
Abstract
Green gram (Vigna radiata (L.) Wilczek) is a widely cultivated pulse crop in India, and it holds a key importance in food supplement due to its protein-rich edible seeds. This crop usually experiences abiotic stresses like drought and salinity during the cultivation period. The investigation was carried out to study the effect of hydropriming on osmotic stress tolerance potential of three Vigna radiata varieties, with varied abiotic stress tolerance potential. Hydropriming enhances the photosynthetic activities of the seedlings. Moreover, hydropriming resulted in reduction of proline content in Pusa Ratna and Pusa 9531 seedlings, whereas the salt-tolerant variety Pusa Vishal showed increase in proline content, especially under stressed conditions. Osmotic stress caused increase in MDA content in all the varieties studied, and this increase was significantly minimized by the influence of hydropriming. Hydropriming also enhanced the activities of antioxidant enzymes in the seedlings. From the results of this study, it is concluded that hydropriming of seeds improved the drought and salinity stress tolerance potential of all the three green gram varieties and it was significantly evident in the abiotic stress-sensitive variety Pusa Ratna as compared to Pusa 9531 (drought tolerant) and Pusa Vishal (NaCl tolerant).
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Abiotic stresses lead to substantial and unpredictable loss in crop production in agriculture. Of the different abiotic stresses, the major ones, drought and salinity causes a variety of biochemical, physiologic and metabolic changes in plants, ultimately compromising on yield [22]. Green gram (Vigna radiata (L.) Wilczek) is an important dietary pulse crop enriched with protein. Various abiotic stresses, especially drought and salinity are the major limitations to its production. Till date, various strategies are employed to generate plants which can withstand abiotic stresses. Seed priming has been developed as an indispensable method to produce tolerant plants against various stresses. Seed priming is the induction of a particular physiologic state in plants by the treatment of natural and/or synthetic compounds to the seeds before germination. Plants raised from primed seeds showed various advantages over the non-primed ones, such as sturdy and quick cellular defense response against abiotic stresses [13].
Hydropriming has been reported to be a simple, economical and a safe technique for increasing the capacity of seeds toward osmotic adjustment, enhancing seedling establishment and crop production under stressed conditions [14]. In this priming method, the seeds are immersed in sterilized distilled water and kept at appropriate temperature. The duration of hydropriming is determined by controlling seed imbibitions during germination [15]. After hydropriming, seeds are re-dried to original weight with forced air under shade [6]. The beneficial effects of hydropriming were reported by many research works. According to Umair et al. [21], hydropriming significantly increased the seed yield of V. radiata and also enhanced the antioxidant enzymatic activities. According to Posmyk and Janas [19], hydropriming and hydropriming along with proline can be used as a safe priming method for improving seed germination and growth of V. radiata seedlings at low temperature and also allowing fast repair of injuries caused by stress.
The present investigation was carried out to study the effect of hydropriming on osmotic stress tolerance potential of green gram varieties with varied tolerance potential to NaCl and drought, by analyzing the biochemical changes and primary metabolic changes of primed and non-primed plants subjected to the above stressed conditions.
Materials and Methods
Materials
Green gram belongs to the family Papilionaceae and is an important protein-rich pulse crop. The seeds of V. radiata varieties (Pusa Ratna, Pusa 9531, Pusa Vishal) were procured from Seed Science and Technology Division, Indian Agricultural Research Institute, New Delhi.
