equipped with a SA-300 rotor or equivalent (Thermo Fisher Scientific, Waltham, MS, USA) and 50-mL polycarbonate tubes.
-
13.
Ultracentrifuge (Optima XL-100K) equipped with a SW40 Ti rotor (5–6 g of liver , see Note 2 ) and 14 × 95 mm Ultra-Clear tubes (catalog number 344060) (Beckman Coulter, Brea, CA, USA).
2.2 Isolation of Lysosomes from Cultured Cells
-
1.
Polystyrene square cell culture dishes (Nunc, Thermo Fisher Scientific, catalog number 166508). Culture area: 500 cm2.
-
2.
Heraeus centrifuge (Biofuge 28RS) equipped with a 3745 rotor or equivalent and 50-mL polycarbonate tubes.
-
3.
Thomas Pestle Tissue Grinder, 10-mL (Thomas Scientific, catalog number 3431E45).
-
4.
Polyethylene cell lifters (Corning Incorporated, catalog number 3008, NY, USA).
-
5.
Parr nitrogen bomb (Parr, model 4639, IL, USA) (see Note 3 ).
-
6.
Ultracentrifuge: Optima XL-100K equipped with a SW40 Ti and a 70.1 Ti rotors Optima MAX-130 equipped with a TLA-100 and a TLA-55 rotors. Ultracentrifuge tubes: 14 × 95 mm Ultra-Clear (catalog number 344060), 16 × 76 mm thickwall polycarbonate (catalog number 355630), 11 × 39 mm microcentrifuge polyallomer (catalog number 357448), and 7 × 20 mm thickwall polyallomer (catalog number 343621) (Beckman Coulter).
-
7.
Krebs-Henseleit (KH) medium: 118.4 mM NaCl, 4.75 mM KCl, 1.19 mM KH2PO4, 2.54 mM MgSO4, 2.44 mM CaCl2·2H2O, 28.6 mM NaHCO3, 10 mM glucose, containing 10 mM HEPES, pH 7.4.
-
8.
Phosphate buffered saline (PBS) medium: 150 mM NaCl, 2.7 mM KCl, 1.4 mM KH2PO4, 10 mM Na2HPO4, pH 7.4.
-
9.
Percoll/Metrizamide solutions and 0.25 M sucrose (see Note 4 ).
-
10.
Homogenization buffer (HB) 10×: 2.5 M sucrose, 100 mM HEPES, 10 mM EDTA, pH 7.3.
-
11.
Pasteur pipettes (plastic and glass) and pipette rubber bulbs.
3 Methods
3.1 Isolation of Lysosomes from Rat Liver (Adapted from Wattiaux et al. [15])
Once the liver is obtained all the steps should be performed on ice or at 4 °C to prevent lysosomes from becoming damaged by released enzymes.
-
1.
Obtain liver from a starved Wistar rat. Remove skin and fat tissue to get the liver as clean as possible and weigh the liver.
-
2.
Wash extensively with cold 0.3 M sucrose in a precooled beaker and cut into small pieces with the aid of scissors to facilitate homogenization.
-
3.
Homogenize in 3 volumes of 0.3 M sucrose per gram of tissue by 8 strokes at 500 rpm in a homogenizer.
-
4.
Add 4 additional volumes of 0.3 M sucrose and filter the homogenate through double gauze without pressing it and collect in a precooled Erlenmeyer flask. Separate 60 μL of homogenate in an Eppendorf tube for determination of specific activities of enzymes.
-
5.
Split the homogenate into two 50-mL polycarbonate tubes (use 0.3 M sucrose to equilibrate) and centrifuge at 4800 × g for 5 min at 4 °C in a Heraeus 3745 rotor.
-
6.
After centrifugation, collect the supernatant into clean 50-mL polycarbonate tubes (see Note 5 ).
-
7.
Centrifuge at 17,000 × g for 10 min at 4 °C in a Heraeus 3745 rotor.
-
8.
Remove the supernatant (see Note 6 ). Resuspend the pellet from this second centrifugation with a “cold finger” and wash it with 0.3 M sucrose (3.5 volumes per gram of liver ).
-
9.
