
Abstract
Plants are frequently exposed to a variety of stress conditions such as drought, salinity, heavy metal toxicity, low temperature, flooding, extremes of soil pH and heat which causes reduction of plant growth and productivity. Such abiotic stresses may cause metabolic impairment, nutrient imbalance, reduced synthesis of photosynthetic pigments which are closely related with biomass production in plant, thus, causing serious loss in crop productivity. The present experiment was undertaken to study the biochemical and physiological effects of salinity, drought and heavy metal (copper and lead) stress on seed germination in ricebean [Vigna umbellata (Thunb.) Ohwi and Ohashi] variety Bidhan 1. For studying the effect of iso-osmotic potential of salinity and drought stress, the solutions of NaCl and PEG 6000 with − 0.2, − 0.4 and − 0.8 MPa osmotic potential were used whereas the solutions of 50, 100 and 200 µM Cu and Pb supplemented in the form of CuSO4.·5H2O and Pb(NO3)2 respectively were used to study the effects of equimolar concentrations of copper and lead. Drought was found to produce more adverse effects on germination %, as well as speed of germination, in the seeds of ricebean. The seed protein content was significantly higher under all the treatments of salinity, drought stress as well as metal stress. The highest intensity of copper stress was found to produce more adverse effects than lead in respect of water uptake % in germinating seeds and root elongation rates of ricebean seeds in the present experiment. The presence of copper in the germinating medium produced more detrimental effects on activities of antioxidative enzyme ascorbate peroxidase and guaiacol peroxidase than equimolar concentrations of lead.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Salinity stress can affect plants initially by creating an osmotic stress and then it induces ion toxicity leading to cyto-toxicity, metabolic impairment, nutrient imbalance and finally death of the plant. The presence of high concentration of Na+, Cl−, HCO3−, K+, Ca2+, Mg2+ and SO42− ions usually contribute to the soil salinity. Initially, the presence of salts in high concentration makes it very difficult for plants to withdraw water from soil due to very low osmotic potential. In effect, the plants suffer from a sort of osmotic stress which restricts plant growth causing yield reduction. In the later stages of stress, due to the absorption of sodium and chloride ions in high concentration plants suffer from cyto-toxicity which result in reduction of growth, leaf burn and plant death.
Drought is a meteorological term and defined as a period without significant rainfall. The effects are mainly related to altered metabolic functions, reduced synthesis of photosynthetic pigments, uptake and translocation of ion, carbohydrate biosynthesis, nutrient metabolism and synthesis of growth promoters. A common adverse effect of moisture deficit stress on crop plants is the reduction in fresh and dry biomass (Farooq et al. 2009).
A heavy metal is any one of a number of high atomic weight elements which has the properties of a metallic substance at room temperature. Among heavy metals, copper (Cu) is considered as an essential micronutrient for plants (Gang et al. 2013) and plays important role in CO2 assimilation and ATP synthesis. However excess copper creates toxicity generating oxidative stress and ROS accumulation (Stadtman and Oliver 1991). Oxidative stress causes disturbance of metabolic pathways and damage to macromolecules (Hegedus et al. 2001). Lead (Pb) is one of the most abundant toxic elements in the soil, which exerts adverse effect on morphology, growth and photosynthetic processes of plants when present in excess amount (Yadav 2010). Inhibition of seed germination under lead toxicity may result from the interference of lead with important enzymes.
Ricebean [Vigna umbellata (Thunb.) Ohwi and Ohashi] is a potential leguminous crop with luxuriant growth habit and ability to produce huge quantity of nutritive green fodder as well as high seed yield under limited management inputs (Chatterjee et al. 1978). It has gained attention as a potential under-utilized grain legume owing to its profuse pod bearing habit, wider adaptability, tolerance to biotic and abiotic stresses, high seed yield and high contents of protein, calcium, phosphorus, tryptophan as well as starch in its seeds (Srivastava et al. 2001). It thrives well in marginal lands, rainfed areas, drought-prone areas and exhausted soils. Only a limited number of research works have so far been conducted on abiotic stress responses of ricebean (Subrahmanyam 1998; Pal et al. 2009; Rai 2013). However, such information is still a meager. The present experiment has been designed to study the comparative effects of different levels of salinity, drought, copper and lead stress on some physiological and biochemical parameters of ricebean during seed germination stage. Such a study might find to be useful in promotion of this under-utilized leguminous crop for cultivation in marginal as well as stress-prone areas where other grain legumes may not be suitable for cultivation.
