
Abstract
A remarkable decrease in olive production has been observed in Tunisia since 2000, particularly in the semi-arid region. This downfall was mainly due to a notable change in climatic conditions as a result of wind erosion and over cultivation. The aim of this work was to study the biological, physical, and chemical properties of several semi-arid soils from olive tree fields subjected to different farming practices, such as crop time and type of crop, olive mill wastewater (OMW) application rates, and tillage timing and depth (deep or conventional plowing). We noted that hydraulic conductivity (HC) was proportional to the age of the soil tillage, and the highest values were recorded in the soil cultivated for 100 years, with an average value of 33.05 ± 0.02 mm h−1. An important increment of the iron soil content was observed, especially after the fig tree introduction among the olive trees (12,094 ng μL−1) in 2007. Also, a significant increase of the organic matter (OM) content (up to2.6 mg-OM/g-soil) was identified in soil treated with OMW compared to the lowest OM content (0.83 mg-OM/g-soil) recorded in the soil cultivated since 1901. The bacterial communities of the different soils were characterized by 454 pyrosequencing technology, and showed an important diversity, mainly corresponding to Proteobacteria, Actinobacteria, and Acidobacteria. Many operational taxonomic units (OTUs) are raretons, indicating a high resilience of the soil bacterial communities. Statistical analyses showed significant correlations with the different soil parameters. However, an unexpected correlation was determined between soil respiration and OM (r = − 0.583*), suggesting that OM increases the retention of CO2, a greenhouse gas. The farming techniques analyzed in this study resulted in a reduction of the bacterial diversity, even though the total bacterial biomass augmented.
Graphical abstract

Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Microbial communities play an important role in soil ecosystems and biogeochemical cycles (Le Bodelier 2011). The soil microbial community is affected by both abiotic factors (such as soil pH, temperature, and structure (Drenovsky et al. 2010)) and agricultural soil management (e.g., crop rotation, tillage, silage, and synthetic fertilizer application (Singh et al. 2014)). Consequently, changes in bacterial communities have been considered early indicators of perturbations in the quality of soil ecosystems (Trivedi et al. 2015). Unadvisable farming practices like soil overexploitation, overgrazing, and poor drainage systems can decrease soil microbial diversity (Foley et al. 2005). Therefore, it is necessary to find new techniques for managing and protecting soil fertility, especially in dry soils which suffer from critical problems (Hannachi et al. 2014).
In the South of Tunisia, the olive tree occupies a crucial social and economic place, although climate aridity and warmer climatic conditions have stressed the productivity of the olive oil sector (Nefzaoui 1991; Sahraoui et al. 2012). Tunisia is considered one of the main olive oil producers in the world, with a 90,000 tons production during the 2018/2019 crop season just in the South of Tunisia (CRDA, Mednine). However, despite its fundamental economic importance, olive oil industries have several drawbacks, like olive mill wastewater (OMW) arising from olive processing, which is one of the strongest and most troublesome industrial effluents (Mekki et al. 2006; Magdich et al. 2016). OMW contains high amounts of organic matter, with both biological oxygen demand (BOD) and chemical oxygen demand (COD) (Pierantozzi et al. 2013), as well as suspended solids. In addition, OMW is characterized by large discharge volumes, high salinity, low pH, and presence of toxic compounds such as polyphenols (Chaari et al. 2014). Nevertheless, the use OMW has been recommended for agricultural purposes due to its properties as fertilizer, mainly its richness in different mineral nutrients: e.g., potassium, sodium, calcium, and phosphates (Mahmoud et al. 2010; Magdich et al. 2016). The effects of OMW on the physical and chemical characteristics of the soil are well documented (Sierra et al. 2001; Mekki et al. 2006).
Although the content of organic matter in soils is globally influenced by climatic factors, crops, the texture of the ground, and topographic conditions (Hannachi et al. 2014), the positive effect of OMW irrigation into the soil structure and soil enrichment is well known (Dakhli et al. 2013; Morugán et al. 2015). Indeed, mineral nutrients, released from OMW or the mineralization of OM used as fertilizers, are important factors for the increasing EC values (Kavvadias et al. 2010).
However, the aforementioned studies did not consider the effects of OMW on the soil microbial community. To the best of our knowledge, few researchers have investigated the impact of the use of OMW on the microbiological population of dry land soils (Hannachi et al. 2014; Rusan et al. 2016). Soils can contain up to 1010 bacteria/g, of which more than 95% have not yet been cultured (Di Serio et al. 2008). OMW spreading constitutes one of the most widespread methods to improve soil fertility, particularly in the case of dry regions. On the other hand, this practice could have secondary effects on soils, such as increasing the level of organic matter, decreasing the instability of soil structure, enhancing the biological activity, and limiting the effects of wind erosion. In addition to OMW spreading, other farming techniques, like deep plowing and the introduction of fig trees (Ficus carica L.), can be implemented to improve the physical properties of soil. However, deep plowing in the plowshare during the winter can favor soil infiltration (Abichou et al. 2009), and the introduction of the intercalary fig trees can reduce wind erosion through boosting roughness and favoring soil microbiological activity because of the increase in shade plants.
The objective of this work was to compare the effect of different farming techniques and OMW application rates on the physical and chemical characteristics of semi-arid soils planted with olive trees. To attain this objective, we tested different adaptation measures according to different assumptions, as follows. (1) OMW spreading would improve the rate of organic matter. The working hypothesis here is that the OMW spreading could increase the organic matter in the soils and thus improve the biological activity in the soils and mitigate the effect of wind erosion. (2) Deep tillage would improve the soil physical properties. The working hypothesis was that deep tillage could promote physical soil fertility by a reduction in surface crusting and bulk density. In addition, the deep tillage would improve the chemical soil fertility by an increase in nutrient levels. And (3) the introduction of the fig tree (Ficus carica L.) into the olive-growing system would enhance soil roughness by microbiological activity thanks to the increase in the shading rate. Likewise, the fig tree would present a potential additional source of income to olive tree agriculture.