Centrifuge as in #7. Remove the supernatant.
-
10.
Resuspend the pellet with a “cold finger” in 1 mL of 0.3 M sucrose. This is an LM fraction.
-
11.
Place the LM fraction at the bottom of an Ultra-Clear ultracentrifuge tube and add 3.39 g of 85.6 % Metrizamide to (adjust final Metrizamide concentration to 57 %). Mix gently by vortexing (see Note 7 ).
-
12.
Generate a discontinuous Metrizamide gradient by carefully and consecutively overlaying the following Metrizamide solutions on top of the LM fraction mixed with Metrizamide (Fig. 1):
Fig. 1 
Isolation of lysosomes from rat liver . Once the light mitochondrial (LM) fraction is obtained, it is loaded on the bottom of an Ultra-Clear ultracentrifuge tube and adjusted to 57 % Metrizamide (Mtz). Three layers are overlaid on top to create a discontinuous Mtz gradient and a 0.3 M sucrose layer is finally added. After 90 min centrifugation at the indicated conditions, lysosomes are located at the 0.3 M sucrose/19.8 % Mtz and 19.8 %/26.3 % Mtz interfaces (F1 and F2). F3 and F4 are enriched in mitochondria. Whitish circles represent lysosomes and brown circles mitochondria
-
2.13 mL of 32.8 % Metrizamide.
-
3.50 mL of 26.3 % Metrizamide.
-
3.85 mL of 19.8 % Metrizamide.
Fill the tube up to about 2–3 mm from the border with 0.3 M sucrose, to prevent collapse during centrifugation.
-
-
13.
Centrifuge the gradient at 141,000 × g for 90 min at 4 °C in a SW40 Ti rotor (see Note 8 ).
-
14.
After ultracentrifugation , four bands (F1 to F4, from top to bottom) are distinguished in the gradient interfaces (see Fig. 1). With a glass Pasteur pipette collect the two upper whitish bands (F1 + F2), which contain lysosomes (approximately 2–3 mL).
-
15.
Mix with 10 volumes of ice cold 0.3 M sucrose in 50-mL polycarbonate tubes and centrifuge at 37,000 × g for 10 min at 4 °C (to remove Metrizamide) in a Sorvall centrifuge using a SA-300 rotor. This pellet contains the lysosomes . Resuspend them with a blunt Pasteur pipette (see Note 9 ) in 30 mL of ice cold 0.3 M sucrose and wash two times (Fig. 2a).
Fig. 2 
Purified lysosomes. Representative electron micrographs of lysosomes isolated from rat liver (a) and NIH/3T3 cells (b). Bar: 1 μm
-
16.
Resuspend the last sediment as above in 250–300 μL of the solution suitable for your following experiments (see Note 10 ).
-
17.
Separate 40 μL for quantification of protein and enzymatic activities (see Note 11 ).


3.2 Isolation of Lysosomes from Cultured Cells (Adapted from Storrie and Madden [16])
For optimal isolation use 4–5 square cell culture dishes per condition (see Note 12 ). After washing the cells with 70 mL PBS, treat them for 30 min to 4 h at 37 °C with 70 mL KH medium/dish (see Note 13 ).
-
1.
Remove the KH medium from the plates and carefully detach the cells with a cell lifter in cold PBS (2 mL/ plate). Collect the cells into 50-mL tubes with a plastic Pasteur pipette.
-
2.
Centrifuge cells at 800 × g for 5 min at 4 °C in a Heraeus 3745 rotor.
-
3.
Wash with 0.25 M sucrose (25–30 mL). Centrifuge as in #2.
-
4.
Resuspend the pellet in 3 mL of 0.25 M sucrose. Place the cell suspension in the precooled chamber of the nitrogen bomb. Slowly, open the valve and allow the pressure to reach 35 psi (2.41 Bar) in 1 min. Keep pressure at 35 psi for a total of 7 min with occasional shaking (optimize conditions according to cell type, see Note 3 ).
-
5.
Further homogenize the cells on ice using a precooled 10-mL homogenizer and 8 strokes of its Teflon pestle. Collect the homogenate and wash the homogenizer with 1 mL of 0.25 M sucrose, to get a final volume of about 4 mL. Separate 60 μL of homogenate in an Eppendorf tube for determination of specific activities of enzymes.