Materials and methods
Seeds of ricebean [Vigna umbellata (Thunb) Ohwi and Ohashi] variety Bidhan-1 were used in the experiment. The seeds were collected from AICRP on Forage Crops, Kalyani Centre. All together thirteen treatments involving three doses each of salinity, drought, copper and lead stress, along with unstressed control were considered for the experiment. The salinity and drought stress were imposed by application of 50, 100 and 200 mM of NaCl and 10, 12 and 18% PEG 6000 solution, thus, creating osmotic potential of − 0.2, − 0.4 and − 0.8 MPa, respectively, as per Sosa et al. (2005).The heavy metal stress was imposed by applying 50, 100 and 200 µM Cu and Pb supplemented in the form of CuSO4.·5H2O and Pb(NO3)2, respectively. The seeds were surface sterilized with 0.1% (w/v) HgCl2 for 3 min followed by thorough washing in distilled water. After that, the seeds were set to germinate in petridish of 9 cm diameter lined with Whatman No.1 filter paper at a temperature of 28 ± 1 °C and relative humidity of 80 ± 1% and moistened with 5 ml each of treatment solution. A control set containing distilled water was used for comparison. In each treatment, there were three replications and each replication had four sets of petridish each containing 25 seeds.
On the fourth day of germination, final germination count was taken. From the count of seeds germinated in each day the speed of germination was calculated as per Czabator (1962).Observations were recorded on the water uptake percentage by the germinating seeds at 6 h interval starting from 12 h of germination following the methods described by Foti et al. (2018). The root elongation rate was measured at 24 h interval starting from 24 h of germination upto 120 h and was expressed in cm h−1. The soluble protein content of the cotyledon of germinating seed was measured as per Lowry et al. (1951) at 24 h of germination. Data were also recorded on the activities of three important antioxidative enzymes, viz, ascorbate peroxidase (APX), guaiacol peroxidase (GPOX) and catalase enzyme in the cotyledon at 24 h of germination. The activities of APX and GPOX were measured as per Nakano and Asada (1981) and Siegel and Galston (1967), respectively, while the method described by Cakmak et al. (1993) was followed for the activity of catalase enzyme.
The mean data in all the cases were subjected to statistical analysis following completely randomised design using INDOSTAT version 7.1 software.
Results and discussion
Analysis of variance indicated that the treatments varied significantly among themselves in case of all the germination growth and biochemical parameters under study. All the treatments scored 100% final germination percentage except the 12 and 18% PEG treatment where it was 96 and 92%, respectively (Table 1). However, there was significant negative effects of salinity and drought stress on the speed of germination, especially, at higher concentrations of NaCl and PEG 6000. Thus, the germinating seeds of ricebean cv. Bidhan 1 recorded 34.95% and 44.27% reduction in speed of germination at 200 mM NaCl and 18% PEG solution producing an osmotic potential of − 0.8 MPa, respectively, over that of control. Such reduced speed of germination might be attributed for moisture limitation caused by lower osmotic potential of germination media as well as reduction in the utilization of seed reserves as were also reported earlier by Khan (1980), Promila and Kumar (2000) and Sayar et al. (2010). In contrast, the presence of copper and lead in the germinating media did not cause any remarkable change in germination speed as compared to unstressed control. Thus, copper and lead at their highest concentrations caused only 2.09% and 6.96%, reduction, respectively.
Water uptake by seed is highly related to metabolic events during germination process as it involves the activation of most hydrolytic enzymes that stimulate hydrolysis of starch reserves into sugars for supplying energy source needed for radical emergence and tissue elongation (Bewley et al. 2013).Water potential and hydraulic conductivity of the sowing medium as well as the hydraulic conductivity of the seeds play a major role in water uptake (Ward and Shaykewich 1972). In the present experiment, the drought stress at the lowest osmotic potential was found to produce more adverse effects on water uptake of germinating seeds of ricebean as compared to iso-osmotic potential of salinity (Table 2). In general, the water uptake by seeds increased linearly at all hours of germination in all the cases. The water uptake percentage was found to be remarkably low under drought stress at all osmotic potentials than salinity stress, thus, attributing for slower germination speed under drought treatments. Earlier, the adverse effects of salinity stress (Gamze 2005; Zhang et al. 2010; Fetri et al. 2014) and drought stress (Babu and Rosaiah 2017; Moliehi et al. 2017; Foti et al. 2018) on imbibition of seeds of legumes were reported by different workers. The medium to high concentrations of both the metals registered slower water uptake for initial 12 and 18 h. It might be noted that the low rate of water uptake under Cu 200 µM could not be made up even at 24 h. Thus, at 24 h copper was found to be more drastic than lead for this variety of ricebean in terms of water uptake as was also reported by Marques et al. (2007) and Ling and Jun (2010) in other crops.
