
Abstract
Identification of “donors” possessing superior traits to adapt to low phosphorus (P) soils is a primary requirement to develop efficient crop plants for sustainable agricultural production. Diverse wheat (Triticum aestivum L., 89) and triticale (×Triticosecale Wittmack, 15) genotypes were phenotyped in soil with low and sufficient P for two seasons and once in the hydroponic system. Principal component analysis of relative biomass and grain weight revealed four clusters each for wheat and triticale. In low P soil, genotypes in the efficient cluster showed lesser reduction in total biomass, grain weight, total P uptake, and harvest index (HI), and improved P acquisition efficiency (PAE) and P harvest index (PHI) as compared to the inefficient cluster. In hydroponics, reduction in total biomass was less while root biomass and PAE increased at low P in the efficient genotypes. Out of 22 (soil) and 38 (hydroponics) P-efficient wheat genotypes, nine were overlapping in both screens while in P-inefficient cluster, out of 23 (soil) and 5 (hydroponics), three were common. A few P-efficient genotypes identified were DL784-3, DBW 16, and GW 322 and triticale TREAT, whereas P inefficient were C306 and ABACUS PVR (triticale). The selections in hydroponics are likely to be relevant in the field as demonstrated by overlapping of genotypes in both the mediums. Genotypic variations in P efficiency were strongly associated with P utilization rather than P uptake.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Phosphorus (P) is one of the macro-nutrients essential for crop growth and development, energy transfer reactions, and cellular metabolism. Plants acquire P as orthophosphate anions (HPO42− and H2PO4−) from the soil solution; however, its concentration is <5 μM in most soils (Conyers and Moody 2009). Bioavailability of P is a constraint in the agricultural production system due to its slow diffusion and high P-fixation in soils. Worldwide 50 million tons of P fertilizer per annum is required for crop cultivation (Lopez-Arrendondo et al. 2014). Because of the non-renewable nature of P reserves and extensive consumption, the cost of P fertilizer will continue to increase making the availability of P fertilizer worse in the future (Cordell et al. 2009). The practical and economical solution to P nutrition of crop plants can be achieved through genetic improvement of P efficiency rather than relying on chemical fertilization alone. Thus, the development of crop varieties that can efficiently acquire and utilize soil P would decrease the over-reliance on P fertilizers and, hence, reduce its impact on the ecosystem.
Phosphorus efficiency is defined as the ability of genotypes to grow and yield relatively better even under low P stress as compared to sufficient P conditions. Ideally, an efficient genotype should exhibit comparable yields under low P as well as an efficient response to P fertilization. P efficiency has been defined as relative shoot growth (Gunes et al. 2006; Ozturk et al. 2005) or relative grain yield (Graham 1984) suggesting that screening for P efficiency includes a direct comparison of shoot biomass and grain yield at low and sufficient P levels. Hence, an efficient genotype is the one that produces a high relative biomass and/or grain yield. Further, P efficiency mainly depends on two components, acquiring P from soils involving root traits and belowground processes (Elanchezhian et al. 2015; Lambers et al. 2006; Wang and Lambers 2019), and utilization of P which refers to mechanisms at cellular level involving Pi remobilization (Vance et al. 2003; Wang et al. 2010). For breeding P-efficient grain crops, mostly the P acquisition traits have been widely exploited (Wissuwa et al. 2009); however, there is a need to explore the traits governing P utilization efficiency.
Worldwide, wheat (Triticum aestivum L.) is grown as a staple food crop and consumes large (16.1%) amounts of phosphatic fertilizer which is greater than P consumption in other crops (Heffer 2013). Wheat is mainly cultivated in acidic soils of tropical and subtropical regions and the calcareous soils of temperate regions; hence, an important problem in wheat production is its lack of tolerance to low P stress (stress resulting from low P availability). Similarly, slow response to P fertilization and low P fertilizer recovery are important constraints to enhance P efficiency in wheat genotypes. Improvement of P efficiency in wheat through breeding is imperative for sustainable agricultural production (Godfray et al. 2010). For developing improved varieties, there is a need to identify genotypic variability for specific traits adapted to low P soils. Apart from wheat, high P uptake of triticale along with low P utilization provided better adaptation under low P environment (Osborne and Rengel 2002). Triticale (× Triticosecale Wittmack) is a synthetic self-pollinated crop derived from a cross between wheat (Triticum sp., AABB or AABBDD) and rye (Secale cereale, RR). One of the parents of triticale, rye, was reported to be an excellent performer in soils deficient in micronutrients (Cakmak et al. 1997), acidic conditions (Mergoum and Gómez-Macpherson 2004), Al toxicity (Slaski 1994), and low P stress (Pandey et al. 2005) due to its superior physiological mechanisms such as extensive root growth and root exudation.
In most studies, a limited number of wheat genotypes have been evaluated for genetic variation in P efficiency and tolerance to low P stress (Batten 1986; Bilal et al. 2018; Gunes et al. 2006; Jones et al. 1989; Korkmaz et al. 2009; Pandey et al. 2005; Wang et al. 2008). However, only a few studies have screened more than forty diverse wheat genotypes using soil as well as solution culture (Liao et al. 2008; Manske et al. 2000; McDonald et al. 2015; Osborne and Rengel 2002; Ozturk et al. 2005). The P efficiency mechanisms can vary between the genotypes of a given plant species. Hence, the primary requirement is to use a large number of diverse genotypes for screening to understand the low P tolerance mechanism related to P efficiency. In earlier reports, the screening medium used was either soil or solution culture (hydroponics). Both mediums have their drawbacks: in soil, a precise P concentration is difficult to achieve while in hydroponics, the roots do not have a mechanical support. Results obtained in soil have rarely been compared to hydroponics or vice-versa. In the present study, a diverse set of 89 bread wheat (T. aestivum) and 15 triticale genotypes were phenotyped to understand their response in terms of P efficiency (relative biomass and grain weight) using soil and hydroponics medium. We also demonstrated whether the selections in hydroponic screen would be relevant in the field and whether the growth medium affects the genetic differences in P efficiency.
2 Materials and Methods
2.1 Experiment 1: Phenotyping for P Efficiency in Soil
Eighty-nine wheat (Triticum aestivum) and 15 triticale (× Triticosecale) genotypes (Supplementary Table S1) were grown for two consecutive seasons (2016 and 2017) in sandy loam soil at low and sufficient P levels. Plants were sown in pots in the third week of November each year. Low P soil was collected from the site in our institute mapped as low P (https://www.iari.res.in/files/IARI_Soil_Fertility_Maps.pdf) and divided into two parts; in one-part, single super phosphate was added as per recommended dose (referred to as “sufficient P” treatment), while the other part was used as “low P.” Recommended doses of nitrogen and potassium through urea (116 mg N kg−1 soil) and muriate of potash (30 mg K kg−1 soil) respectively were added to both soils. After thorough mixing and sieving, soil analyses for various properties were carried out. The pH of soil in water (soil:water :: 1:2) was 7.1 and the electrical conductivity was 1.4 mS cm−1. The available P (Olsen et al. 1954) was 42.2 mg kg−1 soil in the sufficient P and 2.67 mg kg−1 soil in the low P treatment. Table 1 shows other soil properties including N, K, S, and DTPA extractable micronutrient (Fe, Mn, Zn, and Cu) contents. Earthen pots of size 30 cm diameter × 30 cm height were filled with low and sufficient P soil and kept in the net-house under natural condition at the Indian Agricultural Research Institute, New Delhi, India, located at 28.08° N and 77.12° E and 228.61 m above mean sea level. The mean maximum and minimum temperatures were 25.0 and 8.9 °C, respectively, in 2016 and 24.7 and 9.1 °C, respectively, in 2017. The total rainfall received during the crop season recorded was 19.2 mm and 81.9 mm in years 2016 and 2017, respectively. Six seeds were sown in each pot, and after emergence of 3–4 leaves, only four healthy plants per pot were retained.
