
Abstract
Resurgence is the reemergence of a previously reinforced behavior following the extinction of an alternative behavior. Although several variables are known to impact resurgence, few studies have examined resurgence when the target response was previously punished. Results of two studies examining resurgence after punishment have been mixed. In the current study, we employed a human-operant procedure to compare resurgence following differential reinforcement of alternative behavior (DRA) with extinction and DRA with extinction and response cost. For the three participants, target responding was eliminated more rapidly and fully following DRA with response cost. Similar levels of resurgence, however, occurred following both procedures. These data provide evidence that arranging response cost during DRA facilitates suppression of target responding without affecting subsequent resurgence. The results further our understanding of the role of negative punishment in resurgence as a process and have implications for the development of clinical intervention packages.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Resurgence refers to the temporary reemergence of a previously reinforced behavior following the extinction of an alternative behavior. Over 50 years of studies have established resurgence as a robust phenomenon that occurs across species and has been replicated with non-humans and humans (see Lattal & St. Peter Pipkin, 2009, for a review). Most evaluations of resurgence involve a three-phase procedure (Lattal & St. Peter Pipkin, 2009). During the first phase, a target behavior is reinforced. During the second phase, the target behavior is placed on extinction, and an alternative behavior is reinforced. In the third phase, some worsening of reinforcement conditions occurs; most often, this worsening consists of both responses being placed on extinction. Resurgence is the transient recurrence of the target response above recent response rates (see Lattal et al., 2017, for an extended discussion of the definition of resurgence).
Resurgence has recently gained attention because the three-phase procedure parallels common behavioral treatments. One such treatment is called differential reinforcement of an alternative response (DRA). DRA is one of the most common treatments for suppressing undesired behavior (Petscher, Rey, & Bailey, 2009). The goal of DRA is to replace undesired target behavior with an alternative response. During DRA, the previously reinforced problem behavior is put on extinction, and a more desirable replacement response is reinforced. One example of a replacement response is a communication response to appropriately request the same reinforcer that was previously maintaining the target behavior (i.e., functional communication training; Carr & Durand, 1985). Previous research has shown that if the DRA treatment is subsequently challenged, recurrence of the problem behavior may occur (e.g., St. Peter, 2015; Volkert, Lerman, Call, & Trosclair-Lasserre, 2009). The resurgence effects seen in clinical populations are similar to demonstrations of resurgence in the nonhuman animal literature (e.g., Lieving & Lattal, 2003).
Because of these parallels, several contemporary resurgence studies have modified aspects of DRA treatments to reduce resurgence during subsequent treatment disruptions (e.g., Fisher, Greer, Fuhrman, Saini, & Simmons, in press; Lambert, Bloom, Samaha, Dayton, & Rodewald, 2015; Romano & St. Peter, 2016). For example, Fisher et al. (in press) used behavioral momentum theory (Nevin & Shahan, 2011) to predict that decreasing reinforcement rate during baseline and treatment, as well as increasing the duration of treatment, would reduce subsequent resurgence. Results matched the predictions for all four participants, who were children with intellectual disabilities who engaged in problem behavior.
Although effects of reinforcement parameters on subsequent resurgence are becoming more widely known (and have resulted in quantitative models of the phenomenon; Shahan & Craig, 2016), less is known about effects of punishment on resurgence. Understanding the role of punishment may be important for the translation of resurgence research to behavior analytic treatment because mild punishment procedures, like reprimands or response cost, are commonly used as intervention components (Conyers et al., 2004; Falcomata, Roane, Hovanetz, Kettering, & Keeney, 2004). Despite the potential importance of understanding the role of punishment on resurgence, only two studies to date have directly examined resurgence following punishment. Kestner, Redner, Watkins, and Poling (2015) evaluated impacts of positive punishment on resurgence using a group design with rats. They implemented DRA alone (control group) compared to DRA in combination with mild electric shock (experimental group) to suppress rates of target responding during the second phase of a resurgence procedure. Both groups of rats were exposed to extinction for all responses in the third phase. Markedly less resurgence occurred for the group that previously experienced punishment than the no-punishment control group, suggesting that including positive punishment procedures in conjunction with DRA may attenuate resurgence. Although these results are promising in regards to the inclusion of punishment procedures as part of treatment packages, positive punishment procedures may be less acceptable in practice than are negative punishment procedures (Kazdin, 1980).