-
6.
Centrifuge at 2500 × g for 15 min at 4 °C in a Heraeus centrifuge using a 3745 rotor (see Note 14 ).
-
7.
In the meantime, prepare the gradient in an Ultra-Clear tube (14 × 95 mm) by adding the following solutions from bottom to top (Fig. 3a):
Fig. 3 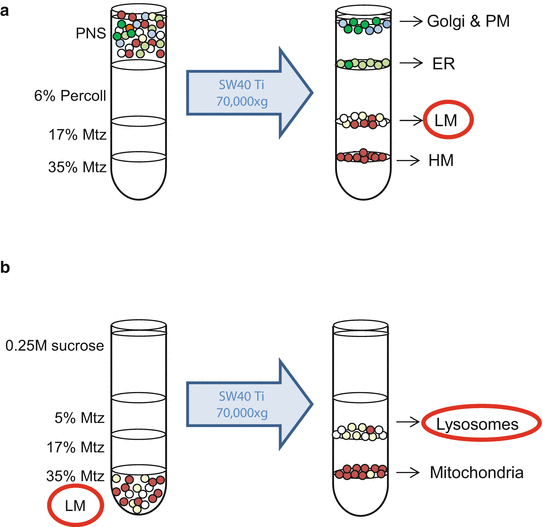
Isolation of lysosomes from cultured cells. (a) Once the postnuclear supernatant (PNS) is obtained, it is layered on top of a Percoll/Metrizamide (Mtz) gradient. After a 35 min centrifugation at the indicated conditions, the light mitochondrial (LM) fraction is recovered from the 17 % Mtz/6 % Percoll interface. (b) This fraction is loaded on the bottom of an Ultra-Clear ultracentrifuge tube and adjusted to 35 % Mtz. Three layers are overlaid on top to create a discontinuous Mtz gradient. After a 35 min centrifugation at the indicated conditions, lysosomes are located at the 17 %/5 % Mtz interface. Green and blue circles represent Golgi complex and plasma membrane (PM); light green circles represent endoplasmic reticulum (ER); whitish circles represent lysosomes and brown circles represent mitochondria (light and heavy (HM))
-
35 % Metrizamide in 0.25 M sucrose: 2.6 mL.
-
17 % Metrizamide in 0.25 M sucrose: 2.6 mL.
-
6 % Percoll in 0.25 M sucrose: 3.9 mL.
-
-
8.
Add the supernatant from #6 (PNS, approximately 3.5 mL) on top of the discontinuous gradient (Fig. 3a).
-
9.
Centrifuge at 70,000 × g for 35 min at 4 °C in a SW40 Ti rotor. LM fraction appears in the 17 % Metrizamide/6 % Percoll interface (see Fig. 3a).
-
10.
Collect the LM band, approximately 1.4 mL (see Note 15 ). Put at the bottom of an Ultra-Clear tube (14 × 95 mm) with 1.1 mL of 80 % Metrizamide and mix gently by vortexing. Add the following solutions from bottom to top (Fig. 3b):
-
17 % Metrizamide in 0.25 M sucrose: 2.6 mL.
-
5 % Metrizamide in 0.25 M sucrose: 2.6 mL.
-
0.25 M sucrose up to 3 mm from the border, approximately 5 mL.
-
-
11.
Centrifuge at 70,000 × g for 35 min at 4 °C in a SW40 Ti rotor.
-
12.
Collect the lysosomes , approximately 1.2 mL (second band from bottom, Fig. 3b). Dilute with 3–4 volumes of 0.25 M sucrose and centrifuge at 100,000 × g for 30 min at 4 °C in a 70.1 Ti rotor using thickwall polycarbonate tubes. Repeat this step.
-
13.
Using a blunt Pasteur pipette, resuspend the lysosomal pellet in 150–200 μL of the solution suitable for your following experiments (see Note 10 ).
-
14.
Separate 20 μL at −80 °C for protein and enzymatic activities measurements (see Note 16 ).
