
Abstract
Salmonella is a gram-negative facultative intracellular pathogen that is capable of infecting a variety of hosts. Inside host cells, most Salmonella bacteria reside and replicate within Salmonella-containing vacuoles. They use virulence proteins to manipulate the host cell machinery for their own benefit and hijack the host cytoskeleton to travel toward the perinuclear area. However, a fraction of bacteria escapes into the cytosol where they get decorated with a dense layer of polyubiquitin, which labels the bacteria for clearance by autophagy. More specifically, autophagy receptor proteins recognize the ubiquitinated bacteria and deliver them to autophagosomes, which subsequently fuse to lysosomes. Here, we describe methods used to infect HeLa cells with Salmonella bacteria and to detect their ubiquitination via immunofluorescence and laser scanning confocal microscopy.
Access provided by CONRICYT – Journals CONACYT. Download protocol PDF
Similar content being viewed by others


Key words
- Salmonella
- Bacteria
- Ubiquitination
- Autophagy
- Confocal microscopy
- Imaging
- Immunofluorescence
- Host–pathogen interaction
- Bacterial clearance
1 Introduction
Salmonella has traditionally been characterized as a vacuolar pathogen since most Salmonella bacteria reside in the so-called Salmonella-containing vacuole (SCV) , a membrane bound compartment derived from the endocytic pathway [1, 2]. Recent work indicates that depending on the cell type, Salmonella can occupy different niches within the cell. In epithelial cells, next to the vacuolar bacteria, a considerable proportion of the total bacteria population has been found to inhabit the cytosol, where they hyper-replicate [3]. However, once the bacteria reach the cytosol they are exposed to the host ubiquitin system [4]. The bacteria are decorated with a dense layer of polyubiquitin chains, which peaks at 4 h post infection [5]. This ubiquitin coat is recognized by autophagy receptors , NDP52, OPTN, and p62 , which can bind to ubiquitin and members of the autophagosomal protein microtubule-associated protein 1A/1B-light chain 3 (LC3) family, leading to targeting of bacteria into autophagosomes and their subsequent degradation in autophagolysosomes [6–8]. Ubiquitination of bacteria is also necessary for the direct recruitment of other important components of the autophagy machinery, including the Atg16L1 complex, the ULK1 complex, and ATG9L1 [9]. Knockdown of autophagy receptors leads to enhanced bacterial replication, indicating that autophagy restricts the growth of cytosolic bacteria and is necessary for their clearance.
It is still unclear whether the formation of a ubiquitin coat on the surface of intracellular bacteria involves the ubiquitination of bacterial surface proteins or the ubiquitination of host cell proteins that bind the bacteria upon membrane rupture. Either way, it is likely that diverse E3 ubiquitin ligases are involved in the recognition and targeting of cytosolic bacteria. Huett et al. reported that the E3 ubiquitin ligase LRSAM1 contributes to the ubiquitination of invading bacteria [10]. However, there is still a substantial amount of bacteria ubiquitinated in LRSAM1-deficient cells, indicating that other E3 ligases are also involved. Moreover, it has been reported that the ubiquitin coat surrounding Salmonella contains at least M1- and K63-linked polyubiquitin chains [11], while in vitro ubiquitination of Salmonella with LRSAM1 primarily produces K6- and K27-linked polyubiquitin [10]. Clearly, determination of the molecular machinery involved in the ubiquitination of invading bacteria and their subsequent clearance by autophagy still needs significant research. In this context, monitoring and quantifying ubiquitinated intracellular bacteria by confocal microscopy is very valuable (Fig. 1). Here we describe methods for the infection of HeLa cells with Salmonella bacteria, immunofluorescence staining with anti-ubiquitin, cellular imaging via confocal microscopy, and image processing.