To better understand the catalytic role that microorganisms play in the fertility of soils, the microbial communities of different soils have been characterized by 454 pyrosequencing technology.
2 Materials and Methods
In this study, we evaluated changes in the physicochemical properties and microbiota of the soils dependent of crop time (0 to 100 years), type of crop (olive trees and fig trees), and treatment with OMW (spreading for 1 to 10 years) (Table 1, Supp. Material).
2.1 Experimental Sites
Sites in this study were located in the region of Chammakh-Zarzis, Southern Tunisia (33° 36′ N, 11° 02′ E). Fields were under the property of the Tunisian Ministry of Agriculture. The climate within the region is generally classified as arid Mediterranean, with a mean annual rainfall of 180 mm/m2, as a long-term average since 1923 (Abichou and Msellem 2015). The soil in this area is very sandy, with a composition of 85.9% fine sand, 7.7% clay, 5.7% coarse sand, and 0.7% silt in the upper 0–15 cm depth (Abichou et al. 2009). The OMW used in this study was supplied from an olive oil factory (Chammakh-Zarzis) located in the region of Chammakh-Zarzis (Southern Tunisia). The OMW was stored in an evaporation pond for 2 months, the time required for degradation of an important phenolic compound, before its use for spreading (Berndt et al. 1996). Its chemical composition is shown in Table 1.
Six different experimental sites (average 1 ha) were studied (Table 1, Supp. Mat.). S1 was an unfarmed field and used as a control site. S2, S3, S5, and S6 were fields previously planted with olive trees (Olea europaea L.) separated 24 m, and S4 planted with a fig tree (Ficus carica L.) in the middle of two olive trees to improve the ratio of shade. S2 and S3 were intensively farmed for 10 years and 115 years, respectively. S5 and S6 were irrigated with 200 m3/ha and 50 m3/ha of OMW, respectively. The duration of OMW spreading was 10 years for all sites, except 1 year for S5 and S6. The OMW was distributed homogeneously on the sandy soil surface, previously tilled to a 20 cm of depth, during the winter period (December–January). A conventional plowing was done on S2, S3, S4, and S5 during 5 times/year. Deep soil tillage, one time/year, coupled to conventional plowing during 2 times/year, was applied for S6. The deep tillage, applied only in the S6, was used to reduce the conventional tillage frequencies.
2.2 Sampling
At S2, S3, S5, and S6, soil samples were taken from between two olive trees, while at S4, they were obtained from the shade of the fig tree. Soil sample cores (6 cm inner diameter × 20 cm length) were collected with a H-4268 hand auger tube. Each sample analyzed was a mix of four different soil cores collected at each site. Soil samples were taken at a depth of 20 cm because the superficial layer is moderately deep (the bedrock is only 1.5 m deep).
2.3 Analytical Measurements
2.3.1 Soil Properties
The pH and electrical conductivity (EC) of each site were determined with a pHmeter, XP pH50 lab model (LabProcess Badalona, Barcelona), and a conductivity meter, inoLab WTW 7110 model (LabProcess Badalona, Barcelona), respectively. Total organic carbon (TOC) was determined following the Walkley-Black method (Walkley 1947), and organic matter (OM) was calculated by multiplying the total carbon by 1.724 (APHA et al. 1997). Total nitrogen was determined by the Kjeldahl standard method (APHA et al. 1997). Available phosphorus (P2O5) was determined by the Olsen standard method (APHA et al. 1994). Potassium K, Sodium Na, Aluminum Al, Iron Fe, Magnesium Mg, Cadmium Cd, Vanadium V, Manganese Mn, Chrome Cr, and Zinc Zn were determined by atomic absorption spectrometry (Avanta, GBC spectrometer, Australia), using air and acetylene as the mode of oxidation. Biological activity in the soil was obtained by measuring CO2 evolution in aerobic conditions. The soil sample was humidified to 50% of its water holding capacity, and then was incubated at 30 °C in the dark during 24 h. The evolved CO2 was trapped in a sodium hydroxide (NaOH) solution and titrated with hydrogen chloride (HCl) (Öhlinger 1995). Dry particle sizes, such as total sand (TS), coarse sand (CS), and fine sand clay slit (C+S), were measured. In addition, the bulk density (BP) and hydraulic conductivity (HC) in the studied soils were determined (Petard 1993).
2.3.2 Analysis of Bacterial Communities: DNA Extraction and 454 Pyrosequencing
Total DNA extractions from each soil sample were performed using the Power Soil Isolation kit (MoBio Lab, CA). The total genomic DNA (10 ng/μL) was sent for pyrosequencing to MacrogenInc (Korea). Pyrosequencing was performed by making use of a 454 FLX+ System (Roche). Primers 27F and 907R were used to construct the libraries. Primers 27F: (5′AGAGTTTGATCMTGGCTCAG3′) and 907R: (5′CCGTCAATTCMTTTGAGTTT3′), were used to construct the libraries. The technical principle used here, a sequencing and data analysis work flow, is outlined in Sanz and Köchling (2019).
2.3.3 Phylogeny and Statistical Data Analysis
All processing of sequences was performed using the Mothur package v.1.36.0 (www.mothur.org, Schloss et al. 2009) as described elsewhere (Sanz et al. 2017). Additional statistical and graphical evaluations were performed with the Vegan package (Okasanen et al. 2011) of the R language program (http://www.R-project.org/). Complete-linkage (furthest neighbor) clustering was implemented to create the sample and the dendrograms. The data set containing the sequences was deposited in the BioSample database at the National Center for Biotechnology Information (NCBI), under the ID numbers SRR5384076 (S1), SRR5384075 (S2), SRR5384074 (S3), SRR5384073 (S4), SRR5384072 (S5), and SRR5384071 (S6).