Methods
Hydropriming Techniques
Plump, healthy and uniform seeds of V. radiata were selected for hydropriming and immersed into distilled water, whose volumes were 3 times to the seed weights. Hydropriming interval was fixed as 6 h, because above this time interval leakage of metabolites from the seeds were observed. During hydropriming treatment in an open beaker, the seeds were swirled regularly to ensure proper aeration. After 6 h of hydropriming treatment, the seeds were washed with distilled water for 2 min and surface dried on absorbent paper. Further, the seeds were placed on a piece of clean filter paper, allowing dehydration under shade at 25 °C to retrieve the original seed moisture before priming treatment. Seed weights were tested repeatedly at fixed time intervals so as to ensure that the seeds attained the original dry weight. The untreated seeds were used as the control. The concentration of stress imparting concentrations of NaCl and PEG was determined through a series of standardization process. Primed as well as non-primed seeds were germinated in light transparent plastic bottles (19 × 11 cm) containing absorbent cotton soaked with distilled water (control), different concentrations of NaCl (0, 25, 50, 75, 100, 125 mM) and poly ethylene glycol (PEG)-6000 (0, 5, 10, 15, 20, 25%) solution. The bottles were kept under a continuous light (120 μmol m−2 s−1) at 25 ± 2 °C and RH 60 ± 5. All the seed materials used for investigation were pre-washed for one min with 0.25% Triton X-100 (Boehringer Mannheim Gmbh). The physiologic and biochemical attributes of primed, non-primed as well as control seedlings were recorded on 7 days after germination.
Physiologic and Biochemical Studies
Chlorophyll estimation was carried out by the method of Arnon [2].Proline content in the seedlings was estimated as per Bates et al. [4].. The MDA content estimation was done according to Heath and Packer [10]. Nitrate reductase activity was measured according to the method suggested by Hageman and Reed [9]. For the estimation of superoxide dismutase (SOD) activity, the protocol of Giannoplitis and Ries [8] was adopted. The peroxidase (PER) activity was measured as per Gaspar et al. [7].
Thylakoids from leaves were isolated according to the method of Puthur [20] and the photochemical activities of thylakoids were assayed polarographically with a Clark-type oxygen electrode (DW1/AD, Hansatech, Norflok, UK) which was connected to a digital control box (OXYG1, Hansatech) at 4 °C. The light-dependent O2 uptake/evolution was measured by irradiating the sample with saturating intensity of white light (∽ 1800 μmol photons m−2 s−1), provided by a 100 W halogen lamp (LS2, Hansatech). The activity of PS I and PS II was calculated in terms of nmol of O2 consumed/evolved min−1 mg−1 chlorophyll.
Statistical Analysis
Univariate analysis of variance (ANOVA) was carried out by using the software SPSS 19. Means were compared using Duncan’s test (P < 0.05).
Results
Determination of Stress Imparting Concentration and Priming Interval
The three different varieties of V. radiata selected for the present study were based on its varied levels of drought/NaCl tolerance potential. Pusa Ratna, a stress-sensitive variety showed 50% growth retardation at 75 mM NaCl and 15% PEG, whereas it was at 75 mM NaCl and 20% PEG for Pusa 9531 (drought tolerant) and for Pusa Vishal (NaCl tolerant), 50% growth retardation was at 100 mM NaCl and 15% PEG. The priming interval for all varieties of V. radiata was fixed at 6 h, as above this interval, the solutes from the seeds leached out.
Primary Metabolic Changes
Photosynthetic Pigment Composition and Photosystem Activities
Hydropriming of seeds caused an increase in the leaf photosynthetic pigment content of the seedlings in all the three varieties of V. radiata. In Pusa Ratna, chlorophyll a, b and total chlorophyll was found to be maximum in the seedlings which were raised from hydroprimed seeds and grown under unstressed and NaCl-stressed conditions, while in those seedlings which were grown under PEG-stressed conditions, the increase in the above parameters was comparatively less. On the other hand, the increase in the carotenoid content of the Pusa Ratna seedlings on hydropriming was nearly equal in all the three conditions (0, NaCl and PEG stress). In the drought-tolerant variety Pusa 9531, significant increase in the photosynthetic pigments was obtained only in the seedlings which were grown under PEG-stressed conditions, while in the NaCl-tolerant variety Pusa Vishal, significant increase in photosynthetic pigment content was obtained in the seedlings which were grown under unstressed conditions (Table 1).