Morpho-physiological observations on plants were recorded at the time of anthesis. Flag leaf area was measured with a leaf area meter (LI-3000; LI-COR, Lincoln, NE, USA) and expressed as cm2 plant−1. Relative amount of chlorophyll in flag leaf was measured by SPAD (SPAD-502 Meter; Konica-Minolta, Japan). Plant height, peduncle length, and number of productive tillers plant−1 were measured after attainment of physiological maturity. Total biomass (g plant−1), total grain weight (g plant−1), number of spikes plant−1, and 1000 grain weight (g) were recorded after harvest. Concentration of P in shoot and grain was estimated by ascorbic acid method (Murphy and Riley 1962) after wet-digestion with diacid mixture (HNO3 9 part: HClO4 4 part) (Bhargava and Raghupathi 1998). Total P content was calculated by multiplying tissue P concentration by total dry weight (shoot dry weight or grain weight) and expressed as mg P plant−1. P acquisition efficiency (PAE) was calculated as the ratio of P content at low P to sufficient P, and expressed as percentage (Lopez-Arrendondo et al. 2014). Relative biomass or grain yield of genotypes was calculated as the ratio of yield (total biomass or total grain yield) at low P to the yield at optimum P condition (Ozturk et al. 2005). Harvest index (HI) was calculated by dividing the grain weight with total biomass (sum of grain weight and shoot weight) while P harvest index (PHI) as the ratio of P content in grain to the P content in grain plus shoot (Jones et al. 1992). Both PHI and HI were expressed as percentage.
2.2 Experiment 2: Phenotyping in Hydroponics Media
Seeds were surface sterilized with 0.1% HgCl2, rolled in germination paper, and kept in the dark at 22 °C for germination. After emergence of coleoptiles (5 days after sowing), seedlings were transferred to Hoagland solution with sufficient (500 μM) and low (5 μM) P (Pandey et al. 2018). The plants were supported on Styrofoam sheets (5 cm thickness) and grown in plastic containers with 10 L of nutrient solution each. There were four replicates of each treatment. The solution was aerated continuously using aquarium pumps and PVC tubing. The solution was changed every third day, and pH (5.6–5.8) was maintained throughout the experiment using 1.0 N HCl or 1.0 N KOH. The composition of nutrient solution used was as follows: macronutrients (in mM) KNO3 5.0, Ca(NO3)2 1.5, NH4NO3 1.0, MgSO4 2.0, H3PO4 0.5; micronutrients (in μM) H3BO3 1.0, MnCl2·4H2O 0.5, ZnSO4·7H2O 1.0, CuSO4·5H2O 0.2, (NH4)6Mo7O24·4H2O 0.075, Fe-EDTA 10.0. Orthophosphoric acid was used to supply P. The experiment was conducted in a glasshouse at the National Phytotron Facility, ICAR-IARI, New Delhi, with day-night temperatures at 22 °C–12 °C, photoperiod of 10 h, and relative humidity maintained at 70%. Plants were sampled at 22 days after transfer to nutrient solution into root and shoot and were dried in hot air oven until a constant weight was obtained. Total P uptake (mg P plant−1) and PAE were estimated as described above.
2.3 Statistical Analyses
The experiment was conducted in a completely randomized design with two factors, P and genotypes. In soil culture, one replication unit consisted of three pots and each pot contained four plants while for hydroponics study, four replications were taken. Data analysis was carried out by two-way analysis of variance (ANOVA). Principal component analysis (PCA) and hierarchical cluster analysis were carried out using statistical software R version 3.5.1 (R Foundation for Statistical Computing, Vienna 2005). PCA was carried out using 16 quantitative traits (pooled mean of 2 years), and the parameters that differentiated genotypes for P efficiency were identified by the components of eigenvectors. Genotype and genotype × trait analysis was carried out in R using the package GGE Biplot GUI version 1.0-9. Clustering based on Ward’s method was executed using squared Euclidean distance matrix of genotypic relative biomass and relative grain yield for soil experiment while for hydroponics experiment, it was based on relative biomass and P acquisition efficiency. Graphs were plotted using GraphPad Prism version 5.01 (GraphPad Software, La Jolla, CA, USA).
3 Results
3.1 Evaluation of Wheat and Triticale Genotypes for P Efficiency in Soil Culture
Traits were recorded for two consecutive years and data was subjected to analysis of variance. Significant (P < 0.001) effects of genotype and P levels were observed for various traits in both years (Supplementary Table S2). The P × genotype interaction for triticale during 2016 was non-significant for the number of tillers, spikes plant−1, and grain P content while in 2017, P level showed no significant effect on 1000 grain weight both in wheat and triticale genotypes. Pooled mean (2 years) for traits like plant height, flag leaf area, number of tillers and spikes, total biomass and grain weight per plant, 1000 grain weight, shoot and grain P concentration and content, total P uptake and HI in low P soil in both triticale and wheat genotypes showed a significant reduction as compared to sufficient P (Table 2). However, a significant increase was noted in SPAD value, PHI, and peduncle length at low P both in wheat and triticale genotypes. The reduction in total biomass of wheat and triticale at low P was 20 and 24% respectively, and the reduction in grain weight was 24 and 30% respectively as compared to sufficient P treatment. Total P content decreased significantly by 53% and 58% in wheat and triticale respectively at low P in comparison to sufficient P. Likewise, the number of tillers was also reduced by 22% both in wheat and triticale genotypes at low P.
Growth responses of wheat and triticale genotypes at sufficient and low P levels were analyzed by principal component analysis (PCA) taking into consideration pooled mean for 2-year (2016 and 2017) data of 16 variables as mentioned in Table 2. Analysis of genotype and genotype × trait exhibited that PC1 and PC2 governed 51.2% variability at sufficient P and 52.8% at low P (Fig. 1a, b). At low P (Fig. 1b), the biplot revealed distinct groups and the most contributing traits to P efficiency were found to be total biomass, HI, PHI, total grain weight, 1000 grain weight, P content in grain and shoot and total plant P uptake. Traits which explained genotypic variability at sufficient P in PC1 were PHI (18.0%), P percentage in shoot (15.4%), shoot P content (11.7%), HI (11.7%), and total grain weight (10.0%). Similarly, under low P, total plant P uptake (17.0%), total grain weight (16.1%), P content in grain (15.7%), total biomass (12.7%), and 1000 grain weight (11.6%) contributed to genotypic variability. Within PC2, the maximum genotypic variation was contributed by total biomass (16.7%) at sufficient P and PHI (19.1%) at low P. Similarly, in PC2, the total plant P uptake contributed to genotypic variability by 13.4% at sufficient P. Other traits which explained genotypic variability at low P in PC2 were HI (16.0%) and number of tillers (13.4%). Since the analysis of biplots at sufficient and low P revealed that grain weight and biomass governed maximum genetic variability, cluster analysis was done using relative values for grain weight and biomass (Fig. 2a, b).