2.4 Statistical Analysis
The statistical analysis was performed using the Statistical Package for the Social Sciences (SPSS, Version 20.0). Data are presented as means ± standard deviation (SD). Values were obtained from triplicate determinations and the differences were examined using one-way analysis of variance (ANOVA) followed by Fischer’s LSD (least significant difference) post hoc test. Statistical significances of the correlations between data sets were calculated using Pearson’s R-values.
3 Results
3.1 Chemical and Physical Soil Proprieties
Results of the chemical and physical analyses of the studied soils are summarized in Tables 2 and 3. A significant difference (p = 0.01) among samples was recorded. The pH ranged from 8.31 to 8.73. An alkaline pH (above 8) was revealed in the different study soil. Electrical conductivity of the soil was higher in S4 (1.73 mS/cm) and in S6 (1.23 mS/cm) samples than in the other soils. The soils in the South of Tunisia can maintain a stable pH because of its high buffering capacity.
The size distribution of soil particles showed that the studied soils had total sand abundances varying between 92 and 98%. Coarse sands have abundances between 10.96 ± 0.03% and 18.55 ± 0.02%, while fine sands are more dominant with contents varying between 76.36 ± 0.03 and 85.13 ± 0.07%. The percentages of silt + clay are the lowest, ranging between 1.84 ± 0.02 and 6.92 ± 0.01%.
A variation in the distribution of TS, CS, FS, and C+S between the different soils studied is also observed. An important level of C+S was noted in S3 (soils planted 100 years ago) compared to other sites. The low BD for this soil planted for 100 years (S3) is a consequence of the working of the soil, which has allowed an increase in porosity which in turn decreases the BD. Consequently, the increase in BD at the level of the soil never plowed (below 20 cm depth) is due to the compaction of this soil, and can be explained by a rearrangement of the soil particles due to the low cohesion between them. The results showed that for the six experimental sites, the values of Ks vary between 10.3 ± 0.11 and 33.05 ± 0.02 mm h−1. HC is proportional to the age of working the soil. Indeed, the highest values were recorded for S1 (cultivated for 100 years) with an average value of 33.05 ± 0.02 mm h−1, and lower Ks were obtained in S2 (planted for 10 years) and S6 (planted for 1 year) with values of 15.2 ± 0.05 mm h−1 and 10.05 ± 0.02 mm h−1, respectively.
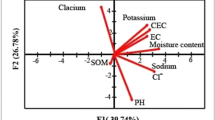
Concerning the soil activity comparison, respiration values in the different soils showed dissimilar behaviors (Tables 2 and 3). A significant difference (p = 0.01) among the samples was registered. The highest soil respiration rates (0.73 mgCO2/g 24 h and 0.68 mgCO2/g 24 h) were recorded in S2 and S6, respectively. The abundances of soil nutrients (N, P, and K), micronutrients, and heavy metals are summarized in Tables 4 and 5. An important difference (p = 0.01) was recorded among the different studied plots with respect to soil nutrient contents, i.e., P2O5, Na, K, Al, Fe, and Mg. The highest contents of P2O5 (4.29 mg P2O5/g-soil) were registered in the soils treated with OMW (S5 and S6). Similarly, the carbon and nitrogen soil contents were significantly higher in the soil treated with the OMW. The heavy metals, e.g., Zn, Cd, and Cr, do not seem to be highly affected by the different treatments (Table 5), except for the V levels registered in S1 (untouched soil) which were lower than in the other soils. Correlation analyses among biological and physical-chemical parameters of the soils studied here indicated several significant trends (Table, SI-2). Significant positive correlations were found: for Al, Mg, Mn, and V with Na (r = 0.811**, 0.932**, 0.781**, and 0.677**, respectively); for Mg, Mn, and V with Al (r = 0.860**, 0.805**, and 0.606**, respectively); for Cd, Mn, and V with Mg (r = 0.546*, 0.920**, and 0.708**, respectively); between Cd and Mn (r = 0.711**); and with Mn and V (r = 0.637**). The different correlations quoted above were produced by ions adsorbed and desorbed into the clay-humus complex, which is selective.
The fertilization using the highest OMW value (200 m3-OMW/ha for 10 years in S5) resulted in an increase of OM up to 2.6 mg-OM/g-soil in S5. This value resembles the value found in the unfarmed soil (S1). The lowest OM soil content was detected in S3: soil cultivated during more than 115 years, with an OM value of 0.83 mg-OM /g-soil.
3.2 Bacterial Communities
The first thing we noted was the high number of OTUs detected in all soils (Table 6). Most of them (more than 90%) were raretons (single- and doubletons, meaning with only one or two reads). The uniformity of the studied soils (Eh and Gini indexes, Table 6) is remarkable, showing a stable and diversified community structure. Raretons most probably do not play any metabolic function, but they could act as a bacterial seederpool, which evolves according to the changing nutritional and physical-chemical conditions. We note that half of the identified sequences could be assigned at the genus level: from 44% in S2 to 55% in S6. These reads encompassed 181 genera. Figure 2-right describes the genera with coverage equal or higher to 1% for, at least, one soil. All these genera are aerobic chemoorganotrophs and many of them have been isolated from soils and agricultural soils.
The high-quality reads were reliably assigned to 11/12 phyla (Fig. 1, left). Proteobacteria (31.5–44.0% of the highest quality reads), Actinobacteria (12.4–35.5%), Acidobacteria (6.8–13.1%), and Bacteroidetes (6.3–12.5%) were predominant in the six sites.

Taxonomic profiles at the phyla phylum (left) and the order (right) levels for the soils studied. All the phyla found have been included. They represent 92–95% of the high-quality reads. Fifty-five orders were identified, covering 86–91% of the reliably taxonomic assigned sequences. In order to clarify the figure: (i) orders with a coverage lower than 1% have been grouped together; (ii) the different orders of the Actinobacteria class considered by Bergey’s Manual 2 ed. have been grouped together in the Actinobacteriales order, according to the Silva Database’s classification (https://www.arb-silva.de/). Note: Six different experimental sites (average one ha) were studied. S2, S3, S5, and S6 were fields previously planted with olive trees (Olea europaea L.) each 24 m, and S4 planted with a fig tree (Ficus carica L.) in the middle of two olive trees. S1 was an unfarmed field and used as a control site
Proteobacteria was the major group of bacteria in five of the six studied soils (Fig. 1, left). This fact should not be surprising because this phylum constitutes the largest phylogenetic lineage and encompasses the largest number of described species. Proteobacteria ranged between 29 and 38.2% of the reads reliably assigned to their order level.