An alternative method to avoid the use of Metrizamide requires modification in the following specific steps of the previous procedure:
-
1.
Wash the cells with 1× HB (about 33 mL/tube) instead of 0.25 M sucrose.
-
2.
Resuspend the pellet in 3 mL of 1× HB and homogenize as in the method above (see Note 17 ).
-
3.
Prepare the gradient by adding the following solutions from bottom to top:
-
10× HB: 1.2 mL.
-
17.5 % Percoll in 1× HB: 8.5 mL.
-
-
4.
Centrifuge at 67,000 × g for 30 min at 4 °C in a 70.1 Ti rotor.
-
5.
Remove the first 1 mL and take 9 different fractions of 1.2 mL in separate Eppendorf tubes. Lysosomes are mainly located in the first three fractions, but it is advisable when using other cell types to recover and analyze the other fractions too. Pool fractions 1–3 in a thickwall polycarbonate tube.
-
6.
Centrifuge at 100,000 × g for 30 min at 4 °C in a 70.1 Ti rotor (see Note 18 ). Continue with #12 of the main protocol.

4 Notes
-
1.
Preparation of Metrizamide solutions. The stock solution (85.6 % Metrizamide) should be dissolved in the dark. To avoid the formation of clumps of undissolved material, start with half of the final volume of water and add small amounts of Metrizamide while stirring. Do not add more Metrizamide until the former has been fully dissolved. Adjust the pH of the solution to 7.0 with 0.01 M NaOH while stirring (since this solution has an elevated density, wait a while for an accurate pH measurement). Using a refractometer, adjust more precisely the concentration of the different solutions by adding water at pH 7.0 or stock solution to bring them to the corresponding refractive index (see below). Store all solutions at –20 °C. The volumes needed for two samples are shown below:
85.6 % Metrizamide (mL)
Water pH 7.0 (mL)
Refractive index (20 °C)
85.6 %
5.00
–
1.4710
32.8 %
1.92 (2.80 g)
3.08
1.3854
26.3 %
2.46 (3.58 g)
5.54
1.3763
19.8 %
1.85 (2.71 g)
6.15
1.3643
Nycodenz, more easily available and less expensive than Metrizamide, could probably replace the use of Metrizamide in a similar protocol [9], but further analyses are recommended.
-
2.
For larger amounts of tissue (about 20 g) use the SW28 rotor, adjusting the volumes of the different Metrizamide layers in the gradient.
-
3.
Parr nitrogen bomb is a cell disruptor by nitrogen decompression. It preserves organelle integrity in cultured cells better than other methods. Conditions provided in this method are optimized for NIH/3T3 cells. Time and nitrogen pressure conditions should be optimized for other cell lines.
-
4.
Percoll/Metrizamide solutions.
-
(a)
80 % Metrizamide in 0.25 M sucrose pH 7.2. All Metrizamide solutions are prepared in 0.25 M sucrose pH 7.2.
Preparation: put 50 mL of 0.25 M sucrose in a glass beaker and slowly add 80 g of Metrizamide. Adjust the pH with 0.01 N NaOH and add 0.25 M sucrose to a final volume of 100 mL. Remember (see Note 1 ) to carry out these preparations in the dark. The volumes needed for two samples are shown below:
80 % Metrizamide (mL)
Percoll (mL)
0.25 M sucrose (mL)
35 % Metrizamide
2.40
–
3.10
17 % Metrizamide
2.34
–
8.66
15 % Metrizamide
0.34
–
5.16
6 % Percoll
–
0.51
8.00
-
(b)
17.5 % Percoll in 1× HB: 18 mL are needed for two samples. Mix 3.15 mL of 100 % Percoll, 1.8 mL of 10× HB and 13.05 mL of water.
-
(a)
-
5.
Take care to avoid collecting portions of the white sediment from this first centrifugation, which consists of erythrocytes, nuclei, large mitochondria, and some cell debris.
-
6.
The supernatants from centrifugation at 17,000 × g for 10 min contain cytosol, microsomes , etc.
-
7.
To collect the LM fraction, Pasteur pipettes or other wide mouth pipettes are better than micropipette tips to reduce damage to lysosomal membranes.
