
Abstract
Assessments of compatibility between targeted insecticides and beneficial organisms are important for integrated pest management programs. In this study, we investigated the effect of some biorational insecticides: D-limonene (applied at 1 g/L), mineral oil (applied at 20 g/L), potassium salts of fatty acid (applied at 0.4 g/L), pyriproxyfen (applied at 0.25 g/L), and potassium salts of fatty acid (C7-C18) (applied at 3 g/L) on the functional response of Cryptolaemus montrouzieri (Mulsant) feeding on Dactylopius opuntiae (Cockerell) females under controlled conditions (26 °C). Previous experiments established that the insecticide rates used in the present study were sublethal, as they did not cause short-term mortality to the predator. In the absence of insecticides, the predator exhibited a type II functional response. All insecticide treatments in this study did not hamper the ladybird functional response, but it altered the shape of the curve compared to the control (tap water). Our results showed that all insecticides treatments residues do not have lethal effects on C. montrouzieri adult, and do not affect significantly his predation. Pyriproxyfen and potassium salts of fatty acid (C7-C18) seem to be the least compatible with C. montrouzieri as it led to both a significant reduction of attack rate (0.01–0.05) and an increase in handling time (2.57–12.77 h). In contrast D-limonene (applied at 1 g/L), mineral oil (applied at 20 g/L) were the most compatible with the predator, as it led to both a significant increase in attack rate (0.05–0.12) and a decrease in handling time (3.91- 5.71 h).
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In the last few years, many insect predators have been consistently observed associated to Dactylopius opuntiae (Cockerell) (Hemiptera: Dactylopiidae) colonies worldwide (Vanegas-Rico et al. 2010; Castro 2011; Lima et al. 2011; Barbosa et al. 2014; Giorgi et al. 2017; El Aalaoui et al. 2019a). For control of D. opuntiae, biological control is usually accompanied by chemical control application in the field. Generally, the effect of insecticides on natural enemies take into consideration by integrated pest management (IPM) programs. Natural enemies are often more sensitive to insecticides compared with other insects (Khan et al. 2012), and the use of pesticides compatible and safe to biological control agents was recommended. Recently many research studies showed the sublethal effects of pesticides that can affect biological and reproduction parameters, and behavior of predators (Halappa et al. 2013; Planes et al. 2013; Anjitha et al. 2013; Martinou et al. 2014; Wanumen et al. 2016; Xiao et al. 2016; Nawaz et al. 2017; El Aalaoui et al. 2021). But, a complete understanding of the impact of plant protection products on predation potential of natural enemies is still lacking (Martinou and Stavrinides 2015).
The predation potential of predators has two components (predator’s numerical and functional response). Functional response defined by the number of prey attacked per predator as a function of prey density (Solomon 1949; Holling 1959), and numerical response can be defined by the number of its progeny in relation to increasing prey density (Solomon 1949). The functional response is a good indicator of the suitability of a predator as a biological control agent against target pest (Wiedenmann and O’Neil 1991; Fernàndez -Arhex and Corley 2004). Many types of functional responses are described for biological control agents (Hodek 1996). Among these types of functional responses, type II and III in which predators cause negative (type II) or positive (type III) density-dependent mortality of their prey have received the most attention (Murdoch et al. 2003), because most natural enemies show these types, and type II response was the most common type for coccinellids predators (Collins et al. 1981; Seo and DeAngelis 2011). Functional response models should not be ignored in integrated pest management (IPM) programs, in order to identify predators that impose positively density-dependent mortality on prey species (type III functional response) because such mortality is thought to stabilize prey populations (Murdoch et al. 2003; Martinou and Stavrinides 2015). These models are also exploited by ethologists to estimate predator dynamics, and to understand prey-predator relationships (Hassell et al. 1977; Murray et al. 2013). Yet, the effects of pesticides on the functional response of many important natural enemies have not been investigated.
Recently, the role of Cryptolaemus montrouzieri (Coccinellidae: Scymninae) as a biological control agent against D.opuntiae is expected to gain further importance. This lady beetle was introduced into Brazil for biological control of D.opuntiae, and primarily mealybug attacking cassava and citrus (Sanches and Carvalho 2010). In Israel, 100,000 naturalists C. montrouzieri were successfully released in infested areas in the north of the country (Protasov et al. 2017). The ladybeetle was also introduced into Morocco to control D.opuntiae. Laboratory studies have shown positive results for successful development, reproduction, and predation on the mealybug (El Aalaoui et al. 2019a, b, c). In the field, the release of 30 C. montrouzieri adults per m2 of ground surface significantly reduced the mealybug population by 92% within 77 days of their release in areas with minimum temperatures above 5 °C (Bouharroud et al. 2018).