Confocal microscopy of ubiquitination of intracellular Salmonella bacteria. Infected HeLa cells were fixed at indicated time points after infection (hours p.i.). Left panels show cellular and bacterial DNA stained with DAPI, middle panels show anti-ubiquitin visualized with Dylight488, and right panels show the overlays of both channels. All images are maximum intensity projections of confocal Z-slices acquired with a step size of 0.5 μm. Scale bar, 5 μm
2 Materials
2.1 Human Cell Culture and Bacterial Infection
-
1.
Culture medium for HeLa cells (human cervical carcinoma cell line): Dulbecco’s Modified Eagle Medium (DMEM ) (Gibco/BRL, Bethesda, MD), supplemented with 10 % fetal calf serum, 0.4 mM sodium pyruvate, 2 mM l-glutamine, 0.1 mM nonessential amino acids.
-
2.
Trypsin/EDTA buffer: 400 ml 0.04 % EDTA (1 mM EDTA, 0.1 mM NaCl, 2.5 mM KCl, 7 mM Na2HPO4⋅12H2O, and 2 mM KH2PO4) and 100 ml trypsin solution (0.1 mM Trypsin, 0.1 M NaCl, 2.5 mM KCl, 66 mM Tris–HCl, 0.7 mM Na2PO4, and 1 % phenol red), pH 7.6. Store at 4 °C.
-
3.
10× Phosphate-buffered saline (PBS): 95.44 g Dulbecco’s Phosphate Buffered Saline (Lonza, Verviers, Belgium) in 1 l water. Store at 4 °C.
-
4.
8-Well μ-Slides (ibidi, Munich, Germany).
-
5.
Luria Broth (LB) medium: 1 % bactotryptone, 0.5 % bactoyeast, 1 % NaCl; autoclave; store at room temperature or 4 °C.
-
6.
LB agar plates: Prepare LB medium as above, add 15 g agar/L before autoclaving. After autoclaving, cool to approximately 55 °C, and pour into petri dishes; let harden, then invert and store at 4 °C.
-
7.
Pathogenic Salmonella enterica ssp. enterica strain LMG3264 was purchased from the Belgian Coordinated Collections of Microorganisms (BCCM)-LMG Bacteria Collection (Ghent University, Belgium).
-
8.
Gentamycin (Gibco/BRL, Bethesda, MD).
2.2 Immuno-fluorescence Staining
-
1.
Fixation buffer: 4 % paraformaldehyde (PFA) in PBS (see Note 1 ). For 1 l of 4 % PFA, heat up 800 ml PBS in a glass beaker to approximately 60 °C while stirring. Do not let the solution boil. Add 40 g of PFA powder to the heated PBS solution. Slowly raise the pH by adding 1 N NaOH drop wise until the solution clears. Once the PFA is dissolved, let the solution cool down and filter through a 0.45 μm filter. Adjust the volume to 1 l with 1× PBS. Recheck the pH and adjust with small amounts of diluted HCl to approximately 6.9. The solution can be aliquoted and frozen or stored at 2–8 °C for up to 1 month.
-
2.
TBS-TX (25 mM Tris–HCl pH 8, 150 mM NaCl, 0.1 % Triton X-100, 1 % bovine serum albumin (BSA)). Triton X-100 and BSA are freshly added prior to use.
-
3.
Primary antibody: mouse monoclonal antibody against mono- and polyubiquitinated conjugates clone FK2 (Enzo Life Sciences, Farmingdale, NY).
-
4.
Secondary antibody: DyLight 488 secondary antibody conjugates (Thermo Fisher Scientific, Waltham, MA).
-
5.
4′,6-Diamidino-2-phenylindole (DAPI) FluoroPure™ grade (Thermo Fisher Scientific, Waltham, MA). A 500 μM stock of DAPI is prepared in deionized water and aliquots are stored at −20 °C.
-
6.
Mounting medium: 1 % N-propyl-gallate in glycerol.
3 Methods
3.1 Seeding of HeLa Cells
-
1.
Plate 104 HeLa cells per well in 200 μl DMEM in 8-well μ-Slides (see Notes 2 and 3 ).
-
2.