Response cost is a negative punishment procedure that is commonly used to reduce challenging behavior. Response cost involves the removal of a reinforcer contingent on a target response (Lerman & Toole, 2011). Response cost procedures have been shown to effectively suppress problem behavior in treatment contexts (e.g., Kahng, Tarbox, & Wilke, 2001; Keeney, Fisher, Adelinis, & Wilder, 2000; Conyers et al., 2004). Multicomponent treatment packages including DRA and punishment may be more efficient than DRA alone at suppressing behavior (Fisher et al., 1993; Hangopian, Fisher, Sullivan, Aquisto, & LeBlanc, 1998). The effects of response cost on future resurgence, however, are less known.
Okouchi (2015) evaluated resurgence following DRA with response cost and DRA in isolation, using a group design with human participants. Participants engaged in arbitrary responses (two-response sequences of touching squares on a computer screen) with points exchangeable for money serving as reinforcers. Two groups experienced response-dependent reinforcement for a particular response sequence during the first phase. Although particular response sequences were individualized for each participant, the contingencies for participants were identical (a terminal variable-interval [VI] 10-s schedule) across groups. During the second phase, previously reinforced (target) sequences were eliminated through a combination of reinforcement of an alternative and punishment of target responding (DRA + RC group) or reinforcement of an alternative only (DRA-only group). During the third phase, the target sequences did not produce points for either group (extinction). Resurgence occurred for both groups, and no statistically significant differences in resurgence were obtained. The results obtained by Okouchi (2015) differ from those obtained by Kestner et al. (2015). Clarifying the role of negative punishment on subsequent resurgence would further enhance our understanding of resurgence as a process as well as inform the development of clinical intervention packages aimed at reducing resurgence during treatment challenges.
The present study was a systematic replication of Okouchi (2015) and aimed to evaluate resurgence following DRA alone (DRA) and DRA with response cost (DRA + RC). There were three main procedural differences between our study and Okouchi (2015). First, we used a simpler DRA-training phase by providing two discrete response options. Second, participants in the current study earned time-based extra credit, independent of their performance, rather than exchanging points for money or other backup reinforcers. Finally, we used a multiple-schedule arrangement to conduct a within-subject comparison of resurgence following DRA and DRA + RC.
Method
Participants and Setting
Three undergraduate students enrolled in lower-division psychology courses participated. Participant A was a 19-year-old Caucasian female. Participant B was a 21-year-old Caucasian male. Participant C was a 20-year-old Caucasian female. Participants received course credit based on the duration of their participation. Sessions took place in a 4.1-m × 3-m laboratory equipped with chairs, two tables, a computer desk, a Dell® Intel desktop computer with a 12-cm × 15-cm screen, and a mouse. We stowed the keyboard in the desk during sessions. Participants sat facing away from a one-way mirror through which we monitored the sessions. Participants completed the experiment in a single appointment consisting of two 60-min blocks separated by a 5- to 10-min break.
Procedure
We provided scripted instructions to the participant at the beginning of the appointment (see Supplemental Materials). During the consent process, we informed the participant that the study involved responding under different contingencies of reinforcement without providing further detail. Before starting the session, we asked the participants leave all personal items, electronic devices, and watches outside of the room to prevent overt timing of the schedules or components. We instructed the participant to use only the mouse to earn as many points as possible.
A custom Visual Basic® program delivered the experimental task. This program was developed in the third author’s research laboratory and has been used in past studies on treatment challenges (e.g., Marsteller & St. Peter, 2012; St. Peter Pipkin, Vollmer, & Sloman, 2010). The computer screen displayed two moving circles (one black and one white). Each circle was 25.4 mm in diameter and moved around the screen at a speed of 25 mm/s. The circles moved randomly and independently across the screen until they reached the side or corner of the screen, at which point they changed direction. The program delivered or removed points according to the reinforcement or punishment schedule in effect. There was a cumulative point counter on the bottom left corner of the screen, which flashed orange briefly when a point was earned or lost. Points were not exchangeable for backup reinforcers. Clicking the black circle was arbitrarily selected as the target response, and clicking the white circle was selected as the alternative response. After the first three-phase sequence, the participant left the laboratory for a 5- to 10-min break. We reset the program, and the participant then completed a replication of the three phases. At the end of the study, we provided a scripted debriefing (see Supplemental Materials).