D-limonene is a monoterpene extracted from citrus peels (Kim et al. 2004) and likely acts on insect nervous system (Karr 1989). D-Limonene has been frequently used to control pest diseases. Indeed, the toxic effect of D-limonene on pests was started by Taylor and Vickery (1974). Also, this botanical insecticide acts as a reproduction inhibitor and growth regulator on many insect species (Karr 1989). Brennan et al. (2013) found that D-limonene causes chitin degradation in pests. D-limonene (60 g/l) has been found to be compatible with C. montrouzieri. Feeding on D. opuntiae treated females with this botanical insecticide does not result in significant mortality for all C. montrouzieri larval stages and pupae compared to the control treatment (tap water), also, does not affect significantly the predator long-term population parameters, including the mean generation time, the intrinsic rate of natural increase, the finite rate of increase compared to the control treatment (tap water) (El Aalaoui et al. 2021).
Mineral oil blocks spiracles in pest adults and nymphs and prevents gas exchange in eggs, causing asphyxiation and death (Cranshaw and Baxendale 2011; Helmy et al. 2012). Mineral oil has been used frequently in agriculture against several pests on a variety of crops (Agnello 2002; El Aalaoui et al. 2019c). Many studies showed that mineral oil has ovicidal activity (Smith and Pearce 1948; Riedl et al. 1995) and sometimes may prevent pest oviposition (Zwick and Westigard 1978; Riedl et al. 1995; Fernandez et al. 2001). Mineral oil reduces the number of eggs laid as well as the level of egg survival of pests (Wins-purdya et al. 2009). Mineral oil has been shown to be highly compatible with C. montrouzieri. This biological insecticide does not significantly affect C. montrouzieri females fertility, does not cause any reduction in egg hatch percentage (100%), and does not affect significantly the predator long-term population parameters compared to the control treatment (tap water) (El Aalaoui et al. 2021).
The potassium salts of fatty acids are contact agents and act by penetrating inside the integument of arthropods, clogging cell membranes, and causing dehydration and death (Tsolakis and Ragusa 2008). Since their discovery as biocidal agents (1947), fatty acid salts have been reported to have very low toxicity to humans and are considered safe to the environment (Prats et al. 1999), although their phytotoxicity limits their use on some crops (Pasini et al. 1997). Potassium salts of fatty acid affect the ovicidal of many insect species (Osborne and Petitt 1985). In addition, potassium salts of fatty acid showed a negative impact on survival rate (direct loss of females) and fecundity (decreased viability of the eggs and population) of many arthropods (Tsolakis and Ragusa 2008). Sublethal concentrations of this insecticide do not alter the development time of C. montrouzieri larvae and pupae compared to the control treatment (tap water) (El Aalaoui et al. 2021).
Pyriproxyfen, an analog of the juvenile insect hormone, causes marked suppression of metamorphosis, embryogenesis, and adult development (Rimoldi et al. 2017). Insect growth regulators (IGRs) generally do not affect egg survival of natural enemies (Rill et al. 2008; Rimoldi et al. 2008). However, the mode of action of pyriproxyfen is more specific and depends on the presence of insecticide-specific endocrine receptors in embryos that develop inside of eggs (Sullivan and Goh 2008).
In the present study, we investigated the effects of sublethal concentrations of the insecticides: D-limonene, mineral oil, potassium salts of fatty acid, and pyriproxyfen on the functional response parameters of C. montrouzieri feeding on D.opuntiae.
Materials and methods
Predator
The C. montrouzieri colony was developed from adults introduced by the Entomology Laboratory of INRA-Morocco, as described by EL-Aalaoui et al. (2020). Cryptolaemus montrouzieri adults were placed in entomological cages (80 × 80x80 cm) consisting of a wooden frame covered with a mesh fabric to allow ventilation and maintained for more than 50 generations under 26 ± 2 °C and 60 ± 10% RH in the laboratory. Access to water was provided by a cotton wick inserted in a 25 ml glass water bottle. Opuntia ficus-indica (L.) (Miller, 1768) cladodes infested with D. opuntiae collected from the fields at Zemamra in Morocco (32°37′48" N, 8°42′0" W) were introduced into the cages weekly to provide food and substrate for C. montrouzieri females oviposition. C. montrouzieri new emerged larvae were moved to another cage with the same characters described above to complete their development and avoid the risk of cannibalism. Also, all reared ladybug beetle adults received supplementary feeding (a mixture of water, honey, and brewer's yeast in a proportion of 20:40:40), which was replaced daily. All trials with C. montrouzieri in this study were carried out at 26 ± 2 °C, 60 ± 10% RH, and 12:12 (L:D) hours.
Insecticides
The effect of five insecticides in the predation potential of C. montrouzieri (a potential predator against D.opuntiae) was evaluated in this study. Limocide (60 g D-limonene per L; applied at 1 g/L; Vivagro, Martillac, France), Insecticide 101 (780 g mineral oil per L; applied at 20 g/L; UPL, Ankleshwar, India), Hamper (500 g potassium salts of fatty acid per L; applied at 0.4 g/L; Gowan Crop Protection, Faenza, Italy). Brai 10 EC (100 g pyriproxyfen per L; applied at 0.25 g/L; Lainco company, Barcelona, Spain), and Nakar (500 g potassium salts of fatty acid (C7-C18) per L; applied at 3 g/L; Seipasa company, Valencia, Spain). The selected insecticides have different modes of action.