Grow cells overnight in a humidified atmosphere at 37 °C and 5 % CO2.
3.2 Infection with Salmonella
-
1.
Two days prior to infection, streak bacteria from frozen glycerol stocks onto a fresh LB agar plate and incubate overnight at 37 °C (see Notes 4 – 6 ).
-
2.
Inoculate a single bacterial colony into 5 ml LB medium and incubate in a shaking incubator overnight at 37 °C (see Note 7 ).
-
3.
Subculture the bacterial suspension 1:33 in fresh LB medium and incubate in a shaking incubator at 37 °C for 2.5–3.5 h until they reach an OD600 of 0.7–1.
-
4.
Prior to infection refresh HeLa cells with new DMEM .
-
5.
Infect cells with 20 μl Salmonella subculture and incubate for 15 min at 37 °C/5 % CO2.
-
6.
After infection remove extracellular bacteria by washing twice with warm PBS and add DMEM containing 100 μg/ml gentamycin (see Note 8 ).
-
7.
After 1 h, change the medium to DMEM containing 20 μg/ml gentamycin.
-
8.
Collect samples at appropriate time points post infection.
3.3 Immuno-fluorescence Staining
-
1.
At appropriate time points wash cells twice with PBS (see Notes 8 and 9 ).
-
2.
Fix cells with fixation buffer for 20 min at room temperature (RT) to insure cellular preservation while imaging (see Notes 10 and 11 ).
-
3.
Wash fixed cells twice with PBS (see Note 12 ).
-
4.
Permeabilize the cells by incubating the slides for 10 min in TBS-TX.
-
5.
Dilute the primary antibody 1:200 in TBS-TX and incubate the slides with primary antibody overnight at 4 °C (see Note 13 ).
-
6.
Wash the slides three times for 5 min and once for 30 min with TBS-TX (see Note 14 ).
-
7.
Dilute the secondary antibody 1:500 in TBS-TX and incubate the slides with secondary antibody for 2 h at RT protected from light.
-
8.
Wash the slides three times for 5 min and once for 30 min with PBS.
-
9.
Incubate the slides with 500 nM DAPI in PBS for 20 min at RT protected from light in order to visualize the DNA present in the HeLa cells as well as the Salmonella bacteria.
-
10.
Rinse three times with PBS.
-
11.
Remove PBS and mount the slides with 200 μl mounting medium per well to prevent photo bleaching (see Notes 15 and 16 ).
3.4 Laser Scanning Confocal Microscopy and Image Processing
-
1.
Images are acquired using a Leica TCS SP5 AOBS confocal system (Leica, Manheim, Germany), with a 63× HCX PLApo 1.4 oil-immersion objective, with a format of 1024 × 1024, line average of 4, and zoom of 3 (pixel size: 70.6 nm by 70.6 nm). Bright-field images are acquired with the HeNe 543 nm laser line, while ubiquitin labeled with DyLight488 is imaged with Ar laser excitation at 488 nm, and nucleic acids with diode laser excitation at 405 nm. Z-stacks acquired for visualization purposes are imaged with axial resolution of 0.5 μm, while the stacks acquired for image analysis are imaged at nyquist Z resolution (step size 0.13 μm).
-
2.
Image analysis is performed with Volocity 3D Image Analysis Software (Perkin Elmer Life Sciences). A protocol can be developed where the bacteria are segmented as objects with fluorescence intensity above background and of predicted size. In the same protocol, a second measurement can be performed to segment for ubiquitin. Finally, an “intersect” algorithm is applied to identify ubiquitin-coated bacteria (Fig. 2). Additionally, the number of nuclei can be determined to give an overall count of the cells in the field of view. This can be useful to determine the average number of bacteria per cell and the number of ubiquitin-coated bacteria per cell.
