
Abstract
Knowledge of plant nutrient strategies is crucial for defining and predicting the patterns and mechanisms resulting from nitrogen (N) deposition. However, the impacts of N enrichment on plant nutrient strategies are unclear in global grasslands. We conducted a meta-analysis of 127 publications to synthesize the pathways underlying the responses of plant nutrient concentration and resorption to N addition across global grassland ecosystems. Our analysis indicated that N addition increased the N concentration in green and senesced leaves, the phosphorus (P) concentration in senesced leaves, and aboveground and belowground biomass by about 32%, 50%, 7%, 74%, and 19%, respectively. Meanwhile, it reduced N resorption efficiency (NRE) and P resorption efficiency (PRE) by about 9% and 6%, respectively. Nitrogen addition did not significantly affect green leaf P concentration. These responses were modulated by N application rates and humidity, and they differed among grassland types, plant groups, fertilizer types, and experimental durations. Nitrogen addition changed the relationship between N and P in green leaves and between NRE and PRE, but it did not alter the N:P ratio in senesced leaves. Our results suggest that N addition affects leaf nutrient concentrations and resorption in global grassland ecosystems, although such effects vary among grassland types and among plant functional groups. Nutrient resorption may be a critical pathway that mediates plant regulation of the coupled N:P balance. Changes in humidity due to climate change also mediate the response of plant nutrients to N addition and thereby affect the soil–plant nutrient cycles of grassland ecosystems under future N enrichment.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Nitrogen (N) is one of the essential and limiting nutrients for plant growth and development in terrestrial ecosystems (Elser et al. 2007; Yuan and Chen 2012; Moreau et al. 2019). Thus, moderate N enrichment may have a positive impact on plant growth and terrestrial ecosystem productivity (LeBauer and Treseder 2008; Bobbink et al. 2010; Tian et al. 2020). However, extensive atmospheric N deposition, owing to human activities (e.g., fertilization and urbanization), has occurred in current terrestrial ecosystems and continues to increase on a global scale (Lamarque et al. 2005; Elser and Bennett 2011; Dietrich et al. 2017). These actions stimulate plant growth and affect plant nutrient cycling (Kallenbach et al. 2017; Bellenger et al. 2020; Iqbal et al. 2020; Tognetti et al. 2021). Therefore, it is crucial to fully understand the changes in terrestrial ecosystem responses to future N deposition (Zaehle and Dalmonech 2011; Greaver et al. 2016; Soong et al. 2020).
Plant nutrient concentration reflects plant growth, nutrient uptake, and use strategies in terrestrial ecosystems, which are collectively affected by N enrichment (Yuan and Chen 2009; Lihavainen et al. 2016). However, this issue is still controversial, although most previous studies have focused on the effect of N enrichment on changes in plant nutrient concentration. For example, field experiments reported varied effects of N enrichment on plant P levels, either increasing (Lu et al. 2013; Carate-Tandalla et al. 2018; Gonzales and Yanai 2019), decreasing (Sardans et al. 2016; Tiruvaimozhi et al. 2018; Liu et al. 2021a), or not affecting (Zong et al. 2018; Tian et al. 2020) P concentration. Nitrogen addition also had positive, negative, and neutral effects on the plant leaf N and P concentrations in the same ecosystem (Lu et al. 2013; Su et al. 2021). These differences may result from the comprehensive effects of multiple factors such as species (Van Heerwaarden et al. 2003), soil nutrient conditions (Gusewell 2005), N addition rates, experimental duration (Hao et al. 2018), and environmental factors. Our study complements the limited knowledge about the complex responses of plant nutrient concentration and nutrient resorption to N addition in varying grassland types and among plant functional groups.
Plants reduce their dependence on external nutrients and enhance their survival and adaptability by resorbing nutrients from senesced tissues (Yuan et al. 2006; Gerdol et al. 2019). This process can be quantified by computing the nutrient resorption efficiency (Milla et al. 2005). Previous studies reported that N addition reduced the nitrogen resorption efficiency (NRE) owing to the increase in soil N availability (Soudzilovskaia et al. 2007; Ren et al. 2015; Li et al. 2016). However, the effect of N addition on phosphorus resorption efficiency (PRE) has not been determined in grassland ecosystems (Lu et al. 2013; Yuan and Chen 2015a, b; Zheng et al. 2018). Nitrogen addition to an ecosystem was thought to transform N limitation to P limitation or co-limitation of N and P (Dong et al. 2019). It is unclear how N addition affects nutrient resorption efficiency in grassland ecosystems, which are usually limited by P (Van Dobben et al. 2017). Therefore, knowledge of nutrient resorption responses to N enrichment has great significance for managing plant nutrient conservation and plant productivity under future global environmental changes.
Grasslands account for approximately 26% of the global terrestrial area (Obermeier et al. 2016). Extensive atmospheric N deposition inevitably affected plant nutrient concentrations and nutrient uptake strategies in grassland ecosystems (Sattari et al. 2016; Shi et al. 2021; Hu et al. 2022). Many studies reported that different N addition rates produced varying responses of leaf nutrient concentrations and nutrient resorption (Cerasoli et al. 2018; Bai et al. 2019; Graff et al. 2020). However, the patterns and mechanisms of leaf nutrient concentrations and nutrient resorption responses to N addition in grassland ecosystems have not been defined, particularly the complex plant responses to changes in multiple biotic and abiotic factors. Therefore, we conducted a meta-analysis to integrate the available data from different field experiments in global grassland ecosystems and define the responses of grassland leaf nutrients and nutrient resorption to N addition.
Here, by using a weighted meta-analysis of a global data set of 1935 observations at 98 sites from 127 publications based on N addition, we examined how leaf nutrient concentration and nutrient resorption respond to N addition in grassland ecosystems. We tested three hypotheses. (1) Nitrogen addition enhances the N concentration in green and senesced leaves due to an N-induced increase in soil N availability (Soudzilovskaia et al. 2007; Li et al. 2016). The corresponding P concentration in green and senesced leaves will also increase to maintain the balance of N and P (You et al. 2018b). (2) Plants will reduce N resorption from senesced tissues due to increased soil N availability (Ren et al. 2018), thereby reducing NRE but not PRE (to balance N and P). (3) Numerous experimental and environmental factors alter leaf nutrient and resorption responses to N addition by modulating plant nutrient cycle patterns.