All the insecticides concentration used in this study had high mortality against nymphs of D.opuntiae (74.46–97.54% mortality, 24 h after treatment) (El Aalaoui et al. 2019c). D-limonene (60 g/L), and mineral oil (780 g/L) had high mortality against both nymphs and adult females (90.28% and 91.94% mortality, 120 h after treatment respectively) of the scale pest under semi-field conditions (El Aalaoui et al. 2019c). Also, the rate of each insecticide used in the current study was sublethal, as they did not cause short-term mortality to the predator (El Aalaoui et al. 2021).
Trail
A determined amount of each treatment was poured into one liter of tap water and sprayed onto the petri dish (9.5 cm diameter) and its lid cover at a rate of 1 ml of pesticide solution, resulting in a deposition of 2.55 mg/cm2 which is similar to what is recommended for biological trials according to the IOBC working group "Pesticides and Beneficial Organisms" (Candolfi et al. 2001; Martinou and Stavrinides 2015). A Potter spray tower (Burkard Scientific Ltd, Uxbridge, UK) was used to apply insecticide solution of a prescribed active ingredient and concentration as a mist over Petri dishes at 150 kPa. We use Petri dishes in this study as a testing substrate because the use of living plants (cladodes) could affect predator–prey consumption and act as a confounding factor in functional response modeling (Martinou and Stavrinides 2015). Control Petri dishes were sprayed with tap water. The predator (C. montrouzieri) and D. opuntiae females were not sprayed. After spraying, the Petri dishes and their lids were allowed to dry out for 24 h in the laboratory condition, then a fixed number of D.opuntiae females collected from infested cactus in fields at Zemamra and El-Gharbia (33°15' N, 8°30' W) localities, Morocco were introduced in each Petri dishes at the following densities: 1, 5, 10, 15, 20, and 25 with a paintbrush. An individual C. montrouzieri adult stage (72 h old) was transferred in each Petri dish and was allowed to forage for 24 h, after which it was removed and the consumed D.opuntiae females were counted. Each density was replicated 10 times for each of the five pesticide treatments and the control, and all experiment was replicated five times. The mean body weights and size of D. opuntiae adult females used in the studies were 5.40 ± 0.5 mg, and 2.33–3.27 mm.
Functional response
Briefly, the functional response was determined by fitting a logistic regression of the proportion of prey consumed (Ne) against prey offered (N0) (Trexler et al. 1988; Pervez 2005). The relationship between \(Ne\)/N0 and N0 was estimated by Juliano equation (Juliano 2001): \(Ne/N_0=\left(exp\left(P_0+P_1N_0{^2}+P_3N_0{^3}\right)\right)/\left(1+exp\left(P_0+P_1N_0{^2}+P_3N_0{^3}\right)\right)\), Where P0, P1, P2, and P3 represent the intercept, linear, quadratic, and cubic coefficients to be estimated, respectively. Juliano (2001) reported that P1 > 0 and P2 < 0, indicate density-dependent predation and describing a type III functional response. If P1 < 0, the proportion of prey consumed declines monotonically with the initial number of prey offered, thus describing a type II functional response.
The results showed that the functional response type exhibited by this predator in all treatments was type II. The type II functional response model is described by the equation: \(Ne\)=\(aTN\) 0/(1 + aN0\(Th)\)—where \(Ne\) is the number of preys consumed, \(a\) is the attack rate or the instantaneous rate of discovery, \(N\) 0 the initial number of prey offered or prey’s density, \(T\) the time when prey and predator are mutually exposed or the experiment duration (24 h) and \(Th\) the handling time associated with each prey consumed. Handling time (Th) and attack rate (\(a\)) were estimated by both the disc equation (Holling 1959) and the random attack equation (Rogers 1972). Mean values of Th were used to calculate maximum attack rate defined as (\(T/Th\)), and searching time (\(Ts\)): \(Ts=T-ThNe\) (Juliano and Williams 1987; Elliot 2003; Flores et al. 2013). The data of handling time (Th), attack rate (\(a\)), and maximum attack rate (\(T/Th\)) were subjected to analysis using Tukey’s LSD test (α = 0.05) with the software package SPSS ver. 18.0 (Carver and Nash 2011).