The phylum Actinobacteria demonstrated very high intra-order biodiversity. Their sequences were distributed among the sub-orders Frankineae, Micrococcineae, Micromonosporineae, Propionibacterineae, Pseudonocardineae (Actinobacteria class), Acidimicrobineae (Acidimicrobiia class), and Rubrobacterineae (Rubrobacteria class).
Sequences affiliated to the orders Rhizobiales, Rhodobacterales, Rhodospirillales and Sphingomonadales (Alpha), Burkholderiales and Nitrosomonadales (Beta), Xantomonadales (Gamma), and Myxococcales (Delta) were retrieved. Nitrosomonadales (fam. Nitrosomonadaceae) was the most abundant order, followed a long way behind by Burkholderiales (fams. Comamonadaceae and Oxalobacteraceae) and Rhodospirillales (fam. Rhodospirillaceae) (Figs. 1 and 2).

Heat maps of the most abundant families (left) and genera (right) identified in the soils studied. 108 families (coverage 66–73% of the high-quality reads) and 181 genera (coverage 44–55% high-quality reads) were identified. Families and genera with coverage lower than 1% have been excluded. Note: Six different experimental sites (average 1 ha) were studied. S2, S3, S5, and S6 were fields previously planted with olive trees (Olea europaea L.) each 24 m, and S4 planted with a fig tree (Ficus carica L.) in the middle of two olive trees. S1 was an unfarmed field and used as a control site
Several OTUs could be identified at the genus level with coverage higher than 1% in, at least, one of the analyzed soils (e.g., Nocardioides, Arthrobacter, Flexibacter, Flavisobacter, Sphingomonas) (Fig. 2, right). Importantly, the number of reads affiliated to the phylum Firmicutes was low and unevenly distributed among the analyzed soils: from 0.3% in S1 to 3.7% and 4.0% in S5 and S3 of the total sequences reliably assigned. Firmicutes (Bacillales, Bacillaceae) seem to be almost absent in never-tilled soils (S1), and well adapted into degraded soils (S3 and S5). Our data show numerous OTUs ascribed to the same genus: e.g., in the case of S5, there were 33 different OTUS of Flexibacter, 25 of Flavisolibacter and Gemmatimonas, 23 OTUS of Candidatus Chloroacidobacterium, and 17 OTUS of Candidatus Solibacter. Consequently, it could be hypothesized that many soil bacterium are in a dormant form (with only a few units), waiting to reactivate when the nutritional and physical-chemical conditions are more appropriate. We identified a different significant correlation using the Simpson vs the Shannon indices with certain parameters (Table SI-3): for example, a significant negative correlation was detected between P and N contents using the Simpson index (− 0.508* and − 0.620**, respectively). We conclude that the abundance of certain nutrients (N and P) seems to be decreasing bacterial biodiversity.
4 Discussion
4.1 Chemical and Physical Soil Proprieties
The percentages of silt plus clay are low, which is consistent with the results of previous studies carried out in this area (Abichou and Msellem 2015). An increase in fine particles can make the soil more susceptible to compaction (Bruand et al. 2005), because the silt and clay particles fill the gaps between the grains of sand, resulting in a tighter assembly and an increase in the mass of soil (Bruand et al. 2005). The decrease in water infiltration in compacted soils (plow soil) for S3 was due to a decrease in soil macro porosity, which plays a key role in infiltration (Ankeny et al. 1990) and pore connectivity, which in turn induces a decrease in water circulation and saturated hydraulic conductivity (Silva et al. 2009). Nimmo and Akstin (1989) reported that, in sandy soils with low water content, soil compaction could increase water retention and decrease hydraulic conductivity. Hence, the importance of preventing these forms of soil degradation by improving plowing techniques (reduction in plowing frequencies, lower pneumatic pressure, finer non-compacting tools), ensuring a ground cover that protects the surface against the formation of crusts, and adding organic fertilization that allows the improvement of the soil structure.
The fertilization using the highest OMW value (200 m3-OMW/ha for 10 years) resulted in an increase of OM. The OM in S5 was up to 2.6 mg-OM/g-soil. This value resembles the value found in the unfarmed soil (S1). Although the content of organic matter in soils is globally influenced by climatic factors, crops, the texture of the ground, and topographic conditions (Hannachi et al. 2014), the positive effect of OMW irrigation into the soil structure and soil enrichment is well known (Dakhli et al. 2013; Morugán et al. 2015). Indeed, mineral nutrients, released from OMW or the mineralization of OM used as fertilizers, are important factors for the increasing EC values (Kavvadias et al. 2010). The lowest OM soil content was detected in the soil (S3): soil cultivated during more than 115 years. The OM value was 0.83 mg-OM /g-soil.