2 Material and Methods
2.1 Data Collection
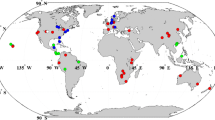
We collected peer-reviewed publications that reported changes in plant nutrient concentrations and resorption efficiency in global grassland ecosystems under the addition of N to fields. We performed Boolean searches of the Web of Science (http://apps.webofknowledge.com) and the China National Knowledge Infrastructure (CNKI, https://www.cnki.net) databases using the following keywords: (a) ‘N addition’ or ‘nitrogen addition’ or ‘nitrogen amendment’ or ‘nitrogen deposition’, (b) ‘leaf N’ or ‘leaf nutrient’, (c) ‘leaf P’ or ‘leaf nutrient’, (d) ‘NRE’ or ‘nutrient resorption’, (e) ‘PRE’ or ‘nutrient resorption’, (f) ‘AGB’ or ‘aboveground biomass’, (g) ‘BGB’ or ‘belowground biomass’, and (h) ‘grassland’ or ‘grassland ecosystem’. Seven criteria were used to select suitable publications: (1) Experiments were conducted on global grassland ecosystems; (2) experiments were conducted in the field; (3) studies reported comparisons between controls (i.e., without nitrogen addition) and treatments (i.e., nitrogen addition); (4) studies reported means, standard deviations (SD) or standard errors (SE), and sample sizes of the selected variables; (5) studies reported the treatment method, magnitude, and duration; (6) the publications were peer-reviewed journal articles, conference collections, theses, or dissertations; and (7) where data were published in different papers for studies at the same site, we only reserved one recent publication to ensure the relative independence of data. These search and selection criteria yielded 1935 experimental observations from 127 papers at 98 sites across the globe (Fig. 1; Supplementary information, data sources list).

Map of sites conducting field studies of nitrogen addition in grasslands that were included in the meta-analysis. Points with different colors represent different N application rates (g N m−2 year−1) in each study site
We performed a stratified analysis to evaluate whether grassland type, plant group, fertilizer type, and experimental duration affected plant nutrient concentration and resorption efficiency responses to N addition. The data classifications included the following categories: Grassland types were categorized into temperate grassland and alpine grassland; plant functional group was categorized into grass and forb; experimental duration was categorized into short-, intermediate- and long-term (i.e., < 3 years, 3–6 years, and > 6 years, respectively), and fertilizer type was categorized into urea and NH4NO3. We extracted site information related to the experiments to construct an integrated database of geographic variables (longitude, latitude, and altitude) and climatic factors [mean annual temperature (MAT) and mean annual precipitation (MAP)]. Data presented in the figures were extracted using GetData version 2.20 (http://www.getdata-graph-digitizer.com). We investigated how the climatic factors affected the plant nutrient concentration and resorption efficiency responses to N addition using the De Martonne aridity index to compute the humidity combined with MAT and MAP data at each experimental site [aridity index = MAP/(MAT + 10)] as described in previous studies (Song et al. 2019; Su et al. 2021).
2.2 Data Analysis
We conducted a meta-analysis to determine the effects of N addition on plant nutrient concentrations and resorption efficiency in global grasslands. The natural logarithm of the response ratio (lnRR) was calculated to indicate the effect size of each treatment (Hedges et al. 1999) [Eq. (1)]:
where \({\overline{X} }_{T}\) and \({\overline{X} }_{C}\) are the mean treatment and control values, respectively. The variance (v) of each lnRR was calculated using Eq. (2):
where \({n}_{T}\) and \({n}_{C}\) are the sample sizes, and \({S}_{T}\) and \({S}_{C}\) are the SD of means for each treatment and control. Most of the selected studies reported SE, which was transformed to SD according to Eq. (3):
where \(n\) was the sample size. Based on previous studies, the lnRR was calculated separately for each control-treatment pair and treated as independent data when data were extracted from multifactor experiments with multiple single-factor treatments and a single control (Lajeunesse 2011; Song et al. 2019).
The weighted response ratio (lnRR++) and bias-corrected 95% bootstrap-confidence interval (CI) were calculated using inverse-variance weighted regressions and random-effects models with the rma function in the “metafor” package version 3.0–2 of R version 4.1.2 (The R Project for Statistical Computing, https://www.r-project.org/) (Hedges et al. 1999). The effects of treatments on selected variables were considered statistically significant if the 95% CI did not overlap zero, whereas the effects between groups or under different conditions differed if their 95% CIs did not overlap. To clarify data interpretation, the percentage change (%) was calculated based on weighted response ratios using the equation [exp (lnRR++) – 1] × 100 (Yan et al. 2020). We computed multiple comparisons to examine differences in treatment effects on different groups or under different conditions. Statistical results were reported as differences among group cumulative effect size (QB) and residual error (QE). Regression analysis (including univariate covariance analysis) was conducted to examine the effects of N addition rate and environmental factors (e.g., aridity index) on the response ratio of plant nutrient concentration and resorption efficiency under N addition and evaluate the relationships of response ratios of objective variables (e.g., green and senesced leaf N and P). Statistical differences were considered significant when P < 0.05. All statistical analyses were performed in R version 4.1.2.
3 Results
3.1 Effects of N Addition on Leaf Nutrient Concentration and Resorption
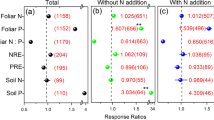
Nitrogen addition significantly increased the concentrations of leaf N, green leaf N, senesced leaf N, senesced leaf P, AGB, and BGB by 29%, 32%, 50%, 7%, 74%, and 19%, respectively (P < 0.001, Fig. 2). By contrast, N addition reduced NRE and PRE by 9% and 5%, respectively (P < 0.001, Fig. 2). Nitrogen addition did not significantly affect the concentrations of total leaf P and green leaf P (P > 0.05, Fig. 2). Regression analysis identified significant and positive relationships between the concentrations of green leaf N and P, senesced leaf N and P, and NRE and PRE. Regression analysis also identified significant differences in the relationships between NRE and PRE, and green leaf N and P in the control and N enrichment plots, but the relationship between N and P in senesced leaves in the control and N enrichment plots did not significantly differ (Fig. 3a–d, Supplementary Table S1).