Type II functional response models for C. montrouzieri for the five insecticide treatments and the control. A) D-limonene (60 g/L) applied at 1 g/l, B) mineral oil (780 g/L) applied at 20 g/L, C) potassium salts of fatty acid (500 g/l) applied at 0.4 g/L, D) Pyriproxyfen (100 g/l) applied at 0.25 g/L, E) potassium salts of fatty acid (C7-C18) (500 g/L) applied at 3 g/L, F control (tap water)
Results
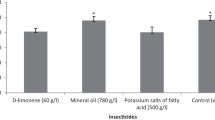
For all the insecticide treatments, and control tested in this study, the values of P1 were negative, indicating a type II functional response (Fig. 1). The insecticides tested did not alter the type of the functional response, but it altered the shape of the curve compared to the control (Fig. 1). The number of prey consumed by C. montrouzieri at different prey densities varied significantly among treatments (f = 52.751, df = 5, p ≤ 0.05). The number of preys consumed was significantly higher for D-limonene, and mineral oil than for the control and other insecticide treatments tested. The model estimated handling time was significantly higher (f = 15.167, df = 5, p ≤ 0.05) for pyriproxyfen, potassium salts of fatty acid (C7-C18), and control treatments, and low for D-limonene, and mineral oil treatments. There were no significant differences in handling time among potassium salts of fatty acid (C7-C18), potassium salts of fatty acid, and control treatments (Table 1). The attack rate of the predator was significantly higher (f = 116.704, df = 5, p ≤ 0.05) for D-limonene treatment than for all the other treatments tested. No significant difference in attack rate was observed between potassium salts of fatty acid, and control treatments (Table 1). The maximum attack rate value was significantly higher (f = 7.029, df = 5, p ≤ 0.05) for D-limonene, and mineral oil, and similar for control and potassium salts of fatty acid, pyriproxyfen, and potassium salts of fatty acid (C7-C18) treatments. Total handling time (Tht) (\(Tht= Th*Ne\)) was significantly higher for D-limonene, and mineral oil than for all the other treatments tested (Table 1). The total searching time (Tst) (\(Tst=Ts*Ne)\) was significantly lower for D-limonene, and mineral oil (f = 6.617, df = 5, p ≤ 0.05). For the total searching time, no significant difference was observed among control, and potassium salts of fatty acid, pyriproxyfen, and potassium salts of fatty acid (C7-C18) treatments. No predator mortality was observed in any treatment tested during the study observation period.
Discussion
The present study tested the effect of some biorational insecticides on predation potential (functional response) of C. montrouzieri feeding on D. opuntiae females. A previous study by (El Aalaoui et al. 2019c) showed that all the tested insecticide concentrations were effective against D. opuntiae, and at the same time had a minimum impact on adults and larvae of C. montrouzieri (did not cause short-term mortality to the predator). For all the insecticide treatments tested in this study, the predator C. montrouzieri exhibited a type II functional response when feeding on D.opuntiae females. In type II functional response the number of preys consumed by the predator does not obligatory increase with prey density (Hassell et al. 1977), because hungry ladybeetles could be consumed completely the first few preys they encounter and utilize subsequent prey with gradually reduced voracity (Hodek and Honek 1996). Many studies reported that type II response is the most common model type for coccinellids (Collins et al. 1981; Murdoch and Oaten 1975; Seo and DeAngelis 2011). The predation potential of the predator (C. montrouzieri) does not affect significantly by pesticide-treated patches. Exposure of C. montrouzieri to either insecticide treatments was associated with a change in the shape of the curve but not in the functional response. The lower change of the predator functional response curve where insecticides are applied is an indication of a change in predation efficiency, either because the predator attacks a different number of prey or because of a change in searching time (Martinou and Stavrinides 2015). The total time of the functional response devise on two components (searching time (Ts) + handling time (Th)) and an increase in the handling time results in a decrease in the time available for prey searching (Martinou and Stavrinides 2015). All insecticides tested in this study do not affect negatively the searching time (Ts) and handling time (Th) of the predator. In all the treatments tested, the prey handling time by the predator (C. montrouzieri) was relatively higher, because D. opuntiae females cover themselves with white waxy cottony filamentous, therefore the predators spent most of the \(Th\) removing the waxy covering to access the cochineal females bodies. Generally, many factors can affect the functional response of predators such as pesticides (Li et al. 2006; Ambrose et al. 2010; He et al. 2012; Malaquias et al. 2014), intraguild predation (Martinou et al. 2010), and mutual interference (Chong et al. 2006). Pyriproxyfen and potassium salts of fatty acid (C7-C18) seem to be the least compatible with C. montrouzieri as it led to both a significant reduction of attack rate and an increase in handling time compared to other insecticide treatments tested. In contrast, D-limonene, and mineral oil were the most compatible with the predator, as they led to both a significant increase in attack rate and a decrease in handling time compared to other treatments tested. D-limonene and mineral oil are biological insecticides that act by contact on many pests of crops (Taylor and Vickery 1974; Helmy et al. 2012). Indeed D-limonene, which has a physical effect on the cuticle of insects (Taylor and Vickery 1974), surely possess a spectrum of action very broad and not very selective, but the lack of studies does not allow us to conclude as to this assumption (El Aalaoui et al. 2019c). The physical effect of these biological insecticides alters the mechanical barrier provided by the waxy cottony-covered D.opuntiae females and helps the predators to access directly to their prey bodies (El Aalaoui et al. 2019c). Our results showed that D-limonene (60 g/L) applied at 1 g/l, mineral oil (780 g/L) applied at 20 g/L, potassium salts of fatty acid (500 g/l) applied at 0.4 g/L, Pyriproxyfen (100 g/l) applied at 0.25 g/L, and potassium salts of fatty acid (C7-C18) (500 g/L) applied at 3 g/L residues do not have lethal effects on C. montrouzieri, and does not affect significantly his predation potential (consumption). The reduction of attack rate in some insecticide treatments may be due to the sublethal effects of these insecticides. The preference of predator between pesticide-treated and non-treated prey patches could be further tested in choice experiments.