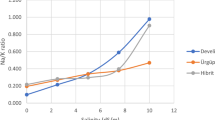
The data on the rate of root elongation were recorded at 24-h interval starting from 24 h of germination upto 120 h and have been presented in Fig. 1A, B. Perusal of the graphical presentation revealed that the rates exhibited a sigmoidal pattern of change under control condition with the rates being almost constant at initial two intervals followed by a peak in between 72 and 96 h of germination and then a decline. However, the elongation rate was higher under unstressed control condition at all the intervals under study in comparison with the stress treatments. Such a reduction in rate of radicle emergence might be attributed to a reduced rate of cell division caused by impairment of reserve mobilization (Foti et al. 2018, 2019).The rate under NaCl 50 mM also showed sigmoidal curve with the increasing rate upto the interval of 48–72 h of germination followed by gradual decline. The corresponding rates under PEG 10% (− 0.2 MPa osmotic potential) treatment increased linearly for the first three intervals with the rate being maximum at 72–96 h of germination followed by slight decrease at 96–120 h of germination. The root elongation under PEG 12% (− 0.4 MPa osmotic potential) treatment exhibited almost constant rates at different intervals of germination. However, among all the treatments, the rates of root elongation were found to be most adversely affected under NaCl 100 and 200 mM treatments. It might be noted that Cu 50 and 100 µM as well as Pb 50 and 100 µM treatments registered higher means for root elongation rate at the very initial interval (24–48 h) as compared to control condition, but then declined sharply as the germination interval progressed. Earlier Peralta et al. (2001) reported stimulatory effects of low doses of heavy metals on root elongation in alfalfa. However, the comparative study in the present experiment indicated more damaging effect of copper on root elongation rate than lead, especially, under higher molar concentration. It might be discussed that although Cu is an essential micronutrient for plant growth, it was found to be more inhibitory than lead in affecting radicle growth in the present experiment. This might be attributed to structural damage in plants in presence of excessive Cu as was suggested earlier by Fernandes and Henriques (1991).

A Effect of salinity and drought stress on rate of root elongation in ricebean during different intervals of germination. B Effect of copper and lead stress on rate of root elongation in ricebean during different intervals of germination
The soluble protein content in the cotyledon of germinating seeds significantly increased at − 0.2 and − 0.4 MPa osmotic potentials under salinity and drought stress in comparison with unstressed control (Table 3), a fact which might be attributed for acclimation strategy to water deficit as was proposed earlier by Nakashima et al. (2009). However, the protein content decreased considerably under the highest stress intensity of − 0.8 MPa created by 200 mM NaCl and 18% PEG solution. The decrease was more in case of PEG than NaCl indicating more detrimental effect of drought stress on soluble protein content of cotyledon during germination. Earlier the adverse effects of salinity stress and drought stress on seed protein content in legumes were reported by Verma et al.(2012) and Bhardwaj and Yadav (2012).The protein content also decreased under the medium and high concentrations of lead, but in different concentrations of copper it significantly increased with the lowest dose registering the highest increase over control. Singh et al. (2011) showed that lead (Pb) toxicity retarded the radical emergence via alteration in protein and carbohydrate contents, affecting the activity of many enzymes. In case of cotyledon protein content drought was found to produce more adverse effects, whereas the effects of lead was found to be more drastic than copper at equimolar concentration.