Genotype and genotype × trait interaction biplot at (a) sufficient (42.2 mg P kg−1 soil), and (b) low (2.67 mg P kg−1 soil) phosphorus. Biplot analysis identified 51.15% of variability at sufficient P and 52.80% of variability at low P. Serial number corresponds to genotype ID in Supplementary Table 1S. PH, plant height; PL, peduncle length; TBM, total biomass; TGW, total grain weight; 1000GW, 1000 grain weight; PCONTS, P content in shoot; PCONTG, P content in grain; SPIKE, number of spikes plant−1; TILLER, number of tillers plant−1; PPERS, P percentage in shoot; PPERG, P percentage in grain; PCONT, total P content; PAE, P acquisition efficiency; SPAD, SPAD value; HI, harvest index; PHI, P harvest index; FLA, flag leaf area

Clustering of (a) wheat (Triticum aestivum), and (b) triticale genotypes by Ward’s method using squared Euclidean distance matrix of relative biomass and relative grain weight. Plants were grown in soil with sufficient (42.2 mg P kg−1 soil) and low (2.67 mg P kg−1 soil) phosphorus
Clustering of genotypes resulted in four distinct groups (Fig. 2a) in wheat as efficient (average relative biomass 88.5 and grain weight 86.0), moderately efficient (average relative biomass 87.3 and grain weight 75.2), moderately inefficient (average relative biomass 76.0 and grain weight 79.7), and inefficient (average relative biomass 71.2 and grain weight 64.2). Out of 89 wheat genotypes, 22 were included in the efficient cluster, 18 were classified as moderately efficient, 26 belonged to moderately inefficient, and 23 were classified as inefficient. A significant difference was found among these groups with respect to total biomass, grain weight, P content in shoot and grain, total P content, HI, PHI, and PAE. In genotypes belonging to efficient cluster, the reduction in total biomass was 11.7% whereas, in moderately efficient cluster, it was 12.6% at low P. Compared to efficient and moderately efficient clusters, low P significantly reduced total biomass of genotypes classified under moderately inefficient (24%) and inefficient (29%) clusters (Fig. 3). Similarly, the efficient, moderately efficient, moderately inefficient, and inefficient clusters showed a reduction in grain weight at low P as compared to sufficient P by 14, 21, 25, and 36% respectively. A significant reduction in shoot and grain P content was noted at low P in comparison to sufficient P in all four clusters, with the highest reduction of 69.7% (shoot P content) and 46.8% (grain P content) in the inefficient cluster. Low P treatment significantly influenced total plant P uptake in all clusters with maximum reduction (60.6%) in inefficient cluster whereas efficient cluster exhibited 47.4% reduction as compared to sufficient P. The HI and PHI did not vary consistently among the groups at either P levels; however, the latter was increased under low P as compared to sufficient P and the contrasting clusters exhibited more than 20% increase. The PAE also varied distinctly among different clusters; it was 54.9, 51, 49.5, and 41.8% in the efficient, moderately efficient, moderately inefficient, and the inefficient cluster respectively. Among other traits, decreases in 1000 grain weight and spike numbers were less in the efficient cluster at low P as compared to sufficient P (Supplementary Fig. S1).

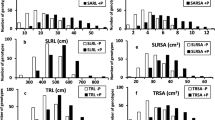
Box and whisker plot showing difference in wheat (Triticum aestivum) genotypes with respect to total biomass, number of tillers plant−1, total grain weight, shoot P content, grain P content, total plant P content, harvest index, P harvest index, and P acquisition efficiency screened in soil with sufficient (42.2 mg P kg−1 soil) and low (2.67 mg P kg−1 soil) P. Horizontal lines inside boxes, median; +, mean; box hinges, first and third quartiles; whiskers, full range of the data; black dots represent outliers. Clustering of genotypes was performed based on Ward’s method using squared Euclidean distance matrix of genotypic relative biomass and relative grain yield
Similar to wheat, triticale genotypes were also grouped into four clusters (Fig. 2b), efficient (average relative biomass 88.4 and grain weight 85.1), moderately efficient (average relative biomass 82.4 and grain weight 69.7), moderately inefficient (average relative biomass 64.8 and grain weight 71.5), and inefficient (average relative biomass 66.7 and grain weight 58.9). Out of 15 triticale genotypes, three belonged to efficient, five in moderately efficient, two as moderately inefficient, and five were in the inefficient cluster. Significant variation was observed among these clusters with respect to total biomass, grain weight, P content in shoot and grain, total plant P uptake, HI, PHI, and PAE (Supplementary Table S2). Low P treatment significantly reduced total biomass in all clusters with maximum reduction observed in inefficient (33.2%) whereas it was only 11% in the efficient cluster (Fig. 4). Likewise, reduction in total grain weight was 15% in the efficient cluster as against 41% in the inefficient cluster. However, in moderately efficient and moderately inefficient clusters, a 30% reduction in grain weight was observed at low P. The total plant P uptake at low P decreased by 52.5, 56.7, 51, and 65.6% in the efficient, moderately efficient, moderately inefficient, and inefficient clusters, respectively. Phosphorus content in shoot and grain decreased significantly at low P compared to sufficient P in all clusters, with maximum reduction (73.2% in shoot P and 55.3% in grain P) observed in the inefficient cluster. Likewise, significant reduction in HI was noted in the inefficient cluster at low P. However, the PHI was enhanced by 26% in the inefficient as against 45.7% for the efficient cluster at low P. The PAE was enhanced by 47.7% in the efficient cluster and 37.5% in the inefficient cluster. Besides these traits, the reduction in plant height observed at low P was less whereas no reduction was observed in the efficient cluster as against inefficient cluster which showed significant decrease in both traits (Supplementary Fig. S2).

Difference in triticale (× Triticosecale) genotypes with respect total biomass, number of tillers plant−1, total grain weight, shoot P content, grain P content, total plant P content, harvest index, P harvest index, and P acquisition efficiency screened in soil with sufficient (42.2 mg P kg−1 soil) and low (2.67 mg P kg−1 soil) P. Clustering of genotypes was performed based on Ward’s method using squared Euclidean distance matrix of genotypic relative biomass and relative grain yield. C I, efficient; C II, moderately efficient; C III, moderately inefficient; C IV, inefficient
3.2 Evaluation of Wheat and Triticale Genotypes for P Efficiency in Hydroponics Culture
The same set of wheat and triticale genotypes were phenotyped in hydroponics solution at seedling stage with precise P concentration. The genotype and P levels significantly (P < 0.001) influenced total biomass, root and shoot dry weight, root-to-shoot ratio, and total P content plant−1 both in wheat and triticale genotypes (Table 3). Total biomass, shoot dry weight, and total P content decreased significantly, but root dry weight and root-to-shoot ratio increased at low P both in wheat and triticale genotypes as compared to sufficient P concentration. Total P uptake was reduced by more than 70% at low P concentration in wheat and triticale genotypes in comparison to sufficient P. The root-to-shoot ratio was increased at low P in both cereals, and it was proportionately more in triticale, inferring that this change contributes to better efficiency in triticale.
Clustering based on relative biomass and PAE revealed four groups each of wheat and triticale genotypes (Fig. 5a, b). In wheat genotypes, the four clusters were as efficient (average values for relative biomass 95.7 and PAE 32.5), moderately efficient (average values for relative biomass 85.7 and PAE 24.9), moderately inefficient (average values for relative biomass 72.1 and PAE 23.0), and inefficient (average values for relative biomass 56.0 and PAE 17.6). Out of 89 wheat genotypes, 38 belonged to the efficient cluster, 28 in moderately efficient, 18 in moderately inefficient, and five were included in the inefficient cluster (Fig. 5a). Significant variation was observed among these clusters with respect to total biomass, total P content, shoot and root dry weight, and root-to-shoot ratio (Fig. 6). The reduction in total biomass at low P treatment in the efficient cluster was non-significant compared to sufficient P, but a significant decrease was observed in moderately efficient (15.0%), moderately inefficient (28.1%), and inefficient clusters (43.6%). Similarly, low P treatment resulted in a marked reduction in total P content which was 83.3% in the inefficient cluster against 68% in the efficient cluster. Root dry weight increased by 34.5% at low P compared to sufficient P in the efficient cluster while genotypes in other clusters did not show any significant increase.