The activity of the photosystem I (PS I) increased in the leaves of seedlings of V. radiata raised from hydroprimed seeds under 0, NaCl and PEG stress. The enhancement of PS I activity was significant in the seedlings of Pusa Ratna raised from hydroprimed seeds and subjected to stress (NaCl/PEG) as compared to other two varieties (Fig. 1a). As a result of seed hydropriming, maximum increase in PS II activity was obtained in the seedlings of Pusa Vishal, followed by Pusa Ratna, both under PEG stress. The seedlings raised from hydroprimed seeds of Pusa Ratna also showed considerable increase in PS II activity under unstressed and NaCl-stressed conditions. In Pusa 9531 and Pusa Vishal, maximum increase in PS II activity was observed in seedlings which were raised from hydroprimed seeds and subjected to stressed conditions while under unstressed condition, the increase in PS II activity was less (Fig. 1b).

Effect of hydropriming on PS I activity (a) and PS II activity (b) of three V. radiata varieties grown under 0-, NaCl- and PEG-stressed conditions. The vertical bars represent SE of the mean value of recordings from three independent experiments each with a minimum of three replicates. Values with different letters are significantly different at 5% level (P < 0.05; ANOVA, Duncan’s test)
Proline Content
The three varieties of V. radiata differentially responded to seed hydropriming in the accumulation pattern of proline content. Among the three varieties of V. radiata, Pusa Ratna showed proline accumulation under unstressed condition while under stressed conditions (NaCl/PEG stress), the proline content got reduced in the seedlings raised from hydroprimed seeds. In the tolerant varieties i.e., in Pusa 9531 and Pusa Vishal, hydropriming resulted in proline accumulation at all three conditions (0, NaCl and PEG stress) (Fig. 2).

Effect of hydropriming on proline content of three V. radiata varieties grown under 0-, NaCl- and PEG-stressed conditions. The vertical bars represent SE of the mean value of recordings from three independent experiments each with a minimum of three replicates. Values with different letters are significantly different at 5% level (P < 0.05; ANOVA, Duncan’s test)
MDA Content
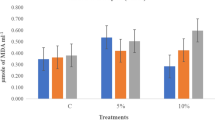
The MDA content was found to decrease in the seedlings of all three varieties raised from hydroprimed seeds as compared to seedlings raised from non-primed seeds, either in the absence or presence of stress (NaCl/PEG). MDA content reduction on hydropriming was evidently occurring in the Pusa Vishal variety when the seedlings were raised under PEG stress (Fig. 3).

Effect of hydropriming on MDA content of three V. radiata varieties grown under 0-, NaCl- and PEG-stressed conditions. The vertical bars represent SE of the mean value of recordings from three independent experiments each with a minimum of three replicates. Values with different letters are significantly different at 5% level (P < 0.05; ANOVA, Duncan’s test)
Enzyme Activities
The activity of nitrate reductase (NR) enzyme was increased in seedlings raised from hydroprimed seeds in all the varieties studied, and it was maximum in the Pusa Ratna seedlings which were grown under unstressed condition (Fig. 4a). Seed hydropriming in Pusa Ratna and Pusa 9531 resulted in an ample increase in superoxide dismutase (SOD) activity but in the Pusa Vishal, seed hydropriming was not able to bring about such an enhancement in the activity of SOD (Fig. 4b). The activity of guaiacol peroxidase (POX) was increased significantly in the Pusa Vishal seedlings which were raised from hydroprimed seeds and grown under stressed (NaCl/PEG) conditions. Pusa 9531 also showed increased activity of POX in the seedlings which were raised from hydroprimed seeds and grown under PEG stress. While in the abiotic stress-sensitive variety Pusa Ratna, hydropriming caused a decrease in the activity of POX under unstressed and stressed (NaCl/PEG) conditions (Fig. 4c).

Effect of hydropriming on nitrate reductase (NR) activity (a), superoxide dismutase (SOD) activity (b) and guaiacol peroxidase (POX) activity (c) of three V. radiata varieties grown under 0-, NaCl- and PEG-stressed conditions. The vertical bars represent SE of the mean value of recordings from three independent experiments each with a minimum of three replicates. Values with different letters are significantly different at 5% level (P < 0.05; ANOVA, Duncan’s test)
Discussion
The three varieties of V. radiata studied were with varied levels of NaCl/PEG tolerance potentials. Thus, it was necessary to check the NaCl/PEG stress tolerance potential of all these varieties in order to identify a concentration which imparted ~ 50% growth retardation(in terms of shoot length, fresh and dry weight), and it varied for each variety [11, 12].