Different abiotic stresses in plants lead to enhanced generation of reactive oxygen species (ROS) leading to oxidative stress. Under this situation, different enzymatic and non-enzymatic mechanisms are induced to combat against such oxidative stress. Several antioxidative enzymes like catalase (CAT), ascorbate peroxidase (APX) and guaiacol peroxidase (GPOX) contribute to plant defense and helpin scavenging of ROS. Catalase (CAT) is tetrameric heme containing enzymes with the potential to directly dismutate H2O2 into H2O and O2 and is indispensable for ROS detoxification during stressed conditions. In the present experiment, catalase (CAT) activity in the cotyledon significantly decreased under most of the salinity and drought treatments as compared to control (Table 3). Only in case of 100 mM NaCl and 10% PEG solution the activity increased over that of control. Thus, mild osmotic stress might trigger the activity of this scavenging enzyme which was inhibited as the intensity of stress increased. Earlier the detrimental effects of salinity stress (Dar et al. 2007) and drought stress (Pratap and Sharma 2010, Jiang-Jing Long et al. 2013) on CAT activity in legumes were reported by different workers. Likewise, the cotyledon CAT activity also decreased under all the concentrations of lead treatments with the effects being more as the molar concentration increased. In contrast, the activity increased at 50 µM of copper followed by significant decrease under higher concentrations of the metal. However, copper was found to be more detrimental than lead for CAT activity. Such negative effects of copper on CAT activity might be attributed for oxidation of CAT-protein structure (Pena et al. 2011).
The activity of APX in the cotyledon of germinating seed of Bidhan-1 significantly increased under salinity and drought with the exception of PEG 18% (− 0.8 MPa osmotic potential) treatment (Table 3). In case of salinity stress the activity registered linear decrease with increase in stress intensity, while in case of drought the pattern of change was a sigmoidal one with PEG 12% (− 0.4 MPa osmotic potential) treatment recording the highest activity. Earlier the adverse effects of salinity stress (Dar et al. 2007) and drought stress (Bhardwaj and Yadav 2012, Jiang-Jing Long et al. 2013) on APX in legumes were reported by different workers. The cotyledon APX activity decreased linearly under increasing concentrations of lead. In contrast, it showed sigmoidal pattern of changes in response to increasing concentrations of copper. However, the highest concentration of copper was found to be more detrimental for APX activity in comparison with lead. It might be further noted that the APX content in ricebean seed registered more adverse effects of salinity stress in comparison with drought stress at iso-osmotic potentials.
The GPOX activity in cotyledon showed sigmoidal pattern of changes under different salinity treatments, while it consistently decreased under increasing levels of drought stress. The drought stress had more negative effects on GPOX activity in the present experiment as compared to iso-osmotic salinity stress (Table 3). Earlier the adverse effects of salinity stress (Amirjani 2010; Verma et al. 2012) and drought stress (Bhardwaj and Yadav 2012) on GPOX activity in legumes were reported by different workers. The activity of GPOX of this variety of ricebean showed increased activities under mild and medium level of metal stress in the present experiment. This finding corroborated the early observation of Gao et al. (2008) who proposed the increased expression of peroxidase enzyme as a biomarker of sub-lethal dose of metal toxicity. However, the activity of GPOX in the present experiment decreased significantly as the metal concentration in the geminating medium reached the maximum. However, copper was found to be more detrimental for GPOX activity in comparison with lead at the highest concentration.
Conclusion
Summarizing the results, it might be concluded that the drought stress was found to register more drastic effects on different biochemical and physiological parameters of germinating seed as compared to iso-osmotic potential of salinity stress, especially, at highest intensity of stress. Also the copper toxicity produced more adverse effects than lead toxicity at equimolar concentration in ricebean cultivar Bidhan 1 during germination stage.
References
Amirjani, M. R. (2010). Effect of salinity stress on growth, mineral composition, proline content, antioxidant enzymes of soybean. American Journal of Plant Physiology, 5, 350–360.
Babu, K., & Rosaiah, G. (2017). A study on germination and seedling growth of blcakgram (Vignamungo L. Hepper) germplasm against polyethylene glycol 6000 stress. IOSR Journal of Pharmacy and Biological Sciences (IOSR-JPBS), 12, 90–98.
Bewley, J. D., Bradford, K. J., Hilhorst, H. W. M., & Nonogaki, H. (2013). Seeds. In J. D. Bewley, K. J. Bradford, H. W. M. Hilhorst, & H. Nonogaki (Eds.), Physiology of development, germination and dormancy (pp. 133–183). London: Springer, New York Heidelberg Dordrecht.
Bhardwaj, J., & Yadav, S. K. (2012). Comparative study on biochemical parameters and antioxidant enzymes in a drought tolerant and a sensitive variety of horsegram (Macrotyloma uniflorum) under drought stress. American Journal of Plant Physiology, 7, 17–29.
Cakmak, I., Strbac, D., & Marschner, H. (1993). Activities of hydrogen peroxide scavenging enzymes in germinated wheat seeds. Journal of Experimental Botany, 44, 127–132.