Clustering of (a) wheat (Triticum aestivum), and (b) triticale genotypes by Ward’s method using squared Euclidean distance matrix of genotypic relative biomass and P acquisition efficiency. The plants were grown in hydroponics at sufficient (500 μM) and low (5 μM) phosphorus concentrations for 22 days

Box and whisker plot showing difference in wheat (Triticum aestivum) genotypes with respect to total biomass, shoot dry weight, root dry weight, root-to-shoot ratio, total P content, and P acquisition efficiency at sufficient (500 μM) and low (5 μM) phosphorus (P). Horizontal lines inside boxes, median; +, mean; box hinges, first and third quartiles; whiskers, full range of the data; black dot represents outliers. Genotypes were classified on the basis of relative biomass and P acquisition efficiency
Clustering of triticale genotypes also revealed four groups (Fig. 5b) as efficient (average relative values for biomass 102.1 and PAE 35.1), moderately efficient (average relative values for biomass 97.3 and PAE 32.3), moderately inefficient (average relative values for biomass 93.2 and PAE 30.5), and inefficient (average relative values for biomass 77.4 and PAE 24.6). Out of 15 triticale genotypes, two were classified as efficient, one as moderately efficient, seven as moderately inefficient, and five as inefficient. In the efficient cluster, total biomass was not influenced by low P while in the inefficient cluster, 22.3% reduction was observed (Fig. 7). The total P content was reduced significantly at low P compared to sufficient P in all clusters with 75.8% reduction observed in the inefficient cluster. Root dry weight increased by 89.8% while shoot dry weight exhibited no significant effect of low P concentration in the efficient cluster.

Difference in triticale (× Triticosecale) genotypes with respect to total biomass, shoot biomass, root biomass, root-to-shoot ratio, total P content, and P acquisition efficiency at sufficient (500 μM) and low (5 μM) phosphorus (P). Genotypes were classified on the basis of relative biomass and P acquisition efficiency in four clusters, CI, efficient; CII, moderately efficient; CIII, moderately inefficient; CIV, inefficient
3.3 Identification of Low P Stress Tolerant and Sensitive Wheat and Triticale Genotypes
From cluster analysis, common genotypes in both cereal species that responded consistently in soil and hydroponic experiments with respect to low P stress tolerance, were identified (Table 4). For cluster analysis, relative biomass was the common trait in both screening mediums. In wheat, the proportion of lines that overlapped in the efficient cluster in soil screen was 40.9% while in hydroponics, it was 23.7%. In both screens, nine genotypes DBW 39, DBW 16, DL 784-3, HD 2967, RAJ 3077, BWL 5202, EC-556434, RAJ 3765, and MACS 6222 were efficient. These common genotypes showed percent reduction in biomass between < 4 and 20% in soil whereas in hydroponics, it was zero to 9% only. Besides total biomass, other contributing traits for these genotypes were total grain weight, HI, total P content, P content in grain, 1000 grain weight, and PAE as evident from Fig. 1b. Similarly, in the moderately efficient cluster, the overlapping genotypes in soil and hydroponics were 16.7 and 10.7% respectively, which were SUNCO, SUNMIST, and STILLETO. In the moderately inefficient cluster, the proportion of common genotypes in soil and hydroponic screen was 30.8 and 44.4% respectively which included DBW 88, HD 2687, BWL 5200, ROLLER-1, NI-5439, OLYMPIC, DATATINE, and KYPO-328. The inefficient cluster exhibited 13.0 and 60.0% of overlapping genotypes in soil and hydroponic screens respectively and those were C 306, CULICANT 89, and ARRINO. These genotypes in the inefficient cluster exhibited poor performance at low P. There was significant reduction in biomass, grain weight, and total P uptake while PAE was < 50% in soil culture. Similar poor performance was exhibited by these genotypes for total biomass, shoot biomass, and total P content in hydroponics culture.
There were a few genotypes which changed the category from efficient in one screen to inefficient in another screen. For example, PBW 502, PBW 550, UP 2425, PBW 677, PBW 660, PBW 644, PBW 396, EC 463394, EC 574376, and SAMNGP 408 were grouped under efficient cluster in hydroponics fell into inefficient cluster in soil screen. In hydroponics screening, these genotypes exhibited < 12% reduction in total biomass and 24 to 184% increase in root biomass but in low P soil, a significant reduction in biomass (21 to 38%) and grain yield (28 to 55%) was observed. Similarly, HD 3090, ATTILA, GUTHA, PBW 175, and RAJ 3777 which were grouped in the efficient cluster in soil were categorized into moderately inefficient cluster in hydroponics. Eight genotypes (VEE/MYNA, CARAZINHO, BENCUBBIN, HD 2891, WH 1105, GW 322, WARIGAL, and RAJ 3777) that were efficient in soil moved to moderately efficient cluster in hydroponics screening. These genotypes exhibited 4 to 18% reduction in biomass with PAE > 45% in low P soil as compared to sufficient P whereas in hydroponics the reduction in biomass was up to 21% with PAE < 30%.
Among triticale genotypes, there was no overlapping between soil and hydroponics in the efficient cluster; however, SPL-TCL-8 and TREAT which were in the efficient and moderately efficient clusters respectively in soil fall under moderately efficient cluster in hydroponics (Table 4). Triticale TREAT exhibited a reduction in total biomass (23%), grain weight (29%), and total P content (60%) in low P soil whereas PAE was > 45%. In hydroponic culture, TREAT showed non-significant change in total biomass, while a reduction in total P content (68%) and low PAE (32%) were observed. The proportion of genotypes overlapped in both soil and hydroponic experiments under inefficient cluster was 60% which included T-3866, ABACUS PVR, and SPL-TCL-4. Among these, ABACUS PVR showed a maximum reduction in total biomass (37%), grain weight (36%), and total P content (67%) while PAE was < 45%. In hydroponics, ABACUS PVR exhibited a significant reduction in total biomass (22%), P content (85%), and PAE (15%). Likewise, the proportion of overlapping in moderately efficient cluster was 20 and 100% in soil and hydroponics respectively whereas in the moderately inefficient cluster, it was 100 and 28.6% in soil and hydroponics respectively (Table 4). It was noted that T-3861 in hydroponic screen changed its group from efficient to inefficient in soil screen, as in former medium, it produced relatively higher biomass with no reduction in shoot dry weight. However, in low P soil medium, it could not sustain growth till maturity and, therefore, reduction in total biomass (31%) and grain yield (40%) as well as P uptake (62%) was higher. Similarly, the genotypes T-3849 and TL-2942 in the efficient group in soil changed to the inefficient in hydroponics screen. These two genotypes in soil showed lesser reduction in relative biomass and grain yield with > 50% decrease in P uptake while in hydroponics, significant reduction in shoot biomass and P uptake (> 75%) was observed.