Responses of leaf nutrient concentrations, nutrient resorption, and biomass to experimental nitrogen addition in global grassland ecosystems. N, leaf nitrogen concentration; P, leaf phosphorus concentration; Ng, green leaf nitrogen concentration; Pg, green leaf phosphorus concentration; Ns, senesced leaf nitrogen concentration; Ps, senesced leaf phosphorus concentration; AGB, aboveground biomass; BGB, belowground biomass; NRE, nitrogen resorption efficiency; PRE, phosphorus resorption efficiency. Error bars represent 95% confidence intervals (CI). The vertical dashed line represents the response ratio = 0. Treatment effects were statistically significant (denoted by *) if 95% CI did not overlap zero. *, **, and *** indicate significant correlations at p < 0.05, p < 0.01, and p < 0.001, respectively. The sample size for each variable is given in parentheses

Relationships between (a) leaf nitrogen (N) and phosphorus (P), (b) green leaf N and P, (c) senesced leaf N and P, and (d) nitrogen resorption efficiency (NRE) and phosphorus resorption efficiency (PRE) under control (N0) and N addition (N) conditions
3.2 Effects of N Enrichment on Leaf Nutrient Concentration and Resorption Among Varying Subgroups
Subgroup analysis indicated that the responses of leaf nutrient concentration and nutrient resorption to N addition differed in subgroups (Fig. 4, Supplementary Table S2). Nitrogen addition increased the green and senesced leaf N concentration in temperate and alpine grasslands (P < 0.001, Fig. 4a,b), but the responses were more sensitive in alpine than in temperate grassland. The N addition impacts on leaf P concentration in different grassland types varied among green and senesced leaves (Fig. 4d,e). Nitrogen addition decreased NRE and PRE in temperate grassland, but did not significantly affect NRE and PRE in alpine grassland (Fig. 4c,f, Supplementary Table S2).

Subgroup analysis of the response of leaf nutrient concentrations and nutrient resorption to experimental nitrogen addition in global grassland ecosystems. Ng, green leaf nitrogen concentration; Ns, senesced leaf nitrogen concentration; Pg, green leaf phosphorus concentration; Ps, senesced leaf phosphorus concentration; NRE, nitrogen resorption efficiency; PRE, phosphorus resorption efficiency. Error bars represent 95% confidence intervals (CI). The vertical dashed line represents the response ratio = 0. Treatment effects were statistically significant (denoted by *) if 95% CI did not overlap zero. *, **, and *** indicate significant correlations at p < 0.05, p < 0.01, and p < 0.001, respectively. The number represents the sample size of each variable
Nitrogen addition enhanced green and senesced leaf N concentration in both grass and forb (P < 0.001, Fig. 4a,b), but grass had a higher response ratio than forb. The effect of N addition on green leaf P concentration was opposite in grass and forb (i.e., decreased in grass and increased in forb; P < 0.05, Fig. 4d), while N addition impact on senesced leaf P concentration was not significantly different between grass and forb (P > 0.05, Fig. 4e). Nitrogen addition significantly reduced NRE and PRE in both grass and forb, while forb had greater responsiveness than grass (P < 0.001, Fig. 4c,f). Ammonium nitrate and urea increased the green and senesced leaf N concentration, but the NH4NO3 effect on plant N concentration was stronger than that of urea (Fig. 4a,b, Supplementary Table S2). Ammonium nitrate increased the senesced leaf P concentration (P < 0.001), whereas urea did not (P > 0.05, Fig. 4e). The effect of NH4NO3 and urea on green leaf P concentration was not significantly different (P > 0.05, Fig. 4d). Ammonium nitrate and urea both reduced NRE (P < 0.001, Fig. 4c), but the effect of urea on NRE was greater than that of NH4NO3. Phosphorus resorption efficiency was only reduced by NH4NO3 (P < 0.001, Fig. 4f). Long-term experiments yielded larger increases in senesced leaf N concentration than short- and intermediate-term experiments. Long-term experiments increased senesced leaf P and reduced green leaf P, whereas short-term and intermediate-term experiments did not affect green leaf P. Larger increases in green leaf N and decreases in NRE were observed in intermediate-term and short-term experiments, respectively (Fig. 4a,c).
3.3 Effects of N Application Rates and Humidity Conditions on Leaf Nutrient Concentration and Nutrient Resorption Responses to N Addition
The response ratios exhibited significant differences under different N application rates (Fig. 5). The aboveground biomass and green and senesced leaf N concentrations enhanced with increasing N addition rates (Fig. 5a,b,g), and the maximum occurred at the N application rate of ~ 40 g N m−2 year−1. Phosphorus resorption efficiency decreased with the increasing N addition rates (Fig. 5f), and NRE also decreased to a minimum at the N application rate of ~ 40 g N m−2 year−1 (Fig. 5e). Our results indicated that the effects of N addition on green and senesced leaf P concentrations were shifted from decrease to increase when the N application rate was ~ 10 g N m−2 year−1 (Fig. 5c,d). The green and senesced leaf N concentration response to N addition was enhanced with increasing aridity index (Fig. 6a,b), whereas the green and senesced leaf P concentration decreased with increasing aridity index (Fig. 6c,d). The aridity index did not significantly affect NRE and PRE responses to N addition (Fig. 6e,f).

Relationships of the natural logarithm of the response ratio (lnRR) of leaf nutrient concentrations, nutrient resorption, and biomass with changes in the nitrogen application rates. Ng, green leaf nitrogen concentration; Ns, senesced leaf nitrogen concentration; Pg, green leaf phosphorus concentration; Ps, senesced leaf phosphorus concentration; AGB, aboveground biomass; BGB, belowground biomass; NRE, nitrogen resorption efficiency; PRE, phosphorus resorption efficiency

Relationships of the natural logarithm of the response ratio (lnRR) of leaf nutrient concentration, nutrient resorption, and biomass with changes in the aridity index [aridity index = MAP/(MAT + 10)]. MAP, mean annual precipitation; MAT, mean annual temperature; Ng, green leaf nitrogen concentration; Ns, senesced leaf nitrogen concentration; Pg, green leaf phosphorus concentration; Ps, senesced leaf phosphorus concentration; AGB, aboveground biomass; BGB, belowground biomass; NRE, nitrogen resorption efficiency; PRE, phosphorus resorption efficiency
4 Discussion
Our global data analyses of N addition experiments indicate that N addition alters leaf nutrient uptake strategies and patterns. The relationship between green leaf N and P and between NRE and PRE changed under N addition, but the relationship between senesced leaf N and P was not altered. Green leaves in grassland ecosystems tend to have higher N:P ratios. These results verify that nutrient resorption may be an important pathway regulating the relationships between leaf N and P concentrations. The multiple responses of varying grassland types and plant groups to N addition also highlight the complex impacts of future N deposition on global grasslands.