Chatterjee, B. N., Mukherjee, A. K., Bhattacharya, K. K., Mandal, S. R., Rana, S. K., & Mandal, B. K. (1978). Production potentiality of forage cropping systems and their effects on soil and crop productivity in the gangetic plains of Eastern India. Forage Research, 4, 73–80.
Czabator, F. J. (1962). Germination value: An index combining speed and completeness of pine seed germination. Forest Science, 8, 386–396.
Dar, Z. M., Hemantaranjan, A., & Panday, S. K. (2007). Antioxidative response of mungbean (Vigna radiata L.) to salt stress. Legume Research, 30, 57–60.
Farooq, M., Wahid, A., Kobayashi, N., Fujita, D., & Basra, S. M. A. (2009). Plant drought stress: Effects, mechanisms and management. Agronomy for Sustainable Development, 29, 185–212.
Fernandes, J. C., & Henriques, F. S. (1991). Biochemical, physiological, and structural effects of excess copper in plants. Botanical Review, 57, 246–273.
Fetri, M., Dargahikhoo, A., & Rajabi, M. (2014). Effect of drought and salinity tensions on germination and seedling growth of common yarrow (Achillea millefolium L.) in laboratory conditions. International Journal of Advanced Biological and Biomedical Research, 2, 383–391.
Foti, C., Khah, E., & Pavli, O. (2018). Response of lentil genotypes under PEG-induced drought stress: Effect on germination and growth. Plant, 6, 75–83.
Foti, C., Khah, E. M., & Pavli, O. I. (2019). Germination profiling of lentil genotypes subjected to salinity Stress. Plant Biology, 21, 480–486.
Gamze, O. K. C. U. (2005). Effects of salt and drought stresses on germination and seedling growth of pea (Pisum sativum L.). Turkish Journal of Agriculture and Forestry., 29, 237–242.
Gang, A., Vyar, A., & Vgas, H. (2013). Toxic effect of heavy metals on germination and seedling growth of wheat. Journal of Environmental Research and Developement, 8, 206–213.
Gao, S., Yan, R., Cao, M., Yang, W., Wang, S., & Chen, F. (2008). Effects of copper on growth, antioxidant enzymes and phenylalanine ammonia-lyase activities in Jatrophacurcas L. seedling. Plant Soil Environment, 54(3), 117–122.
Hegedus, A., Erdei, S., & Horvath, G. (2001). Comparative studies of H2O2 detoxifying enzymes in green and greening barley seedings under cadmium stress. Plant Science, 160, 1085–1093.
Jiang-Jing Long; Su-Miao; Chen-YueRu; Gao-Nan; Jiao-ChengJin; Sun-ZhengXi; Li-FengMin; Wang-ChongYing,. (2013). Correlation of drought resistance in grass pea (Lathyrus sativus) with reactive oxygen species scavenging and osmotic adjustment. Biologia-Bratislava, 68, 231–240.
Khan, A. A. (1980). The physiology and biochemistry of dormancy and germination. North-Holland: Publishing company, oxford.
Ling, T., & Jun, R. (2010). Effect of Hg on seed germination, coleoptile growth and root elongation in seven pulses. Fresenius Environmental Bulletin, 19, 1144–1150.
Lowry, O. H., Rosebrogh, N. J., Farr, L., & Randall, R. J. (1951). Protein measurement with folin phenol reagent. Journal of Biological Chemistry, 193, 265–275.
Marques, A. P. G. C., Rangel, A. O. S. S., & Castro, M. L. (2007). Effect of arsenic, lead and zinc on seed germination and plant growth in black nightshade (Solanum nigrum L.) vs. Clover (Trifolium incarnatum L.). Fresenius Environmental Bulletin, 16, 896–903.
Moliehi, R., Mateboho, M., & Motlatsi, M. (2017). Screening of common bean cultivars (Phaseolus vulgaris l.) for drought tolerance–1. Global Journal of Agricultural Research, 5, 20–29.
Nakano, Y., & Asada, K. (1981). Hydrogen peroxide is scavenged by ascorbate specific peroxidase in spinach chloroplasts. Plant and Cell Physiology, 22, 867–880.
Nakashima, K., Ito, Y., & Yamaguchi-Shinozaki, K. (2009). Transcriptional regulatory networks in response to abiotic stresses in Arabidopsis and grasses. Plant Physiology, 149, 88–95.