4 Discussion
The present study showed that more than 16% of genetic variability to P nutrition was explained by total biomass and grain weight in soil experiment. Under low P, a small reduction in total biomass was observed both in wheat and triticale genotypes belonging to the efficient cluster in comparison to inefficient cluster. Our result corroborates with earlier reports where accumulation of biomass in P-efficient genotypes was higher than that in P-inefficient genotypes of maize (Zea mays) (Fageria and Baligar 1997a), rice (Oryza sativa) (Fageria et al. 1988; Fageria and Baligar 1997b), wheat (Osborne and Rengel 2002; Ozturk et al. 2005), and soybean (Glycine max) (Krishnapriya and Pandey 2016). Low P concentration significantly reduced total biomass in wheat genotypes belonging to the inefficient cluster. Ahmad et al. (2001) reported that a small difference in total biomass between sufficient P and low P conditions in cotton genotypes would be suited for cultivation in P-limited areas. So, total dry matter production can be used as a criterion for screening genotypes of wheat for P efficiency (Fageria and Baligar 1999; Osborne and Rengel 2002). Genotypes with higher relative biomass produced higher dry matter under low P which demonstrates that it is a superior trait for low P stress screening as reported in other crops such as soybean (Furlani et al. 2002), pigeon pea (Cicer arietinum) (Vesterager et al. 2006), dry bean (Phaseolus vulgaris) (Fageria et al. 2010), and cotton (Gossypium hirsutum) (Ahmad et al. 2001).
The total grain weights both in wheat and triticale genotypes were reduced under low P, but a lesser reduction was observed in the efficient cluster, while it was significantly reduced in the inefficient cluster both in wheat and triticale genotypes under low P. Horst et al. (1993) attributed reduction in grain yield to inhibition of tillering causing lack of assimilate production under low P. But in the present study, differences in percent reduction in tiller number in the efficient (18%) and inefficient (20%) clusters were similar. The traits governing grain yield in the efficient genotypes at low P may be attributed to 1000 grain weight, PAE, grain P content, and total plant P uptake (Fig. 1b). Similarly, a less relative reduction in grain yield under low P stress in P-efficient Brassica oleracea genotypes was reported (Hammond et al. 2009). Other reports on P-efficient genotypes of groundnut (Arachis hypogea) showed high pod yield and P uptake at low P (Singh and Basu 2005). Wide variation in sink strength was observed between efficient and inefficient groundnut genotypes. Enhanced sink strength (seed size) was reported due to low P stress tolerance in legumes (Liao and Yan 1999). Likewise, common bean genotypes producing relatively higher dry matter at low P soil (efficient) and also responded well to added P (responsive) were most desirable to yield well under P-limited conditions (Fageria and da Costa 2000).
In the present experiment, P content in shoot and grain reduced significantly at low P as compared to sufficient P in all four clusters but the reduction was higher in the inefficient cluster in both cereal genotypes. Low P treatment also significantly influenced total plant P uptake in all groups with maximum reduction in the inefficient cluster whereas the efficient cluster showed only a smaller reduction in total P uptake in comparison to sufficient P. The P content is dependent on biomass accumulation indicating that the P utilization efficiency decreased in the inefficient genotypes at low P (McDonald et al. 2015). Similar lesser reduction in total biomass exhibited by efficient soybean genotypes grown in low P soil was due to maintaining high P content as compared to the inefficient genotypes (Krishnapriya and Pandey 2016). Ozturk et al. (2005) observed that the total biomass and total P uptake at low P levels were the most reliable traits for assessing wheat genotypes for P efficiency. However, many of the wheat and triticale genotypes with a high P efficiency ratio (relative biomass) exhibited low P content in shoots due to differences in P utilization between P-efficient and P-inefficient genotypes (Rengel 1999). Reports in wheat showed that the P uptake and root dry weight of the efficient genotypes were superior to the inefficient genotypes (Fageria and Baligar 1999). Higher grain yield in wheat was observed as a result of large total P uptake (Jones et al. 1992) which is possibly due to the efficient root system architecture at low P (Gahoonia and Nielsen 2004).
In general, a reduction in HI at low P as compared to sufficient P both in wheat and triticale genotypes was observed. But very less variation was seen in HI among the efficient and inefficient clusters in both groups. Sandana and Pinochet (2011) also observed only slight variation in HI between different P treatments and showed no direct response to P-limited condition. Grain yield response to P supply in wheat was not related to HI (Manske et al. 2001) which is also reflected in our results. Hence, partitioning of photosynthates towards grain filling showed little contribution to grain yield under low P. Aulakh et al. (2003) reported that grain yield was highly correlated with total biomass rather than HI in soybean and wheat genotypes. On the contrary, PHI was found to be enhanced under low P as compared to sufficient P both in wheat and triticale. PHI is a measure of the partitioning of P in crop plants that indicates how efficiently the plant had utilized the acquired P for grain production. It measures the partitioning of P to the grain in relation to total P uptake of the plant, and so the PHI was negatively correlated with shoot P content but positively with grain P content in both cereal species irrespective of soil P concentration. High PHI value was observed for the inefficient wheat and triticale genotypes while low PHI value was exhibited by the efficient genotypes. Our result corroborates with the previous report where low PHI value was observed by efficient wheat genotypes due to its high yielding potential which reduced the grain P concentration as an effect of dilution (Gill et al. 2004).
The PAE of genotypes belonging to the efficient cluster was higher than that of the inefficient cluster in wheat and triticale at low P in both soil and hydroponic media. Generally, plant P uptake under low P stress is the sum of physiological and biochemical adaptations. Physiological adaptations include changes in root system architecture which mainly influences acquisition efficiency (Krishnapriya and Pandey 2016; Lambers et al. 2006; Raghothama and Karthikeyan 2005). The adaptive mechanism under low P stress includes a higher ratio of root to shoot growth, increased root length, and enhanced number of root hairs in wheat (Horst et al. 1993) besides the physiological processes involved in P mining (Krishnapriya and Pandey 2016; Lambers et al. 2013; Pandey et al. 2014; Pandey et al. 2018; Wang et al. 2008). In P-efficient maize genotypes, higher root growth and shoot P content were obtained than P-inefficient genotypes (de Sousa et al. 2012). Our results are in agreement with that of P-efficient rapeseed genotypes which exhibited higher biomass under low P condition due to increased P acquisition (Akhtar et al. 2008; Hu et al. 2010). Similarly, higher yield in P-efficient wheat genotypes was correlated to an increase in PAE and biomass (Fageria and Baligar 1999; Gill et al. 2004). There is a need to study the root system and other P mining traits in these P-efficient wheat and triticale genotypes.
In hydroponics, low P concentration significantly reduced total biomass of genotypes in the moderately inefficient and inefficient clusters as compared to efficient and moderately efficient clusters. Similar results were exhibited by P-efficient soybean genotypes which produced higher biomass compared to P inefficient under low P condition in hydroponics (Krishnapriya and Pandey 2016; Zhou et al. 2016). Low P stress significantly reduced total P uptake in all groups with a maximum reduction in the inefficient cluster. The root dry weight was highest in the efficient cluster in comparison to the inefficient cluster. Our results are in agreement with earlier reports in maize (Zhu et al. 2010) and common bean plants (Ciereszko et al. 1996) where low P stress typically showed enhanced biomass allocation to roots and altered the root system due to increased root length, surface area, and volume. The enhanced biomass partitioning to root under low P concentration causes a reduction in total biomass per plant due to greater respiratory burden of root cells (Hansen et al. 1998). But P-efficient rice plants showed the ability of preferential distribution of P to roots, which enhanced root growth and P uptake, thus improving tolerance to low P stress (Wissuwa et al. 2005).