4.1 Nitrogen Addition Alters Leaf Nutrient Concentrations and Resorption in Grasslands
Consistent with our first hypothesis, N addition significantly enhanced the green and senesced leaf N concentration. This was primarily attributed to enhanced soil N availability (Yuan and Chen 2015a; Taylor et al. 2021). These results are also consistent with previous global-scale synthesis, which demonstrated that N addition significantly increased foliar N concentration under all nutrient-limited conditions and increased the foliar P concentration under P-limited conditions (You et al. 2018a). However, the results in our study demonstrated that N addition only increased P concentration in senesced leaves across global grasslands universally limited by P (Van Dobben et al. 2017). The nonsignificant change of P concentration in green leaves under N addition may be due to the dilution of green leaf P concentration caused by N-induced enhancement of aboveground biomass (Sardans et al. 2016). Overall, these findings suggested that plants alter the nutrient uptake strategies and tend to have higher N:P ratios in green leaves across grassland ecosystems under future N enrichment.
Our analysis also revealed that N addition reduced NRE in grassland ecosystems, which confirmed our second hypothesis. Nitrogen input enhanced soil N availability and reduced N resorption from senesced tissues (Soudzilovskaia et al. 2007; Li et al. 2016). Previous studies also reported the consistent results that N addition significantly reducing the NRE for all plant growth types by about 12–13% on a global scale (Yuan and Chen 2015b; You et al. 2018b). However, this is slightly higher than the 9% reduction of NRE under N addition we found in grassland ecosystems. This may be due to the limitation of soil nutrient availability across global grassland (Craine and Jackson 2010). Plants tend to have high nutrient resorption efficiency to maintain their growth under limited soil nutrient availability (Wright and Cannon 2001; Zong et al. 2018). The results of our study also indicated that N addition reduced PRE, which was in agreement with previous reports of N addition reducing PRE in forest, grassland, and shrubland ecosystems (Yuan and Chen 2015b; Su et al. 2021). This suggested that soil was the primary P source for plants under N addition, even in P-limited grassland ecosystems. Previous studies also showed that N addition leads to an imbalance in N and P cycles in soil and plants during biogeochemical processes (Penuelas et al. 2012) and can decouple N and P cycles in plants (Lü et al. 2016; Yan et al. 2018). However, the results in our analysis showed that plant N and P cycles remained coupled under N enrichment in global grassland ecosystems, whereas the relationships between NRE and PRE, and between green leaf N and P concentrations, changed significantly under N enrichment. Early studies reported that nutrient resorption had a key role in maintaining the coupled balance between N and P cycles in senesced leaves (Lü et al. 2016). Plants uptake critical nutrients from senesced leaves, which reduces their dependence on soil nutrient availability (Killingbeck 1996). This can explain why PRE was reduced under N enrichment. Nutrient resorption from senesced leaves may be an important pathway regulating the coupled relationship between N and P cycles.
4.2 Nitrogen Addition Differentially Affected Leaf Nutrient Concentrations and Nutrient Resorption in Different Subgroups
Our results indicated that N addition differentially affected leaf nutrient concentrations and nutrient resorption in temperate and alpine grasslands. Alpine grasslands displayed more sensitivity in leaf N concentration changes under N addition than temperate grasslands. Plants in alpine environments tend to have efficient nutrient uptake and utilization strategies to adapt to nutrient limitations under extreme environments (Zong et al. 2017; Zimmer et al. 2018). Combined with the decrease of green leaf P and unchanged senesced leaf P under N addition in alpine grasslands, it suggests that alpine plants will tend to have a higher N:P ratio than temperate plants under N addition. Generally, a higher N:P ratio means that N is abundant while P is relatively deficient, which suggests that plant growth is limited by P availability (Zong et al. 2018). Alpine grasslands will probably show a more severe limitation of P availability than temperate grasslands under future N enrichment. By contrast, temperate grasslands displayed greater sensitivity in nutrient resorption efficiency under N addition than alpine grasslands. Nutrient resorption efficiency reflects soil nutrient conditions and the plant adaptation strategies to soil nutrient availability in different environments (Kobe et al. 2005). The nonsignificant response of nutrient resorption efficiency to N addition in alpine grasslands may be due to the low soil availability of N and P, which was likely induced by the inhibition of enzyme and microbial activities caused by low temperature under high altitude (Liu et al. 2021b). The increase of plant biomass in alpine grassland under N enrichment means that plants demand to acquire extensive nutrients, which is obviously difficult for alpine grassland soil to satisfy. Therefore, alpine plants still need to uptake substantial nutrients from senesced tissues to meet their own growth needs even under N addition. Plants in alpine grasslands tended to display stronger dependence on nutrient resorption from senesced tissues than those in temperate grasslands under N addition.
The green leaf N concentration displayed a greater increase under N addition in grasses than in forbs, but nutrient resorption efficiency displayed a greater decrease under N addition in forbs than in grasses. As one of the dominant functional groups in grassland ecosystems, grasses always have higher nutrient resorption efficiency than other species (Zong et al. 2018), which is an effective strategy for the successful competition of grasses with other species. The greater decrease in green leaf P concentration of grass than that of forb under N addition may be due to higher P utilization efficiency in grasses than in forbs (Hayes et al. 2018; Pereira et al. 2018). These results demonstrated that grasses possess a higher N:P ratio under N addition than forbs. This suggests that P limitation in grasses under future N deposition may be more pronounced than in forbs in global grasslands. Long-term N addition and NH4NO3 fertilizer were more likely to shift the leaf nutrient in grassland ecosystems, as NH4NO3 is more soluble in water and more easily acquired and utilized by plants. In summary, our analyses indicated that the responses of leaf nutrient concentrations and nutrient resorption to N addition differed among grassland types, plant groups, and experimental conditions. These results will deepen our understanding of the effects of N enrichment in global grassland ecosystems.
4.3 Factors that Shift the Impact of N Addition on Leaf Nutrient Concentrations and Nutrient Resorption
Consistent with our third hypothesis, the responses of leaf nutrient concentration and nutrient resorption to N addition were modulated by factors such as N application rates and humidity. Our analysis showed that N application rates of ~ 40 g N m−2 year−1 had greater impacts on leaf N concentration and NRE than other N application rates. The plant N uptake gradually increased due to enhanced soil N availability when the N addition rate was below the critical threshold, whereas plant N uptake was limited by the availability of other nutrients such as carbon and phosphorus when the N addition rate was above the critical threshold (Yuan and Chen 2015a, b). This similar critical threshold was also reflected in our analysis of the response of aboveground biomass to N addition. Our results also exhibited that the turning point from decrease to increase in leaf P concentration to N addition was when the N application rate was ~ 10 g N m−2 year−1, which was consistent with the results of a previous study of grasslands across northern China (Su et al. 2021). Low N addition (< 10 g N m−2 year1) tends to inhibit plant uptake of P, but high N addition (> 10 g N m−2 year−1) stimulates plant P uptake. This also may be why soil available P differed due to varying responses of soil phosphatase activity to N application rates (Gong et al. 2020; Widdig et al. 2020). Moreover, previous studies reported that N addition increased aboveground biomass by 20–35% and the effects of N addition on aboveground biomass had a unimodal distribution (Liu and Greaver 2010; Jiang et al. 2019). These responses are also reflected in our analysis. Dilution effects resulting from the increase in plant biomass may also partly explain the complex responses of leaf P concentration to N addition due to mismatched changes in plant biomass and soil phosphatase activity (Yuan and Chen 2015a). However, the mechanism of a critical threshold for the changes in plant P concentration to N addition still needs to be further studied.