We used a multielement design embedded in a reversal design to assess patterns of resurgence following DRA without response cost (DRA) and DRA with response cost (DRA + RC). Each 60-min block included a three-phase sequence. Each phase was 20 min in duration and included two components. The program strictly alternated between components and presented each 2-min component five times during each phase. The initial component was counterbalanced across participants and across replications for each participant. During all phases, the program displayed a blue background during the components associated with DRA during phase 2. The program displayed a red background during components associated with DRA + RC. There were no programmed stimulus changes to signal the beginning of a new phase.
Table 1 displays a summary of the component conditions during each phase. During baseline (phase 1), both components were identical except for background color. Clicks on the black circle resulted in the delivery of one point on a random interval (RI) 5-s schedule, which had a minimum value of 1 and maximum value of 10. Clicks on the white circle resulted in no points. During the alternative reinforcement phase (phase 2), one component consisted of DRA and the other of DRA + RC. During both components, clicks on the white circle resulted in the delivery of one point on an RI 5-s schedule (arranged as in baseline). During the DRA components, clicking on the black circle resulted in no programmed consequences. During the DRA + RC components, clicks on the black circle resulted in the loss of one point on a fixed-ratio 1 schedule. During extinction (phase 3), components were identical except for background color, and there were no programmed consequences for responding on either circle.
Data Analysis
We used visual inspection of graphed data to evaluate responding across components and phases. Of particular interest was detecting resurgence during the extinction phases. We defined resurgence as the rate of black-circle clicks in any extinction component exceeding the rate observed during the last three component presentations in the alternative reinforcement phase (Marsteller & St. Peter, 2012).
Results
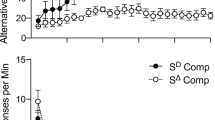
Table 2 shows obtained reinforcement rates (mean reinforcers per min) for each phase and component. The obtained reinforcement rates were similar between participants and between components for each participant. Differences between components were no more than 1 reinforcer per minute and did not vary systematically. Figures 1 and 2, respectively, show response rates for black-circle clicks (target response) and white-circle clicks (alternative response) for each participant. Across phases, participants allocated more responses to the circle that resulted in point deliveries, which supports the notion that points functioned as reinforcers.

Black-circle clicks per min during both component presentations (DRA and DRA + RC) of each phase for all three participants. Note the difference in scaling on the y-axes

White circle clicks per minute during both component presentations (DRA and DRA + RC) of each phase for all three participants. Note the difference in scaling on the y-axes
Figure 1 depicts the black-circle clicks (target response) per minute during each component presentation and phase for all three participants. During baseline (labeled “BL” on the graph), differences in responding across DRA (black symbols) and DRA + RC (white symbols) were small and unsystematic. During the alternative reinforcement phase (labeled “ALT” on the graph), black-circle clicks decreased and both DRA and DRA + RC reduced responding to zero or near-zero levels. Responding was suppressed more rapidly during the component associated with DRA + RC than with DRA. The exception to this pattern was observed during the first treatment phase for participant A, who engaged in a high rate of responding during the first DRA component presentation. This was followed by zero or near-zero levels of responding during all other treatment component presentations for this participant. During the extinction phases (labeled “EXT”), resurgence of the target response occurred during both components for all three participants. Table 3 displays the total number of black-circle responses during each component in the extinction phases. The observed levels of resurgence were relatively similar between components previously associated with DRA and those previously associated with DRA + RC. Participants A and C engaged in slightly more resurgence responses (27 and 19 more black-circle clicks, respectively) during the component previously associated with DRA + RC than the component previously associated with DRA. Participant B engaged in marginally more resurgence responses (28 more black-circle responses) during the component previously associated with DRA. Arranging response cost in conjunction with DRA did not attenuate resurgence.