Our work consisted of a number of common genotypes that have been assessed for P efficiency in earlier experiments (Liao et al. 2008; McDonald et al. 2015; Osborne and Rengel 2002). The genotypes ranked as efficient by Osborne and Rengel (2002) based on shoot dry weight at deficient P supply were CARAZINHO, EGRET, SILVERSTAR, STRETTON, and GUTHA. Out of these, CARAZINHO and GUTHA performed consistently well in terms of relative biomass and grain yield and grouped as P efficient in our study as well as by McDonald et al. (2015). Genotypes EGRET and SILVERSTAR were moderately efficient whereas STRETTON was inefficient in the present study. Another set ranked as moderate to P efficient (SUNMIST, STILLETO, SUNSTATE, BLADE, WARIGAL, DATATINE, ARRINO, CALINGIRI, TAMMIN, and CARNAMAH) by Osborne and Rengel (2002) were comparable with our study. We found SUNMIST, STILLETO, and SUNSTATE grouped as moderately efficient while DATATINE, TAMMIN, CARNAMAH, and BLADE as moderately inefficient. Further, WARIGAL fell in the efficient cluster whereas BROOKTON, ARRINO, and CALINGIRI were grouped as P inefficient. Among genotypes BANKS, JANZ, SUNCO, and BT SCHOMBURGK categorized as inefficient by Osborne and Rengel (2002), we found only JANZ in the inefficient cluster while BANKS and BT SCHOMBURGK were moderately inefficient, and SUNCO as moderately efficient in soil experiment. Interestingly, BROOKTON and JANZ, common in our study and others (Liao et al. 2008; McDonald et al. 2015; Osborne and Rengel 2002), performed consistently poor in biomass production at low P irrespective of growth media. Apart from wheat, triticale TREAT performed well in terms of shoot dry weight at deficient P supply and was grouped as efficient by Osborne and Rengel (2002) which also agrees with our result. However, they grouped triticale ABACUS PVR as moderately P efficient but, in present study, it performed poorly in both mediums. Thus, it provides a strong evidence that a few genotypes performing consistently across the continents should be included in the breeding program for developing P-efficient wheat cultivars.
Earlier reports found no correlation between field and hydroponics screening for low P stress tolerance (Hayes et al. 2004; Rose et al. 2011, 2015), but it was shown that most of the soil “efficient” genotypes were also ranked either efficient or moderately efficient in hydroponic conditions. Meena et al. (2021) evaluated 100 diverse mungbean accessions for low P tolerance in hydroponics, and the selected ten accessions were further validated in soil. Significant relationship between the two methods was observed, and out of ten, eight mungbean accessions stayed in their efficient/inefficient category when evaluated in soil. Panigrahy et al. (2014) showed comparable results in rice (Nagina 22 mutants) under low P conditions in field and nutrient solution experiments. They suggested that traits, viz., root biomass, root length, and root/shoot fresh weight, in response to P deficiency in hydroponically grown seedlings can be used as indicators for identification of P-efficient genotypes under low P soil. Our study showed that in hydroponics, the reduction in biomass under low P stress was higher as compared to soil screen. It may be argued that the P uptake from soil is limited by the rate of P diffusion to root surface, and therefore roots explore greater soil volume whereas in hydroponics, P moves to root surface via mass flow without any hindrance. Moreover, the root exudates and microbial fauna in soil grown plants help to acquire non-available P but such interaction is absent in nutrient solution culture (McNear 2013). Moreover, there is no scope for roots to improve P acquisition in hydroponics since the low P concentration is fixed. All these factors result in faster decrease in plant biomass under limited P supply in solution culture. Despite these differences in P uptake mechanisms, the inherent genetic potential of a variety at seedling stage in hydroponics, and till maturity in soil screen, cannot be ignored.
5 Conclusions
The present study suggests that the selection of genotypes in hydroponics was expected to be relevant in the field, or at least, would not lead to inappropriate selections and rejections as we found some overlapping of genotypes within the contrasting groups in both the mediums. The results showed that seedling traits, viz., root biomass, total biomass, and phosphorus acquisition efficiency, of hydroponically grown plants in response to low P may be used as component traits which are controlled genetically and would be effective in plants grown till harvest in low P soil. Further, it was observed that the P efficiency differences among the genotypes were strongly governed by P utilization rather than uptake as evident from the differences obtained in relative biomass (P efficiency ratio) between the efficient and inefficient genotypes. The enhanced P harvest index under low P also indicated that the acquired P was efficiently utilized for grain production. The genotypes identified in this study may be utilized as “donors” by the breeders to develop low P stress tolerant cultivars in wheat and triticale.
Abbreviations
- P:
-
Phosphorus
- PAE:
-
Phosphorus acquisition efficiency
- HI:
-
Harvest index
- PHI:
-
Phosphorus harvest index
- PCA:
-
Principal component analysis
References
Ahmad Z, Ahmad Z, Gill MA, Qureshi RH, Hamud-Ur-Rehman MT (2001) Phosphorus nutrition of cotton cultivars under deficient and adequate levels in solution culture. Commun Soil Sci Plant Anal 32:171–187. https://doi.org/10.1081/CSS-100103001
Akhtar MS, Oki Y, Adachi T (2008) Genetic diversity of Brassica cultivars in relation to phosphorus uptake and utilization efficiency under P-stress environment. Arch Agron Soil Sci 54:93–108. https://doi.org/10.1080/03650340701663574
Aulakh MS, Pasricha NS, Bahl GS (2003) Phosphorus fertilizer response in an irrigated soybean-wheat production system on a subtropical, semiarid soil. Field Crops Res 80:99–109. https://doi.org/10.1016/S0378-4290(02)00172-7
Batten GD (1986) Phosphorus fractions in the grain of diploid, tetraploid, and hexaploid wheat grown with contrasting phosphorus supplies. Cereal Chem 63:384–387
Bhargava BS, Raghupathi HB (1998) Analysis of plant materials for macro and micronutrients. In: Tandon HLS (ed) Methods of analysis of soils, plants. Waters and Fertilisers. Fertilizer Development and Consultation Organisation, New Delhi, pp 58–59
Bilal HM, Aziz T, Maqsood MA, Farooq M, Yan G (2018) Categorization of wheat genotypes for phosphorus efficiency. PLoS One 13:e0205471. https://doi.org/10.1371/journal.pone.0205471
Cakmak I, Ekiz H, Yilmaz A, Torun B, Koleli N, Gultekin I, Alkan A, Eker S (1997) Different response of rye, triticale, bread and durum wheats to zinc efficiency in calcareous soils. Plant Soil 188:1–10. https://doi.org/10.1023/A:1004247911381