Hydropriming was found to be efficient in increasing the photosynthetic pigment content and photosystem activities of the seedlings. This might be due to the positive influence of hydropriming on the biosynthesis of photosynthetic pigments or due to reduction in the degradation of photosynthetic pigments. The increased amount of these pigments in turn resulted in effective absorption and channelization of light energy into the photosystems, resulting in an enhanced PS I and PS II activities of the seedlings raised from hydroprimed seeds.
Interestingly, Pusa Ratna, which is the abiotic stress-sensitive variety consistently showed significant increase in photosynthetic pigment content and PS I activity of the seedlings under all the three growing conditions (0, NaCl, PEG stress). Hydroprimed Pusa Vishal (NaCl tolerant) showed indication of developing tolerance toward PEG stress by showing an enhancement in PS II activity when subjected to PEG stress. It was already reported that seed priming with salicylic acid and PEG increased the photosynthetic activities of the seedlings in rice [16].
Proline accumulation is known as a common metabolic reaction of plants under stress [5]. Hydropriming of seeds may generate or modulate certain mechanisms inside the seed system which gets carried over to the seedlings that might improve the stress ameliorating character in the seedlings such as the improvement in the status of proline [17]. Pusa Vishal showed a higher dependence on proline for stress tolerance (both in NaCl and PEG) than Pusa Ratna and Pusa 9531, as evident from the increased accumulation of the same.
Under abiotic stresses, peroxidation of membrane lipids occurs which is direct evidence and measure of stress induced damage at the cellular level. On hydropriming, the increased levels of MDA were reduced significantly in all the seedlings of V. radiata varieties. This result was in accordance with the findings of Amooaghaie [1], wherein it was found that MDA content reduced under hydropriming of alfalfa seeds.
Hydropriming resulted in increased activity of NR and SOD in the seedlings of all the three varieties, while the activity of POX was different in the three varieties. The POX activity was found to be increased in the tolerant varieties on hydropriming while in the sensitive variety, the activity was found to be decreased in the seedlings which were raised from hydroprimed seeds. Osmotic stress tolerance and antioxidant enzyme activities are closely related and were reported in many plants [3, 18]. From the present result, it was evident that priming altered the metabolic pathway for the production of antioxidant enzymes. Ashraf and Ali [3] reported the enhanced antioxidant enzyme activities of catalase and peroxidase in canola, conferring salt tolerance to the seedlings.
These results suggest that hydropriming which is the most simple and cheap method of seed priming have a greater potential in imparting osmotic tolerance to V. radiata varieties. This tolerance is achieved by altering the various metabolic pathways. Among the three varieties studied the Pusa Ratna; the abiotic stress-sensitive variety showed maximum positive response toward hydropriming by showing high photosynthetic pigment content, increased activity of photosystems and mitochondria, increased shoot length and fresh weight of the seedlings. Hydropriming also improves the osmotic tolerance potential of the tolerant varieties. Thus, by hydropriming the seeds, we can impart osmotic stress tolerance to stress-sensitive variety and can make it tolerant. In addition to this, we can also improve the stress tolerance of already known tolerant varieties.