Figure 2 depicts the white-circle clicks (alternative responses) per minute during each component presentation and phase for all three participants. During the baseline phases, participants A and B engaged in low levels of responding on the white circles, and participant C engaged in moderate levels of responding on the white circle. Responding on the white circle increased for the participants when it was reinforced during the alternative reinforcement phases. During the extinction phases, white-circle responses quickly returned to baseline levels for participant B. Participant C continued to click the white circle but at a lower rate compared to the alternative reinforcement phase. Participant A continued to click the white circle, with higher rates occurring during the first few component presentations and then decreasing toward the end of the phase.
Discussion
We compared resurgence following DRA with or without response cost, using a human-operant preparation. Although target responding was more rapidly and fully suppressed during DRA with response cost than during DRA in isolation, no differences in subsequent resurgence occurred. Thus, our findings are similar to those from the study by Okouchi (2015), which failed to show a significant difference in resurgence when responding was eliminated via DRA with either extinction alone or extinction and response cost for the target response. Both studies showed quicker suppression of target responding with DRA + RC.
Although our findings are similar to those obtained by Okouchi (2015), our procedures differed in several ways. First, Okouchi evaluated resurgence of a complex response (two-response sequences), which necessitated a more complex DRA-training phase. To shape the response sequences, Okouchi first reinforced target sequences on a fixed-ratio 1 schedule, followed by a VI 2-s schedule, before transitioning to the terminal VI 10-s schedule. In our study, only two-response options were explicitly available to participants (although participants sometimes clicked the computer screen in areas other than the circles or engaged in unmeasured behavior like playing with their hair). This allowed us to more fully control exposure to reinforcers and approximately equate such reinforcer access across participants. Second, participants in the study by Okouchi were paid for their performance. Although our participants earned extra credit in a course for the time they spent participating (independent of performance), all participants acquired target and alternative responses and resurgence occurred reliably for all participants. Computer points functioned as reinforcers in both studies, regardless of whether backup reinforcers were available. Finally, Okouchi evaluated resurgence following different histories using a group design. In our study, we used a multiple schedule to conduct within-subject evaluations of resurgence. The similarity in our findings, despite these differences in experimental preparation, add to the already robust literature suggesting that resurgence is a reliable phenomenon in humans that occurs across a wide array of experimental parameters.
Our findings differed from those obtained by Kestner et al. (2015), who found that inclusion of a positive punishment component during the second phase reduced subsequent resurgence. One obvious difference across the studies evaluating resurgence following punishment is the use of positive or negative punishment. To date, no studies have included a direct comparison of resurgence following response suppression with positive and negative punishment procedures. Such an evaluation may help to inform the extent to which variations on punishment may impact resurgence. At least two other factors may also impact the failure to replicate across studies. First, Kestner et al. used a biologically relevant punisher (electric shock), whereas Okouchi (2015) and the present study used relatively innocuous punishers (point loss). There may be differences in the behavioral effects depending on the salience or biological relevance of a punisher (Crosbie, 1998), and it is currently unknown whether there are differences related to the reduction of subsequent resurgence. Future research should evaluate the type or intensity of punishing stimuli used in the second phase on subsequent resurgence. Such research may have important implications not only for development of effective interventions but also for our understanding of the underlying mechanisms that impact resurgence (such as the resurgence-as-choice model; Shahan & Craig, 2016). Second, the species differed across evaluations. Recall that Kestner et al. evaluated resurgence with rats, but both Okouchi and the present study included human participants. Although resurgence has shown to be a reliable phenomenon across species (and occurred in all three of the studies cited above), it remains possible that variables impacting resurgence may differ to some extent across species. For example, it is possible that the human participants in Okouchi’s study and the present study developed self-generated rules that contributed to equivalent resurgence during the extinction phase (see Dixon & Hayes, 1998 for an example of the effects of verbal rules on resurgence). Future research could explicitly manipulate rule-governed behavior to evaluate the potential impact on subsequent resurgence.
We used an arbitrary response maintained by conditioned reinforcers with a non-clinical population. Although human-operant procedures are beneficial in translating research from the basic laboratory to humans, their generality to other responses, reinforcers, and populations is unclear until the results are replicated. Many negative punishment procedures in clinical contexts involve the use of time out from reinforcement (Lerman & Vorndran, 2002), whereas our procedure involved the removal of a generalized conditioned reinforcer. Differences in the effort of the responses, density and type of reinforcers, and type of response cost used may limit the generality of our findings to clinical contexts. Future studies should evaluate resurgence of socially significant behavior when punishment involves time out rather than point loss.