Conclusion
The study of functional response of C. montrouzieri on D. opuntiae females in this current study shows that this predator (C. montrouzieri) could be more effective against this devastating pest of prickly pear cactus in a suitable IPM program. Also, both biological insecticides D-limonene (60 g/L) applied at 1 g/l, mineral oil (780 g/L) applied at 20 g/L could alter the waxy cottony coved the mealybug body and affect positively the predator predation (consumption). It is important to confirm the results obtained in this study under field conditions. This is because many different factors such as the density and diversity of other non-prey species, pesticides, temperature, and other environmental factors and other factors could affect the predator establishment and functional response in field conditions.
References
Agnello A (2002) Petroleum-derived spray oils: chemistry, history, refining and formulation. In: Beattie GAC, Watson DM, Stevens ML, Rae DJ, Spooner-Hart RN (eds) Spray oils beyond 2000. University of Western Sydney, Sydney, pp 2–18
Anjitha A, Krishnamoorthy SV, Kuttalam S (2013) Toxicity of insecticides to the coccinellid predators, Cryptolaemus montrouzieri Mulsant and Scymnus coccivora Ayyar of papaya mealybug, Paracoccus marginatus Williams and Granara de Willink. J Bio Control 27(1):18–23
Ambrose DP, Rajan SJ, Raja JM (2010) Impacts of Synergy-505 on the functional response and behavior of the reduviid bug. Rhynocoris Marginatus j Insect Sci 10:187. https://doi.org/10.1673/031.010.18701
Barbosa PRR, Oliveira MD, Giorgi JA, Silva-Torres CSA, Torres JB (2014) Predatory behavior and life history of Tenuisvalvae notata (coleoptera: coccinellidae) under variable prey availability conditions. Fla Entomol 97:1026–1034. https://doi.org/10.1653/024.097.0304
Bouharroud R, Sbaghi M, Boujghagh M, El Bouhssini M (2018) Biological control of the prickly pear cochineal Dactylopius opuntiae Cockerell (Hemiptera: Dactylopiidae). EPPO Bulletin 48:300–306. https://doi.org/10.1111/epp.12471
Brennan TCR, Kromer JO, Nielsen LK (2013) Physiological and transcriptional responses of Saccharomyces cerevisiae to d-limonene show changes to the cell wall but not to the plasma membrane. Appl Environ Microbiol 79:3590–3600. https://doi.org/10.1128/AEM.00463-13
Candolfi MP, Barrett KL, Campbell P, Forster R, Grandy N, Huet MC (2001) Guidance document on regulatory testing and risk assessment procedures for plant protection products with non-target arthropods. European Standard Characteristics of Regulatory Testing (ESCORT 2) Workshop. Wageningen, The Netherlands: SETAC Europe, pp. 21–23.
Castro RM (2011) Biologia e exigências térmicas de Zagreus bimaculosus (Mulsant) (Coleoptera: Coccinellidae). Recife: UFRPE (Dissertação de Mestrado).
Carver RH, Nash JG (2011) Doing data analysis with SPSS: version 18.0. Cengage Learning (U.S).