Ciereszko I, Gniazdowska A, Mikulska M, Rychter AM (1996) Assimilate translocation in bean plants (Phaseolus vulgaris L.) during phosphate deficiency. J Plant Physiol 149:343–348. https://doi.org/10.1016/S0176-1617(96)80132-5
Conyers MK, Moody PW (2009) A conceptual framework for improving the P efficiency of organic farming without inputs of soluble P fertiliser. Crop Pasture Sci 60:100–104. https://doi.org/10.1071/CP06327
Cordell D, Drangert JO, White S (2009) The story of phosphorus: global food security and food for thought. Global Environ Chang 19:292–305. https://doi.org/10.1016/2008.10.009
de Sousa SM, Clark RT, Mendes FF, de Oliveira AC, de Vasconcelos MJV, Parentoni SN, Kochian LV, Guimaraes CT, Magalhaes JV (2012) A role for root morphology and related candidate genes in P acquisition efficiency in maize. Funct Plant Biol 39:925–935. https://doi.org/10.1071/FP12022
Elanchezhian R, Krishnapriya V, Pandey R, Rao AS, Abrol YP (2015) Physiological and molecular approaches for improving phosphorus uptake efficiency of crops. Curr Sci 108:1271–1279
Fageria NK, Baligar VC (1997a) Phosphorus-use efficiency by corn genotypes. J Plant Nutr 20:1267–1277. https://doi.org/10.1080/01904169709365334
Fageria NK, Baligar VC (1997b) Upland rice genotypes evaluation for phosphorus use efficiency. J Plant Nutr 20:499–509. https://doi.org/10.1080/01904169709365270
Fageria NK, Baligar VC (1999) Phosphorus-use efficiency in wheat genotypes. J Plant Nutr 22:331–340. https://doi.org/10.1080/01904169909365630
Fageria NK, da Costa JGC (2000) Evaluation of common bean genotypes for phosphorus use efficiency. J Plant Nutr 23:1145–1152. https://doi.org/10.1080/01904160009382088
Fageria NK, Wright RJ, Baligar VC (1988) Rice cultivar evaluation for phosphorus use efficiency. Plant Soil 111:105–109. https://doi.org/10.1007/BF02182043
Fageria NK, Baligar VC, Moreira A, Portes TA (2010) Dry bean genotypes evaluation for growth, yield components and phosphorus use efficiency. J Plant Nutr 33:2167–2181. https://doi.org/10.1080/01904167.2010.519089
Furlani AMC, Furlani PR, Tanaka RT, Mascarenhas HAA, Delgado MDP (2002) Variability of soybean germplasm in relation to phosphorus uptake and use efficiency. Sci Agric 59:529–536. https://doi.org/10.1590/S0103-90162002000300018
Gahoonia TS, Nielsen NE (2004) Root traits as tools for creating phosphorus efficient crop varieties. Plant Soil 260:47–57. https://doi.org/10.1023/B:PLSO.0000030168.53340.bc
Gill HS, Singh A, Sethi SK, Behl RK (2004) Phosphorus uptake and use efficiency in different varieties of bread wheat (Triticum aestivum). Arch Agron Soil Sci 50:563–572. https://doi.org/10.1080/03650340410001729708
Godfray HCJ, Beddington JR, Crute IR, Haddad L, Lawrence D, Muir JF, Pretty J, Robinson S, Thomas SM, Toulmin C (2010) Food security: the challenge of feeding 9 billion people. Science 327:812–818. https://doi.org/10.1126/science.1185383
Graham RD (1984) Breeding for nutritional characteristics in cereals. In: Tinker PB, Lauchli A (eds) Advances in plant nutrition. Preagier Publishers, New York, pp 57–102
Gunes A, Inal A, Alpaslan M, Cakmak I (2006) Genotypic variation in phosphorus efficiency between wheat cultivars grown under greenhouse and field conditions. Soil Sci Plant Nutr 52:470–478. https://doi.org/10.1111/j.1747-0765.2006.00068.x
Hammond JP, Broadley MR, White PJ, King GJ, Bowen HC, Hayden R, Meacham MC, Mead A, Overs T, Spracklen WP, Greenwood DJ (2009) Shoot yield drives phosphorus use efficiency in Brassica oleracea and correlates with root architecture traits. J Exp Bot 60:1953–1968. https://doi.org/10.1093/jxb/erp083
Hansen CW, Lynch JP, Ottosen CO (1998) Response to phosphorus availability during vegetative and reproductive growth in chrysanthemum: whole plant carbon dioxide exchange. J Am Soc Horti Sci 123:215–222. https://doi.org/10.21273/JASHS.123.2.215
Hayes JE, Zhu YG, Mimura T, Reid RJ (2004) An assessment of the usefulness of solution culture in screening for phosphorus efficiency in wheat. Plant Soil 261:91–97. https://doi.org/10.1023/B:PLSO.0000035561.00460.8b
Heffer P (2013) Assessment of fertilizer use by crop at the global level. International Fertilizer Industry Association, Paris
Horst WJ, Abdou M, Wiesler F (1993) Genotypic differences in phosphorus efficiency of wheat. Plant Soil 155:293–296. https://doi.org/10.1007/BF00025040
Hu Y, Ye Z, Shi L, Duan H, Xu F (2010) Genotypic differences in root morphology and phosphorus uptake kinetics in Brassica napus under low phosphorus supply. J Plant Nutr 33:889–901. https://doi.org/10.1080/01904161003658239
Jones GPD, Blair GJ, Jessop RS (1989) Phosphorus efficiency in wheat—a useful selection criterion? Field Crops Res 21:257–264. https://doi.org/10.1016/0378-4290(89)90007-5
Jones GPD, Jessop RS, Blair GJ (1992) Alternative methods for the selection of phosphorus efficiency in wheat. Field Crops Res 30:29–40. https://doi.org/10.1016/0378-4290(92)90054-D
Korkmaz K, Ibrikci H, Karnez E, Buyuk G, Ryan J, Ulger AC, Oguz H (2009) Phosphorus use efficiency of wheat genotypes grown in calcareous soils. J Plant Nutri 32:2094–2106. https://doi.org/10.1080/01904160903308176
Krishnapriya V, Pandey R (2016) Root exudation index: screening organic acid exudation and phosphorus acquisition efficiency in soybean genotypes. Crop Pasture Sci 67:1096–1109. https://doi.org/10.1071/CP15329
Lambers H, Shane MW, Cramer MD, Pearse SJ, Veneklaas EJ (2006) Root structure and functioning for efficient acquisition of phosphorus: matching morphological and physiological traits. Ann Bot 98:693–713. https://doi.org/10.1093/aob/mcl114
Lambers H, Clements JC, Nelson MN (2013) How a phosphorus-acquisition strategy based on carboxylate exudation powers the success and agronomic potential of lupines (Lupinus, Fabaceae). Am J Bot 100:263–288. https://doi.org/10.3732/ajb.1200474
Liao H, Yan X (1999) Seed size is closely correlated to phosphorus use efficiency and photosynthetic phosphorus use efficiency in common bean. J Plant Nutr 22:877–888. https://doi.org/10.1080/01904169909365679
Liao M, Hocking PJ, Dong B, Delhaize E, Richardson AE, Ryan PR (2008) Variation in early phosphorus-uptake efficiency among wheat genotypes grown on two contrasting Australian soils. Aust J Agri Res 59:157–166. https://doi.org/10.1071/AR06311
Lopez-Arrendondo DL, Levya-Gonzalez MA, Gonzalez-Morales SI, Lopez-Bucio J, Herrera-Estrella L (2014) Phosphate nutrition: improving low-phosphate tolerance in crops. Annu Rev Plant Biol 65:95–123. https://doi.org/10.1146/annurev-arplant-050213-035949
Manske GGB, Ortiz-Monasterio JI, Van Ginkel M, Gonzalez RM, Rajaram S, Molina E, Vlek PLG (2000) Traits associated with improved P-uptake efficiency in CIMMYT's semidwarf spring bread wheat grown on an acid Andisol in Mexico. Plant Soil 221:189–204. https://doi.org/10.1023/A:1004727201568
Manske GGB, Ortiz-Monasterio JI, VanGinkel M, Gonzalez RM, Fischer RA, Rajaram S, Vlek PLG (2001) Importance of P uptake efficiency versus P utilization for wheat yield in acid and calcareous soils in Mexico. Eur J Agron 14:261–274. https://doi.org/10.1016/S1161-0301(00)00099-X