Concerning the tillage soil effect, the tillage disintegrates the particles of the soil, reducing the protection of the unstable organic compounds against enzymatic attacks and increases soil porosity, which in turn improves the distribution of gases and access to water for the microorganisms that drive decomposition (La Scala et al. 2008). The soil subjected to an intensive frequency of tillage showed an important soil respiration yield. This yield was usually illustrated by an important CO2 release from the soil (Chatskikh et al. 2008; La Scala et al. 2008). In contrast, in the soil treated with higher doses of OMW (S5), the specific respiration remained very low (0.265 mg CO2/g 24 h). This can be explained by the fact that the phenolic compounds may inhibit soil respiration, which is especially noticeable in soils treated with higher OMW doses (Obied et al. 2005; Mekki et al. 2009). However, the results of these studies are sometimes contradictory and depend on the soil parameters and application rate. Soil respiration inhibition could be also caused by the fact that the largest amount of carbon added to the soil (S5 having a high content of OM) was unavailable to the microflora, due to its strong absorption or its reaction to the components of the soil (Mekki et al. 2006). However, the highest contents of P2O5 were registered in the soils treated with OMW. Previous studies have shown that the soil phosphorous content increases considerably with OMW irrigation (Rusan et al. 2016). Similarly, we found that the carbon and nitrogen soil contents were significantly higher in the soil treated with the OMW. Several studies have indicated that OMW irrigation improves the organic matter content, total N, and C/N ratio, and may have beneficial effects on soil fertility (Mekki et al. 2006; Chaari et al. 2014). Our findings show an important content of potassium in the soil spread with the higher dose of OMW, an important content of iron in the soil planted with a fig tree in the middle of two olive trees (S4), and an increase in nitrogen content in S4 and S5 although the total nitrogen decreased over the year in the other soils. Likewise, the C/N ratio stayed below 5, revealing the occurrence of a mineralization phenomenon (Di Serio et al. 2008). However, the soil nitrogen contents in all studied soils were low: this result was in agreement with other studies, which suggested that the soil total nitrogen contents in the dry region are low due to the soil-leaching phenomenon (Hannachi et al. 2014). In addition, nitrification and nitrogen mineralization can be inhibited by vanadium contents (values shown in Table 3) (Paquelin 1998).
Higher levels of Fe (12,094 mg/L−1) were observed in S4 (soil with fig trees among the olive trees) in contrast to the Fe values found in other soils. This could be attributed to the chelation reactions of Fe with the organic compounds provided by OM (Ylivainio 2010) induced by the fig trees. Moreover, we suspect that the introduction of the fig trees improves the soil roughness by reducing the effect of the wind erosion between two olive trees. In addition, they can increase the shade rate, thereby reducing the soil evaporation and nutrient loss (Abdourhamane Toure et al. 2013).
Also noticeable was the basic pH of the soils studied, despite their calcareous nature, and the null content of Ca2+. This apparent contradiction can be explained by a Ca2+ soil-leaching phenomenon: the calcium is moderately held on the clay-humus complex, and it can be exchanged with other free ions of the soil, such as K+ and Mg2+ (Paquelin 1998). Finally, the relatively high content in Vanadium in S1 can be probably attributed to human activities derived from fertilizer additions (Lambers et al. 2008).
Positive and significant correlations were found for soil respiration (Resp) and soil nutrients, for example, in P2O5, Na, K, and Mg (r = 0.638**, 0.620**, 0.496*, and 0.496*, respectively), supporting the hypothesis that soil respiration improved nutrient solubility, availability, and mobility (Mahmoud et al. 2010). In general, soil pH correlated negatively with EC and Fe (r = − 0.714** and − 0.862**) but positively with Resp (r = 0.627**), which may be attributed to changes in proportions among the different groups of microorganisms (fungi and bacteria) inhabiting the soils due to their different pH requirements (Mohammad and Mazahreh (2003)): bacteria prefer neutral or slightly basic pH, and have higher metabolic activity than fungi. On the other hand, we found a negative correlation between Resp and OM (r = − 0.583*), which seems to be strange. Indeed, this result is in contrast with previous works, which illustrated a highly positive effect between both soil parameters (Brant et al. 2006; Di Serio et al. 2008; Hoorman and Rafiq 2010). The negative correlation we found between the organic matter and the soil Resp could be attributed to carbon soil sequestration, a new strategy to limiting the climate gas effect (Brant et al. 2006; Di Serio et al. 2008; Hoorman and Rafiq 2010). Indeed, the increase in organic matter in this zone would enhance the retention of CO2, a greenhouse gas.
4.2 Bacterial Communities
The majority of the soil microorganisms (80–85%) were in dormant state (Stenström et al. 2001; Fontaine et al. 2003), and the contribution of fresh organic matter can stimulate the microorganisms’ metabolic activity. This short-term modification is known as the priming effect (Buysse and Aubinet 2010).
The dominance of the phylum Proteobacteria can be related to the soil structure. Indeed, the analysis of the spatial distribution of bacteria at microhabitat levels showed that soils characterized by large particle sizes (sand) were dominated by Alpha-Proteobacteria (Torsvik and Ovreas 2002). Most members of Myxococcales develop multicellular spore-filled fruiting bodies in response to starvation (Huntley et al. 2011). In addition, Nitrosomonadales play a critical role in the natural nitrogen cycle (Gaastra et al. 2009). The phylum Actinobacteria demonstrated very high intra-order biodiversity. They are important decomposers in the soil carbon cycle (Di serio et al. 2008). Indeed, many Actinobacteria are usual soil inhabitants, and they have often been isolated from natural environments (soils and waters) (Chouaia et al. 2014). The phylum Acidobacteria is also among the dominant soil phyla (Janssen 2006; Greening et al. 2015). In our study, most reads could be only classified at the family level (Acidobacteriaceae). This was not unexpected because Acidobacteria is a poorly characterized taxon, with less than 10 species currently cultivated and described (Bryant et al. 2007; Greening et al. 2015). Sequences affiliated to Acidobacteria have been retrieved from a wide variety of environments, particularly from soil samples. However, the ecological role of this phylum remains unknown. Lee et al. (2008) suggested that Acidobacteria might be numerically dominant in soils, and might be highly involved in the biogeochemical cycles of the rhizosphere soil. The phylum Bacteroidetes was represented (Figs. 1 and 2) by the orders Sphingobacteriales (fam. Chitinophagaceae) and Cytophagales (fam. Cytophagaceae). The sequences retrieved from our samples are again typical of the soils, i.e., all the species included in Bergey’s Manual of Systematic Bacteriology (Krieg et al. 2011) belonged to the family of Chitinophagaceae, and many of the species affiliated to the Cytophagaceae family were isolated from various types of soils. Other phyla, such as Gemmatiomonadetes (fam. Gemmatiomonadaceae), Planctomycetes (fam. Planctomycetaceae), Chloroflexi (Chloroflexales order), and Verrucomicrobia, were also identified in our soils. Even though in less proportion than the other phyla discussed above, these rarely cultivated taxa could be among the most dominant soil phyla (Greening et al. 2015).