-
8.
It is advisable to carry out all centrifugations of gradients without brake and with low acceleration.
-
9.
To prepare the blunt Pasteur pipette, heat the edge of a glass pipette to make a glass drop at its end.
-
10.
Lysosomes can be used for different experiments and they should be treated accordingly. For example, for chaperone-mediated autophagy studies, freshly isolated lysosomes should be immediately resuspended, after washing, in 10 mM MOPS/0.3 M sucrose pH 7.0 and incubated with the appropriate substrates and reagents as described [18, 20]. For proteomic studies of lysosomal components (e.g., [8]), it is advisable to incubate the lysosomes for 30 min at 37 °C in an iso-osmotic medium (250–300 mOsmol/L) to allow degradation of the cytoplasmic material sequestered by the lysosomes. For isolation of lysosomal membranes follow this procedure:
-
Resuspend lysosomes in 200 μL of water plus protease inhibitors (1 mM phenylmethylsulfonyl fluoride, 100 μM leupeptin, 10 μM pepstatin A, 2 mM EDTA, 2 mM dithiothreitol).
-
Disrupt lysosomes by 6–8 freeze-thaw cycles and homogenize.
-
Centrifuge at 130,000 × g for 10 min at 4 °C in an Optima MAX-130 equipped with a TLA-100 rotor or equivalent.
-
Wash the membranes three times with 100 μL of water plus protease inhibitors to remove residual components from the lysosomal matrix. Centrifuge as above.
-
Resuspend in 100 μL of water plus protease inhibitors (see above) and analyze.
-
-
11.
To calculate the yield and the degree of purification of this lysosomal fraction relative to the original homogenate, we routinely use β-N-acetyl-glucosaminidase and β-hexosaminidase as lysosomal marker enzymes. In our hands, yield and degree of purification are 6–10 % and (70–80)-fold. For experiments with intact lysosomes, calculate lysosomes latency by measuring the activity of a lysosomal enzyme in the presence or not of 0.15 % Triton X-100. Preparations with latency values below 95 % are discarded for those experiments. Protein concentration can be evaluated using a Micro BCA protein assay kit (Thermo Scientific) or an equivalent procedure.
-
12.
We use 90 % confluent NIH/3T3 cells, about 8 × 107cells/plate.
-
13.
KH medium or equivalent starvation media are used to increase the cellular lysosomal mass.
-
14.
The supernatant and the pellet from this centrifugation are the postnuclear supernatant (PNS) and pellet (PNP), respectively. Resuspend the PNP in 4 mL of 0.25 M sucrose and reserve an aliquot for enzymatic activity measurements.
-
15.
To avoid the contamination of the lysosomal fraction with other cellular components, remove the two upper bands with a Pasteur pipette.
-
16.
Calculate yield and degree of purification of the lysosomal fraction as in Note 11 . In our hands, yield and degree of purification are 3–5 % and 20- to 30-fold, respectively.
-
17.
The addition of 10 mM potassium phosphate pH 7.4 alters the sedimentation rate of mitochondria without affecting the lysosomes, improving the purity of the lysosomal fraction [21].
-
18.
If the sediments of this centrifugation are loose, centrifuge at 70,000 × g for 30 min at 4 °C using an Optima MAX-130 equipped with a TLA-55 rotor or equivalent.