Humidity (aridity index) also shifted N addition impacts on leaf nutrient concentrations. Our results indicated that the response of green and senesced leaf N to N addition increased with greater aridity index, whereas the response of green and senesced leaf P to N addition decreased with rising aridity index. This may result from a synergy between N enrichment and humidity, which was reported in previous studies (Grunzweig and Korner 2003; Copeland et al. 2012). Grassland ecosystems can be particularly sensitive to changes in nutrient and water availability (Su et al. 2021). An increase in soil moisture usually led to a higher N mineralization rate and improved the availability of soil inorganic N, which further promoted plant aboveground biomass and nitrogen absorption (Schuster and Dukes 2017; Dijkstra et al. 2018). However, this synergy may also lead to an inhibition of plant growth induced by excessive nitrogen and water. The aridity index did not affect plant biomass and nutrient resorption efficiency responses to N addition, which may be due to the multiple responses of different grassland types and plant groups to the effects of N addition and water availability. For example, N addition and water availability impacts on leaf nutrient concentration and nutrient resorption of grass and forb plants were different in semi-arid grassland, and the distinction also was reflected in varying grassland ecosystems (Lu and Han 2010; Zhang et al. 2019). Overall, our combined results indicated that temperature and precipitation jointly modulate the responses of plant nutrient concentrations and nutrient resorption to N addition in grassland ecosystems.
5 Conclusions
Our meta-analysis synthesized data on changes in leaf nutrient concentration and resorption across global grassland ecosystems under experimental N addition. The results suggested that N enrichment altered leaf nutrient concentration and nutrient resorption in grassland ecosystems, which was modulated by N application rates, temperature, and precipitation. The sensitivity of leaf nutrient concentration and resorption to N enrichment varied greatly across grassland types and plant groups. These findings provide evidence that nutrient resorption plays a significant role in regulating the plant nutrient strategies to respond to future N deposition. Our results contribute to better predictions of changes in the plant nutrient cycle under N enrichment in global grassland. Future changes in temperature and precipitation may affect the soil–plant nutrient cycles of global grassland ecosystems by mediating plant nutrient responses to N addition.
References
Bai W, Wang G, Xi J, Liu Y, Yin P (2019) Short-term responses of ecosystem respiration to warming and nitrogen addition in an alpine swamp meadow. Eur J Soil Biol 92:16–23. https://doi.org/10.1016/j.ejsobi.2019.04.003
Bellenger JP, Darnajoux R, Zhang X, Kraepiel AML (2020) Biological nitrogen fixation by alternative nitrogenases in terrestrial ecosystems: a review. Biogeochemistry 149:53–73. https://doi.org/10.1007/s10533-020-00666-7
Bobbink R, Hicks K, Galloway J, Spranger T, Alkemade R, Ashmore M, Bustamante M, Cinderby S, Davidson E, Dentener F, Emmett B, Erisman JW, Fenn M, Gilliam F, Nordin A, Pardo L, De Vries W (2010) Global assessment of nitrogen deposition effects on terrestrial plant diversity: a synthesis. Ecol Appl 20:30–59. https://doi.org/10.1890/08-1140.1
Carate-Tandalla D, Camenzind T, Leuschner C, Homeier J (2018) Contrasting species responses to continued nitrogen and phosphorus addition in tropical montane forest tree seedlings. Biotropica 50:234–245. https://doi.org/10.1111/btp.12518
Cerasoli S, Campagnolo M, Faria J, Nogueira C, Caldeira MdC (2018) On estimating the gross primary productivity of Mediterranean grasslands under different fertilization regimes using vegetation indices and hyperspectral reflectance. Biogeosciences 15:5455–5471. https://doi.org/10.5194/bg-15-5455-2018
Copeland SM, Bruna EM, Barbosa Silva LV, Mack MC, Vasconcelos HL (2012) Short-term effects of elevated precipitation and nitrogen on soil fertility and plant growth in a Neotropical savanna. Ecosphere 3:1–20. https://doi.org/10.1890/es11-00305.1
Craine JM, Jackson RD (2010) Plant nitrogen and phosphorus limitation in 98 North American grassland soils. Plant Soil 334:73–84. https://doi.org/10.1007/s11104-009-0237-1
Dietrich K, Spohn M, Villamagua M, Oelmann Y (2017) Nutrient addition affects net and gross mineralization of phosphorus in the organic layer of a tropical montane forest. Biogeochemistry 136:223–236. https://doi.org/10.1007/s10533-017-0392-z
Dijkstra FA, Carrillo Y, Blumenthal DM, Mueller KE, LeCain DR, Morgan JA, Zelikova TJ, Williams DG, Follett RF, Pendall E (2018) Elevated CO2 and water addition enhance nitrogen turnover in grassland plants with implications for temporal stability. Ecol Lett 21:674–682. https://doi.org/10.1111/ele.12935
Dong C, Wang W, Liu H, Xu X, Zeng H (2019) Temperate grassland shifted from nitrogen to phosphorus limitation induced by degradation and nitrogen deposition: evidence from soil extracellular enzyme stoichiometry. Ecol Indicators 101:453–464. https://doi.org/10.1016/j.ecolind.2019.01.046
Elser J, Bennett E (2011) A broken biogeochemical cycle. Nature 478:29–31. https://doi.org/10.1038/478029a
Elser J, Bracken ME, Cleland EE, Gruner DS, Harpole WS, Hillebrand H, Ngai JT, Seabloom EW, Shurin JB, Smith JE (2007) Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol Lett 10:1135–1142. https://doi.org/10.1111/j.1461-0248.2007.01113.x