The high number of raretons detected is outstanding: e.g., 48.0% in S6, or 57.5% in S3. According to Hobbie and Hobbie (2013), soil microbes have a starving-survival lifestyle of dormancy, arrested activity, and low activity. In this sense, Greening et al. (2015) stated that “the majority of microbial cells in global soils exist in a spectrum of dormant states.” Moreover, one strategy used by bacteria to survive in a competitive and changing environment, such as the soil, is to phenotypes that may be less adapted to the current conditions, but might confer an advantage in the future (Stum and Dworkin 2015). The comparative analysis between the different types of treated soils by Torsvik et al. (1996) showed that anthropogenic influences, such as agricultural management (tilling, manuring, or cropping) and pollution, can have a profound impact on bacterial communities, leading to reduced bacterial diversity, even though the bacterial biomass may be increased.
5 Conclusions
In our study, the highest content of potassium was detected in the soil spread with the higher dose of olive mill wastewater; the maximum content of iron was identified in the soil with fig trees among olive trees; the total nitrogen was found to decrease over the years; and hydraulic conductivity was proportional to the age of the soil tillage. We could not establish (by analysis of canonical variables) any clear relationships between agricultural practices and the microorganisms found in the different soils. However, two important facts can be highlighted: (i) the high number or sequences retrieved only once or twice (raretons), together with the high number of OTUs affiliated to the same genus, showing a high adaptability and resilience of these microorganisms to nutritional and physical-chemical changes in the soils; and (ii) the significant negative correlation between nutrients and bacterial diversity, which seems to imply that the availability of nutrients favors, over time, the predominance of the bacterial species that are able to use them more efficiently (strategist r), while a low content of nutrients allows the survival of a greater number of species capable of extracting scarce resources (strategist k). The combination of a moderate OMW spreading dose with the introduction of fig trees and an intensive tillage must be taken into consideration in the design of future surveys.
References
Abdourhamane Toure A, Bergametti G, Bielders C, Bouet C, Callot Y, Dupont S, Khatteli H., Labiad MT, Marticorena B, Rajot JL, Valentin C (2013) Erosion éolienne dans les régions arides et semi-arides africaines : processus physiques, métrologie et techniques de lutte Djerba
Abichou M, Msellem A (2015) The effects of climate change on the olive tree phenology. Int J Glob 4:2513–2517. https://doi.org/10.5604/12321966.1167706
Abichou M, Labiadh M, Cornelis W, Gabriels D, Ben Rouina B, Taamallah H, Khatteli H (2009) The olive mills waste water (OMW) as an organic amendment for controlling wind erosion in Southern Tunisia by improving the soil surface structure. Arid Land Stud 19(1):343–346
Ankeny MD, Kaspar TC, Horton R (1990) Characterization of tillage and traffic effects on unconfined infiltration measurements. Soil Sci Soc Am J 54(3):837–840. https://doi.org/10.2136/sssaj1990.03615995005400030037x
APHA, AWWA, WPCF (1994) Standard methods for the examination of water and wastewater. American Public Health Association, Washington DC, USA
APHA, AWWA, WPCF (1997) Standard methods for the examination of water and wastewater. American Public Health Association, Washington DC, USA
Berndt L, Fiestas Ros, JA, De Ursinos K, Geissen M, Kachouri E, Klimm GL, Monpezat D, Xanthoulis D (1996) Les expériences méditerranéennes dans le traitement et l’élimination des eaux résiduaires des huileries d’olives, Ed, Deutsche Gesellschaftfür Technische Zusammenarbeit
Brant JB, Sulzman EW, Myrold DD (2006) Microbial community utilization of added carbon substrates in response to long-term carbon input manipulation. Soil Biol Biochem 38:2219–2232. https://doi.org/10.1016/j.soilbio.2006.01.022
Bruand A, Hartmann C, Lesturgez G (2005) Physical properties of tropical sandy soils: a large range of behaviours
Bryant DA, Garcia Costas AM, Maresca JA, Maqueo Chew AJ, Klatt CG, Bateson MM, Tallon LJ, Hostetler J, Nelson WC, Heidelberg JF, Ward DM (2007) Candidatus Chloracidobacterium thermophilum: an aerobic phototrophic acidobacterium. Science 317:523–526
Buysse P, Aubinet M (2010) La respiration hétérotrophe dans les sols agricoles : description des facteurs importants et comparaison de modèles semi-mécanistes existants. Biotechnol Agron Soc Environ 14(4):707–717
Chaari L, Elloumi N, Mseddi S, Gargouri K, BourouinaB MT, Kallel M (2014) Effects of olive mill wastewater on soil nutrients availability. Int J Interdiscip 2:175–183
Chatskikh D, OlesenJE HEM, Elsgaard L, Petersen BM (2008) Effects of reduced tillage on net greenhouse gas fluxes from loamy sand soil under winter crops in Denmark. Agric Ecosyst Environ 128:117–126. https://doi.org/10.1016/j.agee.2008.05.010
Chouaia B, Gaiarsa S, Crotti E, Comandatore F, DegliEsposti M, Ricci I, Alma A, Favia G, Bandi C, Daffonchio D (2014) Acetic acid bacteria genomes reveal functional traits for adaptation to life in insect guts. Genome Biol Evol 6(4):912–920. https://doi.org/10.1093/gbe/evu062