References
Saftig P (ed) (2005) Lysosomes. Landes Bioscience, Georgetown, TX, USA
Wartosch L, Bright NA, Luzio JP (2015) Lysosomes. Curr Biol 25(8):R315–R316
Knecht E, Aguado C, Cárcel J, Esteban I, Esteve JM, Ghislat G, Moruno JF, Vidal JM, Saez R (2009) Intracellular protein degradation in mammalian cells: recent developments. Cell Mol Life Sci 66(15):2427–2443
De Duve C, Pressman BC, Gianetto R, Wattiaux R, Appelmans F (1955) Tissue fractionation studies. 6. Intracellular distribution patterns of enzymes in rat-liver tissue. Biochem J 60(4):604–617
Hers HG, van Hoof F (eds) (1973) Lysosomes and storage diseases. Academic, New York, NY, USA
Parenti G, Andria G, Ballabio A (2015) Lysosomal storage diseases: from pathophysiology to therapy. Annu Rev Med 66:471–486
Zhang H, Fan X, Bagshaw R, Mahuran DJ, Callahan JW (2008) Purification and proteomic analysis of lysosomal integral membrane proteins. Methods Mol Biol 432:229–241
Ghislat G, Aguado C, Knecht E (2012) Annexin A5 stimulates autophagy and inhibits endocytosis. J Cell Sci 125(Pt 1):92–107
Chapel A, Kieffer-Jaquinod S, Sagne C, Verdon Q, Ivaldi C, Mellal M, Thirion J, Jadot M, Bruley C, Garin J, Gasnier B, Journet A (2013) An extended proteome map of the lysosomal membrane reveals novel potential transporters. Mol Cell Proteomics 12(6):1572–1588
de Duve C (2005) The lysosome turns fifty. Nat Cell Biol 7(9):847–849
Leighton F, Poole B, Beaufay H, Baudhuin P, Coffey JW, Fowler S, De Duve C (1968) The large-scale separation of peroxisomes, mitochondria, and lysosomes from the livers of rats injected with triton WR-1339. Improved isolation procedures, automated analysis, biochemical and morphological properties of fractions. J Cell Biol 37(2):482–513
Horvat A, Baxandall J, Touster O (1969) The isolation of lysosomes from Ehrlich ascites tumor cells following pretreatment of mice with Triton WR-1339. J Cell Biol 42(2):469–479
Henning R, Plattner H (1974) Isolation of rat liver lysosomes by loading with colloidal gold. Biochim Biophys Acta 354(1):114–120
Berg TO, Fengsrud M, Stromhaug PE, Berg T, Seglen PO (1998) Isolation and characterization of rat liver amphisomes. Evidence for fusion of autophagosomes with both early and late endosomes. J Biol Chem 273(34):21883–21892
Wattiaux R, Wattiaux-De Coninck S, Ronveaux-dupal MF, Dubois F (1978) Isolation of rat liver lysosomes by isopycnic centrifugation in a metrizamide gradient. J Cell Biol 78(2):349–368
Storrie B, Madden EA (1990) Isolation of subcellular organelles. Methods Enzymol 182:203–225
Graham JM (2001) Purification of a crude mitochondrial fraction by density-gradient centrifugation. Curr Protoc Cell Biol Chapter 3:Unit 3 4
Kaushik S, Cuervo AM (2008) Chaperone-mediated autophagy. Methods Mol Biol 445:227–244
Seglen PO, Brinchmann MF (2010) Purification of autophagosomes from rat hepatocytes. Autophagy 6(4):542–547
Patel B, Cuervo AM (2015) Methods to study chaperone-mediated autophagy. Methods 75:133–140
Lardeux B, Gouhot B, Forestier M (1983) Improved recovery of rat liver fractions enriched in lysosomes by specific alteration of the sedimentation properties of mitochondria. Anal Biochem 131(1):160–165
Acknowledgements
Current work in the laboratory is supported by grants from the Spanish Ministry of Economy and Competitivity (BFU2011-22630 and SAF2014-54604-C3-2-R), Generalitat Valenciana (Prometeo 2009/051), and Instituto de Salud Carlos III (ACCI2015 action from CIBER on Rare Diseases). The authors would like to acknowledge networking support by the Proteostasis COST Action (BM1307).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer Science+Business Media New York
About this protocol
Cite this protocol
Aguado, C., Pérez-Jiménez, E., Lahuerta, M., Knecht, E. (2016). Isolation of Lysosomes from Mammalian Tissues and Cultured Cells. In: Matthiesen, R. (eds) Proteostasis. Methods in Molecular Biology, vol 1449. Humana Press, New York, NY. https://doi.org/10.1007/978-1-4939-3756-1_19
Download citation
DOI: https://doi.org/10.1007/978-1-4939-3756-1_19
Published:
Publisher Name: Humana Press, New York, NY
Print ISBN: 978-1-4939-3754-7
Online ISBN: 978-1-4939-3756-1
eBook Packages: Springer Protocols