Gerdol R, Iacumin P, Brancaleoni L (2019) Differential effects of soil chemistry on the foliar resorption of nitrogen and phosphorus across altitudinal gradients. Funct Ecol 33:1351–1361. https://doi.org/10.1111/1365-2435.13327
Gong SW, Zhang T, Guo J (2020) Warming and nitrogen deposition accelerate soil phosphorus cycling in a temperate meadow ecosystem. Soil Res 58:109–115. https://doi.org/10.1071/sr19114
Gonzales K, Yanai R (2019) Nitrogen-phosphorous interactions in young northern hardwoods indicate P limitation: foliar concentrations and resorption in a factorial N by P addition experiment. Oecologia 189:829–840. https://doi.org/10.1007/s00442-019-04350-y
Graff P, Gundel PE, Salvat A, Cristos D, Chaneton EJ, Emery N (2020) Protection offered by leaf fungal endophytes to an invasive species against native herbivores depends on soil nutrients. J Ecol 108:1592–1604. https://doi.org/10.1111/1365-2745.13371
Greaver TL, Clark CM, Compton JE, Vallano D, Talhelm AF, Weaver CP, Band LE, Baron JS, Davidson EA, Tague CL, Felker-Quinn E, Lynch JA, Herrick JD, Liu L, Goodale CL, Novak KJ, Haeuber RA (2016) Key ecological responses to nitrogen are altered by climate change. Nat Clim Chang 6:836–843. https://doi.org/10.1038/nclimate3088
Grunzweig JM, Korner C (2003) Differential phosphorus and nitrogen effects drive species and community responses to elevated CO2 in semi-arid grassland. Funct Ecol 17:766–777. https://doi.org/10.1111/j.1365-2435.2003.00797.x
Gusewell S (2005) Nutrient resorption of wetland graminoids is related to the type of nutrient limitation. Funct Ecol 19:344–354. https://doi.org/10.1111/j.0269-8463.2005.00967.x
Hao T, Song L, Goulding K, Zhang F, Liu X (2018) Cumulative and partially recoverable impacts of nitrogen addition on a temperate steppe. Ecol Appl 28:237–248. https://doi.org/10.1002/eap.1647
Hayes PE, Clode PL, Oliveira RS, Lambers H (2018) Proteaceae from phosphorus-impoverished habitats preferentially allocate phosphorus to photosynthetic cells: an adaptation improving phosphorus-use efficiency. Plant Cell Environ 41:605–619. https://doi.org/10.1111/pce.13124
Hedges LV, Gurevitch J, Curtis PS (1999) The meta-analysis of response ratios in experimental ecology. Ecology 80:1150–1156. https://doi.org/10.2307/177062
Hu W, Tan J, Shi X, Lock TR, Kallenbach RL, Yuan Z (2022) Nutrient addition and warming alter the soil phosphorus cycle in grasslands: a global meta-analysis. J Soils Sed. https://doi.org/10.1007/s11368-022-03276-y
Iqbal S, Xu J, Allen SD, Khan S, Nadir S, Arif MS, Yasmeen T (2020) Unraveling consequences of soil micro- and nano-plastic pollution on soil-plant system: implications for nitrogen (N) cycling and soil microbial activity. Chemosphere 260:127578. https://doi.org/10.1016/j.chemosphere.2020.127578
Jiang J, Wang Y-P, Yang Y, Yu M, Wang C, Yan J (2019) Interactive effects of nitrogen and phosphorus additions on plant growth vary with ecosystem type. Plant Soil 440:523–537. https://doi.org/10.1007/s11104-019-04119-5
Kallenbach R, Roberts C, Lory J, Hamilton S (2017) Nitrogen fertilization rates influence stockpiled tall fescue forage through winter. Crop Sci 57:1732–1741. https://doi.org/10.2135/cropsci2016.02.0097
Killingbeck KT (1996) Nutrients in senesced leaves: keys to the search for potential resorption and resorption proficiency. Ecology 77:1716–1727. https://doi.org/10.2307/2265777
Kobe RK, Lepczyk CA, Iyer M (2005) Resorption efficiency decreases with increasing green leaf nutrients in a global data set. Ecology 86:2780–2792. https://doi.org/10.1890/04-1830
Lajeunesse MJ (2011) On the meta-analysis of response ratios for studies with correlated and multi-group designs. Ecology 92:2049–2055. https://doi.org/10.1890/11-0423.1
Lamarque JF, Kiehl JT, Brasseur GP, Butler T, Cameron-Smith P, Collins WD, Collins WJ, Granier C, Hauglustaine D, Hess PG, Holland EA, Horowitz L, Lawrence MG, McKenna D, Merilees P, Prather MJ, Rasch PJ, Rotman D, Shindell D, Thornton P (2005) Assessing future nitrogen deposition and carbon cycle feedback using a multimodel approach: analysis of nitrogen deposition.J Gerontol Ser A Biol Med Sci 110.https://doi.org/10.1029/2005jd005825
LeBauer DS, Treseder KK (2008) Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 89:371–379. https://doi.org/10.1890/06-2057.1
Li L, Gao X, Li X, Lin L, Zeng F, Gui D, Lu Y (2016) Nitrogen (N) and phosphorus (P) resorption of two dominant alpine perennial grass species in response to contrasting N and P availability. Environ Exp Bot 127:37–44. https://doi.org/10.1016/j.envexpbot.2016.03.008
Lihavainen J, Ahonen V, Keski-Saari S, Kontunen-Soppela S, Oksanen E, Keinanen M (2016) Low vapour pressure deficit affects nitrogen nutrition and foliar metabolites in silver birch. J Exp Bot 67:4353–4365. https://doi.org/10.1093/jxb/erw218
Liu L, Greaver TL (2010) A global perspective on belowground carbon dynamics under nitrogen enrichment. Ecol Lett 13:819–828. https://doi.org/10.1111/j.1461-0248.2010.01482.x
Liu M, Shen Y, Li Q, Xiao W, Song X (2021a) Arbuscular mycorrhizal fungal colonization and soil pH induced by nitrogen and phosphorus additions affects leaf C:N: P stoichiometry in Chinese fir (Cunninghamia lanceolata) forests. Plant Soil 461:421–440. https://doi.org/10.1007/s11104-021-04831-1
Liu Y, Li L, Li X, Yue Z, Liu B (2021b) Effect of nitrogen and phosphorus addition on leaf nutrient concentrations and nutrient resorption efficiency of two dominant alpine grass species. J Arid Land 13:1041–1053. https://doi.org/10.1007/s40333-021-0080-7
Lu X-T, Han X-G (2010) Nutrient resorption responses to water and nitrogen amendment in semi-arid grassland of Inner Mongolia, China. Plant Soil 327:481–491. https://doi.org/10.1007/s11104-009-0078-y