Future research on resurgence could replicate these evaluations of punishment and resurgence using different research designs. In the current study, the baseline and extinction phases consisted of two identical component schedules, and similar responding was observed between components during the extinction phases. One possibility is that similar response patterns during extinction may be indicative that adding response cost has no effect on resurgence. Alternatively, it is difficult to rule out a lack of stimulus control between components as a contributing to the similar patterns. Replicating these results with additional designs (e.g., ABCADC) would strengthen the evidence for the current conclusions.
We anticipated that our study might inform our understanding of resurgence as a process as well as the development of effective treatment packages that reduce the likelihood of subsequent relapse during treatment challenges. Our results indicate that punishment may not be an important variable to consider when primarily concerned about relapse, but inclusion of response cost procedures as part of a treatment package may be warranted when rapid suppression of target behavior is necessary during initial treatment. Notably, although including response cost seems unlikely to reduce subsequent resurgence, it also seems unlikely to increase resurgence if the intervention is abruptly discontinued. Thus, clinicians should continue to include response cost components in intervention packages when initial rapid response suppression is a priority.
References
Carr, E. G., & Durand, V. M. (1985). Reducing behavior problems through functional communication training. Journal of Applied Behavior Analysis, 18(2), 1307999–1307126. https://doi.org/10.1901/jaba.1985.18-111.
Conyers, C., Miltenberger, R., Maki, A., Barenz, R., Jurgens, M., Sailer, A., Haugen, M., & Kopp, B. (2004). A comparison of response cost and differential reinforcement of other behavior to reduce disruptive behavior in a preschool classroom. Journal of Applied Behavior Analysis, 37, 411–415. https://doi.org/10.1901/jaba.2004.37-411.
Crosbie, J. (1998). Negative reinforcement and punishment. In K. A. Lattal & M. Perone (Eds.), Handbook of Research Methods in Human Operant Behavior (pp. 163–1890). New York, NY: Plenum Press.
Dixon, M. R., & Hayes, L. J. (1998). Effects of differing instructional histories on the resurgence of rule-following. Psychological Record, 48(2), 275–292.
Falcomata, T. S., Roane, H. S., Hovanetz, A. N., Kettering, T. L., & Keeney, K. M. (2004). An evaluation of response cost in the treatment of inappropriate vocalizations maintained by automatic reinforcement. Journal of Applied Behavior Analysis, 37, 83–87. https://doi.org/10.1901/jaba.2004.37-83.
Fisher, W., Piazza, C., Cataldo, M., Harrell, R., Jefferson, G., & Conner, R. (1993). Functional communication training with and without extinction and punishment. Journal of Applied Behavior Analysis, 26(1), 23–36. https://doi.org/10.1901/jaba.1993.26-23.
Fisher, W. W., Greer, B. D., Fuhrman, A. M., Saini, V., & Simmons, C. A. (in press). Minimizing resurgence of destructive behavior using behavioral momentum theory. Journal of Applied Behavior Analysis.
Hagopian, L.P., Fisher, W.W., Sullivan, M.T., Aquisto, J., LeBlanc, L.A. (1998). Effectiveness of functional communication training with and without punishment: a summary of 21 inpatient cases. Journal of Applied Behaviour Analysis, 31, 211–235. https://doi.org/10.1901/jaba.1998.31-211.
Kahng, S., Tarbox, J., & Wilke, A. E. (2001). Use of a multicomponent treatment for food refusal. Journal of Applied Behavior Analysis, 34, 93–96. https://doi.org/10.1901/jaba.2001.34-93.
Kazdin, A. E. (1980). Acceptability of alternative treatments for deviant child behavior. Journal of Applied Behavior Analysis, 13, 259–273.
Keeney, K. M., Fisher, W. W., Adelinis, J. D., & Wilder, D. A. (2000). The effects of response cost in the treatment of aberrant behavior maintained by negative reinforcement. Journal of Applied Behavior Analysis, 33, 255–258. https://doi.org/10.1901/jaba.2000.33-255.