Chong JH, Oetting RD (2006) Functional response and progeny production of the Madeira mealybug parasitoid, Anagyrus sp. nov. nr. sinope: the effects of host and parasitoid densities. Biol Control 39:320–328. https://doi.org/10.1016/j.biocontrol.2006.08.013
Collins MD, Wards S, Dixon AFG (1981) Handling time and the functional response of Aphelinus thomsoni, a predator and parasite of the aphid Drepanosiphum platanoides. J Anim Ecol 50:479–489. https://doi.org/10.2307/4069
Cranshaw WS, Baxendale B (2011) Insect Control: Horticultural Oils. Colorado State Univirsity, http://www.ext.colostate.edu/pubs/insect/05569.html
El Aalaoui M, Bouharroud R, Sbaghi M, El Bouhssini M, Hilali L (2019a) Predatory potential of eleven native Moroccan adult ladybird species on different stages of Dactylopius opuntiae (Cockerell) (Hemiptera: Dactylopiidae). EPPO Bulletin 49(2):374–379. https://doi.org/10.1111/epp.12565
El-Aalaoui M, Bouharroud R, Sbaghi M, El Bouhssini M, Hilali L (2019b) First study of the biology of Cryptolaemus montrouzieri and its potential to feed on the mealybug Dactylopius opuntiae (Hemiptera: Dactylopiidae) under laboratory conditions in Morocco. Arch Phytopathology Plant Protect 52(13–14):1112–1124. https://doi.org/10.1080/03235408.2019.1691904
El Aalaoui M, Bouharroud R, Sbaghi M, El Bouhssini M, Hilali L, Dari K (2019c) Comparative toxicity of different chemical and biological insecticides against the scale insect Dactylopius opuntiae and their side effects on the predator Cryptolaemus montrouzieri. Arch Phytopathology Plant Protect 52(1–2):155–169. https://doi.org/10.1080/03235408.2019.1589909
El Aalaoui M, Sbaghi M, Bouharroud R, El Bouhssini M, Hilali L (2020) Hyperpredation of local adults ladybirds on the eggs of Cryptolaemus montrouzieri a potential predator of carmine cactus cochineal Dactylopius opuntiae in Morocco Int J Trop Insect Sci 1–6 https://doi.org/10.1007/s42690-020-00282-w
El Aalaoui M, El Bouhssini M, Bouharroud R, Sbaghi M (2021) Lethal and sublethal effects of the insecticides d-limonene, mineral oil, and potassium salts of fatty acid on Dactylopius Opuntiae potential predator Cryptolaemus Montrouzieri Int J Trop Insect Sci 1–10 https://doi.org/10.1007/s42690-021-00473-z
Elliott JM (2003) A comparative study of the functional response of four species of carnivorous stoneflies. Freshw Biol 48(2):191–202. https://doi.org/10.1046/j.1365-2427.2003.00982.x
Fernandez DE, Beers EH, Brunner JF, Doerr ME, Dunley JE (2001) Mineral oil inhibition of white apple leafhopper (Homoptera: Cicadellidae) oviposition. J Entomol Sci 36(3): 237–243. https://doi.org/10.18474/0749-8004-36.3.237
Fernández-Arhex V, Corley JC (2004) La respuesta funcional: una revisión y guía experimental. Ecol Austral 14(1):83–93
Flores A, Olvera H, Rodríguez S, Barranco J (2013) Predation potential of Chilocorus cacti (Coleoptera: Coccinellidae) to the prickly pear cacti pest Dactylopius opuntiae (Hemiptera: Dactylopiidae). Neotrop Entomol 42(4):407–411. https://doi.org/10.1007/s13744-013-0139-z
Giorgi JA, Barbosa PRR, Oliveira JEM, Torres JB (2017) Prodiloides bipunctata Weise (Coccinellidae: Cephaloscymnini): new research on native natural predators of the carmine cochineal, Dactylopius opuntiae (Cockerel) (Hemiptera: Dactylopiidae) in the Brazilian Semi-arid. The Coleopterist Bulletin (in press).
Halappa B, Awaknavar JS, Archana D, Bandi S, Kumar GSA (2013) Laboratory evaluation of insecticides against Australian beetle, Cryptolaemus montrouzieri Mulsant (Coccinellidae: Coleoptera). Current Biotica 7(3):196–201
Hassell MP, Lawton JH, Beddington JR (1977) Sigmoid functional responses by invertebrate predators and parasitoids J Anim Ecol 249–262 https://doi.org/10.2307/3959
Helmy EI, Kwaiz FA, El-Sahn OMN (2012) The usage of mineral oils to control insects. Egypt Acad J Biol Sci : Entomol 5(3):167–174
He Y, Zhao J, Zheng Y, Desneux N, Wu K (2012) Lethal effect of imidacloprid on the coccinellid predator Serangium japonicum and sublethal effects on predator voracity and on functional response to the whitefly Bemisia tabaci. Ecotoxicol 21(5):1291–1300. https://doi.org/10.1007/s10646-012-0883-6
Hodek I, Honek A (1996) Ecology of Coccinellidae. Kluwer Academic Publishers, Dordrecht, Boston, London, p 464
Holling CS (1959) Some characteristics of simple types of predation and parasitism. Can Entomol 91(7):385–398
Juliano SA (2001) Non-linear curve fitting: predation and functional response curves. In: Scheiner SM, Gurevitch J (eds) Design and analysis of ecological experiments, 2nd edn. Chapman and Hall, New York, pp 178–196
Juliano SA, Williams FM (1987) A comparison of methods for estimating the functional response parameters of the random predator equation. J Anim Ecol 52(2):641–653. https://doi.org/10.2307/5074
Karr LL (1989) Toxic properties of d-limonene in insects and the earthworm Eisenia fetida. Dissertations, University of Iowa, Ames, IA (US).