Lu X, Reed S, Yu Q, He N, Wang Z, Han X (2013) Convergent responses of nitrogen and phosphorus resorption to nitrogen inputs in a semiarid grassland. Global Change Biol 19:2775–2784. https://doi.org/10.1111/gcb.12235
Lü X, Reed SC, Yu Q, Han X-G (2016) Nutrient resorption helps drive intra-specific coupling of foliar nitrogen and phosphorus under nutrient-enriched conditions. Plant Soil 398:111–120. https://doi.org/10.1007/s11104-015-2642-y
Milla R, Castro-Diez P, Maestro-Martinez M, Montserrat-Marti G (2005) Does the gradualness of leaf shedding govern nutrient resorption from senescing leaves in Mediterranean woody plants? Plant Soil 278:303–313. https://doi.org/10.1007/s11104-005-8770-z
Moreau D, Bardgett RD, Finlay RD, Jones DL, Philippot L (2019) A plant perspective on nitrogen cycling in the rhizosphere. Funct Ecol 33:540–552. https://doi.org/10.1111/1365-2435.13303
Obermeier WA, Lehnert LW, Kammann CI, Müller C, Grünhage L, Luterbacher J, Erbs M, Moser G, Seibert R, Yuan N, Bendix J (2016) Reduced CO2 fertilization effect in temperate C3 grasslands under more extreme weather conditions. Nat Clim Chang 7:137–141. https://doi.org/10.1038/nclimate3191
Penuelas J, Sardans J, Rivas-Ubach A, Janssens IA (2012) The human-induced imbalance between C, N and P in Earth’s life system. Global Change Biol 18:3–6. https://doi.org/10.1111/j.1365-2486.2011.02568.x
Pereira CG, Clode PL, Oliveira RS, Lambers H (2018) Eudicots from severely phosphorus-impoverished environments preferentially allocate phosphorus to their mesophyll. New Phytol 218:959–973. https://doi.org/10.1111/nph.15043
Ren H, Xu Z, Huang J, Lu X, Zeng D-H, Yuan Z, Han X, Fang Y (2015) Increased precipitation induces a positive plant-soil feedback in a semi-arid grassland. Plant Soil 389:211–223. https://doi.org/10.1007/s11104-014-2349-5
Ren H, Kang J, Yuan Z, Xu Z, Han G (2018) Responses of nutrient resorption to warming and nitrogen fertilization in contrasting wet and dry years in a desert grassland. Plant Soil 432:65–73. https://doi.org/10.1007/s11104-018-3775-6
Sardans J, Alonso R, Janssens IA, Carnicer J, Vereseglou S, Rillig MC, Fernandez-Martinez M, Sanders TGM, Penuelas J (2016) Foliar and soil concentrations and stoichiometry of nitrogen and phosphorous across European Pinus sylvestris forests: relationships with climate, N deposition and tree growth. Funct Ecol 30:676–689. https://doi.org/10.1111/1365-2435.12541
Sattari SZ, Bouwman AF, Martinez Rodriguez R, Beusen AH, van Ittersum MK (2016) Negative global phosphorus budgets challenge sustainable intensification of grasslands. Nat Commun 7:10696. https://doi.org/10.1038/ncomms10696
Schuster MJ, Dukes JS (2017) Rainfall variability counteracts N addition by promoting invasive Lonicera maackii and extending phenology in prairie. Ecol Appl 27:1555–1563. https://doi.org/10.1002/eap.1547
Shi J, Gong J, Baoyin T, Luo Q, Zhai Z, Zhu C, Yang B, Wang B, Zhang Z, Li X (2021) Short-term phosphorus addition increases soil respiration by promoting gross ecosystem production and litter decomposition in a typical temperate grassland in northern China. CATENA 197:104952. https://doi.org/10.1016/j.catena.2020.104952
Song J, Wan S, Piao S, Knapp AK, Classen AT, Vicca S, Ciais P, Hovenden MJ, Leuzinger S, Beier C, Kardol P, Xia J, Liu Q, Ru J, Zhou Z, Luo Y, Guo D, Adam Langley J, Zscheischler J, Dukes JS, Tang J, Chen J, Hofmockel KS, Kueppers LM, Rustad L, Liu L, Smith MD, Templer PH, Quinn Thomas R, Norby RJ, Phillips RP, Niu S, Fatichi S, Wang Y, Shao P, Han H, Wang D, Lei L, Wang J, Li X, Zhang Q, Li X, Su F, Liu B, Yang F, Ma G, Li G, Liu Y, Liu Y, Yang Z, Zhang K, Miao Y, Hu M, Yan C, Zhang A, Zhong M, Hui Y, Li Y, Zheng M (2019) A meta-analysis of 1,119 manipulative experiments on terrestrial carbon-cycling responses to global change. Nat Ecol Evol 3:1309–1320. https://doi.org/10.1038/s41559-019-0958-3
Soong JL, Fuchslueger L, Maranon-Jimenez S, Torn MS, Janssens IA, Penuelas J, Richter A (2020) Microbial carbon limitation: the need for integrating microorganisms into our understanding of ecosystem carbon cycling. Global Change Biol 26:1953–1961. https://doi.org/10.1111/gcb.14962
Soudzilovskaia NA, Onipchenko VG, Cornelissen JHC, Aerts R (2007) Effects of fertilisation and irrigation on ‘foliar afterlife’ in alpine tundra. J Veg Sci 18:755–766. https://doi.org/10.1111/j.1654-1103.2007.tb02591.x
Su Y, Ma X, Gong Y, Li K, Han W, Liu X (2021) Responses and drivers of leaf nutrients and resorption to nitrogen enrichment across northern China’s grasslands: a meta-analysis. CATENA 199:105110. https://doi.org/10.1016/j.catena.2020.105110
Taylor CR, Janes-Bassett V, Phoenix GK, Keane B, Hartley IP, Davies JAC (2021) Organic phosphorus cycling may control grassland responses to nitrogen deposition: a long-term field manipulation and modelling study. Biogeosciences 18:4021–4037. https://doi.org/10.5194/bg-18-4021-2021
Tian Q, Lu P, Ma P, Zhou H, Yang M, Zhai X, Chen M, Wang H, Li W, Bai W, Lambers H, Zhang WH, Wurzburger N (2020) Processes at the soil–root interface determine the different responses of nutrient limitation and metal toxicity in forbs and grasses to nitrogen enrichment. J Ecol 109:927–938. https://doi.org/10.1111/1365-2745.13519
Tiruvaimozhi YV, Varma V, Sankaran M (2018) Nitrogen fixation ability explains leaf chemistry and arbuscular mycorrhizal responses to fertilization. Plant Ecol 219:391–401. https://doi.org/10.1007/s11258-018-0803-9