References
Amooaghaie R (2011) The effect of hydro and osmopriming on alfalfa seed germination and antioxidant defenses under salt stress. Afr J Biotechnol 10:6269–6275
Arnon DI (1949) Copper enzymes in isolated chloroplasts polyphenoloxidase in Beta vulgaris. Plant Physiol 24:1–5
Ashraf M, Ali Q (2008) Relative membrane permeability and activities of some antioxidant enzymes as the key determinants of salt tolerance in canola (Brassica napus L.). Environ Exp Bot 63:266–273
Bates LS, Waldren RP, Teare ID (1973) Rapid determination of free proline for water stress studies. Plant Soil 39:205–208
Behairy RT, El-Danasoury M, Craker L (2012) Impact of ascorbic acid on seed germination, seedling growth and enzyme activity of salt stressed fenugreek. J Med Active Plants 1:106–113
Bennett MA, Waters L (1987) Seed hydration treatments for improved sweet maize germination and stand establishment. JAm Soc Hortic Sci 112:45–49
Gaspar T, Penel C, Greppin H (1975) Peroxidase and isoperoxidase in relation to root and flower formation. Plant Biochem J 2:33–47
Giannopolitis CN, Reis SK (1977) Superoxide dismutase I. Occurence in higher plants. Plant Physiol 59:309–314
Hageman RH, Reed AJ (1980) Nitrate reductase from higher plants. In: Sanpietra A (ed) Methods in enzymology, vol 23. Academic press, New York, pp 491–503
Heath RL, Packer L (1968) Photoperoxidation in isolated chloroplasts. I—kinetics and stoichiometry of fatty acid peroxidation. Arch Biochem Biophys 125:189–198
Jisha KC, Puthur JT (2014) Halopriming of seeds imparts tolerance to NaCl and PEG induced stress in Vigna radiata (L.) Wilczek varieties. Physiol Mol Biol Plants 20:303–312
Jisha KC, Puthur JT (2015) Seed priming with BABA (β-amino butyric acid): a cost-effective method of abiotic stress tolerance in Vigna radiata (L.) Wilczek. Protoplasma. https://doi.org/10.1007/s00709-015-0804-7
Jisha KC, Vijayakumari K, Puthur JT (2013) Seed priming for abiotic stress tolerance: an overview. Acta Physiol Plant 35:1381–1396
Kaur S, Gupta AK, Kaur N (2002) Effect of osmo and hydropriming of chickpea on seedling growth and carbohydrate metabolism under water deficit stress. Plant Growth Regul 37:17–22
Kaya MD, Okcu G, Atak M, Cikili Y, Kolsarici O (2006) Seed treatments to overcome salt and drought stress during germination in sunflower (Helianthus annuus L.). Eur J Agron 24:291–295
Li X, Zhang L (2012) SA and PEG induced priming for water stress tolerance in rice seedling. In: Zhu E, Sambath S (eds) Information technology and agricultural engineering. AISE, vol 134. Springer, Berlin, pp 25–87
Mondal S, Viji P, Bose B (2011) Role of seed hardening in rice variety Swarna (MTU 7029). Res J Seed Sci 4:157–165
Munns R (2002) Comparative physiology of salt and water stress. Plant Cell Environ 25:239–250
Posmyk MM, Janas KM (2007) Effects of seed hydropriming in presence of exogenous proline on chilling injury limitation in Vigna radiata L. seedlings. Acta Physiol Plant 29:509–517
Puthur JT (2000) Photosynthetic events in Sesbania sesban (L.) Merrill in relation to osmotic stress during different developmental stages. Ph.D. Thesis, Jamia Millia Islamia, New Delhi
Umair A, Ali S, Tareen MJ, Ali I, Tareen MV (2012) Effects of seed priming on the antioxidant enzymes activity of mung bean (Vigna radiata) seedlings. Pak J Nutr 11:140–144
Xiong L, Zhu JK (2002) Molecular and genetic aspects of plant responses to osmotic stress. Plant Cell Environ 25:131–139
Acknowledgements
We thank Sh. Manjunath Prasad C. T., Scientist, Seed Science and Technology Division, IARI, New Delhi, for providing seeds of V. radiata varieties. We also acknowledge Dr. Veenalakshmi, Assistant Professor, MES Asmabi College, Thrissur, for editing the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Jisha, K.C., Puthur, J.T. Seed Hydropriming Enhances Osmotic Stress Tolerance Potential in Vigna radiata. Agric Res 7, 145–151 (2018). https://doi.org/10.1007/s40003-018-0306-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40003-018-0306-x