Khan HAA, Sayyed AH, Akram W, Razald S, Ali M (2012) Predatory potential of Chrysoperla carnea and Cryptolaemus montrouzieri larvae on different stages of the mealybug, Phenacoccus solenopsis: A threat to cotton in South Asia. J Insect Sci 12(1):147. https://doi.org/10.1673/031.012.14701
Kim M, Shin D, Suh E, Cho K (2004) An assessment of the chronic toxicity of fenpyroximate and pyridaben to Tetranychus urticae using a demographic bioassay. Appl Entomol Zool 39(3):401–409. https://doi.org/10.1303/aez.2004.401
Li DX, Tian J, Shen ZR (2006) Effects of pesticides on the functional response of predatory thrips, Scolothrips takahashii to Tetranychus viennensis. J Appl Entomol 130(5):314–322. https://doi.org/10.1111/j.1439-0418.2006.01056.x
Lima MS, da Silva DMP, Falcão HM, Ferreira WM, Silva LD, Paranhos BAJ (2011) Predators of the Dactylopius opuntiae (Hemiptera: Dactylopiidae) on fodder opuntia in the Pernambuco State, Brazil. Rev Chil Entomol 36:51–54
Malaquias JB, Ramalho FS, Omoto C, Godoy WAC, Silveira RF (2014) Imidacloprid affects the functional response of predator Podisus nigrispinus (Dallas)(Heteroptera: Pentatomidae) to strains of Spodoptera frugiperda (JE Smith) on Bt cotton. Ecotoxicol 23(2):192–200. https://doi.org/10.1007/s10646-013-1162-x
Martinou AF, Raymond B, Milonas PG, Wright DJ (2010) Impact of intraguild predation on parasitoid foraging behaviour. Ecol Entomol 35(2):183–189. https://doi.org/10.1111/j.1365-2311.2009.01171.x
Martinou AF, Seraphides N, Stavrinides MC (2014) Lethal and behavioral effects of pesticides on the insect predator Macrolophus pygmaeus. Chemosphere 96:167–173. https://doi.org/10.1016/j.chemosphere.2013.10.024
Martinou AF, Stavrinides MC (2015) Effects of sublethal concentrations of insecticides on the functional response of two mirid generalist predators. PLoS ONE 10(12):e0144413. https://doi.org/10.1371/journal.pone.0144413
Murdoch WW, Briggs CJ, Nisbet RM (2003) Consumer-resource dynamics. Princeton University Press, Princeton, NJ
Murdoch WW, Oaten A (1975) Predation and population stability. Adv Ecol Res 9:1–131. https://doi.org/10.1016/S0065-2504(08)60288-3
Murray GP, Stillman RA, Gozlan RE, Britton JR (2013) Experimental predictions of the functional response of a freshwater fish. Ethol 119(9):751–761. https://doi.org/10.1111/eth.12117
Nawaz M, Cai W, Jing Z, Zhou X, Mabubu JI, Hua H (2017) Toxicity and sublethal effects of chlorantraniliprole on the development and fecundity of a non-specific predator, the multicolored Asian lady beetle, Harmonia axyridis (Pallas). Chemosphere 178:496–503. https://doi.org/10.1016/j.chemosphere.2017.03.082
Osborne LS, Petitt FL (1985) Insecticidal soap and the predatory mite, Phytoseiulus persimilis (Acari: Phytoseiidae), used in management of the twospotted spider mite (Acari: Tetranychidae) on greenhouse grown foliage plants. J Econ Entomol 78(3):687–691. https://doi.org/10.1093/jee/78.3.687
Pasini C, D'Aquila F, Curir P, Gullino ML (1997) Effectiveness of antifungal compounds against rose powdery mildew (Sphaerotheca pannosa var. rosae) in glasshouses. Crop Prot 16(3): 251–256. https://doi.org/https://doi.org/10.1016/S0261-2194(96)00095-6
Pervez A, Omkar, (2005) Functional responses of coccinellid predators: an illustration of a logistic approach. J Insect Sci 5(5):1–6. https://doi.org/10.1093/jis/5.1.5
Planes L, Catalán J, Tena A, Porcuna JL, Jacas JA, Izquierdo J, Urbaneja A (2013) Lethal and sublethal effects of spirotetramat on the mealybug destroyer. Cryptolaemus Montrouzieri J Pest Sci 86(2):321–327. https://doi.org/10.1007/s10340-012-0440-3
Prats D, Rodrguez M, Varo P, Moreno A, Ferrer J, Berna JL (1999) Biodegradation of soap in anaerobic digesters and on sludge amended soils. Water Res 33(1):105–108. https://doi.org/10.1016/S0043-1354(98)00199-7
Protasov A, Mendel Z, Spodek M, Carvalho CJ (2017) Management of the Opuntia cochineal scale insect, Dactylopius opuntiae (Cockerell) in Israel. Alon Hanotea 71:48–51
Riedl H, Halaj J, Kreowski WB, Hilton RJ, Westigard PH (1995) Laboratory evaluation of mineral oils for control of codling moth (Lepidoptera: Tortricidae). J Econ Entomol 88(1):140–147. https://doi.org/10.1093/jee/88.1.140
Rill SM, Grafton-Cardwell EE, Morse JG (2008) Effects of two insect growth regulators and a neonicotinoid on various life stages of Aphytis melinus (Hymenoptera: Aphelinidae). Biocontrol 53(4):579–587. https://doi.org/10.1007/s10526-007-9097-x
Rimoldi F, Schneider MI, Ronco AE (2008) Susceptibility of Chrysoperla externa eggs (Neuroptera: Chrisopidae) to conventional and biorational insecticides. Environ Entomol 37(5):1252–1257. https://doi.org/10.1093/ee/37.5.1252
Rimoldi F, Fogel MN, Ronco AE, Schneider MI (2017) Comparative susceptibility of two Neotropical predators, Eriopis connexa and Chrysoperla externa, to acetamiprid and pyriproxyfen: short and long-term effects after egg exposure. Environ Pollut 231:1042–1050. https://doi.org/10.1016/j.envpol.2017.08.097
Rogers D (1972) Random search and insect population models. J Anim Ecol 41:369–383. https://doi.org/10.2307/3474
Sanches NF, Carvalho RS (2010) Procedimentos para manejo da criação e multiplicação do predador exótico Cryptolaemus montrouzieri. Cruz dasAlmas: BrasilEmbrapa Mandioca e Fruticultura (Circular Técnica, 99).