Dakhli R, Lamouri R, Taamallah H, Ouessar M (2013) Short term effects of olive mill waste water on soil chemical properties under semi-arid Mediterranean conditions. J Life Sci 7(11):1209–1218
Di Serio MG, Lanza B, Mucciarella MR, Russi F, Iannucci E, Marfisi P, Madeo A (2008) Effects of olive mill wastewater spreading on the physico-chemical and microbiological characteristics of soil. Int Biodeterior Biodegradation 62:403–407. https://doi.org/10.1016/j.ibiod.2008.03.006
Drenovsky RE, James JJ, Richards JH (2010) Variation in nutrient resorption by desert shrubs. J. Arid Environ 74:1564–1568. https://doi.org/10.1016/j.jaridenv.2010.05.030
Foley JA, De Fries R, Asner GP, Barford C, Bonan G, Carpenter SR, Chapin FS, Coe MT, Daily GC, Gibbs HK, Helkowski JH, Holloway T, Howard EA, Kucharik CJ, Monfreda C, Patz JA, Prentice IC, Ramankutty N, Snyder PK (2005) Global consequences of land use. Science 309:570–574. https://doi.org/10.1126/science.1111772
Fontaine S, Mariotti A, Abbadie L (2003) The priming effect of organic matter: a question of microbial competition? Soil Biol Biochem 35:837–843. https://doi.org/10.1016/S0038-0717(03)00123-8
Gaastra W, Boot R, Ho HT, Lipman LJ (2009) Rat bite fever. Vet Microbiol 133:211–228. https://doi.org/10.1016/j.vetmic.2008.09.079
Greening C, Carere CR, Rushton-Green R, Harold LK, Hards K, TaylorMC MSE, Stott MB, Cook GM (2015) Persistence of the dominant soil phylum Acidobacteria by trace gas scavenging. PNAS 112(33):10497–10502. https://doi.org/10.1073/pnas.1508385112
Hannachi N, Cocco S, Fornasier F, Agnelli A, Brecciaroli G, Massaccesi L, Weindorf D, Corti G (2014) Effects of cultivation on chemical and biochemical properties of dryland soils from southern Tunisia. Agric Ecosyst Environ 199:249–260. https://doi.org/10.1016/j.agee.2014.09.009
Hobbie JE, Hobbie EA (2013) Microbes in nature are limited by carbon and energy: the starving-survival life style in soil and consequences for estimating microbial rates. Front Microbiol 4:324. https://doi.org/10.3389/fmicb.2013.00324
Hoorman J, Rafiq I (2010) Understanding soil microbes and nutrient recycling. Agriculture and Natural Resources, the Ohio State University
Huntley S, Hamann N, Wegener-Feldbrugge S, Treuner-Lange A, Kube M, Reinhardt R, Klages S, Muller RM, Ronning CC, Nierman W, Søgaard-Andersen L (2011) Comparative genomic analysis of fruiting body formation in myxococcales. Mol Biol Evol 28(2):1083–1097. https://doi.org/10.1093/molbev/msq292
Janssen PH (2006) Identifying the dominant soil bacterial taxa in libraries of 16S rRNA and 16S rRNA genes. Appl Environ Microbiol 72(3):1719–1728. https://doi.org/10.1128/AEM.72.3.1719-1728.2006
Kavvadias V, Doula MK, Komnitsas K, Liakopoulou N (2010) Disposal of olive oil mill wastes in evaporation ponds: effects on soil properties. J Hazard Mater 182:144–155. https://doi.org/10.1016/j.jhazmat.2010.06.007
Krieg NR, Staley JT, Brown DR, Hedlund BP, Paster BJ, Ward NL, Ludwig W, Whitman WB (2011) Bergey’s manual of systematic bacteriology. In: Vol. Four: The Bacteroidetesand other phyla, 2nd edn. Springer, New York
La Scala JN, Lopes A, Spokas K, Bolonhezi D, Archer DW, Reicosky DC (2008) Short-term temporal changes of soil carbon losses after tillage described by a first-order decay model. Soil Till Res 99:108–118. https://doi.org/10.1016/j.still.2008.01.006
Lambers H, Chapin FS, Pons TL (2008) Plant physiological ecology, 2nd edn. Springer, New York
Le Bodelier P (2011) Interactions between nitrogenous fertilizers and methane cycling in wetland and upland soils. Curr Opin Environ Sustain 3(5):379–388. https://doi.org/10.1016/j.cosust.2011.06.002
Lee SH, Ka JO, Cho JC (2008) Members of the phylum Acidobacteria are dominant and metabolically active in rhizosphere soil. FEMS Microbiol Lett 285(2):263–269. https://doi.org/10.1111/j.1574-6968.2008.01232.x
Magdich S, Abid W, Boukhris M, Ben Rouina B, Ammar E (2016) Effects of long-term olive mill wastewater spreading on the physiological and biochemical responses of adult Chemlali olive trees (Oleaeuropaea L.). Ecol Eng 97:122–129. https://doi.org/10.1016/j.ecoleng.2016.09.004
Mahmoud M, Janssen M, Haboub N, Nassour A, Lennartz B (2010) The impact of olive mill wastewater application on flow and transport properties in soils. Soil Till Res 107:36–41. https://doi.org/10.1016/j.still.2010.01.002
Mekki A, Dhouib A, Sayadi S (2006) Changes in microbial and soil properties following amendment with treated and untreated olive mill wastewater. Microbiol Res 161:93–101. https://doi.org/10.1016/j.micres.2005.06.001
Mekki A, Dhouib A, Sayadi S (2009) Evolution of several soil properties following amendment with olive mill wastewater. Prog.Nat.Sci. 19:1515–1521. https://doi.org/10.1016/j.pnsc.2009.04.014
Mohammad MJ, Mazahreh N (2003) Changes in soil fertility parameters in response to irrigation of forage crops with secondary treated wastewater. Commun Soil Sci Plan 34(9–10):1281–1294. https://doi.org/10.1081/CSS-120020444
Morugán C, Orenes F, Cerdà A (2015) Changes in soil microbial activity and physicochemical properties in agricultural soils in Eastern Spain. Spanish J Soil Sci 5(3)
Nefzaoui A (1991) Contribution à la rentabilité de l’oléiculture par une valorisation optimale des sous-produits. Option méditerranéennes, 16 : 101–108