Pal, A. K., Rai, D., & De, D. K. (2009). Evaluation of ricebean genotypes for salt tolerance during early seedling growth and physiological basis of tolerance. Forage Research, 35, 73–79.
Pena, L. B., Azpilicueta, C. E., & Gallego, S. M. (2011). Sunflower cotyledons cope with copper stress by inducing catalase subunits less sensitive to oxidation. Journal of Trace Elements in Medicine and Biology, 25, 125–129.
Peralta, J. R., Gardea-Torresdey, J. L., Tiemann, K. J., Gomez, E., Arteaga, S., Rascon, E., et al. (2001). Uptake and effects of five heavy metals on seed germination and plant growth in alfalfa (Medicago sativa L.). Bulletin of Environmental Contamination Toxicology, 66, 727–734.
Pratap, V., & Sharma, Y. K. (2010). Impact of osmotic stress on seed germination and seedling growth in blackgram (Phaseolus mungo). Journal of Environmental Biology, 31, 721–726.
Promila, K., & Kumar, S. (2000). Vigna radiata seed germination under salinity. BiologiaPlantarum, 43, 423–426.
Rai, D. (2013). Changes in growth, enzyme activities and osmolytes of ricebean [Vigna umbellata (Thunb.) Ohwi and Ohashi] seedlings during abiotic stress and recovery. Ph.D. Thesis, Bidhan Chandra KrishiViswavidyalaya, Mohanpur, Nadia.
Sayar, R., Bchini, H., Mosbahi, M., & Ezzine, M. (2010). Effects of salt and drought stresses on germination, emergence and seedling growth of durum wheat (Triticum durum Desf.). African Journal of Agricultural Research, 5, 2008–2016.
Siegel, B. Z., & Galston, A. W. (1967). Theisoperoxidases of Pisum sativum. Plant Physiology, 42, 212–226.
Singh, H. P., Kaur, G., Batish, D. R., & Kohli, R. K. (2011). Lead (Pb)-inhibited radicle emergence in Brassica campestris involves alterations in starch-metabolizing enzymes. Biological Trace Elemental Research, 144, 1295–1301.
Sosa, L., Llanes, A. A., Herminda, R., Mariana, R., & Virginia, L. (2005). Osmotic and specific ion effects on the germination of Prosopis strombulifera. Annals of Botany, 96, 261–267.
Srivastava, R. P., Srivastava, G. K., & Gupta, R. K. (2001). Nutritional quality of ricebean (Vigna umbellata). Indian Journal of Agricultural Biochemistry, 14, 55–56.
Stadtman, E. R., & Oliver, C. N. (1991). Metal-catalyzed oxidation of proteins physiological consequences. Journal of Biological Chemistry, 266, 2005–2008.
Subrahmanyam, D. (1998). Effect of aluminium on growth, lipid peroxidation, superoxide dismutase and peroxidase activities in rice and French bean seedlings. Indian Journal of Plant Physiology, 3, 240–242.
Verma, S. K., Chaudhary, M., & Prakash, V. (2012). Study of the alleviation of salinity effect due to enzymatic and non enzymatic antioxidants in glycine max. Research-Journal-of-Pharmaceutical,-Biological-and-Chemical-Sciences, 3, 1177–1185.
Ward, J., & Shaykewich, C. (1972). Water absorption by wheat seeds as influenced by hydraulic properties of soil. Canadian Journal of Soil Science, 52, 99–105.
Yadav, S. K. (2010). Heavy metals toxicity in plants: An overview on the role of glutathione and phytochelatins in heavy metal stress tolerance of plants. South African Journal of Botany, 76, 167–179.
Zhang, H. X., Irving, L. J., McGill, C., Matthew, C., DaoWei, Z., & Kemp, P. (2010). The effects of salinity and osmotic stress on barley germination rate: Sodium as an osmotic regulator. Annals of Botany, 106, 1027–1035.
Acknowledgement
The authors acknowledge the assistance extended by AICRP on Forage Crops, Kalyani Centre, for supplying plant materials.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Atta, K., Pal, A.K. & Jana, K. Effects of salinity, drought and heavy metal stress during seed germination stage in ricebean [Vigna umbellata (Thunb.) Ohwi and Ohashi]. Plant Physiol. Rep. 26, 109–115 (2021). https://doi.org/10.1007/s40502-020-00542-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40502-020-00542-4