Seo G, DeAngelis DL (2011) A predator–prey model with a Holling type I functional response including a predator mutual interference. J Nonlinear Sci 21(6):811–833. https://doi.org/10.1007/s00332-011-9101-6
Smith EH, Pearce GW (1948) The mode of action of petroleum oils as ovicides. J Econ Entomol 41(2):173–180. https://doi.org/10.1093/jee/41.2.173
Solomon ME (1949) The natural control of animal populations. J Anim Ecol 18:1–35. https://doi.org/10.2307/1578
Sullivan JJ, Goh KS (2008) Environmental fate and properties of pyriproxyfen. J Pestic Sci 33(4):339–350. https://doi.org/10.1584/jpestics.R08-02
Taylor WE, Vickery B (1974) Insecticidal properties of limonene, a constituent of citrus oil. Ghana J Agric Sci 7:61–62
Trexler JC, McCulloch CE, Travis J (1988) How can the functional response best be determined? Oecologia 76(2):206–214
Tsolakis H, Ragusa S (2008) Effects of a mixture of vegetable and essential oils and fatty acid potassium salts on Tetranychus urticae and Phytoseiulus persimilis. Ecotoxicol Environ Saf 70(2):276–282. https://doi.org/10.1016/j.ecoenv.2007.10.001
Vanegas-Rico JM, Lomeli-Flores JR, Rodríguez-Leyva E, Mora-Aguilera G, Valdez JM (2010) Enemigos naturales de Dactylopius opuntiae (Cockerell) en Opuntia ficus-indica (L.) Miller en el centro de México. Acta Zool Mex 26(2): 415–433.
Wanumen AC, Sánchez-Ramos I, Viñuela E, Medina P, Adán Á (2016) Impact of feeding on contaminated prey on the life parameters of Nesidiocoris tenuis (Hemiptera: Miridae) adults. J Insect Sci 16(1). https://doi.org/10.1093/jisesa/iew084
Wiedenmann RN, O’Neil RJ (1991) Laboratory measurement of the functional response of Podisus maculiventris (Say) (Heteroptera: Pentatomidae). Environ Entomol 20(2):610–614. https://doi.org/10.1093/ee/20.2.610
Wins-Purdy AH, Whitehouse C, Judd GJR, Evenden ML (2009) Effect of horticultural oil on oviposition behavior and egg survival in the obliquebanded leafroller (Lepidoptera: Tortricidae). Can Entomol 141(1):86–94. https://doi.org/10.4039/n08-042
Xiao D, Zhao J, Guo X, Chen H, Qu M, Zhai W, Desneux N, Biondi A, Zhang F, Wang S (2016) Sublethal effects of imidacloprid on the predatory seven-spot ladybird beetle Coccinella septempunctata. Ecotoxicol 25(10):1782–1793. https://doi.org/10.1007/s10646-016-1721-z
Zwick RW, Westigard PH (1978) Prebloom petroleum oil applications for delaying pear psylla (Homoptera: Psyllidae) oviposition. Can. Entomol 110(3): 225–236. https://doi.org/10.4039/Ent110225-3
Funding
The research of this study was supported by the National Institute of Agronomic Research (INRA), Morocco.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interests.
Ethical approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Aalaoui, M.E., Sbaghi, M. Effects of sublethal concentrations of some biorational insecticides in predation potential of Cryptolaemus montrouzieri on Dactylopius opuntiae. Int J Trop Insect Sci 42, 519–526 (2022). https://doi.org/10.1007/s42690-021-00567-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42690-021-00567-8