Kestner, K., Redner, R., Watkins, E. E., & Poling, A. (2015). The effects of punishment on resurgence in laboratory rats. The Psychological Record, 65, 315–321. https://doi.org/10.1007/s40732-014-0107-y.
Lambert, J. M., Bloom, S. E., Samaha, A. L., Dayton, E., & Rodewald, A. M. (2015). Serial alternative response training as intervention for target response resurgence. Journal of Applied Behavior Analysis, 48, 765–780. https://doi.org/10.1002/jaba.253.
Lattal, K. A., & St. Peter Pipkin, C. (2009). Resurgence of previously reinforced responding: Research & application. The Behavior Analyst Today, 10, 254–265.
Lattal, K. A., Cançado, C. R., Cook, J. E., Kincaid, S. L., Nighbor, T. D., & Oliver, A. C. (2017). On defining resurgence. Behavioural Processes, 141, 85–91. https://doi.org/10.1016/j.beproc.2017.04.018.
Lerman, D. C., & Toole, L. M. (2011). Developing function-based punishment procedures for problem behavior. In W. W. Fisher, C. C. Piazza, & H. S. Roane (Eds.), Handbook of Applied Behavior Analysis (pp. 348–369). New York, NY: Guilford Press.
Lerman, D. C., & Vorndran, C. M. (2002). On the status of knowledge for using punishment: Implications for treating behavior disorders. Journal of Applied Behavior Analysis, 35, 431–464. https://doi.org/10.1901/jaba.2002.35-431.
Lieving, G. A., & Lattal, K. A. (2003). Recency, repeatability, and reinforcer retrenchment: An experimental analysis of resurgence. Journal of the Experimental Analysis of Behavior, 80(2), 217–233. https://doi.org/10.1901/jeab.2003.80-217.
Marsteller, T. M., & St. Peter, C. C. (2012). Resurgence during treatment challenges. Revista Mexicana de Análisis de la Conducta, 38, 7–23.
Nevin, J. A., & Shahan, T. A. (2011). Behavioral momentum theory: Equations and applications. Journal of Applied Behavior Analysis, 44(4), 877–895. https://doi.org/10.1901/jaba.2011.44-877.
Okouchi, H. (2015). Resurgence of two-response sequences punished by point-loss response cost in humans. Revista Mexicana de Análisis de la Conducta, 41, 137–154.
Petscher, E. S., Rey, C., & Bailey, J. S. (2009). A review of empirical support for differential reinforcement of alternative behavior. Research in Developmental Disabilities, 30(3), 409–425. https://doi.org/10.1016/j.ridd.2008.08.008.
Romano, L. M., & St. Peter, C. C. (2016). Omission training results in more resurgence than alternative reinforcement. The Psychological Record, 67, 315–324. https://doi.org/10.1007/s40732-016-0214-z.
Shahan, T. A., & Craig, A. R. (2016). Resurgence as choice. Behavioural Processes, 141, 100–127. https://doi.org/10.1016/j.beproc.2016.10.006.
St. Peter, C. C. (2015). Six reasons why applied behavior analysts should know about resurgence. Revista Mexicana De Análisis De La Conducta, 41, 252–268.
St. Peter Pipkin, C., Vollmer, T. R., & Sloman, K. N. (2010). Effects of treatment integrity failures during differential reinforcement of alternative behavior: A translational model. Journal of Applied Behavior Analysis, 42, 47–70.
Volkert, V. M., Lerman, D. C., Call, N. A., & Trosclair-Lasserre, N. (2009). An evaluation of resurgence during treatment with functional communication training. Journal of Applied Behavior Analysis, 42(1), 145–160. https://doi.org/10.1901/jaba.2009.42-145.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Ethical Approval
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki declaration and its later amendments or comparable ethical standards.
Informed Consent
Informed consent was obtained from all individual participants included in the study.
Additional information
Authorship among the first two authors should be considered equal.
Rights and permissions
About this article

Cite this article
Kestner, K.M., Romano, L.M., St. Peter, C.C. et al. Resurgence Following Response Cost in a Human-Operant Procedure. Psychol Rec 68, 81–87 (2018). https://doi.org/10.1007/s40732-018-0270-7
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40732-018-0270-7