Fig. 2 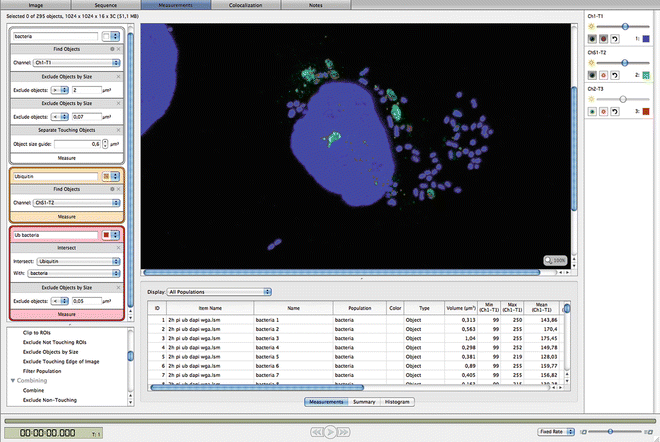
Image analysis to identify ubiquitin-coated Salmonella bacteria. A protocol was developed in Volocity software to segment bacteria and ubiquitin in order to identify ubiquitin-coated bacteria . On the left side the protocol is shown, on the right side the result of the segmentation is visible in the image. The boundaries of the segmented areas are shown in a different color for each of the three populations

4 Notes
-
1.
PFA is toxic. Gloves should be worn and solutions should be made in the fume hood.
-
2.
The cells need to be sub-confluent and nicely stretched out at the time of infection to enable an adequate analysis. As different cells in different laboratories tend to grow at different rates, it is advisable to determine the optimal cell concentration of the used cell line.
-
3.
Up to five 8-well μ-Slides can be stored in one 14 cm petri dish for easy handling and transportation.
-
4.
Salmonella enterica is classified as a biosafety level 2 (BL2) organism. All handling of these bacteria must be performed in a BL2 facility.
-
5.
Submerge an inoculation needle into the frozen glycerol stock and apply several streaks on an LB plate. Use a fresh inoculation needle and streak through the applied lines. Repeat several times in order to obtain single bacteria colonies.
-
6.
The LB plate containing bacterial colonies can be stored at 4 °C for several weeks. For new experiment, a single colony should be streaked on a fresh LB plate.
-
7.
We use 50 ml Falcon tubes to grow overnight cultures.
-
8.
For all washing steps remove the liquid carefully with a pipette from the corner of the well. Avoid scratching the monolayer with the tip of the pipette. When adding new liquid, pipette carefully against the wall of the chamber to avoid detachment of the cells.
-
9.
Do not let your cells dry at any point during the staining.
-
10.
Once the slides have been fixed in fixation buffer, all following steps may be performed outside the BL2 facility.
-
11.
Do not exceed the fixing time as over-fixation can lead to artifacts.
-
12.
Fixed cells can be stored in PBS at 4 °C for approximately 2 weeks without reducing the staining efficiency.
-
13.
Always include a negative staining control without primary antibody using only the secondary antibody to exclude nonspecific binding of the secondary antibody.
-
14.
Extensive washing is required to reduce background.
-
15.
Cut the tip off for pipetting the mounting medium.
-
16.
Once stained and covered with mounting medium, the fluorescent staining will stay for up to 1 month if you have a strong signal and if stored at 4 °C in the dark.