Nimmo JR, Akstin KC (1989) Hydraulic conductivity of a sandy soil at low water content after compaction by various methods. Soil Sci Am J 52:303–310. https://doi.org/10.2136/sssaj1988.03615995005200020001x
Obied HK, Allen MS, Bedgood DR (2005) Bioactivity and analysis of biophenols recovered from olive mill waste. J Agri Food Chem 53:823–837. https://doi.org/10.1021/jf048569x
Öhlinger R (1995) Soil respiration by titration. In: Margesin (ed) Methods in soil biology. Springer, Berlin, pp 95–98
Okasanen J, Blanchet FG, Kindt R, Legendre P, O’Hara RB (2011) Vegan: community ecology package. R Package Version 2.3-5. The Comprehensive R Archive Network Website
Paquelin D (1998) Largile et lamotte : la fertilité de sol maîtrisée. Educagri:E-212-34
Petard J (1993) Les Méthodes d’analyse Tome1 Analyse de Sol; 192p
Pierantozzi P, Torres M, Verdenelli R, Basanta M, Maestri DM, Meriles JM (2013) Short-term impact of olive mill wastewater (OMWW) applications on the physico-chemical and microbiological soil properties of an olive grove in Argentina. J Environ Sci Heal A 48:393–401. https://doi.org/10.1080/03601234.2013.742398
Rusan MMJ, Albalasmeh AA, Malkawi HI (2016) Treated olive mill wastewater effects on soil properties and plant growth. Water Air Soil Pollut 227:135. https://doi.org/10.1007/s11270-016-2837-8
Sahraoui H, Jrad A, Mellouli HJ (2012) Épandage des margines sur les sols agricoles : impacts environnementaux microbiologiques. Afrique Science : Revue Internationale des Sciences et Technologie 8(1):97–106
Sanz JL, Köchling T (2019) Next-generation sequencing and waste/wastewater treatment: a comprehensive overview. Rev Environ Sci aBio 18:635–680. https://doi.org/10.1007/s11157-019-09513-0
Sanz JL, Rojas P, Morato A, Méndez L, Ballesteros M, González-Fernández C (2017) Microbial communities of biomethanization digesters fed with raw and heat pre-treated microalgae biomasses. Chemosphere 168:113–121. https://doi.org/10.1016/j.chemosphere.2016.10.109
Schloss PD, Westcott SL, Ryabin T, Hall JR, Hartmann M, Hollister EB, Lesniewski RA, Oakley BB, Parks DH, Robinson CJ (2009) Introducing Mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl Environ Microbiol 75(23):7537–7541. https://doi.org/10.1128/AEM.01541-09
Sierra J, Marti E, Montserrat G, Cruanas R, Garau MA (2001) Characterisation and evolution of a soil affected by olive oil mill wastewater disposal. Sci Total Environ 279:207–214. https://doi.org/10.1016/S0048-9697(01)00783-5
Silva J, Nichols J, Theunissen TW, Anouk GG, Oosten LV, Barrandon O, Wray J, Yamanaka S, Chambers I, Smith A (2009) Nanog is the gateway to the pluripotent ground state.2008. Cell Press. Volume 138, Issue 4, Pages 722–737. https://doi.org/10.1016/j.cell.2009.07.039
Singh ND, Koerich LB, Carvalho AB, Clark AG (2014) Positive and purifying selection on the Drosophila y chromosome. Mol Biol Evol 31(10):2612–2623. https://doi.org/10.1093/2Fmolbev%2Fmsu203
Stenström J, Svensson K, Johansson M (2001) Reversible transition between active and dormant microbial states in soil. FEMS Microbiol Ecol 36:93–104. https://doi.org/10.1111/j.1574-6941.2001.tb00829.x
Stum A, Dworkin J (2015) Phenotypic diversity as a mechanism to exit cellular dormancy. Curr Biol 25:2272–2277. https://doi.org/10.1016/j.cub.2015.07.018
Torsvik V, Ovreas L (2002) Microbial diversity and function in soil: from genes to ecosystems. Curr Opin Microbiol 5(3):240–245 https://www.ncbi.nlm.nih.gov/pubmed/12057676
Torsvik V, Sorheim R, Goksoyr J (1996) Total bacterial diversity in soil and sediment communities. J Ind Microbiol 17:170–178. https://doi.org/10.1007/BF01574690
Trivedi P, Rochester IJ, Trivedi C, Van Nostrand JD, Zhou J, Karunaratne S, Anderson IC, Singh BK (2015) Soil aggregate size mediates the impacts of cropping regimes on soil carbon and microbial communities. Soil Biol Biochem 91:169–181. https://doi.org/10.1016/j.soilbio.2015.08.034
Walkley A (1947) A critical examination of a rapid method for determining organic carbon in soils- effect of variations in digestion conditions and inorganic soil constituents. Soil Sci 63:251–264
Ylivainio K (2010) Effects of iron (III) chelates on the solubility of heavy metals in calcareous soils. Environ Pollut 158:3194–3200. https://doi.org/10.1016/j.envpol.2010.07.004
Acknowledgments
We are very thankful to Ana I. Morato (Department of Molecular Biology, Universidad Autonoma de Madrid, Spain) for providing her skillful technical assistance. The authors would like to thank Esther Ruiz-Uceda for her valuable review of the spelling and grammar of the manuscript.
Funding
This work was supported by grants from the Tunisian Ministry of Agriculture and Water Resources, and by the International Fund for Agricultural Development (IFAD).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
ESM 1
(DOCX 22 kb)
Rights and permissions
About this article

Cite this article
Rajhi, H., Sanz, J.L., Abichou, M. et al. The Impact of Different Techniques of Soil Management on Soil Fertility and the Associated Bacterial Communities in Semi-arid Olive Tree Fields. J Soil Sci Plant Nutr 21, 547–558 (2021). https://doi.org/10.1007/s42729-020-00382-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42729-020-00382-z