Tognetti PM, Prober SM, Baez S, Chaneton EJ, Firn J, Risch AC, Schuetz M, Simonsen AK, Yahdjian L, Borer ET, Seabloom EW, Arnillas CA, Bakker JD, Brown CS, Cadotte MW, Caldeira MC, Daleo P, Dwyer JM, Fay PA, Gherardi LA, Hagenah N, Hautier Y, Komatsu KJ, McCulley RL, Price JN, Standish RJ, Stevens CJ, Wragg PD, Sankaran M (2021) Negative effects of nitrogen override positive effects of phosphorus on grassland legumes worldwide. Proc Natl Acad Sci USA 118:e2023718118. https://doi.org/10.1073/pnas.2023718118
Van Dobben HF, Wamelink GWW, Slim PA, Kaminski J, Piorkowski H (2017) Species-rich grassland can persist under nitrogen-rich but phosphorus-limited conditions. Plant Soil 411:451–466. https://doi.org/10.1007/s11104-016-3021-z
Van Heerwaarden LM, Toet S, Aerts R (2003) Nitrogen and phosphorus resorption efficiency and proficiency in six sub-arctic bog species after 4 years of nitrogen fertilization. J Ecol 91:1060–1070. https://doi.org/10.1046/j.1365-2745.2003.00828.x
Widdig M, Heintz-Buschart A, Schleuss P-M, Guhr A, Borer ET, Seabloom EW, Spohn M (2020) Effects of nitrogen and phosphorus addition on microbial community composition and element cycling in a grassland soil. Soil Biol Biochem 151:108041. https://doi.org/10.1016/j.soilbio.2020.108041
Wright IJ, Cannon K (2001) Relationships between leaf lifespan and structural defences in a low-nutrient, sclerophyll flora. Funct Ecol 15:351–359. https://doi.org/10.1046/j.1365-2435.2001.00522.x
Yan Z, Li X, Tian D, Han W, Hou X, Shen H, Guo Y, Fang J (2018) Nutrient addition affects scaling relationship of leaf nitrogen to phosphorus in Arabidopsis thaliana. Funct Ecol 32:2689–2698. https://doi.org/10.1111/1365-2435.13219
Yan C, Yuan Z, Shi X, Lock TR, Kallenbach RL (2020) A global synthesis reveals more response sensitivity of soil carbon flux than pool to warming. J Soils Sed 20:1208–1221. https://doi.org/10.1007/s11368-019-02513-1
You C, Wu F, Yang W, Xu Z, Tan B, Yue K, Ni X (2018a) Nutrient-limited conditions determine the responses of foliar nitrogen and phosphorus stoichiometry to nitrogen addition: a global meta-analysis. Environ Pollut 241:740–749. https://doi.org/10.1016/j.envpol.2018.06.018
You C, Wu F, Yang W, Xu Z, Tan B, Zhang L, Yue K, Ni X, Li H, Chang C, Fu C (2018b) Does foliar nutrient resorption regulate the coupled relationship between nitrogen and phosphorus in plant leaves in response to nitrogen deposition? Sci Total Environ 645:733–742. https://doi.org/10.1016/j.scitotenv.2018.07.186
Yuan ZY, Chen HYH (2009) Global-scale patterns of nutrient resorption associated with latitude, temperature and precipitation. Global Ecol Biogeogr 18:11–18. https://doi.org/10.1111/j.1466-8238.2008.00425.x
Yuan ZY, Chen HYH (2012) A global analysis of fine root production as affected by soil nitrogen and phosphorus. Proc R Soc B-Biol Sci 279:3796–3802. https://doi.org/10.1098/rspb.2012.0955
Yuan ZY, Chen HYH (2015a) Decoupling of nitrogen and phosphorus in terrestrial plants associated with global changes. Nat Clim Chang 5:465–469. https://doi.org/10.1038/nclimate2549
Yuan ZY, Chen HYH (2015b) Negative effects of fertilization on plant nutrient resorption. Ecology 96:373–380. https://doi.org/10.1890/14-0140.1
Yuan ZY, Li LH, Han XG, Chen SP, Wang ZW, Chen QS, Bai WM (2006) Nitrogen response efficiency increased monotonically with decreasing soil resource availability: a case study from a semiarid grassland in northern China. Oecologia 148:564–572. https://doi.org/10.1007/s00442-006-0409-0
Zaehle S, Dalmonech D (2011) Carbon-nitrogen interactions on land at global scales: current understanding in modelling climate biosphere feedbacks. Curr Opin Env Sust 3:311–320. https://doi.org/10.1016/j.cosust.2011.08.008
Zhang H, Gao Y, Tasisa BY, Baskin JM, Baskin CC, Lu X-T, Zhou D (2019) Divergent responses to water and nitrogen addition of three perennial bunchgrass species from variously degraded typical steppe in Inner Mongolia. Sci Total Environ 647:1344–1350. https://doi.org/10.1016/j.scitotenv.2018.08.025
Zheng J, She W, Zhang Y, Bai Y, Qin S, Wu B (2018) Nitrogen enrichment alters nutrient resorption and exacerbates phosphorus limitation in the desert shrub Artemisia ordosica. Ecol Evol 8:9998–10007. https://doi.org/10.1002/ece3.4407
Zimmer A, Meneses RI, Rabatel A, Soruco A, Dangles O, Anthelme F (2018) Time lag between glacial retreat and upward migration alters tropical alpine communities. Perspect Plant Ecol 30:89–102. https://doi.org/10.1016/j.ppees.2017.05.003
Zong N, Chai X, Shi P, Yang X (2017) Effects of warming and nitrogen addition on plant photosynthate partitioning in an alpine meadow on the Tibetan Plateau. J Plant Growth Regul 37:803–812. https://doi.org/10.1007/s00344-017-9775-6
Zong N, Shi P, Chai X (2018) Effects of warming and nitrogen addition on nutrient resorption efficiency in an alpine meadow on the northern Tibetan Plateau. Soil Sci Plant Nutr 64:482–490. https://doi.org/10.1080/00380768.2018.1467727
Funding
This study was supported by the National Key Research and Development Program of China (2016YFA0600801), the Hundred Talents Program of Shaanxi Province (A289021701), the Natural Science Basic Research Plan in Shaanxi Province of China (2018JZ3002), the Special Fund from the State Key Laboratory of Soil Erosion and Dryland Farming on the Loess Plateau (A314021403-C9), and the Double First Class University Plan of NWSUAF (Z102021829). The funders had no role in the study design, data collection, data analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Hu, W., Yuan, Z., Shi, X. et al. A Global Meta-analysis Reveals that Nitrogen Addition Alters Plant Nutrient Concentration and Resorption in Grassland Ecosystems. J Soil Sci Plant Nutr 22, 4960–4971 (2022). https://doi.org/10.1007/s42729-022-00973-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42729-022-00973-y