References
Drecktrah D, Knodler LA, Howe D, Steele-Mortimer O (2007) Salmonella trafficking is defined by continuous dynamic interactions with the endolysosomal system. Traffic 8(3):212–225. doi:10.1111/j.1600-0854.2006.00529.x
Steele-Mortimer O, Meresse S, Gorvel JP, Toh BH, Finlay BB (1999) Biogenesis of Salmonella typhimurium-containing vacuoles in epithelial cells involves interactions with the early endocytic pathway. Cell Microbiol 1(1):33–49
Malik-Kale P, Winfree S, Steele-Mortimer O (2012) The bimodal lifestyle of intracellular Salmonella in epithelial cells: replication in the cytosol obscures defects in vacuolar replication. PloS One 7(6):e38732. doi:10.1371/journal.pone.0038732
Perrin AJ, Jiang X, Birmingham CL, So NS, Brumell JH (2004) Recognition of bacteria in the cytosol of Mammalian cells by the ubiquitin system. Curr Biol 14(9):806–811. doi:10.1016/j.cub.2004.04.033
Birmingham CL, Smith AC, Bakowski MA, Yoshimori T, Brumell JH (2006) Autophagy controls Salmonella infection in response to damage to the Salmonella-containing vacuole. J Biol Chem 281(16):11374–11383. doi:10.1074/jbc.M509157200
Thurston TL, Ryzhakov G, Bloor S, von Muhlinen N, Randow F (2009) The TBK1 adaptor and autophagy receptor NDP52 restricts the proliferation of ubiquitin-coated bacteria. Nat Immunol 10(11):1215–1221. doi:10.1038/ni.1800
Wild P, Farhan H, McEwan DG, Wagner S, Rogov VV, Brady NR, Richter B, Korac J, Waidmann O, Choudhary C, Dotsch V, Bumann D, Dikic I (2011) Phosphorylation of the autophagy receptor optineurin restricts Salmonella growth. Science 333(6039):228–233. doi:10.1126/science.1205405
Zheng YT, Shahnazari S, Brech A, Lamark T, Johansen T, Brumell JH (2009) The adaptor protein p62/SQSTM1 targets invading bacteria to the autophagy pathway. J Immunol 183(9):5909–5916. doi:10.4049/jimmunol.0900441
Fujita N, Morita E, Itoh T, Tanaka A, Nakaoka M, Osada Y, Umemoto T, Saitoh T, Nakatogawa H, Kobayashi S, Haraguchi T, Guan JL, Iwai K, Tokunaga F, Saito K, Ishibashi K, Akira S, Fukuda M, Noda T, Yoshimori T (2013) Recruitment of the autophagic machinery to endosomes during infection is mediated by ubiquitin. J Cell Biol 203(1):115–128. doi:10.1083/jcb.201304188
Huett A, Heath RJ, Begun J, Sassi SO, Baxt LA, Vyas JM, Goldberg MB, Xavier RJ (2012) The LRR and RING domain protein LRSAM1 is an E3 ligase crucial for ubiquitin-dependent autophagy of intracellular Salmonella Typhimurium. Cell Host Microbe 12(6):778–790. doi:10.1016/j.chom.2012.10.019
van Wijk SJ, Fiskin E, Putyrski M, Pampaloni F, Hou J, Wild P, Kensche T, Grecco HE, Bastiaens P, Dikic I (2012) Fluorescence-based sensors to monitor localization and functions of linear and K63-linked ubiquitin chains in cells. Mol Cell 47(5):797–809. doi:10.1016/j.molcel.2012.06.017
Acknowledgement
M.L. holds a predoctoral fellowship from the Fund for Scientific Research Flanders (FWO). Work in the authors’ lab is supported by grants from the FWO, “Belgian Foundation Against Cancer”, “Agency for Innovation by Science and Technology”, “Interuniversity Attraction Poles”, “Concerted Research Actions”, and “Group-ID Multidisciplinary research partnership” from Ghent University, VIB. The authors would like to acknowledge networking support by the Proteostasis COST Action (BM1307).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer Science+Business Media New York
About this protocol
Cite this protocol
Lork, M., Delvaeye, M., Gonçalves, A., Van Hamme, E., Beyaert, R. (2016). Monitoring Ubiquitin-Coated Bacteria via Confocal Microscopy. In: Matthiesen, R. (eds) Proteostasis. Methods in Molecular Biology, vol 1449. Humana Press, New York, NY. https://doi.org/10.1007/978-1-4939-3756-1_14
Download citation
DOI: https://doi.org/10.1007/978-1-4939-3756-1_14
Published:
Publisher Name: Humana Press, New York, NY
Print ISBN: 978-1-4939-3754-7
Online ISBN: 978-1-4939-3756-1
eBook Packages: Springer Protocols