McDonald G, Bovill W, Taylor J, Wheeler R (2015) Responses to phosphorus among wheat genotypes. Crop Pasture Sci 66:430–444. https://doi.org/10.1071/CP14191
McNear DH Jr (2013) The rhizosphere-roots, soil and everything in between. Nat Edu Knowledge 4:1
Meena SK, Pandey R, Sharma S, Gayacharan KT, Singh MP, Dikshit HK (2021) Physiological basis of combined stress tolerance to low phosphorus and drought in a diverse set of mungbean germplasm. Agronomy 11:99. https://doi.org/10.3390/agronomy11010099
Mergoum M, Gómez-Macpherson H (2004) Triticale improvement and production. Food & Agriculture Org, Rome, p 179
Murphy J, Riley JP (1962) A modified single solution method for the determination of phosphate in natural waters. Anal Chim Acta 27:31–36. https://doi.org/10.1016/S0003-2670(00)88444-5
Olsen SR, Cole CV, Watanabe FS, Dean LA (1954) Estimation of available phosphorus in soils by extraction with sodium bicarbonate. USDA Circular, p 939
Osborne LD, Rengel Z (2002) Screening cereals for genotypic variation in efficiency of phosphorus uptake and utilization. Crop Pasture Sci 53:295–303. https://doi.org/10.1071/AR01080
Ozturk L, Eker S, Torun B, Cakmak I (2005) Variation in phosphorus efficiency among 73 bread and durum wheat genotypes grown in a phosphorus-deficient calcareous soil. Plant Soil 69:69–80. https://doi.org/10.1007/s11104-004-0469-z
Pandey R, Singh B, Nair TVR (2005) Phosphorus use efficiency of wheat, rye and triticale under deficient and sufficient levels of P. Indian J Plant Physiol 10:292–296
Pandey R, Meena SK, Krishnapriya V, Ahmad A, Kishora N (2014) Root carboxylate exudation capacity under phosphorus stress does not improve grain yield in green gram. Plant Cell Rep 33:919–928. https://doi.org/10.1007/s00299-014-1570-2
Pandey R, Lal MK, Vengavasi K (2018) Differential response of hexaploid and tetraploid wheat to interactive effects of elevated [CO2] and low phosphorus. Plant Cell Rep 37:1231–1244. https://doi.org/10.1007/s00299-018-2307-4
Panigrahy M, Rao DN, Yugandhar P, Raju NS, Krishnamurthy P, Voleti SR, Singh K (2014) Hydroponic experiment for identification of tolerance traits developed by rice Nagina 22 mutants to low-phosphorus in field condition. Arch Agron Soil Sci 60:565–576. https://doi.org/10.1080/03650340.2013.821197
R Development Core Team (2005). R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3–900051–07-0, URL: http://www.R-project.org
Raghothama KG, Karthikeyan AS (2005) Phosphate acquisition. Plant Soil 274:37–49. https://doi.org/10.1007/s11104-004-2005-6
Rengel Z (1999) Physiological mechanisms underlying differential nutrient efficiency of crop genotypes. In: Rengel Z (ed) Mineral nutrition of crops: fundamental mechanisms and implications. Food Products Press, New York, pp 227–265
Rose TJ, Rose MT, Tanaka JP, Heuer S, Wissuwa M (2011) The frustration with utilization: why have improvements in internal phosphorus utilization efficiency in crops remained so elusive? Front Plant Sci 2:73. https://doi.org/10.3389/fpls.2011.00073
Rose TJ, Mori A, Julia CC, Wissuwa M (2015) Screening for internal phosphorus utilisation efficiency: comparison of genotypes at equal shoot P content is critical. Plant Soil 401:79–91. https://doi.org/10.1007/s11104-015-2565-7
Sandana P, Pinochet D (2011) Ecophysiological determinants of biomass and grain yield of wheat under P deficiency. Field Crops Res 120:311–319. https://doi.org/10.1016/j.fcr.2010.11.005
Singh AL, Basu MS (2005) Integrated nutrient management in groundnut- a farmers manual. National Research Center for Groundnut (ICAR). Junagadh, Indian, p 54
Slaski JJ (1994) Differences in the metabolic responses of root tips of wheat and rye to aluminium stress. Plant Soil 167:165–171. https://doi.org/10.1007/BF01587612
Vance CP, Uhde-Stone C, Allan DL (2003) Phosphorus acquisition and use: critical adaptations by plants for securing a non-renewable resource. New Phytol 157:423–447. https://doi.org/10.1046/j.1469-8137.2003.00695.x
Vesterager J, HØgh-Jensen H, Nielsen NE (2006) Variation in phosphorus uptake and use efficiencies between pigeon pea genotypes and cowpea. J Plant Nutr 29:1869–1888. https://doi.org/10.1080/01904160600899436
Wang Y, Lambers H (2019) Root-released organic anions in response to low phosphorus availability: recent progress, challenges and future perspectives. Plant Soil 447:135–156. https://doi.org/10.1007/s11104-019-03972-8
Wang X, Tang C, Guppy CN, Sale PWG (2008) Phosphorus acquisition characteristics of cotton (Gossypium hirsutum L.), wheat (Triticum aestivum L.) and white lupin (Lupinus albus L.) under P deficient conditions. Plant Soil 312:117–128. https://doi.org/10.1007/s11104-008-9589-1
Wang X, Shen J, Liao H (2010) Acquisition or utilization, which is more critical for enhancing phosphorus efficiency in modern crops? Plant Sci 179:302–306. https://doi.org/10.1016/j.plantsci.2010.06.007
Wissuwa M, Gamat G, Islamail AM (2005) Is root growth under phosphorus deficiency affected by source or sink limitation? J Exp Bot 56:1943–1950. https://doi.org/10.1093/jxb/eri189
Wissuwa M, Mazzola M, Picard C (2009) Novel approaches in plant breeding for rhizosphere-related traits. Plant Soil 321:409–430. https://doi.org/10.1007/s11104-008-9693-2
Zhou T, Du Y, Ahmed S, Liu T, Ren M, Liu W, Yang W (2016) Genotypic differences in phosphorus efficiency and the performance of physiological characteristics in response to low phosphorus stress of soybean in southwest of China. Front Plant Sci 7:1776. https://doi.org/10.3389/fpls.2016.01776
Zhu J, Zhang C, Lynch JP (2010) The utility of phenotypic plasticity of root hair length for phosphorus acquisition. Funct Plant Bio 37:313–322. https://doi.org/10.1071/FP09197
Acknowledgements
We acknowledge the Indian Institute of Wheat & Barley Research, Karnal (India), Punjab Agricultural University, Ludhiana (India), Australian Winter Cereals Collection (Australia), and CIMMYT (Mexico) for providing the seed material.
Funding
This study was funded by the Council of Scientific and Industrial Research, New Delhi, India (Grant # 38(1494)/19/EMR-II).
Author information
Authors and Affiliations
Contributions
Conceptualization—RP; funding acquisition—RP; investigation—PRS, DS; methodology—PRS, DS, SS; project administration—RP; resources—RP, AMS; supervision—RP, AMS; validation—SPR; roles/writing–original draft—PRS; writing–review and editing—RP, SS, AMS, DS.
Corresponding author
Ethics declarations
Ethical Approval
Not applicable.
Conflict of Interest
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
ESM 1
(DOCX 1180 kb)
Rights and permissions
About this article

Cite this article
Soumya, P.R., Singh, D., Sharma, S. et al. Evaluation of Diverse Wheat (Triticum aestivum) and Triticale (× Triticosecale) Genotypes for Low Phosphorus Stress Tolerance in Soil and Hydroponic Conditions. J Soil Sci Plant Nutr 21, 1236–1251 (2021). https://doi.org/10.1007/s42729-021-00436-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42729-021-00436-w